
Abstract
Predator search efficiency can be enhanced by anthropogenic landscape change, leading to increased predator–prey encounters and subsequent prey population declines. Logging increases early successional vegetation, providing ungulate forage. This increased forage, however, is accompanied by linear feature networks that increase predator hunting efficiency by facilitating predator movement and increasing prey vulnerability. We used integrated step selection analyses to weigh support for multiple hypotheses representing the combined impact of logging features (cutblocks and linear features) on wolf (Canis lupus) movement and habitat selection in interior British Columbia. Further, we examine the relationship between logging and wolf kill-sites of moose (Alces alces) identified using spatiotemporal wolf location cluster analysis. Wolves selected for linear features, which increased their movement rates. New (0–8 years since harvest) cutblocks were selected by wolves. Moose kill-sites had a higher probability of occurring in areas with higher proportions of new and regenerating (9–24 years since harvest) cutblocks. The combined selection and movement responses by wolves to logging features, coupled with increased moose mortality sites associated with cutblocks, indicate that landscape change increases risk for moose. Cumulative effects of landscape change contribute to moose population declines, stressing the importance of cohesive management and restoration of anthropogenic features.
Similar content being viewed by others
Introduction
Anthropogenic landscape change modifies predator–prey dynamics, which has implications for both predator and prey populations1,2. Predators respond to prey through numerical responses (changes in predator density) and functional responses (changes in consumption rate), and mathematical models describing the functional response identify limits to the rate of prey consumption, prey encounter rate, and handling time3,4. Holling’s disc equation, which models the functional response, postulates that kill rate is limited by handling time at high prey densities, but by search efficiency at low prey densities3,4. Growing evidence indicates that anthropogenic landscape change influences predator search efficiency and thus, predator–prey encounter rate by facilitating predator movement and/or altering prey vulnerability1,5,6. Unless prey alter their behavior to avoid predation (e.g. sheltering in human-created refugia7) or landscape change bolsters prey populations by increasing habitat quality (e.g., increasing available forage), anthropogenic landscape change could lead to declining prey populations due to increased predator foraging efficiency.
Predators exploit specific anthropogenic features to increase search efficiency, which intensifies predation risk for prey8. Large carnivores often select and travel quickly on linear features, which improves foraging efficiency by increasing potential predator–prey encounters1,5,9. Logging—which creates both roads and cutblocks—increases predator travel efficiency, reduces hiding cover, concentrates prey in remaining patches, and creates predictable, small areas for predators to search10,11. Additionally, prey species are attracted to polygonal features such as cutblocks, where early seral vegetation offers abundant forage12,13,14. If predators hunt more efficiently due to linear features linking cutblocks5,15, these anthropogenic features could function cumulatively to increase predation risk for prey across disturbed landscapes.
Predation risk could be elevated if prey select for cutblocks (e.g. for increased forage) but logging features also increase predator search efficiency, possibly leading to an ecological trap16,17,18. In areas with extensive logging, such as forests infested with outbreaks of bark beetles (Scolytinae) which are subsequently logged to salvage timber19,20, the opportunity for such scenarios to manifest may be intensified16,17,18. Landscapes highly modified by salvage logging of beetle-killed forests are characterized by cutblocks that are significantly larger than conventional cutblocks, linked by extensive linear feature networks and interspersed with patches of forests relatively homogeneous in structure, age, and composition21,22. If functioning cumulatively to increase predation risk, salvage logging features could lead to prey population declines. This may be the case in the western sub-boreal, where extensive salvage logging of forest killed by mountain pine beetle (MPB; Dendroctonus ponderosae) outbreaks linked to climate change coincided with declines in moose (Alces alces) populations20,23,24,25.
One mechanism hypothesised for the moose population decline within interior BC could be increased movement rates of wolves (Canis lupus)—a primary predator of moose—and altered habitat selection, resulting in increased predation risk for moose near logging features25. Evidence suggests that anthropogenic landscape change—particularly, linear feature networks—facilitates predation by wolves1,5,26. Selection for linear features increases wolf movement efficiency, affecting predator–prey encounter rates and subsequent predation rates on ungulates1,5,9,27,28,29,30. Additionally, wolves select forest edges, cutblocks, and areas with new forage created by logging, due to increased availability of prey associated with these features14,15,31,32. We argue that anthropogenic features facilitating wolf travel and creating predictable prey locations is a concern for moose inhabiting areas undergoing extensive logging.
We examined whether the two dominant forms of anthropogenic landscape change associated with salvage logging—linear features and cutblocks—work to cumulatively influence wolf movement and habitat selection, and are tied to moose kill-site locations within our study area—interior British Columbia (BC), Canada. We ask: (1) do wolves select for salvage logging features; (2) do salvage logging features facilitate wolf movement; and (3) are salvage logging features linked to wolf kill-sites of moose? We hypothesized that the impacts of cutblocks and linear features function together to affect predator–prey dynamics. Specifically, we predict that wolves will select for cutblocks and linear features and have increased movement rates associated with these disturbance features. We expect that cutblock age and size will influence wolf habitat selection, with increased selection for smaller, regenerating (9–24 years since harvest) cutblocks due to increased prey availability14,33,34. Lastly, we expect these landscape features to facilitate wolf predation on moose, such that there is a positive relationship between salvage logging features and wolves’ kill-sites of moose.
Methods
Study area
BC’s Interior Plateau has experienced the cumulative effects of significant land conversion and habitat loss, with impacts to forests including a recent severe MPB outbreak35,36,37. This outbreak began in the 1990s, killing over 53% of merchantable pine (723 million m3 of pine)35,36,38. To mitigate economic effects, the Government of BC increased the annual allowable cut (timber amount sustainably harvested per year for a region) by approximately 30% from levels prior to the outbreak, resulting in extensive linear feature networks and the removal of large areas of beetle-killed trees36.
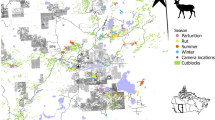
Our study area, Prince George South (PGS), is located southwest of the city of Prince George, on BC’s Interior Plateau (Fig. 1; Supplementary Information S1). PGS is one of five study areas in a long-term provincial moose monitoring project, selected for additional research on predation dynamics due to its continued moose declines and the identified role of wolves as a leading cause of mortality for both adult females and 8–12 month old calves25.

The Prince George South (PGS) study area, located in interior British Columbia, Canada. PGS is heavily altered with linear features (in grey) and cleared forest. The municipal boundary for the city of Prince George is outlined in red. Waterbodies are shown in blue. The map was created in ArcGIS v.10 (www.esri.com).
Integrated step selection analysis
Adult wolves were captured using standard protocols for aerial darting or net-gunning (December-March), or soft-catch, foot-hold trapping (June–July), 2018–2020 (Supplementary Information S1). All wolf captures and protocols were completed following Canadian Council on Animal Care and Animal Research: Reporting In Vivo Experiments (ARRIVE) guidelines, as well as BC Ministry of Environment Standards for Live Animal Capture and Handling. Approval for experimental protocols and animal care guidelines was issued in accordance with the British Columbia Wildlife Act and BC Ministry of Forest, Lands and Natural Resource Operations Animal Care Ethics Committee (permit: PG17-272811).
Captured wolves were fitted with satellite GPS collars (Vectronic Aerospace, Berlin) with a 60-min fix rate and 2-year drop-off mechanisms. Wolf movement data was divided into ‘winter’ (October 1-March 31; snow present, no pups) and ‘summer’ (April 1-September 30; denning, rearing pups, ungulate calving, snow-free) periods. We were interested in wolf habitat selection and movement during all periods except those associated with denning and rendezvous sites, where we assumed there would be limited prey searching behaviors by wolves39. We removed wolf locations within 1 km of these sites40, determined using GPS cluster analysis41 and ground truthing. We removed GPS locations within the first 48-h after capture to account for altered behavior following handling and only included wolves with > 7 days of movement data.
Integrated step selection analyses (iSSA) compare used (1) to available (0) locations of steps (connection between successive relocations), integrating habitat selection and movement within a conditional logistic regression model framework42. We used the R package ‘amt’ (Animal Movement Tools Version 0.0.6.) to generate ten random steps for every used wolf step43,44, drawn from population-level parametric distributions of step lengths (Euclidean distance between successive relocations) and turn angles (angle between consecutive relocations). Because we had a limited sample size of wolves and packs, we used individual wolves as the sampling unit and retained all individuals regardless of pack within the analysis. While this decision could lead to biased results due to pseudo-replication and territory restrictions, there is evidence suggesting that individual resource use varies between pack members45.
We included the following habitat covariates: cutblock use (new cutblock [0–8 years since harvest], regenerating cutblock [9–24 years since harvest] or outside of cutblock [reference category]33) and size; distance to, and density of, linear features; distance to edge habitat; land cover type (deciduous-leading stands, coniferous-leading stands, mixed forest stands, pine-leading stands, and non-forest); plant productivity (normalized difference vegetation index, NDVI); and distance to the nearest waterbody (Supplementary Information S1). All environmental covariates that were included as an interaction with ln(Step length) were extracted from the start of the step, while all other covariates were extracted from the end of the step. Distance covariates were log-transformed to account for skewness.
We developed candidate models with each model representing a competing hypothesis (Table 1), and modeled iSSAs for each individual in each season separately42,46. As step length may vary with time of day, we created a harmonic interaction term, hereafter referred to as sin(hour), to represent activity peaks at dawn and dusk, using the following formula: sin(\(\frac{4*\pi *(hour-6)}{24}\))47,48. All models included ln(Step length) and an interaction between sin(hour) and ln(Step length) to control for varying movement rates at different times of day (Supplementary Fig. S1).
Akaike’s Information Criterion (AIC) was used to determine the best-supported model for each individual wolf in each season. Performance of models was assessed using cross-validation, with data subset by step ID. For model selection, we determined the best overall model for each season by assessing the distribution of AIC weights. Then, we used bootstrapping to estimate population β coefficients and associated confidence intervals from the best-supported model46,47,49,50. This two-stage approach of fitting separate individual models and then post-hoc estimating population averages via bootstrapping is commonly used for iSSAs when sample sizes for individual steps are sufficient42. This approach allows for unbiased estimation of habitat selection variability and fewer assumptions than mixed-effects models42,46. For bootstrapping, we weighted samples by individual wolf i.d., which ensured equal probability of sampling for each individual wolf. From 2000 repetitions, we obtained the median and confidence interval for beta coefficient estimates (using 2.5th and 97.5th quantiles) which were used for population-level inferences. To quantify selection responses, we calculated relative strength of selection which estimates probability of selecting one resource unit over another51 (Supplementary Information S1).
Moose kill-site analysis
Moose mortality sites were determined by ground-truthing potential kill-sites identified by cluster analysis of wolf GPS locations, using the Find Points Cluster Identification Program Version 241 (Supplementary Information S1). We used logistic regressions to compare habitat features at sites of successful wolf kills of moose to random sites selected within the study area. We used variance inflation factors (VIF) to check for multicollinearity and excluded variables with VIF > 4. We weighed evidence for competing hypotheses relating landscape features to sites where moose were killed by wolves, following a set of a priori candidate models similar to the iSSA set (Supplementary Table S2), and selected the best supported model using AIC. For the top model, we used k-fold cross validation with k = 10 and Spearman’s rank correlation (rs) to assess model fit52.
Results
We deployed satellite GPS collars on ten wolves in five packs (Supplementary Table S1) and collected hourly location data between February 24, 2018 and July 31, 2020. Wolf collars were staggered in deployment and end date, so data was not available from all individuals through the study duration (Supplementary Table S1).
For both seasons, the ‘Global’ model outperformed the alternate models (Fig. 2, Table S3), indicating wolf movement and habitat selection is influenced by a combination of cutblocks, linear features and natural features. All remaining models received minimal support based on AIC weights.

Akaike’s information criterion (AIC) weight distribution for summer (April 1–September 30) and winter (October 1–March 31) integrated step selection analysis candidate models for Prince George South, 2018–2020. Outliers are represented by points.
Wolf selection for salvage logging features
In both seasons, wolves selected habitat closer to linear features (Table 2, Fig. 3). There was no clear trend in selection of varying linear feature densities for both seasons (Table 2).

Seasonal wolf log-transformed relative selection strength (RSS) with 95% confidence intervals for distance to linear features (m) in summer (April 1–September 30) and winter (October 1–March 31) for Prince George South, 2018–2020.
Wolf selection of logged areas was dependent on cutblock size and age in summer, but only on cutblock age in winter (Table 2). In both seasons, wolves selected for new cutblocks. In summer, selection of new cutblocks decreased as cutblock size increased. There was no clear trend in wolf selection of regenerating cutblocks during both seasons.
In summer, coniferous-leading forests were selected, and in winter, wolves selected for areas with high NDVI values (Table 2). Edge habitats and areas closer to water were selected for in both seasons.
Impact of salvage logging features on wolf movement
The impact of cutblocks on displacement rates varied between seasons (Table 2). In summer, wolf step lengths were shorter in regenerating cutblocks, but no trend existed in relation to new cutblocks or size of regenerating cutblocks. In winter, displacement rates were only associated with size of new cutblocks, with shorter step lengths as cutblock size increased.
In both seasons, wolves had faster displacement rates when closer to linear features (Fig. 4A). However, wolf step length decreased as the density of linear features increased (Fig. 4B).

Seasonal mean displacement rates (km/h) with 95% confidence intervals of Prince George South wolves in comparison to (A) distance to linear features (m) and (B) linear feature density (km/km2) for summer (April 1–September 30) and winter (October 1–March 31), 2018–2020.
Relationship between salvage logging features and moose kill-sites
We identified 158 moose kill-sites using cluster analysis of wolf GPS locations (Supplementary Information S1). A single top model was best supported: “Prey + Cutblocks” (Table S4; rs = 0.953). Moose kill-sites were more likely to occur in areas with higher proportions of new and regenerating cutblocks (Fig. 5A,B; Table 3). As mean NDVI increased, the probability of a moose kill-site occurring increased (Fig. 5C; Table 3). Moose kill-sites had a lower probability of occurring in areas with a higher proportion of deciduous-leading stands (Fig. 5D) and further from waterbodies (Table 3).

Predicted probability with 95% confidence intervals of a wolf kill-site of a moose occurring based on (A) proportion of new (0–8 years old) cutblocks, (B) proportion of regenerating (9–24 years old) cutblocks, and (C) mean normalized difference vegetation index (NDVI), and (D) proportion of deciduous-leading stands within a 883 m buffer around the location, in Prince George South, 2018–2020.
Discussion
Large-scale logging affects predator–prey dynamics by modifying predator search efficiency, elevating predation risk for prey near disturbance features. In our study, landscape change—cutblocks and linear feature networks—impacted wolf habitat selection and movement, altering the distribution of predation events to cutblocks. Based on our results, we suggest that extensive logging potentially creates an ecological trap mediated by season and cutblock age based on patterns of moose habitat selection identified in other studies within interior BC14,33,34. While this hypothesis requires further investigation, this scenario could contribute to moose population declines observed within the study area.
Wolf selection of new cutblocks, combined with an increased likelihood of moose kill-sites in areas with higher proportions of new and regenerating cutblocks, indicates that cutblocks are a risky feature for moose. In both seasons, wolves selected new cutblocks, suggesting better predation opportunities either due to higher prey availability or visibility53,54. Forage biomass increases post-harvest due to more solar insolation and nutrients available to plants, subsequently attracting ungulates12,15,55,56,57. However, forage biomass and ungulate use peaks a decade post-harvest12,55, and while moose kill-sites were linked to higher proportions of regenerating cutblocks, there was no trend in wolf selection for regenerating blocks. Regenerating cutblocks attract moose for the increased forage biomass and cover12,14,33, but increased vegetative cover would reduce prey visibility. Possibly, wolves are balancing prey availability and visibility in their selection of cutblocks, which is supported by our results: wolf movement rates were lower in regenerating cutblocks in summer, when wolf sightlines would be most obscured by vegetation. While wolves hunt with both olfactory and visual cues, areas with reduced cover (i.e. new cutblocks) are more likely to lead to a wolf successfully killing a moose due to both prey visibility and availability58,59, potentially leading to the observed selection of new cutblocks by wolves. However, adult female moose selection of new cutblocks—and thus, prey availability for wolves in these features—appears to vary based on season, with increased selection for new cutblocks in winter and avoidance in other seasons14,33,34. Consequently, moose vulnerability in new cutblocks is likely highest in winter due to their selection of these features, in addition to the presence of deeper snow. Wolf selection for new cutblocks throughout the year could indicate increased foraging success despite reduced moose availability in some seasons, which is supported by our kill-site analysis results. To clarify this, further studies could compare seasonal and demographic effects on spatial occurrence of wolf predation events of moose, which we were unable to do here due to limited sample sizes and data.
Consistent with previous research1,5,27,28,29, wolves selected for habitat near linear features and increased their displacement rates there. Linear features likely increase predation risk across the landscape by allowing predators to increase their search efficiency by facilitating movement1,5,31. Animals are predicted to spend less time in a foraging patch if the travel time between patches is reduced60 and therefore, linear features could promote faster searching of more habitat patches. Further, linear features provide travel corridors into refugia or biologically important habitat for ungulates, increasing spatial overlap between prey and predators27,29. As a result, predation risk may increase and homogenize across the landscape due to linear features.
We suggest that wolves use linear features as travel corridors into moose habitat which could enhance their chance of successfully detecting moose; however, linear features were not an important predictor of moose kill-sites. Unless used as human-created refugia61, ungulates generally avoid linear features due to perceived predation risk or limited forage availability relative to other habitats34,50,62. The combination of wolf selection for and moose avoidance of linear features5 likely interacts such that kill-sites are not necessarily close to linear features. Mumma and Gillingham63 also found that adult female moose were more likely to be killed by wolves in areas of low linear feature density. Therefore, kill-sites are not a function of linear features alone and our results suggest the cumulative effects of linear features and polygonal early-seral features produce the effect on kill-sites.
Despite selection for linear features, we observed no significant trend in wolf selection for areas of high linear feature densities. Previous research has identified inconsistent responses of wolves to varying densities of linear features64,65,66,67, which could be attributed to levels of human use—data which we lacked for PGS. While linear features may increase hunting efficiency of wolves, high linear feature densities are indicative of urban areas and increased accessibility of the landscape for human activities. If perceived as risky, areas with increased human activity would be avoided by wolves65,68. Alternatively, we were unable to differentiate between varying linear feature conditions (e.g., degree of vegetation growth) in the analysis and therefore, it is possible that this lack of trend in selection is an artefact of the dataset.
We propose that the behavioural responses to logging features by wolves coupled with cutblock forage attracting moose create conditions synonymous with an ecological trap for moose, mediated by season and cutblock age, although more research is required to conclude that a trap exists. Salvage logging creates a landscape with patches of attractive foraging habitats for ungulates (cutblocks), connected by a network of linear features that enable predator movement through the system, facilitating predation. Ungulates are attracted to the increased forage offered by regenerating vegetation in cutblocks12,55,56,57, but are more vulnerable to predation due to reduced cover58 and the ease of movement of predators through the system due to linear features1,5. Linear features increase spatial overlap of wolves and their prey by increasing accessibility of previously isolated habitat patches27,29 and allow wolves to efficiently search more of the landscape for prey1,5. If this potential ecological trap exists, it is likely mediated by season and cutblock age due to patterns in habitat selection by moose (i.e., increased selection for regenerating cutblocks; avoidance of new cutblocks except in winter; increased selection for cutblocks in winter) identified by previous moose research within interior BC14,33,34. Further investigation is required to characterize this potential ecological trap, by further assessing habitat preference and appropriate fitness and demographic measures for moose69.
Linear features and cutblocks function together to increase predation risk for prey, and effective management should target decoupling these disturbance features to reduce predator search efficiency. This could be accomplished by restoring linear features (e.g., felling trees, planting vegetation of > 1 m height70,71) that link cutblocks, to reduce wolf movement rates and access into moose habitat. Linear features linking biologically important but disjunct patches of moose habitat should be prioritized and if possible, construction of linear features should proactively avoid linking critical prey habitats. Habitat enhancement (e.g., planting palatable vegetation) should occur in areas where linear feature access is limited. Deciduous-leading stands may act as refuges for moose due to reduced wolf selection and fewer associated kill sites, and replanting or retention of these stands should be prioritized. Maintaining adequate cover for prey is important, by manipulating cutblock configuration to limit sightlines and decrease distance to cover, maintaining patches of intact forest (even dead standing pine), and allowing fast-growing shrubs to establish. However, shrub establishment may be a double-edged sword: while shrubs would disrupt predator sightlines and provide browse, they would encourage moose to use new cutblocks and potentially increase wolf-moose encounters. Overall, we emphasize the need to cohesively consider restoration and management of cutblocks and linear features in order to implement successful management programs, particularly in highly disturbed landscapes.
Data availability
Data are owned by the Government of British Columbia and are not publicly available at this time. Data requests can be directed to the Ministry of Environment and Climate Change Strategy Wildlife Species Inventory Team (SPI_Mail@gov.bc.ca). All spatial data is publicly available.
References
Dickie, M., Serrouya, R., McNay, R. S. & Boutin, S. Faster and farther: wolf movement on linear features and implications for hunting behaviour. J. Appl. Ecol. 54, 253–263 (2017).
Owen-Smith, N., Fryxell, J. M. & Merrill, E. H. Foraging theory upscaled: The behavioural ecology of herbivore movement. Philos. Trans. R. Soc. B Biol. Sci. 365, 2267–2278. https://doi.org/10.1098/rstb.2010.0095 (2010).
Holling, C. S. The functional response of predators to prey density and its role in mimicry and population regulation. Mem. Entomol. Soc. Can. 97, 5–60 (1965).
Holling, C. The components of predation as revealed by a study of small-mammal predation of the European pine sawfly (1959).
Dickie, M., McNay, S. R., Sutherland, G. D., Cody, M. & Avgar, T. Corridors or risk? Movement along, and use of, linear features varies predictably among large mammal predator and prey species. J. Anim. Ecol. https://doi.org/10.1111/1365-2656.13130 (2019).
DeCesare, N. J. Separating spatial search and efficiency rates as components of predation risk. Proc. Biol. Sci. 279, 4626–4633. https://doi.org/10.1098/rspb.2012.1698 (2012).
Muhly, T. B., Semeniuk, C., Massolo, A., Hickman, L. & Musiani, M. Human activity helps prey win the predator-prey space race. PLoS ONE 6, e17050. https://doi.org/10.1371/journal.pone.0017050 (2011).
Fleming, P. A. & Bateman, P. W. Novel predation opportunities in anthropogenic landscapes. Anim. Behav. 138, 145–155. https://doi.org/10.1016/j.anbehav.2018.02.011 (2018).
Whittington, J. et al. Caribou encounters with wolves increase near roads and trails: A time-to-event approach. J. Appl. Ecol. 48, 1535–1542. https://doi.org/10.1111/j.1365-2664.2011.02043.x (2011).
Larivière, S. & Messier, F. Effect of density and nearest neighbours on simulated waterfowl nests: Can predators recognize high-density nesting patches?. Oikos 83, 12–20. https://doi.org/10.2307/3546541 (1998).
Taitt, M. J. & Krebs, C. J. Predation, cover, and food manipulations during a spring decline of Microtus townsendii. J. Anim. Ecol. 52, 837–848. https://doi.org/10.2307/4458 (1983).
Fisher, J. T. & Wilkinson, L. The response of mammals to forest fire and timber harvest in the North American boreal forest. Mammal. Rev. 35, 51–81 (2005).
Fisher, J. T. & Burton, A. C. Wildlife winners and losers in an oil sands landscape. Front. Ecol. Environ. 16, 323–328. https://doi.org/10.1002/fee.1807 (2018).
Francis, A. L., Procter, C., Kuzyk, G. & Fisher, J. T. Female Moose Prioritize Forage Over Mortality Risk in Harvested Landscapes. J. Wildl. Manag. (2021).
Hebblewhite, M., Munro, R. H. & Merrill, E. H. Trophic consequences of postfire logging in a wolf–ungulate system. For. Ecol. Manag. 257, 1053–1062. https://doi.org/10.1016/j.foreco.2008.11.009 (2009).
Pulliam, H. R. Sources, sinks, and population regulation. Am. Nat. 132, 652–661 (1988).
Battin, J. When good animals love bad habitats: Ecological traps and the conservation of animal populations. Conserv. Biol. 18, 1482–1491 (2004).
Nielsen, S. E., Stenhouse, G. B. & Boyce, M. S. A habitat-based framework for grizzly bear conservation in Alberta. Biol. Conserv. 130, 217–229 (2006).
Bentz, B. et al. Salt Lake City 42 (University of Utah Press, 2005).
Carroll, A. L., Taylor, S. W., Régnière, J. & Safranyik, L. in Mountain pine beetle symposium: challenges and solutions. 223–232 (Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre).
Lindenmayer, D. B. & Noss, R. F. Salvage logging, ecosystem processes, and biodiversity conservation. Conserv. Biol. 20, 949–958. https://doi.org/10.1111/j.1523-1739.2006.00497.x (2006).
Leverkus, A. B., Lindenmayer, D. B., Thorn, S. & Gustafsson, L. Salvage logging in the world’s forests: Interactions between natural disturbance and logging need recognition. Glob. Ecol. Biogeogr. 27, 1140–1154. https://doi.org/10.1111/geb.12772 (2018).
Kuzyk, G. et al. Moose population dynamics during 20 years of declining harvest in British Columbia. Alces 54, 101–119 (2018).
Kuzyk, G. W. Provincial population and harvest estimates of moose in British Columbia. Alces J. Devot. Biol. Manag. Moose 52, 1–11 (2016).
Procter, C. et al. Factors affecting moose population declines in British Columbia. 2020 Progress Report: February 2012-May 2020. B.C. Ministry of Forests, Lands, Natural Resource Operations and Rural Development, Victoria, B.C., Wildlife Working Report No. WR-128. Pp. 89. https://www2.gov.bc.ca/gov/content/environment/plants-animals-ecosystems/wildlife/wildlife-conservation/moose/moose-conservation/moose-research. (2020).
Wittmer, H. U., Sinclair, A. R. E. & McLellan, B. N. The role of predation in the decline and extirpation of woodland caribou. Oecologia 144, 257–267. https://doi.org/10.1007/s00442-005-0055-y (2005).
Latham, A. D. M., Latham, M. C., Boyce, M. S. & Boutin, S. Movement responses by wolves to industrial linear features and their effect on woodland caribou in northeastern Alberta. Ecol. Appl. 21, 2854–2865 (2011).
James, A. R. C. & Stuart-Smith, A. K. Distribution of caribou and wolves in relation to linear corridors. J. Wildl. Manag. 64, 154–159. https://doi.org/10.2307/3802985 (2000).
DeMars, C. A. & Boutin, S. Nowhere to hide: Effects of linear features on predator–prey dynamics in a large mammal system. J. Anim. Ecol. 87, 274–284. https://doi.org/10.1111/1365-2656.12760 (2018).
McKenzie, H. W., Merrill, E. H., Spiteri, R. J. & Lewis, M. A. How linear features alter predator movement and the functional response. Interface Focus 2, 205–216. https://doi.org/10.1098/rsfs.2011.0086 (2012).
Houle, M., Fortin, D., Dussault, C., Courtois, R. & Ouellet, J.-P. Cumulative effects of forestry on habitat use by gray wolf (Canis lupus) in the boreal forest. Landsc. Ecol. 25, 419–433. https://doi.org/10.1007/s10980-009-9420-2 (2010).
Kuzyk, G. W., Kneteman, J. & Schmiegelow, F. K. Winter habitat use by wolves, Canis lupus, in relation to forest harvesting in west-central Alberta. Can. Field Nat. 118, 368–375 (2004).
Mumma, M. A. et al. Regional moose (Alces alces) responses to forestry cutblocks are driven by landscape-scale patterns of vegetation composition and regrowth. For. Ecol. Manag. 481, 118763 (2021).
Scheideman, M. Use and selection at two spatial scales by female moose (Alces alces) across central British Columbia following a mountain pine beetle outbreak MSc thesis, University of Northern British Columbia (2018).
Alfaro, R. I., van Akker, L. & Hawkes, B. Characteristics of forest legacies following two mountain pine beetle outbreaks in British Columbia Canada. Can. J. For. Res. 45, 1387–1396 (2015).
Dhar, A., Parrott, L. & Hawkins, C. D. B. Aftermath of mountain pine beetle outbreak in British Columbia: Stand dynamics, management response and ecosystem resilience. Forests 7, 171 (2016).
Shackelford, N., Standish, R. J., Ripple, W. & Starzomski, B. M. Threats to biodiversity from cumulative human impacts in one of North America’s last wildlife frontiers. Conserv. Biol. 32, 672–684 (2018).
Corbett, L. J., Withey, P., Lantz, V. A. & Ochuodho, T. O. The economic impact of the mountain pine beetle infestation in British Columbia: Provincial estimates from a CGE analysis. For. Int. J. For. Res. 89, 100–105. https://doi.org/10.1093/forestry/cpv042 (2015).
Latham, A. D. M. Wolf ecology and caribou-primary prey-wolf spatial relationships in low productivity peatland complexes in northeastern Alberta PhD thesis, University of Alberta, (2009).
Person, D. K. & Russell, A. L. Reproduction and den site selection by wolves in a disturbed landscape. Northw. Sci. 83, 211–224. https://doi.org/10.3955/046.083.0305 (2009).
Gillingham, M. Documentation for using Find Points Cluster Identification Program (Version 2) (University of Northern British Columbia, 2009).
Avgar, T., Potts, J. R., Lewis, M. A. & Boyce, M. S. Integrated step selection analysis: Bridging the gap between resource selection and animal movement. Methods Ecol. Evol. 7, 619–630. https://doi.org/10.1111/2041-210X.12528 (2016).
Signer, J., Fieberg, J. & Avgar, T. Animal movement tools (amt): R package for managing tracking data and conducting habitat selection analyses. Ecol. Evol. 9, 880–890 (2019).
Thurfjell, H., Ciuti, S. & Boyce, M. S. Applications of step-selection functions in ecology and conservation. Mov. Ecol. 2, 4. https://doi.org/10.1186/2051-3933-2-4 (2014).
Benson, J. F. & Patterson, B. R. Spatial overlap, proximity, and habitat use of individual wolves within the same packs. Wildl. Soc. Bull. (2011-) 39, 31–40 (2015).
Fieberg, J., Matthiopoulos, J., Hebblewhite, M., Boyce, M. S. & Frair, J. L. Correlation and studies of habitat selection: problem, red herring or opportunity?. Philos. Trans. R. Soc. B Biol. Sci. 365, 2233–2244 (2010).
Ladle, A. et al. Grizzly bear response to spatio-temporal variability in human recreational activity. J. Appl. Ecol. 56, 375–386. https://doi.org/10.1111/1365-2664.13277 (2019).
Kohl, M. T. et al. Diel predator activity drives a dynamic landscape of fear. Ecol. Monogr. 88, 638–652 (2018).
Scrafford, M. A., Avgar, T., Heeres, R. & Boyce, M. S. Roads elicit negative movement and habitat-selection responses by wolverines (Gulo gulo luscus). Behav. Ecol. 29, 534–542. https://doi.org/10.1093/beheco/arx182 (2018).
Prokopenko, C. M., Boyce, M. S. & Avgar, T. Characterizing wildlife behavioural responses to roads using integrated step selection analysis. J. Appl. Ecol. 54, 470–479. https://doi.org/10.1111/1365-2664.12768 (2017).
Avgar, T., Lele, S. R., Keim, J. L. & Boyce, M. S. Relative selection strength: Quantifying effect size in habitat- and step-selection inference. Ecol. Evol. 7, 5322–5330. https://doi.org/10.1002/ece3.3122 (2017).
Boyce, M. S., Vernier, P. R., Nielsen, S. E. & Schmiegelow, F. K. A. Evaluating resource selection functions. Ecol. Model. 157, 281–300. https://doi.org/10.1016/S0304-3800(02)00200-4 (2002).
Visscher, D. R. & Merrill, E. H. Temporal dynamics of forage succession for elk at two scales: Implications of forest management. For. Ecol. Manag. 257, 96–106. https://doi.org/10.1016/j.foreco.2008.08.018 (2009).
Stelfox, J. G., Lynch, G. M. & McGillis, J. R. Effects of clearcut logging on wild ungulates in the Central Albertan foothills. For. Chron. 52, 65–70. https://doi.org/10.5558/tfc52065-2 (1976).
Gagné, C., Mainguy, J. & Fortin, D. The impact of forest harvesting on caribou–moose–wolf interactions decreases along a latitudinal gradient. Biol. Conserv. 197, 215–222. https://doi.org/10.1016/j.biocon.2016.03.015 (2016).
Potvin, F., Breton, L. & Courtois, R. Response of beaver, moose, and snowshoe hare to clear-cutting in a Quebec boreal forest: a reassessment 10 years after cut. Can. J. For. Res. 35, 151–160 (2005).
Rempel, R. S., Elkie, P. C., Rodgers, A. R. & Gluck, M. J. Timber-management and natural-disturbance effects on moose habitat: landscape evaluation. J. Wildl. Manag. 61, 517–524. https://doi.org/10.2307/3802610 (1997).
Kunkel, K. E. & Pletscher, D. H. Habitat factors affecting vulnerability of moose to predation by wolves in southeastern British Columbia. Can. J. Zool. 78, 150–157. https://doi.org/10.1139/z99-181 (2000).
Mech, L. D. & Boitani, L. Wolves: behavior, ecology, and conservation. (University of Chicago Press, 2007).
Charnov, E. L. Optimal foraging, the marginal value theorem. (1976).
Hebblewhite, M. & Merrill, E. H. Trade-offs between predation risk and forage differ between migrant strategies in a migratory ungulate. Ecology 90, 3445–3454. https://doi.org/10.1890/08-2090.1 (2009).
Lendrum, P. E., Anderson Jr, C. R., Long, R. A., Kie, J. G. & Bowyer, R. T. Habitat selection by mule deer during migration: effects of landscape structure and natural-gas development. Ecosphere 3, art82. https://doi.org/10.1890/ES12-00165.1 (2012).
Mumma, M. & Gillingham, M. Determining factors that affect survival of moose in Central British Columbia. Technical report to the Habitat Conservation Trust Foundation for Grant Agreement CAT19-0-522 (1 April 2017 through 31 March 2019). 56 (2019).
Roffler, G. H., Gregovich, D. P. & Larson, K. R. Resource selection by coastal wolves reveals the seasonal importance of seral forest and suitable prey habitat. For. Ecol. Manag. 409, 190–201. https://doi.org/10.1016/j.foreco.2017.11.025 (2018).
Lesmerises, F., Dussault, C. & St-Laurent, M.-H. Wolf habitat selection is shaped by human activities in a highly managed boreal forest. For. Ecol. Manag. 276, 125–131. https://doi.org/10.1016/j.foreco.2012.03.025 (2012).
Muhly, T. B. et al. Functional response of wolves to human development across boreal North America. Ecol. Evol. 9, 10801–10815. https://doi.org/10.1002/ece3.5600 (2019).
Mladenoff, D. J., Sickley, T. A. & Wydeven, A. P. Predicting gray wolf landscape recolonization: logistic regression models vs. new field data. Ecol. Appl. 9, 37–44. https://doi.org/10.1890/1051-0761(1999)009[0037:PGWLRL]2.0.CO;2 (1999).
Rogala, J. K. et al. Human activity differentially redistributes large mammals in the Canadian Rockies National Parks. Ecol. Soc. 16 (2011).
Robertson, B. A. & Hutto, R. L. A framework for understanding ecological traps and an evaluation of existing evidence. Ecology 87, 1075–1085. https://doi.org/10.1890/0012-9658(2006)87[1075:AFFUET]2.0.CO;2 (2006).
Finnegan, L. et al. Natural regeneration on seismic lines influences movement behaviour of wolves and grizzly bears. PLoS ONE 13, e0195480. https://doi.org/10.1371/journal.pone.0195480 (2018).
Dickie, M., Serrouya, R., DeMars, C., Cranston, J. & Boutin, S. Evaluating functional recovery of habitat for threatened woodland caribou. Ecosphere 8, e01936. https://doi.org/10.1002/ecs2.1936 (2017).
Acknowledgements
We thank FLNRORD staff and volunteers who assisted with field work, and Rob Altoft and Matt Scheideman for assistance with captures. This study was funded by the Ministry of Forests, Lands, Natural Resource Operations and Rural Development, Habitat Conservation Trust Foundation, Natural Sciences and Engineering Research Council, the Ian McTaggart Cowan Professorship at the University of Victoria, and the Forest Enhancement Society of British Columbia.
Funding
This article was funded by Natural Sciences and Engineering Research Council of Canada, University of Victoria, Ministry of Forests, Lands, Natural Resource Operations and Rural Development.
Author information
Authors and Affiliations
Contributions
All authors conceived and designed the study. M.A., C.P., S.M. and G.K. collected the data. N.B. and A.L. analyzed the data. N.B. led the writing of the manuscript. All authors provided significant contributions to manuscript drafts and approved the final version for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Boucher, N.P., Anderson, M., Ladle, A. et al. Cumulative effects of widespread landscape change alter predator–prey dynamics. Sci Rep 12, 11692 (2022). https://doi.org/10.1038/s41598-022-15001-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-15001-3
- Springer Nature Limited
This article is cited by
-
Advances in biologging can identify nuanced energetic costs and gains in predators
Movement Ecology (2024)