
Abstract
Drug-induced off-target cardiotoxicity, particularly following anti-cancer therapy, is a major concern in new drug discovery and development. To ensure patient safety and efficient pharmaceutical drug development, there is an urgent need to develop more predictive cell model systems and distinct toxicity signatures. In this study, we applied our previously proposed repeated exposure toxicity methodology and performed 1H NMR spectroscopy-based extracellular metabolic profiling in culture medium of human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) exposed to doxorubicin (DOX), an anti-cancer agent. Single exposure to DOX did not show alteration in the basal level of extracellular metabolites while repeated exposure to DOX caused reduction in the utilization of pyruvate and acetate, and accumulation of formate compared to control culture medium. During drug washout, only pyruvate showed reversible effect and restored its utilization by hiPSC-CMs. On the other hand, formate and acetate showed irreversible effect in response to DOX exposure. DOX repeated exposure increased release of lactate dehydrogenase (LDH) in culture medium suggesting cytotoxicity events, while declined ATP levels in hiPSC-CMs. Our data suggests DOX perturbed mitochondrial metabolism in hiPSC-CMs. Pyruvate, acetate and formate can be used as metabolite signatures of DOX induced cardiotoxicity. Moreover, the hiPSC-CMs model system coupled with metabolomics technology offers a novel and powerful approach to strengthen cardiac safety assessment during new drug discovery and development.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Drug-induced cardiotoxicity is a leading cause of drug attrition in drug discovery and development. Hence cardiac safety assessment remains a critical component in preclinical drug development. In addition to electrophysiological studies, multiple parameters need to be considered to evaluate series of toxic events of pharmaceutical compounds. Currently used safety assessment parameters are informative but not sufficient to exclude the potential cardiotoxic effects of new drugs for human. Moreover, till now the experimental in vitro and in vivo animal models cannot truly predict cardiotoxicity in humans because of physiological differences between different species. Such model systems can identify false negative compounds, which can cause serious structural and functional damage to the human heart. Therefore pharmaceutical industries urgently need novel and diverse biomarkers, and also clinically relevant in vitro experimental models. Toxicological studies routinely use biomarkers to evaluate harmful effects of compounds. Advancement in ‘-omics’ technologies like transcriptomics and proteomics has accelerated the process of new biomarker discovery. Metabolomics is an emerging ‘-omics’ technology that offer new parameter of global metabolic profiling and is a promising tool for biomarker discovery (Johnson et al. 2016; Lewis et al. 2008). Nuclear Magnetic Resonance (NMR) and Mass Spectroscopy (MS) are routinely used in the metabolic profiling of biological samples. Compared to MS, NMR is less sensitive but highly reproducible, with other advantages over MS (Markley et al. 2016). NMR spectroscopy-based metabolomics can quantify low-molecular weight metabolites in biological samples (cells/tissues/organism), biological fluids (serum/urine) and cell culture media. NMR-based metabolite profiling has been used to detect drug responsive to early changes in metabolite levels contributing to the subsequent toxicity (Andreadou et al. 2009; Park et al. 2009; Zhang et al. 2015). Such toxicity-responsive metabolite signatures can be used to predict adverse effect of drug candidates at an early stage of preclinical drug development (Clarke and Haselden 2008; Robertson et al. 2011; Wishart 2008).
Human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) beat spontaneously, recapitulate human cardiac biology and show clinically relevant physiological response upon exposure to drugs (Liang et al. 2013; van Meer et al. 2016). Since hiPSC-CMs may overcome the limitations of existing experimental models used in cardiac safety assessment, they are becoming reliable source for high-throughput drug discovery screening, toxicity prediction, cardiac disease modeling, regenerative medicine and for wide range of life sciences studies (Chow et al. 2013; Ebert et al. 2012; Sharma et al. 2013).
Doxorubicin (DOX) is a highly effective chemotherapeutic drug used in the treatment of various types of cancer including solid tumors, hematological malignancies and soft tissue sarcoma. However, its chemotherapeutic use is mainly limited by the occurrence of dose-dependent cardiotoxicity (Gianni et al. 2008; Zhang et al. 2012). The DOX induced cardiotoxicity is manifested by acute cardiac dysfunction, chronic cardiomyopathy and progressively leading finally to congestive heart failure. Although extensive work has been done on DOX, the mechanism of DOX mediated cardiotoxicity is still not completely understood. Different types of molecular mechanisms are proposed including oxidative stress, topoisomerase II beta inhibition and alteration in mitochondrial energetics (Minotti et al. 2004; Octavia et al. 2012; Zhang et al. 2012). Cellular energetics plays a critical role in DOX induced cardiomyopathy (Tokarska-Schlattner et al. 2006).
In this study, 1H NMR spectroscopy-based metabolite profiling was performed in extracellular culture medium of hiPSC-CMs exposed to DOX or solvent control. The metabolic profiling was performed to identify early metabolic signatures in response to DOX exposure. The present findings from metabolomics measurements are consistent with DOX-induced disturbances in mitochondrial metabolism. Our study demonstrates that metabolic profiling of media from hiPSC-CM cultures in the presence of cardiotoxicants can provide a unique platform to predict the cardiotoxic potential of pharmaceutical drug candidates.
Materials and methods
Cardiomyocytes cell culture
Highly pure population (>98%) of human induced pluripotent stem cell-derived cardiomyocytes (iCell Cardiomyocytes®, Cellular Dynamics International, Madison, WI, USA) were used to perform all the experiments. These spontaneously beating cardiomyocytes are mixture of atrial, ventricular and nodal like myocytes. For metabolomics studies, cryopreserved hiPSC-CMs were thawed as per manufactured instructions and resuspended in iCell cardiomyocytes plating medium (iCell-PM; Cellular Dynamics International, Madison, WI, USA) and plated on fibronectin (5 µg/cm2, 2 h at 37 °C) (Sigma-Aldrich, Steinheim, Germany) coated 6-well cell culture grade plate at a density of approximately 0.4 × 106 cells/well. Two days post plating; hiPSC-CMs were refreshed with and maintained in iCell-cardiomyocytes maintenance medium (iCell-MM; Cellular Dynamics International, Madison, Wi, USA) with fresh media change after every 2 days. Four days post-plating hiPSC-CMs were used for compound exposure. The cardiomyocytes were cultured in standard cell culture incubator under culture conditions of 5% CO2 and 37 °C.
Reference compound
10 mM stock solution of Doxorubicin (Sigma-Aldrich, Steinheim, Germany) was prepared in high quality dimethyl sulfoxide (DMSO) solvent. Stock solution was dispensed into small volume aliquots and stored at −20 °C for long-term use. iCell-MM was brought to room temperature prior to drug dilutions.
Experimental design and compound exposure
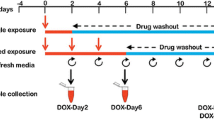
For metabolic profiling studies, we used our previously established in vitro repeated exposure toxicity methodology (Fig. 1) (Chaudhari et al. 2016). In short, hiPSC-CMs were given single exposure to DOX (156 nM) for 2 days (DOX-Day2) or three consecutive repeated exposures to DOX (DOX-Day6) (DOX supplemented media refreshed every 2 days). Following DOX exposure (single/repeated), hiPSC-CMs were cultured in drug free iCell-MM till day 14 after the start of drug exposure (DOX-Day2WO and DOX-Day6WO). Control-Day2 and -Day6 cardiomyocytes were exposed to DMSO as a solvent control. Culture medium was refreshed every 2 days.

A schematic view of experimental design with timeline of the study. Synchronously beating monolayer of hiPSC-CMs was exposed to DOX for 2 days or for 6 days in a repeated way (DOX-supplemented culture media was refreshed every 2 days). Following 2 and 6 days exposure to DOX, hiPSC-CMs were further cultured in drug free culture medium until day 14 (after the start of drug exposure). Culture media was refreshed every 2 days. For metabolic profiling, culture media samples were collected on day 2, 6 and 14. Experimental design was also used in cytotoxicity and ATP content determination studies
Sample collection and metabolomics analysis
Metabolomics samples were collected from three independent experiments. As shown in Fig. 1 the experimental media samples were collected from the cardiomyocytes culture medium both during drug exposure (Day 2 and Day 6) and drug washout (Day 14 after start of first drug exposure). Media samples from control cardiomyocytes (vehicle control) were also harvested at Day 0, 2, 6 and 14 time points. Media samples were collected in sterile tubes and immediately stored at −80 °C. Culture media samples represent the metabolite changes in the spent culture media over a culture period of the previous 2 days. Media that had not been exposed to cells was analysed to determine the initial concentration of individual metabolites, which allowed us to determine if the change in concentration observed was a result of cellular influx or efflux. For metabolomics studies, the media samples were thawed on ice. A DSA (4,4-dimethyl-4-silapentane-1-ammonium trifluoroacetate [D2O (50 µL)] solution was added to the media sample (550 µL) to give a final DSA concentration of 0.97 mmol/L. The samples were centrifuged to remove any particulate matter and transferred to a 5 mm NMR tube (Norell, USA). 1D Carr–Purcell–Meiboom–Gill (CPMG) NMR spectroscopy and data processing was carried out as previously described (Ellis et al. 2011; Keun and Athersuch 2011).
Determination of ATP content and LDH release
High energy ATP molecules functions as a biomarker for metabolically active cells, and lactate dehydrogenase (LDH) release is general cytotoxicity biomarker. The iPSC-CMs were plated in fibronectin coated white 96-well plate at a density of approximately 20 × 103 cells/well using iCell-PM. After 2 days of culture, iCell-PM was refreshed with iCell-MM. Four days post-plating, cardiomyocytes were exposed to DOX (156 nM) as per experimental design shown in Fig. 1. On day 2, 6 and 14, cardiomyocytes were used for ATP content determination, while culture media was used to measure enzymatic activity of LDH, released by cardiomyocytes in response to DOX exposure.
ATP amount was measured using ATPlite Luminescence ATP detection assay system kit (Perkin Elmer, Netherlands), according to the manufacturer’s instructions. In short, viable cardiomyocytes in the plate were lysed with mammalian cell lysis solution for 5 min in an orbital shaker. Lysed cells were immediately incubated with substrate solution for 5 min on an orbital shaker and then kept it undisturbed for 10 min in the dark place. After incubation, luminescence was measured on Softmax Pro M5e 96-well plate reader (Molecular Devices, Sunnyvale, CA, USA). Background luminescence was also measured from blank wells containing culture medium without cardiomyocytes. Emitted luminescence values are directly proportional to the amount of ATP.
Extracellular LDH activity was assessed using the Thermo Scientific™ Pierce™ LDH Cytotoxicity Assay Kit (colorimetric method) according to the manufacturer’s instructions. In brief, each media sample was gently mixed with reaction mixture, followed by incubation in the dark for 30 min at room temperature. The absorbance was measured at 490 nm using Softmax Pro M5e 96-well plate reader. Increase in absorbance is directly proportional to the LDH activity in the culture media. LDH activity in the culture medium without cardiomyocytes was considered as baseline activity.
Results
Analysis of the metabolomics data
For metabolic profiling studies, we used our previously established in vitro repeated exposure toxicity methodology (Fig. 1) (Chaudhari et al. 2016). 1H NMR spectroscopy was applied in metabolic profiling of culture medium of hiPSC-CMs model. A representative 1H CPMG spectrum of the spent culture media (Control-Day2) is shown in Fig. 2. Specific resonances are assigned to metabolites where possible and a full list of assigned spectral regions is listed in Table 1 with chemical shift ranges indicated. The chemical shift, in the units of ppm, describes where in the NMR spectrum a specific metabolite resonance is detected. In addition to the characterization of the extracellular metabolites at baseline, we investigated the influx and efflux of individual metabolites in culture media of hiPSC-CMs during and after DOX exposure (drug washout). Control media samples were used to determine changes in metabolite profile of DOX exposed media samples. No significant difference was observed between metabolic profile of Control-Day2 and DOX-Day2 groups. However, media samples from DOX-Day6 exhibited distinct changes in the metabolic profile compared to Control-Day6. Specifically, of the 20 metabolites studied, 3 showed a significant DOX-related effect in at least one treatment group; formate, pyruvate and acetate (Fig. 3). Either reversible or irreversible DOX-induced changes in the levels of these three metabolites were detected by 1H NMR metabolomics.

An extracellular metabolic profile of hiPSC-CMs model. The regions of the spectrum shown in the figure are a typical full resolution 1D 600 MHz 1H CPMG spin-echo NMR spectrum from the control media samples after 2 days of incubation. 1 3-methyl-2-oxovalerate, 2 isoleucine, 3 Leucine, 4 valine, 5 3-hydroxyisobutyrate, 6 ethanol, 7 lactate, 8 threonine, 9 2-hydroxyisobutyrate, 10 alanine, 11 lysine, 12 acetate, 13 pyroglutamate, 14 glutamine, 15 methionine, 16 pyruvate, 17 T-methylhistidine, 18 phenylalanine, 19 galactose, 20 tyrosine, 21 formate

NMR spectroscopy detected significant alterations in extracellular metabolites levels of hiPSC-CMs. DOX reduced uptake of a pyruvate, b acetate, and c reduced efflux of formate. During drug washout pyruvate showed a reversible effect, while acetate and formate showed a irreversible effect. Error bars in the bar graphs represents standard deviation (SD). n = 3 biological replicates, *p value 0.05
DOX exposure showed a temporal effect on the uptake of only one media component; pyruvate. Unlike single exposure, media samples collected from DOX-Day6 group showed significant reduction in the utilization of pyruvate and acetate (Fig. 3a, b). During drug washout, a reversible effect was observed in the utilization of pyruvate, while acetate showed an irreversible effect with high concentration levels in the culture medium. Control media samples exhibited increasing concentration levels of formate (effluxed by hiPSC-CMs) in a culture condition dependent manner. Compared to Control-Day6, DOX exposure in DOX-Day6 group significantly inhibited the accumulation of formate in the culture media (Fig. 3c). In addition, formate was the only metabolite for which production was irreversibly impaired by DOX, as evidenced by a significantly reduced level of formate in DOX-Day2WO and DOX-Day6WO media samples after drug washout. These findings suggest that DOX impaired mitochondrial metabolism in hiPSC-CMs.
No significant changes in the utilization of amino acids; valine, isoleucine and methionine (Fig. 4) could be observed. Leucine and lactate concentrations could not be calculated due to interference from other metabolites. In addition, no significant changes were found in the 3-methyl-2-oxovalerate, 2-Hydroxyisobutyrate and 3-hydroxyisobutyrate concentration levels (Fig. 4).

Response of extracellular metabolites- valine, 3-methyl-2-oxovalerate, isoleucine, tyrosine, 3-hydroxyisobutyrate, methionine and 2-hydroxybutyrate to DOX exposure in hiPSC-CMs model. 1H NMR spectroscopy was used to calculate the concentration (mM) of the metabolites in the culture media at day 2, 6 and 14 of DOX study. The levels of the metabolite are the net accumulation of influx and efflux over the previous 2 day period
DOX induced LDH leakage from hiPSC-CMs
LDH is impermeable to viable cell membrane, but it is released by cells in culture medium/blood stream in response to membrane damage and compound induced cytotoxicity. Compared to Control-Day2, media samples of DOX-Day2 did not show an increase in LDH activity (Fig. 5a). On the other hand, in media samples of DOX-Day6 group LDH release was observed with significant increase in LDH activity as compared to Control-Day6. Increased LDH activity indicates DOX induced cell membrane damage and cytotoxicity. Relative to Control-Day14, no significant increase in LDH activity was observed in drug washout media samples (DOX-Day2WO and DOX-Day6WO).

Determination of DOX induced cytotoxicity and energy levels in hiPSC-CMs. a DOX on repeated exposure induced cytotoxicity, as evidenced by increased LDH activity in the culture media. b DOX exposure declined energy level in cardiomyocytes with depletion of high energy ATP molecules. Data in bar graphs presented as mean ± SD. n = 3 technical replicates, *p value 0.05
DOX reduced ATP levels in hiPSC-CMs
ATP measurement was performed to determine cellular energy level after DOX exposure. Compared to controls, DOX exposure dependent depletion in the ATP levels was observed in DOX-Day2 and DOX-Day6 (Fig. 5b). Interestingly during drug washout, hiPSC-CMs from DOX-Day2WO group could not restore ATP levels, but a further decrease in ATP content was observed. Similarly hiPSC-CMs repeatedly exposed to DOX could not restore ATP levels after the removal of DOX (DOX-Day6WO). These data indicate a prolonged effect of DOX on depletion of the ATP-pool in hiPSC-CMs.
Discussion
NMR-based metabolic profiling is a flexible approach to identify pharmaceutical compounds with potential physiological toxicity. Moreover, metabolomics platform may offer metabolites as toxicity biomarkers in preclinical predictive toxicity screening. For the first time we applied the NMR-based spectroscopy in metabolic profiling of hiPSC-CMs model system to detect extracellular metabolite signatures of DOX induced toxicity.
In this context, we applied our in vitro repeated exposure toxicity methodology (Chaudhari et al. 2016) to identify DOX response early, intermediate and long-term affected extracellular metabolites in hiPSC-CMs model system. Our previous cytotoxicity studies combined with functional studies based on impedance measurements demonstrated that DOX at a concentration of 156 nM was non-toxic, while its repeated exposure induced significant arrhythmicity and cytotoxicity to hiPSC-CMs (Chaudhari et al. 2016). These impedance based data were consistent with our LDH-based cytotoxicity data. The present metabolomics data suggests that hiPSC-CMs are non-responsive to a single exposure of DOX as assessed by extracellular metabolite levels. DOX responsive metabolic profiling of intracellular metabolites could be conducted in hiPSC-CMs to detect early toxicity metabolite signatures. On the other hand hiPSC-CMs induced distinct changes in the concentration level of certain extracellular metabolites in response to repeated exposure to DOX.
High energy demands of normal beating heart largely depend on a steady supply of pyruvate and fatty-acyl coenzyme A (Kolwicz et al. 2013). Acetate is also an important source of acetyl Coenzyme A (acetyl-CoA) and plays a significant role in maintaining energy homeostasis in mammalian cells (Shimazu et al. 2010). Acetyl-CoA is primarily utilized in anabolic processes such as fatty acid biosynthesis and an energy generating catabolic process; tricarboxylic acid (TCA) cycle (Gaspar et al. 2014). In the mitochondria, pyruvate and acetate derived acetyl-CoA, enters into the TCA cycle to meet energy requirements (Gaspar et al. 2014). In this study, decreased uptake of pyruvate and acetate from the culture media was observed after repeated exposure to DOX. This may reflect reduced ability of mitochondria to metabolize pyruvate and acetate to generate energy. Studies in rat-heart mitochondria indicated that the rate of pyruvate transport was inhibited by DOX (Paradies and Ruggiero 1988). However, during drug washout the reversible effect in the uptake of pyruvate suggests DOX dependent disturbances in the uptake/utilization of pyruvate. Interestingly, hiPSC-CMs repeatedly exposed to DOX exhibit irreversible effect in the uptake of acetate during drug washout which remained at high levels in culture media. In addition, hiPSC-CMs given single exposure to DOX showed reduction in the uptake of acetate, as evidenced by higher levels of acetate compared to controls (not statistically significant). Our data suggest that DOX induced disturbances in acetate relevant metabolic energy pathways. Acetate utilization and production could play important role in ageing process too (Shimazu et al. 2010). In this context, a previous in vivo study (Andreadou et al. 2009) utilized NMR spectroscopy to demonstrate significantly increased acetate (and succinate) levels in heart tissue exposed to DOX and the authors proposed these intracellular metabolites as a novel biomarker for DOX induced cardiotoxicity.
Formate is produced in the mitochondria by folate-mediated metabolic processes, and it is utilized in purine biosynthesis (Lamarre et al. 2013; Morrow et al. 2015). However, our knowledge about formate metabolism in the context of cardiac metabolism is limited. Formate, the only measured metabolite produced (effluxed) by hiPSC-CMs, was irreversibly affected during repeated exposure to DOX and after removal of DOX (drug washout). Our data gave evidence that DOX inhibits the folate-mediated mitochondrial production of formate. Therefore, changes in the formate metabolism may represent an early indicator of general mitochondrial damage. Reduced accumulation of formate during drug washout suggested a prolonged effect of DOX on the formate production and anabolic processes such as purine biosynthesis. However, the reduced production of formate by DOX may also occur due to an inhibition of one or more transporters responsible for the formate efflux. ATP molecules are mostly synthesized in the mitochondria and used as energy source in various cellular functions. DOX induced depletion in ATP levels was reported in rat and mice cardiomyocytes (Octavia et al. 2012; Pointon et al. 2010). Consistent with these observations, we demonstrated that DOX induced a decline in ATP levels in hiPSC-CMs. In general, the ATP depletion and mitochondrial dysfunction induced by DOX may be responsible for the DOX induced cardiotoxicity. In this context, impaired ATP production in mitochondria has been linked to cardiac hypertrophy and heart failure (Ingwall and Weiss 2004; Rosca et al. 2013).
Conclusions
In summary, our study demonstrates that 1H NMR spectroscopy-based profiling of extracellular metabolites can be used to identify metabolic signatures associated with cardiotoxicity and cardiomyopathies. Pyruvate, acetate and formate can be proposed as candidate biomarkers to predict DOX induced physiological toxicity in the heart. Alterations in extracellular metabolite concentrations of these three metabolites may deepen our knowledge in understanding the mechanism of DOX induced cardiotoxicity. In general, the ATP depletion and mitochondrial dysfunction induced by DOX may be the main causes for the DOX induced cardiotoxicity. Our data suggest that metabolic profiling in hiPSC-CMs model system is an effective approach to strengthen preclinical cardiac safety assessment in new drug discovery and development.
Abbreviations
- Acetyl-CoA:
-
Acetyl-coenzyme A
- ATP:
-
Adenosine triphosphate
- CPMG:
-
Carr–Purcell–Meiboom–Gill
- DMSO:
-
Dimethyl sulfoxide
- DOX:
-
Doxorubicin
- DSA:
-
4,4-Dimethyl-4-silapentane-1-ammonium trifluoroacetate
- hiPSC-CMs:
-
Human induced pluripotent stem cell-derived cardiomyocytes
- iCell-MM:
-
iCell cardiomyocytes maintenance medium
- iCell-PM:
-
iCell cardiomyocytes plating medium
- LDH:
-
Lactate dehydrogenase
- MS:
-
Mass spectroscopy
- NMR:
-
Nuclear magnetic resonance
- SD:
-
Standard deviation
- TCA cycle:
-
Tricarboxylic acid cycle
References
Andreadou I, Papaefthimiou M, Zira A, Constantinou M, Sigala F, Skaltsounis AL, Tsantili-Kakoulidou A, Iliodromitis EK, Kremastinos DT, Mikros E (2009) Metabonomic identification of novel biomarkers in doxorubicin cardiotoxicity and protective effect of the natural antioxidant oleuropein. NMR Biomed 22:585–592
Chaudhari U, Nemade H, Wagh V, Gaspar JA, Ellis JK, Srinivasan SP, Spitkovski D, Nguemo F, Louisse J, Bremer S, Hescheler J, Keun HC, Hengstler JG, Sachinidis A (2016) Identification of genomic biomarkers for anthracycline-induced cardiotoxicity in human iPSC-derived cardiomyocytes: an in vitro repeated exposure toxicity approach for safety assessment. Arch Toxicol 90:2763–2777
Chow M, Boheler KR, Li RA (2013) Human pluripotent stem cell-derived cardiomyocytes for heart regeneration, drug discovery and disease modeling: from the genetic, epigenetic, and tissue modeling perspectives. Stem Cell Res Ther 4:97
Clarke CJ, Haselden JN (2008) Metabolic profiling as a tool for understanding mechanisms of toxicity. Toxicol Pathol 36:140–147
Ebert AD, Liang P, Wu JC (2012) Induced pluripotent stem cells as a disease modeling and drug screening platform. J Cardiovasc Pharmacol 60:408–416
Ellis JK, Athersuch TJ, Cavill R, Radford R, Slattery C, Jennings P, McMorrow T, Ryan MP, Ebbels TM, Keun HC (2011) Metabolic response to low-level toxicant exposure in a novel renal tubule epithelial cell system. Mol BioSyst 7:247–257
Gaspar JA, Doss MX, Hengstler JG, Cadenas C, Hescheler J, Sachinidis A (2014) Unique metabolic features of stem cells, cardiomyocytes, and their progenitors. Circ Res 114:1346–1360
Gianni L, Herman EH, Lipshultz SE, Minotti G, Sarvazyan N, Sawyer DB (2008) Anthracycline cardiotoxicity: from bench to bedside. J Clin Oncol 26:3777–3784
Ingwall JS, Weiss RG (2004) Is the failing heart energy starved? On using chemical energy to support cardiac function. Circ Res 95:135–145
Johnson CH, Ivanisevic J, Siuzdak G (2016) Metabolomics: beyond biomarkers and towards mechanisms. Nat Rev Mol Cell Biol 17:451–459
Keun HC, Athersuch TJ (2011) Nuclear magnetic resonance (NMR)-based metabolomics. Methods Mol Biol 708:321–334
Kolwicz SC Jr, Purohit S, Tian R (2013) Cardiac metabolism and its interactions with contraction, growth, and survival of cardiomyocytes. Circ Res 113:603–616
Lamarre SG, Morrow G, Macmillan L, Brosnan ME, Brosnan JT (2013) Formate: an essential metabolite, a biomarker, or more? Clin Chem Lab Med 51:571–578
Lewis GD, Asnani A, Gerszten RE (2008) Application of metabolomics to cardiovascular biomarker and pathway discovery. J Am Coll Cardiol 52:117–123
Liang P, Lan F, Lee AS, Gong T, Sanchez-Freire V, Wang Y, Diecke S, Sallam K, Knowles JW, Wang PJ, Nguyen PK, Bers DM, Robbins RC, Wu JC (2013) Drug screening using a library of human induced pluripotent stem cell-derived cardiomyocytes reveals disease-specific patterns of cardiotoxicity. Circulation 127:1677–1691
Markley JL, Bruschweiler R, Edison AS, Eghbalnia HR, Powers R, Raftery D, Wishart DS (2016) The future of NMR-based metabolomics. Curr Opin Biotechnol 43:34–40
Minotti G, Menna P, Salvatorelli E, Cairo G, Gianni L (2004) Anthracyclines: molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol Rev 56:185–229
Morrow GP, MacMillan L, Lamarre SG, Young SK, MacFarlane AJ, Brosnan ME, Brosnan JT (2015) In vivo kinetics of formate metabolism in folate-deficient and folate-replete rats. J Biol Chem 290:2244–2250
Octavia Y, Tocchetti CG, Gabrielson KL, Janssens S, Crijns HJ, Moens AL (2012) Doxorubicin-induced cardiomyopathy: from molecular mechanisms to therapeutic strategies. J Mol Cell Cardiol 52:1213–1225
Paradies G, Ruggiero FM (1988) The effect of doxorubicin on the transport of pyruvate in rat-heart mitochondria. Biochem Biophys Res Commun 156:1302–1307
Park JC, Hong YS, Kim YJ, Yang JY, Kim EY, Kwack SJ, Ryu DH, Hwang GS, Lee BM (2009) A metabonomic study on the biochemical effects of doxorubicin in rats using (1)H-NMR spectroscopy. J Toxicol Environ Health A 72:374–384
Pointon AV, Walker TM, Phillips KM, Luo J, Riley J, Zhang SD, Parry JD, Lyon JJ, Marczylo EL, Gant TW (2010) Doxorubicin in vivo rapidly alters expression and translation of myocardial electron transport chain genes, leads to ATP loss and caspase 3 activation. PLoS One 5:e12733
Robertson DG, Watkins PB, Reily MD (2011) Metabolomics in toxicology: preclinical and clinical applications. Toxicol Sci 120(Suppl 1):S146–S170
Rosca MG, Tandler B, Hoppel CL (2013) Mitochondria in cardiac hypertrophy and heart failure. J Mol Cell Cardiol 55:31–41
Sharma A, Wu JC, Wu SM (2013) Induced pluripotent stem cell-derived cardiomyocytes for cardiovascular disease modeling and drug screening. Stem Cell Res Ther 4:150
Shimazu T, Hirschey MD, Huang JY, Ho LT, Verdin E (2010) Acetate metabolism and aging: an emerging connection. Mech Ageing Dev 131:511–516
Tokarska-Schlattner M, Zaugg M, Zuppinger C, Wallimann T, Schlattner U (2006) New insights into doxorubicin-induced cardiotoxicity: the critical role of cellular energetics. J Mol Cell Cardiol 41:389–405
van Meer BJ, Tertoolen LG, Mummery CL (2016) Concise review: measuring physiological responses of human pluripotent stem cell derived cardiomyocytes to drugs and disease. Stem Cells 34:2008–2015
Wishart DS (2008) Applications of metabolomics in drug discovery and development. Drugs R D 9:307–322
Zhang S, Liu X, Bawa-Khalfe T, Lu LS, Lyu YL, Liu LF, Yeh ET (2012) Identification of the molecular basis of doxorubicin-induced cardiotoxicity. Nat Med 18:1639–1642
Zhang A, Sun H, Yan G, Wang P, Wang X (2015) Metabolomics for biomarker discovery: moving to the clinic. Biomed Res Int 2015:354671
Acknowledgements
This work was supported by the ‘Detection of endpoints and biomarkers for repeated dose toxicity using in vitro systems (DETECTIVE)’ Project (FP7 Health Programme, European Commission).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Funding
This work was supported by the ‘Detection of endpoints and biomarkers for repeated dose toxicity using in vitro systems (DETECTIVE) Project, a part of SEURAT-1 cluster.
Additional information
Handling Editors: C.-A.A. Hu, Y. Yin, Y. Hou, G. Wu, Y. Teng.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Chaudhari, U., Ellis, J.K., Wagh, V. et al. Metabolite signatures of doxorubicin induced toxicity in human induced pluripotent stem cell-derived cardiomyocytes. Amino Acids 49, 1955–1963 (2017). https://doi.org/10.1007/s00726-017-2419-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00726-017-2419-0