
Abstract
Key message
The Douglas fir provenance Three Valley was found most suitable for planting in mid-Sweden. Greenhouse tests can most likely predict how different Douglas fir provenances will perform in the field.
Abstract
The need for species that will grow well through ongoing climate change has increased the interest in Douglas fir [Pseudotsuga menziesii (Mirb.) Franco] in Sweden. One of the most common problems seen in plantations of Douglas fir seedlings is damage caused by late spring frost, known to be highly correlated with the timing of bud burst. The objective of this study was to investigate spring-related bud development under Nordic conditions of seven Douglas fir provenances and to compare data with a local provenance of Norway spruce (Picea abies (L.) Karst). Results from a field trial and a greenhouse-based study were compared. The interior Douglas fir provenances exhibited an earlier bud burst than coastal provenances, both in the greenhouse and in the field trial. When comparing differences within the groups of interior and coastal Douglas fir provenances, no differences could be found. The local Norway spruce, only grown in the greenhouse, showed an intermediate bud development profile similar to the interior Douglas fir provenance Three Valley. We therefore suggest that Three Valley could be planted at the same locations as the investigated local provenance of Norway spruce in mid-Sweden. To avoid spring frost damage the Douglas fir seedlings need to be frozen stored and planted late in spring. Planting under shelterwood can also help protect the seedlings from spring frost damages. As similar results for bud development patterns of Douglas fir and Norway spruce provenances were obtained from the greenhouse and field trials, greenhouse tests could facilitate selection of provenances.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The need to find suitable species to grow through ongoing climate change (IPCC 2014) has increased interest in Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) in Sweden. The most common tree species in Sweden, Norway spruce (Picea abies L. Karst) is one of the species that will have difficulty coping with higher temperatures and increased risk of wind damage (Spiecker et al. 1996). Douglas fir is attractive, because its commercially valuable wood (Hermann and Lavender 1999) and potentially fast growth (Karlberg 1961; Nord-Larsen et al. 2009). The species also has a wide site adaptability, and capacity to adapt to changing environmental conditions (Isaac-Renton et al. 2014). Douglas fir has been grown, to a small extent, in the very south of Sweden since the early 1900s (Martinsson and Winsa 1986; Lemoine and Wirten 1988). However, more extensive use has been hindered by, among other reasons, access to seedlings of suitable provenances and concern about damage by agents such as pine weevil (Hylobius abietis) and frost (Witte 1948; Kardell 2013, Wallertz and Malmqvist 2013; Wallertz et al. 2014).
Frost damage to seedlings is a well-known problem when growing Douglas fir in Sweden as well as its natural range (Newsome et al. 1990; Wallertz et al. 2013). The susceptibility to frost, both in spring and autumn, differs widely among provenances (Snäll 2000; O’Neill et al. 2001; Hansen 2007). In Europe, excluding the most northern parts, provenance trials and practical experience cover mostly coastal provenances. In a future climate these experiences would be advantageous for more northern growing sites (Eilmann et al. 2013; Isaac-Renton et al. 2014; Chakraborty et al. 2016). In Norway, Finland and Sweden, provenance trials found the interior provenances to be superior to the coastal (Kurkela 1981; Magnesen 1987; Martinsson and Kollenmark 2001). Since spring frost hardiness and bud burst occurrence are highly correlated (Howe et al. 2003; Søgaard et al. 2008), the time of bud burst is a useful indicator of a species suitability for a particular location (Hannerz 1999; Howe et al. 2003; Anekonda et al. 2004; Gould et al. 2011). Christophe and Birot (1979) and Edman (1997) were unable to show a correlation between bud burst occurrence and latitude or elevation of Douglas fir provenances. Instead, Edman (1997) found a strong correlation between longitude and bud burst occurrence; families from the most eastern parts of British Columbia showed an earlier bud burst in spring than those from the western parts, also shown earlier by, e.g., White et al. (1979). However, Lavadinovic et al. (2009) concluded that increasing elevation of Douglas fir results in earlier bud burst, but this is unaffected by latitude and longitude. The climate of Pacific Northwest America, where Douglas fir provenances suitable for Nordic conditions grow, differs significantly between regions (Hermann and Lavender 1990). It varies from extreme maritime in coastal areas to continental in the Rocky Mountains with an intermediate climate between those regions. The climate in southern Sweden is characterised by mild winters, occasionally interrupted by cold periods caused by continental weather systems from the east or north. In winter and spring, this can make the temperature periodically shift between being above-zero and sub-zero over a few hours, increasing the risk of frost damage (SMHI 2016).
As temperatures rise during spring, many biological processes start in plants, for example bud burst in conifers, which could be described as the start of the growing season (Sarvas 1972). The annual process of bud development includes several stages, from bud set in autumn to bud burst in spring the following year. For conifers, the most important parameter initiating bud set is an increase in night length. The “critical night length” (when 50% of a population starts to set bud) depends on the origin or provenance of the trees (Ekberg et al. 1979; Dormling and Lundkvist 1983). The temperature during bud set is known to affect bud size as well as the time of bud burst the following spring (Dormling et al. 1968; Grossnickle 2000). After termination of bud set, seedlings start to build up their cold hardiness (Bigras et al. 2001) and enter a state of dormancy (Dormling et al. 1968; Ekberg et al. 1979; Fuchigami and Nee 1987), where no apical shoot growth occurs. About 15–30 days of chilling is required for Norway spruce seedlings to break the dormancy state, and in southern Scandinavia, this condition has normally been met by November or December (Hannerz et al. 2003). The next step in the annual growth cycle is bud burst, a process that starts after some weeks with temperatures ≥+5 °C and, thereafter, a period with increased temperature >+5 °C (Sarvas 1972; Cannell and Smith 1983; Hannerz 1999). When the number of accumulated degree days (daily mean temperature >5 °C) reaches a certain threshold, specific for different species of conifers, buds start growing (Grossnickle 2000). The sum of accumulated temperature (T Sum) is commonly used as a measure for describing the difference in time for bud burst among different temperate tree species (Cannell and Smith 1983) as well as between provenances of, for example, Norway spruce (Hannerz 1994b; Morén and Perttu 1994; Hannerz 1999).
Terminal buds of Douglas fir seedlings, from many seed sources, have a chilling requirement of about 50 days at 0–5 °C (Campbell 1974, Grossnickle 2000) to break dormancy. Campbell and Sugano (1979) found that interior provenances of Douglas fir have a lower chilling requirement than coastal provenances, the same trend as in Silver birch and Scots pine (Leinonen 1996). When dormancy is broken, temperatures higher than +5 °C force bud burst through accumulation of heat units (Bailey and Harrington 2006), and accumulated heat sums can be used to describe geographical variations in the timing of bud burst if the chilling requirements have been met. Hannerz et al. (2003) reported that provenances of Norway spruce from Sweden had a shorter chilling requirement compared to provenances from Denmark and Germany.
Previously, it has been shown that freezing tolerance in autumn varies among different provenances of Douglas fir (Malmqvist et al. 2016). For successful establishment of Douglas fir seedlings under Nordic conditions, they must be able to withstand early spring frost as well as autumn frost. As little is known about how spring-related bud development of Douglas fir provenances are affected by Nordic conditions, the objective of this study was (1) to investigate spring-related bud development of seven Douglas fir provenances, (2) to compare data with the native Norway spruce (Picea abies (L.) Karst) to relate the two species to each other, and 3) to study the freezing tolerance of buds at different development stages.
Materials and methods
Seven provenances of Douglas fir from British Columbia, Canada, and one Swedish provenance of Norway spruce were used in a greenhouse study. Six Douglas fir provenances were used for the field study. Table 1 shows the details of each provenance. The greenhouse study was carried out at the Vassbo research station, Sweden (60°31′N, 15°31′E), and the field study near the Asa Experimental forest and research station, Sweden (57°10′N, 14°45′E).
The greenhouse study
At the Vassbo research station, seeds were sown on May 13, 2013 in 90 ml containers, 40 cavities per container tray (Svepot Air 90, Svenska Skogsplantor AB, Sweden). The seedlings were grown in a greenhouse at mean day/night temperatures of 25/20 °C until June 28 (Fig. 1). After that, the seedlings were moved outdoors. On September 25, 2013, the seedlings were moved back into the greenhouse, which allowed natural night length, with the air temperature set to a minimum of +7 °C to avoid sub-zero temperatures. The air temperatures in a ventilated radiation shield at 1.3 m above ground outside and 1.6 m above ground in the greenhouse (Fig. 1) were measured and logged every 5 min, using an automated data logger (Campbell Scientific CR1000, UK). Daily values were based on the mean of 15 min averages for 24 h from midnight to midnight. From March 12 to May 19, 2014, bud development of the terminal bud was measured once a week on eight randomly selected seedlings in each of four container trays. The four container trays of each provenance were randomly distributed. The T Sum were calculated using the threshold value +5 °C in accordance with Hannerz (1994a):
where T Sum is the temperature sum in °C and T i is the daily mean temperature in °C.

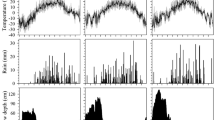
Air temperature, daily average, maximum and minimum temperatures recorded from May 8, 2013 to July 28, 2014 (above). From May 8, 2013 to June 28, 2013, the temperature was measured inside the greenhouse and from June 29, 2013 to September 25, 2013, it was measured at the outdoor holding area. Greenhouse temperatures were logged as the seedlings were moved indoors on September 25, 2013. The temperature sums (lower graph), that is the sum of daily mean temperatures exceeding + 5 °C from January 1, 2014 to July 31, 2014, were calculated in accordance with Hannerz (1994a)
T Sum in the greenhouse were calculated from January 1, 2014 (Fig. 1).
The field study
The field site near the Asa research station is included in a large survey study of survival and damage of Douglas fir in Sweden, described by Wallertz et al. (2013). The site was planted with 2-year-old Douglas fir seedlings of six different provenances (Table 1) in May 2009, after site preparation the same spring. The following year, the bud development of the terminal bud was measured once a week on 20 healthy seedlings for each provenance, i.e., seedlings with visible damage were excluded at the first inventory. The seedlings were randomly selected among the seedlings measured in the survey study (Wallertz et al. 2013). For one of the provenances, Caycuse River, only 10 healthy seedlings could be found and measured. Air temperatures were recorded with a data logger (Campbell Scientific CR10, UK) from the beginning of January until the end of July 2010 at the Asa research station, 6 km from the field study site. The air temperature (°C) was measured at 1.7 m above ground in a ventilated radiation shield in an open clearing. Daily values were based on 10 min averages using a 60 s scan interval, using the mean value from midnight to midnight for each day. T Sum were calculated from the beginning of the growing season, defined by Morén and Perttu (1994) as 4 days in a row with a daily mean temperature above +5 °C. This condition was met on April 25, 2010 (Fig. 2).

Air temperature, daily average, maximum and minimum temperatures recorded from January 1, 2010 to July 31, 2010 (above). The temperature sums (lower graph), that is the sum of daily mean temperatures exceeding + 5 °C from April 25, 2014 to July 31, 2014, were calculated in accordance with Hannerz (1994a)
Bud development in the greenhouse and in the field
Bud development was assessed using the Krutzsch index (Krutzsch 1973). The index was originally designed for Norway spruce, but was in this study used for Douglas fir, as well (Fig. 3). The reason for using the Krutzsch index instead of scales specifically developed for Douglas fir (Thomson and Moncrieff 1982; Bailey and Harrington 2006; Harrington et al. 2010) was that we had a special interest in comparing the early bud burst patterns of Norway spruce and Douglas fir, which are included in the Krutzsch index (stages 1 and 2) but not in the specific Douglas fir scales.

Classification of bud development stages was used for both Norway spruce and Douglas fir Krutzsch (1973). Stages 1–8 (shown in these pictures for Douglas fir) are defined and scored as follows: 1 buds slightly swollen, 2 buds swollen, green to grey–green in colour, bud scales still closed, 3 burst of bud scales, tips of needles emerging, 4 needles elongated to about double the bud length, 5 first spread of needles, buds now have the appearance of a painter’s brush, 6 elongation of shoot, basal needles not yet spread, 7 differentiation of shoot, basal needles spread, 8 all needles more or less spread, new buds developing. The first development stage 0, a dormant bud, is not shown
Freezing tolerance of buds
On April 7, 2014, the freezing tolerance of the buds prior to bud burst (stages 1 and 2 of the Krutzsch index) was tested by measuring the electrolyte conductance caused by freezing to −5 °C. The freezing test, carried out on the local Norway spruce, interior Douglas fir provenance Three Valley, and coastal Douglas fir provenance Ladysmith (Table 1), was based on methods described by Lindström et al. (2014) and Malmqvist et al. (2016). In this study, only buds at stages 1 and 2 were frozen. The buds were randomly collected from seedlings grown in the same container trays as the seedlings designated for the greenhouse study of bud development measurements. To prepare three replicates of each bud development stage, 30 buds of each development stage and seed origin were picked from the upper half of the seedlings. Each sample contained 10 buds. The samples were slowly frozen (2.5 °C h−1) to the target temperature of −5 °C. After 1 h at −5 °C, the samples were slowly thawed (2.5 °C h−1) to +5 °C. To each sample, 40 ml of deionised water were added, and thereafter, the samples were put in a shaker for 24 h. The electrolytic conductivity was measured using a conductivity meter (Model Hach SensIon 5). To calculate the electrolytic leakage from cells caused by freezing the buds to −5 °C, the samples were autoclaved and the total electrolytic leakage measured. The electrolytic leakage caused by freezing the buds to −5 °C was calculated as the difference between the total electrolytic leakage of frozen buds and the natural leakage from unfrozen buds (see Lindström et al. 2014).
Statistical analyses
Differences in bud development between provenances at certain times were carried out using Fisher’s exact test (Zar 2010). Fisher’s exact test was used due to a small sample size in one of the provenances in the field study and it also fitted the experimental design in the greenhouse study. Differences in freezing tolerance of buds were statistically analysed using the independent samples t test with equal variances assumed according to Leven’s test for equality of variances. The analyses were carried out using IBM SPSS Statistics, version 23.
Results
Bud development in the greenhouse
Bud burst appeared earlier for the interior Douglas fir provenances compared to both the coastal Douglas fir provenances and Norway spruce (Table 2). On April 1, 2014, the bud burst (reached or passed stage 3 of the Krutzsch index) of the interior Larch Hills was 81%, Anstey Arm 71%, and Three Valley 38%, while, among the coastal provenances, the proportion of seedlings that showed bud burst ranged between 5 and 14% at this date. Norway spruce showed an intermediate timing of bud burst with 28% of the seedlings reaching or passing stage 3 on April 1 (Fig. 4).

Proportion (%) of greenhouse-grown seedlings in different spring-related bud development stages, using the Krutzsch index (1973), for the seven Douglas fir provenances and Norway spruce in spring 2014, on March 12, upper left, March 19, upper right, March 25, middle left, April 1, middle right, April 10, bottom left, and April 16, bottom right. T Sum in degree days (dd) indicated for each date. T Sum accumulated from January 1. N = 32
One week later, on April 8, more than 80% of all seedlings of the interior Douglas fir provenances had burst buds (reached or passed stage 3). By mid-April, all provenances as well as Norway spruce had a proportion of 80% or more of seedlings that exhibited bud burst (Fig. 4).
Bud development in the field
When the field measurements started on May 19, 2010, more than 50% of the seedlings from all provenances had reached or passed stage 1, indicating that bud development had started (Fig. 5). No seedlings of coastal origin exhibited bud burst at this date, in contrast to the interior provenances, where the percentages exhibiting bud burst were Larch Hills 45%, Anstey Arms 35%, and Three Valley 13%. One week later, on May 26, all provenances except Caycuse River had a high proportion of seedlings which showed bud burst and many of them had begun their shoot elongation. On May 26, the differences in bud burst patterns between coastal and interior provenances were significant (Table 3).

Proportion (%) of field-grown seedlings in different spring-related bud development stages, according to the Krutzsch index (1973), for six Douglas fir provenances (Table 1) in late spring 2010, on May 19, upper left, May 26, upper right, June 2, bottom left, and June 10, bottom right. T Sum in degree days (dd) indicated for each date. T Sum accumulated from the date (April 25) when the vegetation period started according to Morén and Perttu (1994). N = 10–20
Summer bud set in the field
The seedlings in the field study were monitored until they reached stage 8 of the Krutzsch index (stage 8: all needles more or less spread, new buds developing). The interior provenances formed their new buds earlier in comparison to the coastal provenances. On June 24, 2010, more than 70% of the seedlings of the interior provenances had set bud, while the coastal provenances did so later. Among the coastal provenances, no bud set was recorded for Caycuse River at this date, while 40% of the Bella Coola and 55% of Ladysmith had set bud (Fig. 6).

Proportion (%) of field-grown seedlings that reached stage 8 of the Krutzsch index (all needles more or less spread, new buds developing) (Krutzsch 1973), during June and early July 2010 at the Asa experimental site. N = 10–20
Freezing tolerance of buds
The freezing test of buds to −5 °C in early spring showed that the buds of Norway spruce and Douglas fir were already sensitive to freezing when they were slightly swollen (stage 1, see Fig. 3). The buds of the Norway spruce and Douglas fir provenances (Ladysmith, coastal, and Three Valley, interior) were severely damaged after freezing to −5 °C, as electrolytic leakage from freezing exceeded 50% on average for all tested treatments. The average electrolyte leakage of unfrozen control samples of Norway spruce was 41% and for Douglas fir 28%. No significant differences regarding electrolyte leakage after freezing were found between species (p = 0.258) or between the two provenances of Douglas fir (p = 0.650). However, for buds that had reached stage 2 of bud development (stage 2, see Fig. 3), the electrolytic leakage was significantly higher than for buds that had reached stage 1, regardless of species or provenance (p = 0.035).
Discussion
In our study, the interior Douglas fir provenances showed an earlier bud burst in spring than the coastal provenances, which was in agreement with earlier findings (Campbell and Sugano 1979; Edman 1997). This trend could be compared with differences among Norway spruce seedlings where northern provenances showed earlier bud burst compared to southern provenances (Hannerz et al. 2003). Bud burst patterns were similar for the local Norway spruce and the interior Douglas fir provenances in the greenhouse study. Of the interior Douglas fir provenances, Anstey Arms and Three Valley had a similar shoot development pattern during spring. Apart from bud burst, a previous study by Malmqvist et al. (2016) showed similar timing of bud set by the Three Valley provenance and the local Norway spruce. In addition, Malmqvist et al. (2016) showed that the development of freezing tolerance in autumn occurred at the same time for these specific provenances of Norway spruce and Douglas fir. The similar timing of the start and termination of growth, as well as the timing of the winter hardening processes, both suggests that the Three Valley provenance of Douglas fir could be successfully planted at the same locations as the local Norway spruce in mid-Sweden. This method to relate two species to each other by studying, for example, bud development in spring might be a way of predicting performance of species or provenances to a new site using knowledge from a well-known native species of local origin.
No significant differences in bud burst could be seen within the groups of interior and coastal provenances, even though the origin of the provenances differs in elevation and latitude. However, bud development had already begun at the first measurement, so more frequent observations might have been able to detect eventual differences in bud burst patterns.
If the chilling requirements have been met for dormancy release, temperature sums can be used to describe variations in the timing of bud burst among provenances of Douglas fir (Cannell and Smith 1983; Bailey and Harrington 2006) as well as for Norway spruce (Hannerz 1994a, b, 1999). When calculating the accumulated temperature sum, the threshold value of +5 °C for when the growing season starts has been commonly used by, for example, Sarvas (1972), Cannell and Smith (1983), Morén and Perttu (1994), Hannerz (1994a, b, 1999), and Hannerz et al. (2003). Hannerz (1994a) showed a sharp decrease in frost risk when bud burst was delayed from T Sum 120 dd to T Sum 180 dd. The Belarusian provenances of Norway spruce (T Sum 180 dd to burst of bud) are often used in southern Sweden to reduce damage from late spring frost. In our field study, all Douglas fir provenances except the coastal Caycuse River had a majority of the seedlings burst bud at T Sum 122, which is comparable to the local Norway spruce provenance from latitude 60° (Hannerz 1994a). Already at 68 dd, a high proportion of the interior provenances had reached stage 2 in the Krutzsch index and consequently susceptible to frost. In the greenhouse study, the accumulated temperature sum is not comparable to outdoor values, because the temperature never falls below +5 °C. The total accumulated temperature sum in the greenhouse, therefore, depends on when we decide to start measuring. In our case, we chose to start on January 1, 2014. This resulted in very high T Sum before buds started to grow, despite the early start of bud burst (mid-March) in the greenhouse study compared to the field study. A possible explanation for the high requirement of accumulated temperature before the buds started to burst in the greenhouse could be that the chilling requirement took longer to be met due to non-optimal chilling temperatures. This could have slowed bud development despite the high forcing temperatures during early spring in the greenhouse. The effect of non-optimal chilling temperatures postponing the start of bud development has previously been shown by Bailey and Harrington (2006). Harrington et al. (2010) also suggested that temperatures that are some degrees above freezing are the most effective in satisfying the chilling requirement for dormancy release, while higher autumn temperatures, as occurred in the greenhouse, would not be as effective.
Despite the differences in environmental prerequisites for seedlings in the greenhouse and the field, the time laps between the bud development stages (1–4) were approximately the same once the buds had started to grow. As the same trends and relationships between the growth of provenances during bud development can be seen in the greenhouse trial and the field trial, we suggest that greenhouse trials in some cases might replace field trials. Greenhouse trials could be used as an easy way to map differences between provenances and predict field performance. This could facilitate more rapid selection of Douglas fir provenances in Sweden.
The observations of shoot elongation and bud set during early summer showed that all provenances of Douglas fir and Norway spruce formed new buds. This early bud set suggests a possible further growth, appearing as Lammas growth or free growth (von Wühlisch and Muhs 1986), which can make seedlings more susceptible to frost damage during autumn and winter (Anekonda et al. 1998).
In spring, during bud burst, Langvall and Ottosson Löfvenius (2002) observed shoots of Norway spruce to be sensitive to frost when newly flushed (from stage 3 of the Krutzsch index). For Douglas fir, Aitken and Adams (1997) showed that spring frost both killed buds and also severely damaged newly flushed shoots. In our study, buds from Norway spruce as well as Douglas fir were already very sensitive to sub-zero temperatures when slightly swollen (from stage 1 of the Krutzsch index), indicating that the period of high sensitivity to late spring frost starts at the earliest bud development stages. The freezing test showed high leakage values from freezing buds of both species and both provenances of Douglas fir in their earlier bud development stages (1–2), and the leakage increased significantly between stages 1 and 2. We also noticed high leakage levels from unfrozen control samples that could be due to leakage caused by cutting off the buds from the stem. The open surface area in the cross section will be proportionally large compared to cutting the top 2 cm of the shoot, which is a common procedure when using the SELdiff-25-method (Lindström et al. 2014). Usually, the natural leakage from unfrozen controls in this latter case varies between 5 and 10% which should be compared with 28–41% natural leakage from unfrozen buds in this study. Even though we have had high natural leakage values from buds frozen to −5 °C, increased leakage values definitely indicate severe damage from freezing.
Since climate change might increase the frequency of temperature backlashes causing frost damage in spring (Jönsson et al. 2004; Langvall 2011), it is increasingly important to consider the bud development patterns of provenances when choosing seed sources. In addition, actions to protect seedlings at the planting site should be considered. Silvicultural measures such as use of shelterwood systems, planting on slopes, and mechanical site preparation can reduce the damage from late spring frost and increase seedling survival (Newsome et al. 1990; Langvall 2000; Langvall and Örlander 2001; Nilsson et al. 2010). The use of frozen stored seedlings planted late in spring will diminish the risk of serious damage from late spring frost the first year, and the years to come, the increasing tree height will conduce towards reduced risk. At a tree height taller than 1–2 m, the apical shoot and upper branches are above the coldest air (Morén and Perttu 1994).
Author contribution statement
CM conceived the experiment. CM and EW designed and carried out the experiment as well as analysed the data, conducted the statistical analyses, and wrote the paper with support from AL and HS. CM, 45%, EW, 40%, AL, 10%, and HS, 5%.
References
Aitken SN, Adams WT (1997) Spring cold hardiness under strong genetic control in Oregon populations of Pseudotsuga menziesii var. menziesii. Can J For Res 27(11):1773–1780. doi:10.1139/x97-151
Anekonda TS, Adams WT, Aitken SN (1998) Influence of second flushing on genetic assessment of cold hardiness in coastal Douglas-fir (Pseudotsuga menziesii var. menziesii (Mirb.) Franco). For Ecol Manage 111:119–126
Anekonda T, Jones C, Smith BN, Hansen LD (2004) Differences in physiology and growth between coastal and inland varieties of Douglas-fir seedlings in a common garden. Thermochim Acta 422:75–79. doi:10.1016/j.tca.2004.05.036
Bailey JD, Harrington CA (2006) Temperature regulation of bud-burst phenology within and among years in a young Douglas-fir (Pseudotsuga menziesii) plantation in western Washington USA. Tree Physiol 26:421–430. doi:10.1093/treephys/26.4.421
Bigras FJ, Ryyppö A, Lindström A, Stattin E (2001) Cold acclimation and deacclimation of shoots and roots of conifer seedlings. In: Bigras FJ, Colombo SJ (eds) Conifer cold hardiness. Kluwer Academic, Dordrecht, pp 57–89
Campbell RK (1974) Use of phenology for examining provenance transfers in reforestation of Douglas-fir. J Appl Ecol 11(3):1069–1080
Campbell RK, Sugano AI (1979) Genecology of bud-burst phenology in Douglas-fir: response to flushing temperature and chilling. Bot Gaz 140(2):223–231
Cannell MGR, Smith RI (1983) Thermal time, chill days and prediction of budburst in Picea sithchensis. J Appl Ecol 20(3):951–963
Chakraborty D, Wang T, Andre K, Konnert M, Lexer MJ, Matulla C, Weißenbacher L, Schueler S (2016) Adapting Douglas-fir forestry in Central Europe: evaluation, application, and uncertainty analysis of a genetically based model. Eur J For Res 135:919–936. doi:10.1007/s10342-016-0984-5
Christophe C, Birot Y (1979) Genetic variation within and between populations of Douglas fir. Silvae Genet 28:197–206
Dormling I, Lundkvist K (1983) Vad bestämmer skogsplantors tillväxt och härdighet i plantskolan? Skogsfakta nr 8. Swedish University of Agricultural Sciences, Garpenberg (in Swedish)
Dormling I, Gustavsson Å, von Wettstein D (1968) The experimental control of the life cycle in Picea abies (L.) Karst. 1. Some basic experiments on the vegetative cycle. Silvae Genet 17:44–64
Edman M (1997) The phenology of the interior Douglas fir. Examensarbete i ämnet skogsskötsel 1997–1. Institutionen för skogsskötsel SLU Swedish University of Agricultural Sciences Department of Forest Ecology and Management
Eilmann B, de Vries SMG, den Ouden J, Mohren GMJ, Sauren P, Sass-Klaassen U (2013) Origin matters! Differens in drought tolerance and productivity of coastal Douglas-fir (Pseudotsuga menziesii (Mirb.) provenances. For Ecol Manag 302:133–143. doi:10.1016/j.foreco.2013.03.031
Ekberg I, Eriksson G, Dormling I (1979) Photoperiodic reactions in conifer species. Holarct Ecol 2:255–263
Fuchigami LH, Nee CC (1987) Degree growth stage model and rest-breaking mechanisms in temperate woody perennials. Hort Sci 22(5):836–845
Gould PJ, Harrington CA, St Clair JB (2011) Incorporating genetic variation into a model of bud burst phenology of coast Douglas-fir (Pseudotsuga menziesii var. menziesii). Can J For Res 41(1):139–150. doi:10.1139/X10-191
Grossnickle SC (2000) Ecophysiology of northern spruce species—the performance of planted seedlings. NRC Research, Canada
Hannerz M (1994a) Damage to Norway spruce (Picea abies (L.) Karst) seedlings caused by a late spring frost. Forestry Research Institute of Sweden, Uppsala (report 5)
Hannerz M (1994b) Predicting the risk of frost occurrence after budburst of Norway spruce in Sweden. Silva Fenn 28(4):243–249
Hannerz M (1999) Evaluation of temperature models for predicting bud burst in Norway spruce. Can J For Res 29(1):9–19. doi:10.1139/x98-175
Hannerz M, Ekberg I, Norell L (2003) Variation in chilling requirements for completing bud rest between provenances of Norway spruce. Silvae Genet 52(3–4):161–168
Hansen JK (2007) Dyrkningssikker Douglasgran—en evaluering af forsög med douglasgranfrökilder. University of Copenhagen Forestry & Landscape, Copenhagen (in Danish)
Harrington CA, Gould PJ, St. Clair JB (2010) Modeling the effects of winter environment on dormancy release of Douglas-fir. For Ecol Manage 259:798–808. doi:10.1016/j.foreco.2009.06.018
Hermann RK, Lavender DP (1990) Pseudotsuga menziesii (Mirb.) Franco. In: Burns RM, Russell M, Honkala BH (Technical coordinators) Silvics of North America, vol 1. Conifers. US Department of Agriculture, Agricultural Handbook, Washington DC, p 654, pp 527–554
Hermann RK, Lavender DP (1999) Douglas-fir planted forests. New For 17:53–70
Howe GT, Aitken SN, Neale DB, Jermstad KD, Wheeler NC, Chen THH (2003) From genotype to phenotype: unravelling the complexities of cold adaption in forest trees. Can J Bot 81(12):1247–1266. doi:10.1139/b03-141
IPCC (2014) Climate change 2014: synthesis report. In: Core Writing Team, Pachauri RK, Meyer LA (eds) Contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change. IPCC, Geneva, p 151
Isaac-Renton MG, Roberts DR, Hamann A, Spiecker H (2014) Douglas-fir plantations in Europe: a retrospective test of assisted migration to address climate change. Glob Change Biol 20:2607–2617. doi:10.1111/gcb.12604
Jönsson AM, Linderson M-L, Stjernquist I, Schlyter P, Bärring L (2004) Climate change and the effect of temperature backlashes causing frost damage in Picea abies. Global Planet Change 44:195–207. doi:10.1016/j.gloplacha.2004.06.012
Kardell L (2013) Åkermarksplanteringar på Tagel, Steninge och Em 1998–2012 The Swedish University of Agricultural Sciences Department of Environmental Forestry Report, ISRN SLU-SLV-R-118-SE. ISSN 1101-0525 (in Swedish)
Karlberg S (1961) Development and yield of Douglas fir (Pseudotsuga taxifolia (Pior.) Britt.) and Sitka spruce (Picea sitchensis (Bong.) Carr.) in southern Scandinavia and on the Pacific Coast. Bulletin of the Royal School of Forestry 34, Stockholm
Krutzsch P (1973) Norway spruce development of buds. Internal report. IUFRO S2.02.11, Vienna
Kurkela T (1981) Growth reduction in Douglas fir caused by Rhabdocline needle cast. Commun Inst For Fenn 102:1–16
Langvall O (2000) Interactions between near-ground temperature and radiation, silvicultural treatments and frost damage to Norway spruce seedlings. Swed Univ Agric Sci Acta Universitatis Agriculturae Sueciae Silvestria 330:1–34
Langvall O (2011) Impact of climate change, seedling type and provenance on the risk of damage to Norway spruce (Picea abies (L.) Karst.) seedlings in Sweden due to early summer frosts. Scand J For Res 26:56–63. doi:10.1080/02827581.2011.564399
Langvall O, Ottosson Löfvenius M (2002) Effect of shelterwood on nocturnal near-ground temperature, frost injury risk and budburst date of Norway spruce. For Ecol Manage 168:149–161. doi:10.1016/S0378-1127(01)00754-X
Langvall O, Örlander G (2001) Effects of pine shelterwoods on microclimate and frost damage to Norway spruce seedlings. Can J For Res 31(1):155–164. doi:10.1139/x00-149
Lavadinovic V, Isajev V, Markovic N, Rakonjak L, Lucic A (2009) Douglas-fir provenance differentiation based on bud burst. Lucrările sesiunii ştiinţifice bienale cu participare internaţională Pădurea şi Dezvoltarea Durabilă Braşov, 17–18 octombrie 2008, pp 49–54
Leinonen I (1996) Dependence of dormancy release on temperature on different origins of Pinus sylvestris and Betula pendula seedlings. Scand J For Res 11(1–4):122–128. doi:10.1080/02827589609382919
Lemoine C, Wirten H (1988) Douglasgran i Sverige. SLU, Inst. för skogsskötsel, examensarbete, 1988-1 Swedish University of Agricultural Sciences Department of Forest Ecology and Management (in Swedish)
Lindström A, Stattin E, Gräns D, Wallin E (2014) Storability measures of Norway spruce and Scots pine seedlings and assessment of post-storage vitality by measuring shoot electrolyte leakage. Scand J For Res 29(8):717–724. doi:10.1080/02827581.2014.977340
Magnesen S (1987) The international short term Douglas fir (Pseudotstuga menziesii) provenance experiment in west Norway. In: FBVA Berichte Proceedings of the IUFRO Working Party on Breeding Strategi for Douglas-fir as an Introduced Species 21
Malmqvist C, Wallertz K, Lindström A (2016) Storability and freezing tolerance of Douglas fir and Norway spruce seedlings grown in mid-Sweden. Scand J For Res. doi:10.1080/02827581.2016.1183704
Martinsson O, Kollenmark R (2001) Growth and survival in family tests of Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) eight or nine years after establishment in southern and central Sweden. Swedish University of Agricultural Sciences Working paper 168 (in Swedish with English summary)
Martinsson O, Winsa H (1986) Främmande trädslag i svenskt skogsbruk. Swedish University of Agricultural Sciences, Faculty of Forest Sciences, Report (in Swedish)
Morén AS, Perttu KL (1994) Regional temperature and radiation indices and their adjustments to horizontal and inclined forest land. Stud For Suec 194:1–19
Newsome TA, Sutherland DC, Vyse A (1990) Establishing Douglas fir plantations in the dry belt of interior British Columbia. In: Interior Douglas-fir, the species and its management, symposium proceedings
Nilsson U, Luoranen J, Kolström T, Örlander G, Puttonen P (2010) Reforestation with planting in northern Europe. Scand J For Res 25:283–294. doi:10.1080/02827581.2010.498384
Nord-Larsen T, Meilby H, Skovsgaard JP (2009) Site-specific height growth models for six common tree species in Denmark. Scand J For Res 24:194–204. doi:10.1080/02827580902795036
O’Neill GA, Adams WT, Aitken SN (2001) Quantitative genetics of spring and fall cold hardiness in seedlings from two Oregon populations of coastal Douglas fir. For Ecol Manage. doi:10.1016/S0378-1127(00)00564-8
Sarvas R (1972) Investigations on the annual cycle of development of forest trees. Active period. Communicationes Instituti Forestalis Fennaie 76.3. Helsinki
SMHI (2016) Sveriges klimat. http://www.smhi.se/kunskapsbanken/klimat/sveriges-klimat-1.6867. Accessed 9 Nov 2016 (in Swedish)
Snäll K (2000) Frosttolerans hos douglasgran av olika ursprung och tillväxttyp. Frost tolerance of Douglas fir from different origins. Degree project in mathematical statistics. Uppsala University, Department of Mathematics (in Swedish)
Søgaard G, Johnsen Ö, Nilsen J, Junttila O (2008) Climatic control of bud burst in young seedlings of nine provenances of Norway spruce. Three Physiol 28(2):311–320. doi:10.1093/treephys/28.2.311
Spiecker H, Mielikäinen K, Köhl M, Skovsgaard J eds. (1996) Growth trends in European forests. European Forest Institute Research Report 5. Springer, Berlin
Thomson AJ, Moncrieff SM (1982) Prediction of bud burst in Douglas-fir by degree-day accumulation. Can J For Res 12(2):448–452. doi:10.1139/x82-069
von Wühlisch G, Muhs HJ (1986) Influence of age on sylleptic and proleptic free growth of Norway spruce seedlings. Silvae Genet 35:1–7
Wallertz K, Malmqvist C (2013) The effect of mechanical site preparation methods on the establishment of Norway spruce (Picea Abies (L.) Karst.) and Douglas fir (Pseudotsuga Menziesii (Mirb.) Franco) in southern Sweden. Forestry 86:71–78
Wallertz K, Frisk J, Johansson U, Örlander G (2013) Odlingstester och proveniensförsök med douglasgran i Södra Sverige. Dokumentation av etablering och tidig utveckling för planteringar 2009 och 2010. Swedish University of Agricultural Sciences. Enheten för skoglig fältforskning. Report 6. Asa (in Swedish with English summary)
Wallertz K, Nordenhem H, Nordlander G (2014) Damage by the pine weevil Hylobius abietis to seedlings of two native and five introduced tree species in Sweden. Silva Fenn 48:1–14. doi:10.14214/sf.1188
White TL, Ching KK, Walters J (1979) Effects of provenance, years, and planting location on bud burst of Douglas fir. For Sci 25:161–167
Witte A (1948) Skadegörelse på Douglas-gran. Svenska Skogsvårdsföreningens Tidskrift 46:10–18 (in Swedish)
Zar JH (2010) Biostatistical analysis, 5th edn. Pearson Education, Upper Saddle River
Acknowledgements
We wish to thank Marianne Vemhäll for all her help with laboratory work. In addition, we wish to thank Johan Henriksson and Göran Örlander at Södra for providing experimental seeds. We also want to thank Claudia von Brömssen at SLU for comments and support regarding the statistics, and Claes Hellqvist who helped us with tables and gave us good advice. The comments from anonymous reviewers were gratefully received. We also thank SITES Asa and the Reference climate monitoring programme at SLU experimental forests for providing the climatic data.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by M. Buckeridge.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Malmqvist, C., Wallin, E., Lindström, A. et al. Differences in bud burst timing and bud freezing tolerance among interior and coastal seed sources of Douglas fir. Trees 31, 1987–1998 (2017). https://doi.org/10.1007/s00468-017-1603-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-017-1603-x