
Abstract
This review on platelet research focuses on defects of adhesion, cytoskeletal organisation, signal transduction and secretion. Platelet defects can be studied by different laboratory platelet functional assays and morphological studies. Easy bruising or a suspected platelet-based bleeding disorder is of course the most obvious reason to test the platelet function in a patient. However, nowadays platelet research also contributes to our understanding of human pathology in other disciplines such as neurology, nephrology, endocrinology and metabolic diseases. Apart from a discussion on classical thrombopathies, this review will also deal with the less commonly known relation between platelet research and disorders with a broader clinical phenotype. Classical thrombopathies involve disorders of platelet adhesion such as Glanzmann thrombastenia and Bernard-Soulier syndrome, defective G protein signalling diseases with impaired phospholipase C activation, and abnormal platelet granule secretion disorders such as gray platelet disorder and delta-storage pool disease. Other clinical symptoms besides a bleeding tendency have been described in MYH9-related disorders and Duchenne muscular dystrophy due to adhesion defects, and also in disorders of impaired Gs signalling, in Hermansky Pudlack disease and Chediak Higashi disease with abnormal secretion. Finally, platelet research can also be used to unravel novel mechanisms involved in many neurological disorders such as depression and autism with only a subclinical platelet defect.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
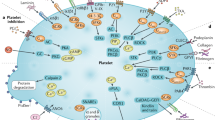
Normal hemostasis prevents spontaneous bleeding and traumatic hemorrhage by a coordinated sequence of cellular and biochemical reactions to the ultimate formation of a stable platelet-fibrin aggregate [20]. Platelets, under normal circumstances, circulate in close contact to the endothelial cell lining of the vessel wall, and respond to vascular damage by adhering to subendothelial structures. Platelet adhesion is the first step in the hemostatic plug formation [11]. The major platelet receptors involved in this process are the von Willebrand factor (vWF) receptor GP(glycoprotein)Ib/IX/V, the collagen integrin receptor α2β1, and the fibrinogen integrin receptor αIIBβ3 (Fig. 1a). Subsequent platelet spreading is conducted by cytoskeleton proteins including the structural subunit of the microtubules, the αβ-tubulin heterodimer, filamin and actin (Fig. 1a). The cytoskeleton is responsible for the shape of the resting platelet and carries out contractile events such as the secretion of granules and retraction of clots by activated cells.

a Schematic model of the main components involved in platelet adhesion and the cytoskeleton proteins. Platelet adhesion and its subsequent activation by calcium release is mainly regulated by the platelet receptor αIIbβ3 after binding to fibrinogen or the RGD domain of vWF, the main vWF receptor GPIb/IX/V and the collagen receptor α2β1. Microtubules together with the cytoplasmic, actin-rich cytoskeleton are responsible for the platelet structure. Different actin binding proteins have been identified in platelets such as filamin A, myosin and dystrophin. b Schematic model of G protein signal transduction in platelets regulated by Gq for platelet activation by the ultimate step of calcium release. Gi and Gs further influence the platelet activation by respectively inhibiting and stimulating the intracellular cAMP formation. c Schematic model of platelet secretion. The second amplification step in platelet activation is the release of alpha and dense granules in platelets guarantying irreversible platelet activation
Platelet adhesion also initiates multiple intracellular G protein-coupled signalling pathways (Fig. 1b). Stimulation of Gq by different ligands such as adenosine diphosphate (ADP) and thromboxane A2 (TXA2) results in platelet activation by stimulating phospholipase C and a release of calcium from the intracellular stores. This platelet activation process is enhanced when Gi is activated and inhibited when Gs is activated both by modulating the intracellular cAMP level.
Platelet adhesion and activation eventually results in secretion from platelet organelles (Fig. 1c) [37]. Resting platelets circulate as discoid anuclear cells and consist of a lipid bilayer and an internal dense tubular system, where calcium is sequestered. The platelet cytoplasm contains mitochondria, glycogen particles, lysosomes, and the platelet-specific storage granules: the α-granules and dense granules (Fig. 2). The α-granules contain proteins such as platelet factor 4, β-thromboglobulin, platelet derived growth factor, fibrinogen, fibronectin, thrombospondin, plasminogen activator inhibitor I and vWF. Dense bodies are rich in serotonin, ATP, ADP and calcium.

Electron microscopy (original magnification ×22,500) of platelets showing the dense tubular system (DTS), microtubules (MT), open canalicular system (OCS), alpha granules (G), glycogen (Gly) and the dense bodies (DB)
The study of platelet adhesion, G protein signalling and secretion is particularly useful for our understanding of several clinical disorders (Table 1). The most obvious reason is of course the study of platelet defects in patients with an isolated platelet disorder leading to bleeding or thrombosis in order to gain additional information on the different pathways involved in platelet function. The analysis of human disorders in which the defective platelet phenotype is just one part of its clinical spectrum to unravel novel biological and genetic mechanisms involved in the disease. Also, using platelet functional and morphological studies can be used as a tool to find novel pathways involved in more complex disorders usually caused by more than one gene defect. Recently, it became obvious that the molecular pathways involved in more complex human disorders such as diabetes type 2 or neurological disorders such as schizophrenia, migraine, bipolar disorder, and depression can also be better understood by studying platelet signalling and secretion. Due to space limitations, it is impossible to give a full overview of all human disorders studied today by means of platelet research. This review describes some well-known as well as some less common disorders to illustrate how platelet research contributes to the understanding of thrombopathies [13, 35] as well as the broader future of this research outside its classical field of thrombosis and hemostasis (Table 1).
When considering type of platelet tests
A clinical platelet-based bleeding problem is of course the main reason to investigate platelet function and morphology. The diagnostic approach to easy bruising or a suspected platelet-based bleeding disorder includes a careful history and physical examination of the patient as well as different laboratory investigations such as the Ivy bleeding time, platelet aggregation tests, ATP secretion, platelet adhesion by the platelet function analyzer (PFA100) and platelet morphology by electron microscopy [13, 40]. Reviewing the medical history can already establish whether the disorder is hereditary or acquired. The specific clinical findings useful in the differential diagnosis of coagulation versus platelet-based disorders are summarized in Table 2. Frequently, mucocutaneous bleedings characterize abnormalities of platelet function. In contrast, hemorrhage into synovial joints and deep muscular hemorrhage are signs of severe hereditary coagulation disorders and very rare events in disorders of platelets, vessels or acquired coagulation disorders. Inherited disorders of platelet function are further subdivided based on the functions or responses that are abnormal and therefore can belong to different subgroups including abnormal platelet adhesion, signalling and secretion [36]. Platelet-based bleeding disorders are usually classified according to abnormalities of platelet function, platelet number (thrombocytopenia) or both [36].
Functional and morphological platelet studies in patients with mainly a neurological, metabolic or another clinical problem but no obvious bleeding problem are usually not performed for diagnostic purposes but rather for research aims. In such patients, novel insights are expected to result from the platelet research studies, which are still preliminary today but in the future hopefully will help to better define when to ask for what type of platelet tests in a given patient.
Thrombopathies
Glanzmann thrombasthenia and the Bernard-Soulier syndrome (BSS) are two rare inherited disorders of platelet adhesion. Glanzmann thrombasthenia (MIM 273800) is an autosomal recessive disorder, characterized by prolonged bleeding time and abnormal clot retraction [30, 31, 35]. The hallmark of this disease is severely reduced or absent platelet aggregation in response to various physiological platelet agonists such as ADP, thrombin and collagen. The defect is caused by mutations in one of the integrin genes, ITGA2 or ITGB3, encoding the αIIbβ3 receptor complex. Lack of expression or qualitative defects in αIIbβ3 results in a disturbed interaction between activated platelets and adhesive glycoproteins (fibrinogen at low shear and vWF at high shear) that bridge adjacent platelets during platelet aggregation (Fig. 1a). The Bernard-Soulier syndrome (MIM 231200) is caused by abnormalities in the GP Ib/IX/V receptor complex due to mutations in the genes for GPIbα, GPIbβ or GPIX (but there are no reports of BSS affecting the GPV gene) [22]. It is an autosomal recessive disorder with moderate to severe macrothrombocytopenia, decreased platelet survival and often a spontaneous bleeding tendency. The bleeding events can be severe but are usually controlled by platelet transfusion. Most heterozygotes, with a few exceptions, do not have a bleeding diathesis. BBS platelets aggregate normally in response to physiological agonists (ADP and collagen), have a weak response towards low concentrations of thrombin and do not agglutinate when platelet rich plasma is stirred with ristocetin or botrocetin [22].
Defects in G protein signaling resulting in an isolated platelet defect [36] are expected to be caused by a mutant G protein-coupled receptor (GPCR) since these can be cell-specific while the G proteins and their downstream effectors in this pathway are ubiquitously expressed. A dominantly inherited mutation (Arg60Leu) in the Gq-coupled TXA2 receptor was described in patients with a mild bleeding disorder characterized by defective platelet aggregation responses to TXA2 and its analogues (MIM 188070) [15]. In cultured cells, the Arg60Leu mutant was shown to impair phospholipase C (PLC) activation. Patients can be heterozygous (with some PLC activation left) or homozygous (without PLC activation) for this mutation but all have a life-long history of mucosal bleeding and easy bruising but no episodes of major bleeding such as hematuria, gastrointestinal bleeding or hemarthrosis [14]. The Gi-coupled ADP receptor P2Y12 (Fig. 1b) is responsible for the sustained, full aggregation response to ADP. P2Y12 deficiency (MIM 609821) is an autosomal recessive bleeding disorder characterized by excessive bleeding, prolonged bleeding time and abnormalities that are very similar to those observed in patients with secretion defects (reversible aggregation in response to weak agonists and impaired aggregation towards low concentrations of collagen and thrombin), except for the severely impaired response to ADP [2]. Study of the heterozygous P2Y12 defect revealed platelets that undergo a normal first wave of ADP induced aggregation but abnormal ATP secretion with different agonists [3, 42].
A defective platelet secretion is described for patients with absent alpha granules (gray platelet syndrome) or abnormal dense granules (delta-storage pool disease, δ-SPD) [37]. Gray platelet syndrome or α-SPD (MIM 139090) owes its name to the fact that the typically enlarged platelets devoid of α-granule staining, present with a gray color in a Wright-stained blood smear [32]. Most cases are sporadic though some family studies suggest an autosomal dominant inheritance. Affected members have a life-long history of mucocutaneous bleeding, which may vary from mild to moderate in severity, prolonged bleeding time, mild thrombocytopenia, abnormally large platelets and an isolated reduction of the platelet α-granule content. The molecular defect(s) in α-SPD have not yet been defined and further insights into the molecular mechanisms responsible for platelet exocytosis (as the SNARE proteins) will help in the search for causes of human platelets secretory disorders. δ-SPD (MIM 185050) may present as an isolated platelet function defect or can be associated with a variety of other congenital defects (see further). δ-SPD is characterized by a bleeding diathesis of variable degree, mildly to moderately prolonged skin bleeding time (fully related to the amount of ADP or serotonin contained in the granule), abnormal platelet secretion induced by several agonists and a reduced platelet aggregation. The δ-SPD platelets have decreased levels of the dense granule contents: ATP and ADP, serotonin, calcium and pyrophosphate (Fig. 1c). It was estimated that 10–18% of patients with a congenital abnormality of the platelet function have δ-SPD [12]. The inheritance pattern is autosomal recessive in some families while autosomal dominant in others but the molecular players responsible for δ-SPD are still unknown.
Human disorders comprising a platelet defect
Defects in platelet adhesion and subsequent platelet activation can also be due to an alteration in the platelet cytoskeletal organization, which consists of the microtubules and F-actin coupled to myosin, filamin and dystrophin [29]. Mutations in these widely expressed proteins result in a broader clinical phenotype. May-Hegglin anomaly (MIM 155100), Fechtner syndrome (MIM 153640), Epstein syndrome (MIM 153650), and Sebastian syndrome (MIM 605249) are characterized by macrothrombocytopenia, with or without different types of leukocyte inclusions, which can only be differentiated by an accurate ultrastructural examination [38]. In addition, patients with Epstein or Fechtner syndrome also suffer from nephritis, deafness, and congenital cataracts. Recently it became obvious that Sebastian platelet syndrome, May-Hegglin anomaly, Fechtner and Epstein syndrome have mutations in the same gene MYH9, encoding the 224-kD nonmuscle myosin heavy chain 9 polypeptide [26]. This gene is expressed in platelets, monocytes, granulocytes, the kidney, the auditory system but also in a lot of other tissues. MYH9 deficiency results in an alteration of the composition and agonist-induced reorganization of the platelet cytoskeleton [1, 6]. The cytoskeletal defect could also explain the abnormal platelet formation from megakaryocytes, resulting in thrombocytopenia and giant platelets in MYH9 deficiency. Why patients with May-Hegglin anomaly, Fechtner syndrome, Epstein syndrome and Sebastian platelet syndrome have different signs and symptoms in other tissues than their common defect in platelets still remains to be elucidated.
Duchenne muscular dystrophy (DMD) is an X-linked recessive disease (MIM 310200) characterized by progressive degeneration of muscle resulting in early death from respiratory or cardiac failure. DMD is caused by mutations in the dystrophin gene, a 427-kDa membrane-associated cytoskeletal protein. Evidence for a role of dystrophin in platelets started with the observation that DMD patients tend to bleed more during spinal surgery for scoliosis than do patients during the same surgery with other underlying conditions [7]. Other C-terminal isoforms of dystrophin due to differential promoter usage and/or alternative splicing at the 3′-end of the gene have been identified in platelets (Dp71), the retina (Dp260) and in the peripheral (Dp116) and central nervous systems (Dp140). It is well established that platelets contain a complex membrane cytoskeleton that resembles, at least in part, the cytoskeleton found in muscle, but a role for dystrophin during platelet activation still remains to be clarified [18, 27]. Recent studies showed a role for dystrophin in normal controls during platelet spreading and adhesion by regulating the α2β1 receptor but this was not studied in DMD patients [4, 5]. Another study describes a normal platelet function in DMD patients and blames the selective defect of primary hemostasis in DMD to impaired vessel reactivity [43].
Patients with an abnormal signal transduction are a heterogeneous group combined of defects in platelet G protein-coupled receptors (GPCR), the G proteins, and their effectors. Due to an extreme complex regulation between those key components (Fig. 1b), the incidence of this class of defects is definitely underestimated and the underlying molecular defects for the signaling problems are still largely unknown. Platelet Gs activity is easily determined using the platelet aggregation-inhibition test which gives a value for the Gs activity based on the inhibition of platelet aggregation by the rapid generation of cAMP after incubation with different Gs agonists such as prostacyclin or prostaglandin (Fig. 1b). A congenital Gs hyperfunction syndrome was described in three patients of two unrelated families due to a paternally inherited functional polymorphism in the extra-large stimulatory G-protein gene (XLαs) and its overlapping cofactor ALEX [8]. This XLαs variant is associated with Gs hyperfunction in platelets, leading to an increased trauma-related bleeding tendency but is also accompanied by neurological problems and brachydactyly (MIM 139320). A subsequent study revealed eight additional patients who paternally inherited the same XLαs polymorphism presenting with platelet Gs hyperfunction, brachydactyly, increased alkaline phosphatase and neurological problems or growth deficiency [9, 17].
Megakaryocytes and platelets express the Gs-coupled VPAC1 receptor, for which both the pituitary adenylyl cyclase-activating polypeptide (PACAP) and the vasointestinal peptide (VIP) are specific agonists. Studies in two related patients with a partial trisomy 18p revealed three copies of the PACAP gene and elevated PACAP concentrations in plasma. The patients suffer from multiple neurological (epilepsy, hypotonia, convulsions, mental retardation, tremor, psychotic, hyperactive behavior), gastro-intestinal (diarrhea, vomiting) and endocrinological (hypoplasia of the pituitary gland, hypogonadotropic hypogonadism) problems and have a pronounced bleeding tendency (MIM 102980) [10]. The basal cAMP level in the patients' platelets was strongly elevated, providing a basis for the strongly reduced platelet aggregation. The VPAC1 signalling pathway also mediates megakaryocyte maturation and platelet formation (unpublished results). Patients with PACAP overexpression have a mild thrombocytopenia, a normal platelet survival, relatively small platelets and their bone marrow examination reveals almost no mature megakaryocytes.
There exist two rare syndromic forms of the δ-SPD: the Hermansky-Pudlak syndrome (HPS) and the Chediak-Hygashi syndrome (CHS). HPS (MIM 203300) consists of several genetically different autosomal recessive disorders, which share the clinical manifestations of oculocutaneous albinism, bleeding, and lysosomal ceroid storage resulting from defects of multiple cytoplasmic organelles: melanosomes, platelet-dense granules, and lysosomes hypopigmentation [25, 36]. HPS can arise from mutations in at least eight different genes known to date (HSP1 to HSP8), all coding for proteins involved in the formation, trafficking or fusion of intracellular vesicles of the lysosomal lineage [44]. CHS (MIM 214500) is also an autosomal recessive disorder, characterized by variable degrees of oculocutaneous albinism, large peroxidase-positive cytoplasmic granules in hematopoietic and non-hematopoietic cells, δ-SPD, recurrent infections, neutropenia, and an accelerated chronic lymphohistiocytic infiltration phase. The only known CHS-causing gene, LYST, codes for a large protein of unknown function but it seems that CHS is a disease of vesicle trafficking [16]. Most CHS patients present in early childhood and die before the age of 7 years unless treated by bone marrow transplantation [21]. About 10–15% of patients exhibit a much milder clinical phenotype and survive to adulthood but develop progressive and often fatal neurological dysfunction.
Human disorders examined by functional and morphological platelet assays
Some mainly polygenetic disorders can also be studied by using platelets although the patients only present with a subtle subclinical platelet phenotype. It is not easy to define disorders such as diabetes type 2 or some neurological diseases according to a defective adhesion, G protein signalling and secretion since the platelet defect is not yet well studied and usually overlaps different pathways. This part of the review will only briefly focus on the use of platelet research in our understanding of neurological disorders. It has been known for years that certain cellular functions are very similar in platelets and in neurosecretory cells [33] but the link between functional platelet studies and neurological defects is novel. A platelet and a neuron contain both mitochondria and dense core vesicles in which transmitters (such as serotonin) are stored. Platelets release serotonin upon activation (Fig. 2c) and the neuronal membrane is facilitated by a calcium dependent excitation-excretion coupling mechanism. In addition, both platelets and neurons contain functional neurotransmitter and neuromodulator receptor sites on their outer membrane such as adrenoceptors, serotonin receptors and serotonin transporters. More recently it was shown that platelets also express GABA and glutamate receptors [24, 34]. Serotonin uptake and release by platelets and serotonin plasma levels have been quantified and found to be altered in patients with bipolar disorder, schizophrenia, depression, aggression, autism, migraine, etc. [19, 23, 28]. Many epidemiological studies try to link these changes in activity of the serotonin transporter or changes in the density and responsiveness of the serotonin 2A receptor and the alpha2 adrenoceptor on the platelet membrane of these patients with genetic polymorphism in the corresponding genes. As for other epidemiological studies solid evidence for any linkage is not obvious but it is irrefutable that platelet studies have been invaluable in enabling an insight to be gained into the role of serotonin in a number of psychiatric and neurological disease [39, 41]. Major advances are expected of platelet research for this field in the near future since it is now obvious that besides the serotonin pathway, which was only the tip of the iceberg, many other pathways are chaired between platelets and neurons as well as the many gene products responsible for the regulation of granule formation, transport, secretion and endocytosis.
Conclusions
Platelet research is an expanding field originally studying isolated thrombopathies caused by the imbalance between thrombosis and hemostasis but more recently being able to bring novel insights in our understanding of human pathology in other clinical disciplines such as neurology, endocrinology and metabolic diseases. Platelets are easily accessible cells, and different techniques are possible to study platelet function and morphology under basal and activated conditions. Defects in platelet adhesion, G protein signalling and secretion can result from mutations in platelet-specific genes leading to isolated thrombopathies or from mutations in widely expressed genes leading to broader clinical phenotype including a platelet defect. In addition to using platelet research for diagnostic purposes, these platelet functional and morphological studies can also be used for research aims. From the close collaboration between clinicians of different disciplines, geneticists and the functional platelet research unit, novel insights in the pathogenesis of different human disorders are to be expected in the near future by using this strategy.
Abbreviations
- ADP:
-
Adenosine diphosphate
- BSS:
-
Bernard-Soulier syndrome
- GABA:
-
Gamma-aminobutyric acid
- GP:
-
Glycoprotein
- MYH9:
-
Myosin heavy chain 9
- PACAP:
-
Pituitary adenylyl cyclase-activating polypeptide
- PFA100:
-
Platelet function analyzer 100
- PLC:
-
Phospholipase C
- SNARE:
-
Soluble NSF attachment receptor
- SPD:
-
Storage pool disease
- TXA2:
-
Thromboxane A2
- VIP:
-
Vasointestinal peptide
- vWF:
-
Von Willebrand factor
References
Canobbio I, Noris P, Pecci A, Balduini A, Balduini CL, Torti M (2005) Altered cytoskeleton organization in platelets from patients with MYH9-related disease. J Thromb Haemost 3:1026–1035
Cattaneo M, Zighetti ML, Lombardi R, Martinez C, Lecchi A, Conley PB, Ware J, Ruggeri ZM (2003) Molecular bases of defective signal transduction in the platelet P2Y12 receptor of a patient with congenital bleeding. Proc Natl Acad Sci USA 100:1978–1983
Cattaneo M (2005) The P2 receptors and congenital platelet function defects. Semin Thromb Hemost 31:168–173
Cerecedo D, Martinez-Rojas D, Chavez O, Martinez-Perez F, Garcia-Sierra F, Rendon A, Mornet D, Mondragon R (2005) Platelet adhesion: structural and functional diversity of short dystrophin and utrophins in the formation of dystrophin-associated-protein complexes related to actin dynamics. Thromb Haemost 94:1203–1212
Cerecedo D, Mondragon R, Cisneros B, Martinez-Perez F, Martinez-Rojas D, Rendon A (2006) Role of dystrophins and utrophins in platelet adhesion process. Br J Haematol 134:83–89
Di Pumpo M, Noris P, Pecci A, Savoia A, Seri M, Ceresa IF, Balduini CL (2002) Defective expression of GPIb/IX/V complex in platelets from patients with May-Hegglin anomaly and Sebastian syndrome. Haematologica 87:943–947
Forst J, Forst R, Leithe H, Maurin N (1998) Platelet function deficiency in Duchenne muscular dystrophy. Neuromuscul Disord 8:46–49
Freson K, Hoylaerts MF, Jaeken J, Eyssen M, Arnout J, Vermylen J, Van Geet C (2001) Genetic variation of the extra-large stimulatory G protein alpha-subunit leads to Gs hyperfunction in platelets and is a risk factor for bleeding. Thromb Haemost 86:733–738
Freson K, Jaeken J, Van Helvoirt M, de Zegher F, Wittevrongel C, Thys C, Hoylaerts MF, Vermylen J, Van Geet C (2003) Functional polymorphisms in the paternally expressed XLalphas and its cofactor ALEX decrease their mutual interaction and enhance receptor-mediated cAMP formation. Hum Mol Genet 12:1121–1130
Freson K, Hashimoto H, Thys C, Wittevrongel C, Danloy S, Morita Y, Shintani N, Tomiyama Y, Vermylen J, Hoylaerts MF, Baba A, Van Geet C (2004) The pituitary adenylate cyclase-activating polypeptide is a physiological inhibitor of platelet activation. J Clin Invest 113:905–912
Gibbins JM (2004) Platelet adhesion signalling and the regulation of thrombus formation. J Cell Sci 117:3415–3425
Gunay-Aygun M, Huizing M, Gahl WA (2004) Molecular defects that affect platelet dense granules. Semin Thromb Hemost 30:537–547
Hayward CP, Rao AK, Cattaneo M (2006) Congenital platelet disorders: overview of their mechanisms, diagnostic evaluation and treatment. Haemophilia 12:128–136
Higuchi W, Fuse I, Hattori A, Aizawa Y (1999) Mutations of the platelet thromboxane A2 (TXA2) receptor in patients characterized by the absence of TXA2-induced platelet aggregation despite normal TXA2 binding activity. Thromb Haemost 82:1528–1531
Hirata T, Kakizuka A, Ushikubi F, Fuse I, Okuma M, Narumiya S (1994) Arg60 to Leu mutation of the human thromboxane A2 receptor in a dominantly inherited bleeding disorder. J Clin Invest 94:1662–1667
Introne W, Boissy RE, Gahl WA (1999) Clinical, molecular, and cell biological aspects of Chediak-Higashi syndrome. Mol Genet Metab 68:283–303
Jaeken JC, Freson K, Goemans NM, De Cock PP, De Vos RM, Lagae LG, Van Geet CA (2003) G protein diseases: newly recognized causes of metabolic encephalopathy. Eur J Paediatr Neurol 7:211–215
Jagadeesh G, Lavanya M, Anandaraj MP, Anjaneyulu A (1990) Altered protein kinase C and protein kinase A activities in erythrocyte membrane, platelets and lymphocytes of Duchenne muscular dystrophy (DMD) patients. Clin Chim Acta 193:79–84
Janusonis S, Anderson GM, Shifrovich I, Rakic P (2006) Ontogeny of brain and blood serotonin levels in 5-HT receptor knockout mice: potential relevance to the neurobiology of autism. J Neurochem 99:1019–1031
Jurk K, Kehrel BE (2005) Platelets: physiology and biochemistry. Semin Thromb Hemost 31:381–392
Karim MA, Suzuki K, Fukai K, Oh J, Nagle DL, Moore KJ, Barbosa E, Falik-Borenstein T, Filipovich A, Ishida Y, Kivrikko S, Klein C, Kreuz F, Levin A, Miyajima H, Regueiro J, Russo C, Uyama E, Vierimaa O, Spritz RA (2002) Apparent genotype-phenotype correlation in childhood, adolescent, and adult Chediak-Higashi syndrome. Am J Med Genet 108:16–22
Kunishima S, Kamiya T, Saito H (2002) Genetic abnormalities of Bernard-Soulier syndrome. Int J Hematol 76:319–327
Lance JW (1991) 5-hydroxytryptamine and its role in migraine. Eur Neurol 31:279–281
Langer SZ, Galzin AM (1988) Studies on the serotonin transporter in platelets. Experientia 44:127–130
Li W, Rusiniak ME, Chintala S, Gautam R, Novak EK, Swank RT (2004) Murine Hermansky-Pudlak syndrome genes: regulators of lysosome-related organelles. Bioessays 26:616–628
Martignetti J (2002) Five (un)easy pieces: the MYH9-related giant platelet syndromes. Haematologica 87:897–898
Maurin N, Forst J, Leithe H, Forst R (1998) Deficiency of platelet glass bead adhesion and platelet membrane glycoprotein IV (CD36) in Duchenne muscular dystrophy. Thromb Haemost 79:1067
Mendelson SD (2000) The current status of the platelet 5-HT(2A) receptor in depression. J Affect Disord 57:13–24
Nakamura F, Pudas R, Heikkinen O, Permi P, Kilpelainen I, Munday AD, Hartwig JH, Stossel TP, Ylanne J (2006) The structure of the GPIb-filamin A complex. Blood 107:1925–1932
Nurden AT (2005) Qualitative disorders of platelets and megakaryocytes. J Thromb Haemost 3:1773–1782
Nurden AT (2006) Glanzmann thrombasthenia. Orphanet J Rare Dis 1:10
Nurden AT, Nurden P (2007) The gray platelet syndrome: clinical spectrum of the disease. Blood Rev 21:21–36
Pletscher A (1986) Blood platelets as neuronal models: use and limitations. Clin Neuropharmacol 9:344–346
Rainesalo S, Keranen T, Saransaari P, Honkaniemi J (2005) GABA and glutamate transporters are expressed in human platelets. Brain Res Mol Brain Res 141:161–165
Ramasamy I (2004) Inherited bleeding disorders: disorders of platelet adhesion and aggregation. Crit Rev Oncol Hematol 49:1–35
Rao AK, Jalagadugula G, Sun L (2004) Inherited defects in platelet signaling mechanisms. Semin Thromb Hemost 30:525–535
Reed GL (2004) Platelet secretory mechanisms. Semin Thromb Hemost 30:441–450
Seri M, Cusano R, Gangarossa S, Caridi G, Bordo D, Lo Nigro C, Ghiggeri GM, Ravazzolo R, Savino M, Del Vecchio M, d’Apolito M, Iolascon A, Zelante LL, Savoia A, Balduini CL, Noris P, Magrini U, Belletti S, Heath KE, Babcock M, Glucksman MJ, Aliprandis E, Bizzaro N, Desnick RJ, Martignetti JA (2000) Mutations in MYH9 result in the May-Hegglin anomaly, and Fechtner and Sebastian syndromes. The May-Heggllin/Fechtner syndrome consortium. Nat Genet 26:103–105
Serretti A, Calati R, Mandelli L, De Ronchi D (2006) Serotonin transporter gene variants and behavior: a comprehensive review. Curr Drug Targets 7:1659–1669
Sham RL, Francis CW (1994) Evaluation of mild bleeding disorders and easy bruising. Blood Rev 8:98–104
Sutcliffe JS, Delahanty RJ, Prasad HC, McCauley JL, Han Q, Jiang L, Li C, Folstein SE, Blakely RD (2005) Allelic heterogeneity at the serotonin transporter locus (SLC6A4) confers susceptibility to autism and rigid-compulsive behaviors. Am J Hum Genet 77:265–279
Storey RF (2006) Biology and pharmacology of the platelet P2Y12 receptor. Curr Pharm Des 12:1255–1259
Turturro F, Rocca B, Gumina S, De Cristofaro R, Mangiola F, Maggiano N, Evangelista A, Salsano V, Montanaro A (2005) Impaired primary hemostasis with normal platelet function in Duchenne muscular dystrophy during highly-invasive spinal surgery. Neuromuscul Disord 15:532–540
Wei ML (2006) Hermansky-Pudlak syndrome: a disease of protein trafficking and organelle function. Pigment Cell Res 19:19–42
Acknowledgements
This work was supported by the ‘Excellentie financiering KULeuven’ (EF/05/013), by research grants G.0453.05 and G.0124.02 from the FWO-Vlaanderen (Belgium) and by GOA/2004/09 from the Research Council of the University of Leuven (Onderzoeksraad K.U.Leuven, Belgium).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Freson, K., Labarque, V., Thys, C. et al. What’s new in using platelet research? To unravel thrombopathies and other human disorders. Eur J Pediatr 166, 1203–1210 (2007). https://doi.org/10.1007/s00431-007-0543-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00431-007-0543-7