
Abstract
Oligodendrocytes are the myelinating cells of the central nervous system (CNS). They are the end product of a cell lineage which has to undergo a complex and precisely timed program of proliferation, migration, differentiation, and myelination to finally produce the insulating sheath of axons. Due to this complex differentiation program, and due to their unique metabolism/physiology, oligodendrocytes count among the most vulnerable cells of the CNS. In this review, we first describe the different steps eventually culminating in the formation of mature oligodendrocytes and myelin sheaths, as they were revealed by studies in rodents. We will then show differences and similarities of human oligodendrocyte development. Finally, we will lay out the different pathways leading to oligodendrocyte and myelin loss in human CNS diseases, and we will reveal the different principles leading to the restoration of myelin sheaths or to a failure to do so.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
General introduction
Most of our knowledge about the biology of oligodendrocytes and myelin derives from studies in rodents. This fact can be easily explained, since cells or tissue slices of these animals are readily available and can be easily cultured, and since a large number of genetically modified animals allows it to dissect the function of individual proteins in the proper spatial and temporal context of development and disease. Studies in rodents led to the identification of important principles of oligodendrocyte development and myelin formation.
The development of oligodendrocytes and myelin—a rodent's perspective
Spinal cord and brain contain different subclasses of oligodendrocytes which derive from multiple sources
In the spinal cord, most oligodendrocytes derive from a specialized domain of the ventral ventricular zone, which first gives rise to motor neuron precursors, and then, after the neurogenic/gliogenic switch, to oligodendrocyte precursor cells/progenitors (OPCs) [107, 181, 183, 214]. From there, OPCs migrate all through the spinal cord and finally differentiate into myelin-forming oligodendrocytes. Later, an additional source of OPCs arises in the dorsal spinal cord, contributing to 10–15% of the final oligodendrocyte population in the spinal cord [25, 49, 195].
In the forebrain, the first OPCs originate in the medial ganglionic eminence and anterior entopeduncular area of the ventral forebrain. These OPCs populate the entire embryonic telencephalon including the cerebral cortex, and are then joined by a second wave of OPCs derived from the lateral and/or caudal ganglionic eminences. The third wave of OPCs, finally, arises within the postnatal cortex [86]. These different populations of OPCs are functionally redundant: When any one of them is destroyed at source by the targeted expression of a toxin gene in mice, the remaining cells spread into the vacant territory and restore the normal distribution of OPCs. As a result of this, a normal complement of oligodendrocytes and myelin can be produced, and the mice develop, survive, and behave normally [86]. In spite, or maybe even due to this functional redundancy, the different OPC populations seem to be fierce competitors.
Different OPC lines compete with each other during development
In the developing rodent spinal cord, the competition is won by OPCs derived from the ventral ventricular zone: These cells represent the first wave of cells of the oligodendrocyte lineage generated in the spinal cord, and eventually give rise to 85–90% of the final oligodendrocyte population found in this organ [25, 49, 195]. In this case, competition for limiting quantities of growth factors like platelet-derived growth factor (PDGF) might be a determining factor for winning the competition [9, 26, 196].
In the developing forebrain, however, the situation seems to be quite different. Here, the competition is clearly lost by the first wave OPCs: although these cells are the first to distribute and to occupy vacant territories, their contribution to the total OPC population rapidly declines, until they are almost completely eliminated from the adult forebrain.
The reasons for the eventual loss of one OPC population in the developing forebrain remain completely unclear, and one can only speculate about possible causes and consequences [157].
According to such speculations, the first wave of forebrain OPCs emerging from the medial ganglionic eminence and the anterior entopeduncular area might represent the most “primitive source” of oligodendrocytes, a relic that lost its importance during evolution, when new sources of OPCs developed in the expanding brain. This assumption is supported by the observation that birds, which have much less cortical volume than mammals derive their OPCs only from the anterior entopeduncular area [157]. Alternatively, the first wave OPCs could be lost in the course of a (not yet demonstrated) oligodendrocyte turnover, or as a result of changes in the availability/responsiveness to growth and differentiation factors. Moreover, they might become dispensable if they have functions during development, that are not required in the adult [157].
No matter whether OPCs belong to the first, second, or third wave of cells, they have one point in common: They have to travel long distances in order to end up in their final place of destination. This migration is tightly controlled.
OPC migration is guided by regulatory signals
To date, three different classes of secreted molecules seem to be involved in the migration of OPCs: growth factors like PDGF, FGF [155, 174] or hepatocyte growth factor [211, 138]; chemotropic molecules like netrins and secreted semaphorins [74, 193]; and the chemokine CXCL1 [37]. Although there is no doubt that these factors play a role in OPC migration, the exact mode of action of these factors is still a matter of controversy, in part due to differences in experimental models, culture systems, and time points studied.
OPC migration is not only controlled by secreted molecules, but is also regulated by contact-mediated mechanisms involving many different extracellular matrix proteins and cell surface molecules [52, 88, 199], N-cadherins [125, 133, 145, 167, 190], and possibly even additional, yet unidentified molecules.
From all these different molecules, a common theme evolves, demonstrating contact-based migration of OPCs over extracellular matrices, axonal tracts, and astrocytic surfaces [37]. Once located at their final destination, some OPCs persist into adulthood (see Appendix 1), while the vast majority differentiates to myelin-producing oligodendrocytes.
The differentiation of OPCs to oligodendrocytes and the onset of myelination are spatially and temporally regulated
This process involves signaling processes between the Notch1 receptor, its ligand Jagged 1 located on the axonal surface [55], and γ-secretase [150, 202]. Interestingly, oligodendrocytes have only a brief period of time for myelination early during differentiation, and are relatively incapable of myelinating once they are mature [202]. Moreover, the ensheathment of multiple axons by a single oligodendrocyte is a highly coordinated event: oligodendrocytes do not ensheath different axons sequentially at different time points, but are done within a brief window of time, typically within 12–18 h [8, 202].
Oligodendrocytes do not randomly wrap plasma membrane around neuronal processes
Oligodendrocytes select axons with diameters above 0.2 μm [171]. The molecular cues for this recognition remain unknown. In the peripheral nervous system, the critical axonal signal for myelination by Schwann cells is provided by neuronal neuregulin-1 (NRG1) type III, interacting with glial ErbB receptors, and it was thought for a long time that NRG1/ErbB signaling might also regulate myelination in the CNS. This point of view was supported by the observation that not only Schwann cells, but also oligodendrocytes express erbB receptors (oligodendrocytes erbB2 and erbB4, Schwann cells erbB2 and erbB3, but only little erbB4 [197]). It was further observed, that oligodendrocytes fail to develop in spinal cord explants derived from mice lacking neuregulin [198], that erbB2 is required for the development of terminally differentiated spinal cord oligodendrocytes [141], and that erbB2 plays a role in governing the properly timed exit from the cell cycle during development into myelinating oligodendrocytes [89]. Moreover, mice with dominant negative erbB4 have more differentiated oligodendrocytes, but each of them seems to myelinate less axonal surface and to produce thinner myelin sheaths than its wild-type counterpart [159]. And last, mice haploinsufficient for type III NRG1 are hypomyelinated in the brain, and normally myelinated in the optic nerve and spinal cord [187]. However, recent observations challenge the assumption that NRG1/ErbB signaling might also regulate myelination in the CNS. Instead, they suggest that NRG1/ErbB signaling serves distinct functions in myelination of the peripheral and central nervous system [19]: Mice completely lacking NRG1 beginning at different stages of neural development assemble normal amounts of myelin, on schedule [19]. When neuregulin signaling is completely abolished in oligodendrocytes of double mutants lacking ErbB3 and ErbB4 (and carrying ErbB2 without ligand-binding activity), CNS axons are nevertheless myelinated without delay, and at the same level as in control mice, at least until postnatal day p11, the latest time point studied [19]. However, while myelination was not delayed even in the absence of NRG1, there was clear evidence for premature, and even for hyper-myelination in NRG1 type III overexpressing mice [19]. Cumulatively, these data suggest that normal myelination occurs independently of NRG1 signaling in vivo. They also demonstrate that excess NRG1 can initiate the myelination program in CNS development, but that this function is normally provided by different, still unknown axonal signaling systems.
Myelination is a regulated process
The data summarized above led to the conclusion that the onset of CNS myelination in normal development might be determined by the degree of neuronal differentiation, and not by the timing of an intrinsic oligodendrocyte differentiation program [19].
One essential signal for the onset of myelination seems to be provided by the electrical activity of neurons. For example, the optic nerves of mice which had been reared in the dark developed fewer myelinated axons than control optic nerves [62], optic nerves of naturally blind cape-mole rats are hypomyelinated [139], and blockade of sodium-dependent action potentials in developing optic nerves inhibits myelination [38]. Vice versa, increasing neuronal firing with α-scorpion toxin enhances myelination [38], and premature eye opening accelerates myelination in rabbit optic nerves [186]. Action potential firing leads to the release of ATP [177] and adenosine [96, 115], which can mediate neuron-glial communications. In the CNS, adenosine inhibits the proliferation of OPCs, stimulates their differentiation, and promotes the formation of myelin [178]. The ATP released from axons firing axon potentials does not directly act on OPCs or oligodendrocytes. Instead, it triggers the release of leukemia inhibitory factor (LIF) from astrocytes, which promotes myelination by mature oligodendrocytes [67]. This fact could explain why LIF−/− mice display impaired myelin formation [23], and why interfering with astrocyte biology leads to myelin abnormalities, as seen in mice with knock-out of the astrocytic protein GFAP [104], and in patients with Alexander disease, a fatal white matter disorder of childhood caused by mutations in the GFAP gene [124].
Although the electrical activity of neurons in the CNS is an essential promyelinating factor, additional changes on axons seem to be needed to drive efficient myelin formation. Some of these changes seem to be induced by electrical activity as well. For example, during development, all growing nerve fibers express the polysialylated-neural cell adhesion molecule (PSA-NCAM), which prevents homophilic NCAM–NCAM adhesion and, more generally, cell–cell interactions. Only when this form of NCAM is downregulated, as it is in electrically active neurons [91, 97], myelination can proceed [32]. Another molecule correlating with the extent of oligodendrocyte maturation and myelination is LINGO-1, a transmembrane protein with leucine-rich repeats and an immunoglobulin domain expressed in neurons and oligodendrocytes. Loss of LINGO-1 function in oligodendrocytes leads to increased myelination, whereas its overexpression inhibits myelin formation [122].
There are certainly more, yet unidentified molecular mechanisms enabling oligodendrocytes to recognize, ensheath, and wrap axons [8].
Myelin assembly is under neuronal control
The window of time available for the onset of myelination seems to be very narrow, in the range of 12–18 h during which oligodendrocytes have to wrap layer after layer of plasma membrane around multiple axons [8]. Hence, they have to synthesize, sort, and traffick an enormous amount of proteins in short time. The machinery for these processes is rather complex. For example, one major myelin protein, myelin basic protein (MBP, Table 1), is targeted by transport of its mRNA. The MBP mRNA is assembled into granules in the perikaryon of oligodendrocytes, transported along processes, and then localized to the myelin membrane [2, 3]. The other major protein, proteolipid protein (PLP)/DM20 is transported to myelin by vesicular transport through the biosynthetic pathway. Two different observations suggest that both delivery systems might be under the control of neuronal signals.
First, oligodendrocyte cultures without neurons express MBP, PLP, and galactosylceramide (GalC). However, under these conditions, PLP is predominantly intracellularly localized, and shows only little co-localization with MBP or GalC in the membrane sheets. In coculture with neurons, however, PLP, MBP, and GalC are co-localized. This rearrangement of the oligodendroglial plasma membrane is critically dependent on the presence of MBP. MBP seems to act as a lipid coupler by bringing the different layers of myelin in close position (a process termed “vertical membrane coupling”), and by clustering the lipid bilayer in lateral dimensions (“horizontal membrane coupling”). This neuron- and MBP-dependent clustering mechanism might be responsible for the concentration of membrane components within myelin [48].
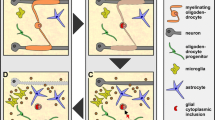
Second, in the absence of neurons, PLP is produced by oligodendrocytes, but immediately internalized by endocytosis. After receiving neuronal signals, the rate of endocytosis is reduced, and PLP is transported from the late endosomes/lysosomes to the plasma membrane by exocytosis [192]. This regulation of PLP trafficking might represent a mechanism to store membrane produced in advance, before the onset of myelination, and to release this membrane on demand in a regulated fashion [171] (Fig. 1).

a–d Detection of oligodendrocytes in paraffin embedded tissue sections. a Mouse cortex, stained by immunocytochemistry for CNPase shows numerous process bearing oligodendrocytes and staining of myelin sheaths, ×120; b high magnification of a CNPase positive oligodendrocyte in the mouse cortex (layer I), showing a small round cell body and few cell processes connected to myelin sheaths, ×1,000; c rat spinal cord stained by in situ hybridization for myelin basic protein mRNA; staining is seen in the cell bodies (e.g., in the gray matter) as well as in oligodendrocyte processes associated with myelin sheaths (dark staining in the white matter), ×50; d rat spinal cord stained by in situ hybridization for proteolipid protein (PLP); the mRNA for PLP is only located within the perinuclear cytoplasm, there is no staining of myelin, ×50; e–h oligodendrocyte pathology in transgenic animals overexpressing proteolipid protein; e and f hemizygous animal, which shows normal myelination, stained by immunocytochemistry for PLP; in the normal animal PLP protein is rarely detected in the cytoplasm of oligodendrocytes; in hemizygous PLP transgenic animals a variable extent of PLP expression is seen in the cytoplasm of oligodendrocytes (e), and this is associated with aberrant formation of myelin-like structures within and adjacent to the cells (f); g–h homozygous animal, which shows extensive dys-myelination; only few oligodendrocytes are preserved, which are connected to myelin sheaths and contain abundant PLP immunoreactivity in their cytoplasm (g); some of the PLP reactive oligodendrocytes show nuclear condensation and fragmentation, consistent with apoptosis (h), ×1,200; i–o neuropathology of progressive multifocal leukoencephalopathy; i multiple small confluent demyelinating lesions in the white and gray matter, giving the impression of a moth eaten pattern of demyelination (×2); k edge of an active demyelinating lesion with numerous macrophages, containing recent (luxol fast blue positive) degradation products; l and m pathologically altered nuclei in PML showing giant nuclei of astrocytes (l) and small oligodendrocyte nuclei with intranuclear inclusion (m); n and o similar nuclei, as shown in l and m contain virus antigen as revealed by immunocytochemistry; ×1,200
The development of oligodendrocytes and myelin—what is similar/different in humans?
The key findings of rodent oligodendrocyte development and myelin formation described above include: (1) a common progenitor cell for neurons and oligodendrocytes; (2) a ventral to dorsal progression of oligodendrogenesis; (3) multiple origins of oligodendrocytes; (4) the dependency of differentiation and migration on regulatory factors; and (5) the interrelationship between axonal signaling and myelination.
In general, all these principles seem to apply to the human system as well, although evidence for it is mostly circumstantial and beyond the scope of this review to discuss. Instead, we refer the reader to an excellent review on this matter [72], and select for further demonstration just one particularly important aspect: data indicating that also human oligodendrocytes have multiple origins. These data derive from studies of the human fetal forebrain at midgestation, and reveal the simultaneous presence of three different OPC populations [70, 71, 152]. The first population consists of cortical OPCs which express Dlx2 and Nkx2.1 [152], typical transcription factors of ventrally derived OPCs in rodents [72]. The second population consists of OPCs which do not express Dlx2 and Nkx2.1 and most likely represent dorsally derived oligodendrocytes in the human brain [152]. The third population, finally, expresses typical OPC markers like PDGFRα, NG2, and Olig1, and is found in a stream of cells migrating between the ganglionic eminences and cortical subventricular zone [152]. As in rodents, it remains unclear whether oligodendrocytes from these different sources have different roles, myelinate different axonal pathways, or affect the outcome of CNS pathologies.
Since our knowledge about human oligodendrocyte biology is limited, results from rodent studies are often extrapolated to the human situation. However, several essential differences between the rodent and the human brain strongly argue against such a simple and uncritical approach:
-
1.
Key regions of the human brain might be underdeveloped in rodents and vice versa. For example, humans have neocortical regions which are completely lacking in mice, while prominent structures of the rodent brain, the olfactory bulbs, are underdeveloped in humans [72].
-
2.
The time scale for myelination is different between humans and rodents. Due to the greater complexity of the human brain, myelination in the human forebrain takes decades, compared to weeks in rodents [126].
-
3.
The sheer numbers of oligodendrocytes in humans are dramatically increased (although the density of oligodendrocytes per mm3 is remarkably similar between rodents and humans) [140, 176].
-
4.
Human and rodent OPCs might respond differently to certain factors [6, 168, 205, 213]. This can be shown best in the case of CXCL1. Approximately 85% of all rodent OPCs carry receptors for CXCL1 [37], while only very few human OPCs do so [47]. Hence, CXCL1 can act directly on rodent OPCs, but has an indirect mode of action in humans, where it induces astrocytes to secrete OPC mitogens [47].
Consequences of myelination
Oligodendrocytes not only ensheath axons to electrically insulate these structures, but also induce a clustering of sodium channels along the axon, at the node of Ranvier, which is one important prerequisite for saltatory nerve conduction [78, 79]. Even normal axonal transport processes and neuronal viability seem to depend on proper myelination, since axons with modified myelin sheaths have altered axonal transport rates and changes in the microtubule number or stability [45, 90], or are swollen and show signs of degeneration [44, 53, 60, 82, 83, 98, 194]. The presence of intact myelin sheaths could even lead to an increase in axon diameter [35, 116, 161], possibly mediated by the local accumulation and phosphorylation of neurofilament [161, 162]. And last, oligodendrocytes can provide trophic support for neurons by the production of glial cell line-derived neurotrophic factor (GDNF) [203], brain-derived neurotrophic factor (BDNF) [41], or insulin-like growth factor-1 (IGF-1) [42].
Oligodendrocyte metabolism as risk factor for oligodendrocyte pathology
It has been estimated that during the peak of myelination, oligodendrocytes elaborate about three times its weight in membrane per day, and eventually support membrane up to 100× the weight of its cell body [36, 111, 120, 121]. This particular feature renders oligodendrocytes vulnerable at several different “Achilles′ heels”.
First, in order to myelinate properly, oligodendrocytes must have extremely high metabolic rates, and must consume large amounts of oxygen and ATP [121]. The production of ATP leads to the formation of hydrogen peroxide as a toxic byproduct, and a high cellular metabolism also creates reactive oxygen species, both of which must be properly metabolized [121]. Second, myelination is under control of many myelin synthetic enzymes, which require iron as a co-factor [36]. This may contribute to the observation that OPCs and oligodendrocytes have the largest intracellular stores of iron in the brain [34, 189], which can, under unfavorable conditions, evoke free radical formation and lipid peroxidation [18, 76]. On top of this, oligodendrocytes have only low concentrations of the anti-oxidative enzyme glutathione [189]. And last, during myelination, the capacity of the endoplasmic reticulum to produce and fold proteins properly seems to be a cellular “bottle neck”, since even slight variations in the amount of a single protein can mess up the entire system and result in the retention, misfolding, and accumulation of many other proteins in this oligodendrocytic organelle [10, 16].
Taken together, just being an oligodendrocyte seems already enough to put these cells at greater risk of damage under pathological conditions.
Mechanisms of oligodendrocyte death
Due to the combination of a high metabolic rate with its toxic byproducts, high intracellular iron, and low concentrations of the antioxidative glutathione, oligodendrocytes are particularly vulnerable to oxidative damage [76, 189]. Hence, oxidative damage is a common contributor to oligodendrocyte loss under many pathological conditions like MS and ischemia. It can act in concert with the sphingomyelinase/ceramide pathway: ceramide is the core component of sphingolipids, the major lipid components of myelin sheaths [11]. It is released by the action of sphingomyelinase which is normally inactive, but gets activated in response to oxidative stress [73, 172], inflammatory mediators [20, 172, 188], injury or infection [165]. Once released within oligodendrocytes, ceramide can activate pro-apoptotic signaling cascades eventually culminating in oligdendrocyte loss [20, 121].
Oligodendrocytes also express an arsenal of molecules rendering them susceptible to excitotoxic cell death [40, 103, 117, 119, 163]: they carry AMPA [184], kainate [4, 163], and NMDA [80, 123, 160] receptors which make them vulnerable to glutamate toxicity, and the ATP receptor P2X7 [118] which predisposes them to the damaging action of sustained levels of extracellular ATP [118].
Oligodendrocyte loss can also occur as a result of exposure to inflammatory cytokines. For example, tumor necrosis factor α (TNFα) can induce apoptosis of oligodendrocytes by binding to their p55 TNF receptor [75]. The situation is more complex in the case of interferon gamma (IFNγ). This cytokine is highly toxic for actively proliferating OPCs, much less so for immature oligodendrocytes, and not at all for mature oligodendrocytes [65]. Besides these direct actions, inflammatory mediators may also damage oligodendrocytes indirectly through stimulation of radical production in microglia and possibly also in astrocytes. Oxygen- and nitric oxide-radicals are particularly toxic for mitochondria through interaction and blockade of various proteins of the respiratory chain [113, 173]. Indeed, recent studies on changes of gene expression in glia cells revealed that many different pro-inflammatory cytokines can induce mitochondrial injury [106].
As mentioned above, oligodendrocytes are particularly vulnerable to oxidative damage and mitochondrial injury. This is probably reflected by the profound oligodendrocyte damage in certain toxic states, which interfere with mitochondrial function. Examples for this are the selective oligodendrocyte apoptosis and demyelination induced by cuprizone, a copper chelator interfering with complex IV of the mitochondrial respiratory chain [191], and by the intoxication with cyanides also blocking the respiratory chain at the level of complex IV [27, 63].
All these mechanisms described above do not destroy oligodendrocytes specifically, but may also impair function and viability of other cells, such as neurons and astrocytes. However, oligodendrocytes and their myelin sheaths are in general more susceptible to damage than other cellular components of the nervous system. This explains the so-called “bystander damage” of myelin and oligodendrocytes observed in many inflammatory disease states, in which the immune reaction is not specifically directed against these cells [206]. In fact, demyelination and oligodendrocyte death is a common feature of inflammatory white matter lesions, both in humans and experimental models. A particularly illustrative example is Devic’s neuromyelitis optica (NMO). This disease has originally been classified as an inflammatory demyelinating disease due to the presence of widespread primary demyelination in the spinal cord and optic nerves [110]. Recent immunological studies, however, provide clear evidence that the primary targets of the pathogenic immune (autoantibody) response in NMO are not oligodendrocytes, but astrocytes [100, 101]. Time course studies on lesion development in NMO patients revealed that astrocytes are destroyed first, but that this is followed by profound demyelination and oligodendrocyte, axons and nerve cells destruction [127, 128, 158]. Whether oligodendrocyte injury in this disease is only a bystander reaction of the inflammatory process or whether specific disturbance of the homeostatic interaction between astrocytes and oligodendrocytes plays an additional role, is currently unresolved.
Besides by non-specific bystander mechanisms, oligodendrocytes can also be destroyed by specific, cell selective immune mechanisms. Autoantibodies directed against an epitope on the extracellular surface of myelin or oligodendrocytes can induce demyelination either through activation of complement or through their recognition by Fc-receptors of activated macrophages. The most compelling examples for such autoantibodies are those directed against myelin oligodendrocyte glycoprotein (MOG, [105]) and galactocerebroside [43]. Antibody-mediated demyelination is an important mechanism in models of autoimmune encephalomyelitis and seems to play a role also in a subset of patients with MS-like inflammatory demyelinating diseases [109, 137]. Similarly, cytotoxic T-lymphocytes, directed against a myelin or oligodendrocyte antigen, or against a foreign (e.g., virus) antigen expressed in oligodendrocytes, can induce oligodendrocyte apoptosis, followed by selective demyelination [66, 130, 164]. As other glia cells and neurons, oligodendrocytes are able to express major histocompatibility (MHC) class I antigens under inflammatory conditions, which is an essential pre-requisition for antigen recognition and cytotoxicity by MHC class I restricted cytotoxic T-cells [64].
Different pathological patterns of white matter injury reflect different pathogenetic mechanisms of myelin and oligodendrocyte damage
Although pathogenetic events that target myelin and oligodendrocytes invariably result in primary demyelination, the structural patterns of tissue injury in the initial stages of lesion formation differ, depending upon the mechanism involved. Three main patterns of tissue injury can be differentiated.
Simultaneous destruction of oligodendrocytes and myelin
If the inciting injury is simultaneously directed against myelin and oligodendrocytes, sharply demarcated plaques of primary demyelination are induced [180]. Myelin sheaths are completely lost, while oligodendrocyte cell bodies may be partly preserved within active lesion areas. The paradigmatic example for such a mechanism is demyelination triggered by specific antibodies against MOG, an antigen expressed in highest density at the peripheral processes of oligodendrocytes covering the myelin sheath [22]. Upon binding of these antibodies, myelin sheaths are disintegrated by vesicular dissolution in case of massive complement deposition [54] or by phagocytosis of myelin fragments by macrophages (antibody dependent cellular cytotoxicity [21]. Acute injury of oligodendrocytes follow the pathway of necrosis. However, mature oligodendrocytes which have lost their myelin sheaths but survived the initial attack are then slowly removed from the lesions by apoptosis [208]. Following these initial changes, sharply demarcated focal demyelinated lesions are formed. A similar pattern of lesion formation is also seen when myelin and oligodendrocytes are destroyed through antigen-specific cytotoxic T-cells [164] or non-selectively by toxic products of activated macrophages (bystander demyelination). Under the latter conditions, however, the demyelinated lesions are associated with much more widespread damage to other cellular components as well, in particular axons.
Primary oligodendrocyte injury
Distinct types of lesions are seen, when the primary injury is due to a metabolic disturbance of oligodendrocytes, not directly affecting myelin. In this situation, demyelination is frequently incomplete. Such lesions not only contain areas of complete demyelination, but also diffuse myelin pallor is observed. At the edges of the lesions, a moth eaten pattern of demyelination is observed which may reflect the loss of single oligodendrocytes with their respective myelin sheaths. This pattern of demyelination is mainly seen in conditions of infections, which target oligodendrocytes, such as progressive multifocal leukoencephalopathy. In such conditions, virus antigen is abundant in oligodendrocytes and the cells are destroyed both by apoptosis or necrosis [129]. Toxic damage of oligodendrocytes, for example by cuprizone results in similar patterns of demyelination [191].
Demyelination and oligodendrocyte damage induced by mitochondrial injury and/or energy deficiency
Energy deficiency in the white matter leads to a fundamentally different pattern of tissue injury. Also in this condition, oligodendrocytes are highly vulnerable, but they die by a process termed “distal/dying back oligodendrogliopathy”. In initial lesions, the most severely damaged parts of the cells are the most distal (periaxonal) oligodendrocyte processes [1]. This is reflected by a selective loss of proteins, which are predominantly located in this location, such as myelin-associated glycoprotein (MAG) and cyclic nucleotide phosphodiester [1, 68]. Conventional staining for myelin with luxol fast blue shows a diffuse or focal myelin pallor, while immunocytochemistry for proteins located within compact myelin (myelin basic protein or proteolipid protein) is unaffected. With progression of the lesions the majority of oligodendrocytes reveal nuclear condensation and in part nuclear fragmentation in the absence of the expression of activated caspase 3 (caspase independent apoptotic like cell death). Additional characteristic features of such lesions are the preferential destruction and loss of small caliber axons and a remarkable preservation of axons and myelin around larger blood vessels (arterioles and veins). Such tissue changes are the hallmark of initial ischemic lesions of the white matter, and occur within the first hours or days in a white matter stroke lesion [1]. Similar lesions, however, can also be seen in severe inflammatory brain lesions, for example in a subset of patients with acute multiple sclerosis or with virus infections of the white matter like herpes simplex virus encephalitis, cytomegalovirus encephalitis, or progressive multifocal leukoencephalopathy [1]. In the latter lesions, energy deficiency is associated with profound mitochondrial damage, which may at least in part be induced by disturbance of the mitochondrial respiratory chain through reactive oxygen and nitrogen species [114]. Upregulation of molecules which are induced by (hypoxic) tissue preconditioning, such as hypoxia inducible factor 1 alpha or stress proteins in the periphery of such lesions may exert local neuroprotective effects. Moreover, tissue areas with increased resistance to energy deficiency may alternate in the periphery of the lesions with more vulnerable areas, and can give rise to concentric layering of demyelinated and preserved tissue zones typically found in the lesions of Balo’s type of concentric sclerosis [175] (Fig. 2).

Myelin and oligodendrocyte pathology in autoimmune encephalomyelitis, multiple sclerosis, and stroke. a–d Chronic autoimmune encephalomyelitis, induced in DA rat by active sensitization with MOG fusion protein; a massive demyelination is seen in the cerebellar white matter, ×6; b–d oligodendrocytes in different lesion stages of EAE; in the peri-plaque white matter myelin (red) is present and multiple oligodendrocytes with PLP mRNA (black) are seen (b); in the active lesions myelin falls apart, myelin fragments are taken up by macrophages (red granules) and oligodendrocytes are lost (c); in more advanced lesions no macrophages with early myelin degradation products are present; numerous oligodendrocytes re-appear in the lesions, apparently recruited from progenitor cells (black cells), followed by rapid and extensive remyelination; immunocytochemistry for PLP and in situ hybridization for PLP mRNA, ×1000; e–h chronic multiple sclerosis case with extensive remyelination within the CNS; This hemispheric brain section contains 3 active lesions, 4 demyelinated plaques, and 8 remyelinated shadow plaques, ×1.2; f–h double staining for PLP protein (red) and PLP mRNA (black) in one of the active lesions shows a similar pattern as described before in EAE; many oligodendrocytes in the peri-plaque white matter (f); oligodendrocyte loss in the zone of active demyelination (g) and reappearance of oligodendrocytes in the inactive zone, closely adjacent to the zone of activity (h), ×500; i–o myelin changes in the initial stage of a lesion in white matter stroke; LFB shows pale myelin staining (i); the axons, stained with Bielschowsky silver impregnation are largely preserved (k); MAG (l) and CNPase (m) are completely lost from the lesions, while the myelin proteins within the compact sheath (PLP; n) or on the oligodendrocyte surface (MOG; o) are preserved, ×20; p–s acute multiple sclerosis with lesions following a pattern of hypoxia-like tissue injury (Pattern III, [109]). p The section contains areas of initial demyelination (i), early active demyelination (a) and late active or inactive portions (d); q serial section of p, stained by immunocytochemistry for PLP; Only the late active and inactive lesions show loss of PLP; in the active portions (a) a minor loss of PLP reactivity is seen, while in the initial lesions PLP reactivity is the same as in the normal appearing white matter, ×3; r and s edge of an active lesion showing partial preservation of immunoreactivity for MOG (r), but extensive and complete loss of MAG(s), ×20
It has to be emphasized that these distinct patterns of demyelination segregate well in the initial stages of lesion formation and in patients with rapidly progressive white matter disease. However, in more slowly expanding lesions, these morphological features may in part be lost. Then, it may become difficult to determine the mechanism of tissue injury purely on morphological grounds. The final outcome of all the lesions described above is focal or diffuse areas of primary demyelination in the white matter.
Remyelination
Remyelination, the restoration of new myelin sheaths to demyelinated axons, is not performed by pre-existing mature oligodendrocytes [84, 144, 170, 185], but involves in most cases the generation of new mature oligodendrocytes from the adult, quiescent OPC pool distributed throughout the CNS [28, 46, 56, 61, 102, 136, 200, 201, 212]. In the corpus callosum, remyelinating oligodendrocytes can also be derived from stem and precursor cells of the adult subventricular zone [50, 131]. The process of remyelination takes place in several different steps. First, local adult OPCs must switch from an essentially quiescent state to a regenerative phenotype [50]. This transition seems to be triggered by factors derived from activated microglia cells and astrocytes [57, 156], and not by the demyelination per se [134], and leads to OPC proliferation and recruitment to demyelinated areas [50]. Then, the differentiation of OPCs to remyelinating oligodendrocytes starts. All following steps—the interactions with unmyelinated axons, the expression of myelin genes, the elaboration, wrapping and compacting of myelin membrane to form myelin sheaths are similar in myelinating OPCs during development, and in remyelinating OPCs during the regenerative process [50]. However, some differences between myelination and remyelination exist:
-
1.
Adult OPCs have a longer cell cycle time and a slower rate of migration [209].
-
2.
The requirements for transcription factor usage seem to be different. Studies in genetically modified mice clearly revealed that the lack of the oligodendrocyte-lineage specific transcription factor olig1 is incompatible with myelination of the brain [210]. However, when this lack is compensated by the overexpression of the oligodendrocyte-lineage specific transcription factor olig2 (as was probably the case in earlier studies, due to the usage of a particular gene targeting cassette [107, 210]), the mice were able to myelinate during development [7], but were unable to repair demyelinated lesions by remyelination [7].
-
3.
Notch, the regulator of oligodendrocyte differentiation in development (see above), is dispensable during remyelination [179].
-
4.
The correlation between axon diameter and myelin sheath thickness and length seen during developmental myelination is less apparent in remyelination, resulting in thinner and shorter sheath segments [50, 112]. The mechanisms underlying this observation remain unclear, but could involve signals obtained from dynamically growing and changing axons with a need for myelination along their entire length during development, or from mature axons focally lacking myelin sheaths during remyelination [50].
Thus, the pathological hallmark of remyelination in the CNS is the presence of axons with unusually thin myelin sheaths in relation to their caliber [182]. This is best seen by an increase of the G-ratio (the ratio between axonal diameter and myelinated fiber diameter). Unequivocal identification of remyelination in conditions of diffuse demyelination is possible at early stages, but very difficult in older established lesions. In the latter situation detailed quantitative electron microscopic studies may be necessary to show differences in the G-ratio in affected areas. In areas of focal demyelination, such as those occurring in multiple sclerosis, remyelination is reflected by shadow plaques (Markschattenherde, [166]). These are MS-typical focal, sharply demarcated white matter lesions, characterized by uniformly thin myelin sheaths [149].
Remyelination has been extensively studied in MS. In accordance with the basic concepts described above, the recruitment of OPCs and early remyelination is extensive in very early stages of demyelination, in lesions which are still infiltrated by macrophages and lymphocytes [99, 147, 151], and in plaques formed at early stages of the disease. In these fresh lesions, remyelination might be facilitated by inflammation and infiltrating macrophages which provide the tissue with growth factors [39, 92]. Remyelination largely fails at the later (progressive) stage of the disease [59]. This failure of remyelination may be additionally ascribed to age [153, 169], to age-associated changes in the growth factor responsiveness of adult OPCs, and to less efficient clearance of myelin debris from the lesions which has been shown to inhibit remyelination in experimental models [93, 169]. Progressive axonal loss in the lesions, an inability of demyelinated axons to interact with myelinating cells [33], or the presence of myelination inhibiting factors in the extracellular space [204] may additionally impair the capacity of remyelination. These speculations are further corroborated by the observations that mature oligodendrocytes found in active lesions slowly disappear from established lesions [208], and that the OPCs found in late demyelinated lesions seem to be impaired in their further differentiation to mature myelin forming oligodendrocytes [31, 94]. Moreover, it has also been observed that both the numbers and the differentiation stages of OPCs and mature oligodendrocytes are highly variable within lesions of different patients and in different lesion stages. This suggests that different mechanisms of demyelination may have different effects on the remyelinating capacity of lesions [108].
Thus, major efforts are invested to find new neuroprotective therapies, which stimulate myelin repair and by this halt progressive degeneration of chronically demyelinated axons. However, recent studies suggest that the situation in MS patients might be more complicated than previously anticipated. These studies show that extensive remyelination is even seen in a subset of patients, who died at the late progressive stage of the disease. The extent of remyelination in these patients was variable, depending upon the location of the plaques in the brain and spinal cord [142, 143]. Extensive remyelination was predominantly seen in forebrain lesions, located in the subcortical and deep white matter, while it was rather sparse in periventricular areas, the brain stem, and the spinal cord. These data indicate that the capacity of OPCs to differentiate into remyelinating cells is regionally different, possibly related to intrinsic differences in different oligodendrocyte populations as discussed above.
Another important factor is the instability of newly formed myelin in MS lesions, which still show inflammatory and demyelinating activity. New demyelinating activity in previously remyelinated shadow plaques has been unequivocally documented in MS [148], and areas of remyelination are more frequently affected by new inflammatory demyelination than the normal appearing white matter [17]. However, the instability of newly formed myelin in MS lesions also crucially depends on an active inflammatory environment. When inflammation subsides at very late stages of the disease [51], myelin repair seems to be long lasting and stable [143].
Conclusions
Oligodendrocyte biology, myelination, and maintenance of myelin sheaths are very complex processes and their disturbances are associated with major diseases of the nervous system. Intensive research efforts, performed during the last decades have clarified basic principles of these processes and offer new avenues for therapeutic interventions. Much less, however, is known so far on the exact role of these processes in the different diseases of the nervous system. Addressing these questions will be the major challenge for the near future.
References
Aboul-Enein F, Rauschka H, Kornek B, Stadelmann C, Stefferl A, Bruck W, Lucchinetti C, Schmidbauer M, Jellinger K, Lassmann H (2003) Preferential loss of myelin-associated glycoprotein reflects hypoxia-like white matter damage in stroke and inflammatory brain diseases. J Neuropathol Exp Neurol 62:25–33
Ainger K, Avossa D, Morgan F, Hill SJ, Barry C, Barbarese E, Carson JH (1993) Transport and localization of exogenous myelin basic protein mRNA microinjected into oligodendrocytes. J Cell Biol 123:431–441
Ainger K, Avossa D, Diana AS, Barry C, Barbarese E, Carson JH (1997) Transport and localization elements in myelin basic protein mRNA. J Cell Biol 138:1077–1087
Alberdi E, Sánchez-Gómez MV, Torre I, Domercq M, Pérez-Samarín A, Pérez-Cerdá F, Matute C (2006) Activation of kainate receptors sensitizes oligodendrocytes to complement attack. J Neurosci 26:3220–3228
Alonso G (2005) NG2 proteoglycan-expressing cells of the adult rat brain: possible involvement in the formation of glial scar astrocytes following stab wound. Glia 49:318–338
Anderson SA, Eisenstat DD, Shi L, Rubenstein JL (1997) Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science 278:474–476
Arnett HA, Fancy SPJ, Alberta JA, Zhao C, Plant SR, Kaing S, Raine CS, Rowitch DH, Franklin RJM, Stiles CD (2004) bHLH transcription factor olig1 is required to repair demyelinated lesions in the CNS. Science 306:2111–2115
Barres BA (2008) The mystery and magic of glia: a perspective on their roles in health and disease. Neuron 60:430–440
Barres BA, Raff MC (1993) Proliferation of oligodendrocyte precursor cells depends on electrical activity in axons. Nature 361:258–260
Bauer J, Bradl M, Klein M, Leisser M, Deckwerth TL, Wekerle H, Lassmann H (2002) Endoplasmic reticulum stress in PLP-overexpressing transgenic rats: gray matter oligodendrocytes are more vulnerable than white matter oligodendrocytes. J Neuropathol Exp Neurol 61:12–22
Baumann N, Pham-Dinh D (2001) Biology of oligodendrocyte and myelin in the mammalian central nervous system. Physiol Rev 81:871–927
Belachew S, Chittajallu R, Aguirre AA, Yuan X, Kirby M, Anderson S, Gallo V (2003) Postnatal NG2 proteoglycan-expressing progenitor cells are intrinsically multipotent and generate functional neurons. J Cell Biol 161:169–186
Berger T, Frotscher M (1994) Distribution and morphological characteristics of oligodendrocytes in the rat hippocampus in situ and in vitro: an immunocytochemical study with the monoclonal Rip antibody. J Neurocytol 23:61–74
Bergles DW, Roberts JD, Somogyi P, Jahr CE (2000) Glutamatergic synapses on oligodendrocyte precursor cells in the hippocampus. Nature 405:187–191
Borrelli E, Langley OK, Ghandour MS, Delaunoy JP, Gombos G (1982) Immunocytology of carbonic anhydrase II in the central nervous system of jimpy mutant mice. Neurosci Lett 32:321–327
Bradl M, Bauer J, Inomata T, Zielasek J, Nave KA, Toyka K, Lassmann H, Wekerle H (1999) Transgenic Lewis rats overexpressing the proteolipid protein gene: myelin degeneration and its effect on T cell-mediated experimental autoimmune encephalomyelitis. Acta Neuropathol (Berl) 97(6):595–606
Bramow S, Frischer JM, Sorensen PS, Lassmann H, Koch-Henriksen N, Laursen H (2009) Remyelinated plaques in progressive multiple sclerosis are susceptible to second hit active demyelination. Am Acad Neurol 61. Ann. Meeting, P03.058
Braughler JM, Duncan LA, Chase RL (1986) The involvement of iron in lipid peroxidation. J Biol Chem 261:10282–10289
Brinkmann BG, Agarwal A, Sereda MW, Garratt AN, Müller T, Wende H, Stassart RM, Nawaz S, Humml C, Velanac V, Radyushkin K, Goebbels S, Fischer TM, Franklin RJ, Lai C, Ehrenreich H, Birchmeier C, Schwab MH, Nave KA (2008) Neuregulin-1/ErbB signaling serves distinct functions in myelination of the peripheral and central nervous system. Neuron 59:581–595
Brogi A, Strazza M, Melli M, Costantino-Ceccarini E (1997) Induction of intracellular ceramide by interleukin-1 beta in oligodendrocytes. J Cell Biochem 66:532–541
Brosnan CF, Stoner GL, Bloom BR, Wisniewski HM (1977) Studies on demyelination by activated lymphocytes in the rabbit eye. II. Antibody-dependent cell-mediated demyelination. J Immunol 118:2103–2110
Brunner C, Lassmann H, Waehneldt TV, Matthieu JM, Linington C (1989) Differential ultrastructural localization of myelin basic protein, myelin/oligodendroglial glycoprotein, and 2′, 3′-cyclic nucleotide 3′-phosphodiesterase in the CNS of adult rats. J Neurochem 52:296–304
Bugga L, Gadient RA, Kwan K, Stewart CL, Patterson PH (1998) Analysis of neuronal and glial phenotypes in brains of mice deficient in leukemia inhibitory factor. J Neurobiol 36:509–524
Butt AM, Hamilton N, Hubbard P, Pugh M, Ibrahim M (2005) Synantocytes: the fifth element. J Anat 207:695–706
Cai J, Qi Y, Hu X, Tan M, Liu Z, Zhang J, Li Q, Sander M, Qiu M (2005) Generation of oligodendrocyte precursor cells from mouse dorsal spinal cord independent of Nkx6 regulation and Shh signaling. Neuron 45:41–53
Calver AR, Hall AC, Yu WP, Walsh FS, Heath JK, Betsholtz C, Richardson WD (1998) Oligodendrocyte population dynamics and the role of PDGF in vivo. Neuron 20:869–882
Carelli V, Ross-Cisneros FN, Sadun AA (2002) Optic nerve degeneration and mitochondrial dysfunction: genetic and acquired optic neuropathies. Neurochem Int 40:573–584
Carroll WM, Jennings AR, Ironside LJ (1998) Identification of the adult resting progenitor cell by autoradiographic tracking of oligodendrocytes precursors in experimental CNS demyelination. Brain 121:293–302
Cassiani-Ingoni R, Coskaygan T, Xue H, Reichert-Scrivner SA, Wiendl H, Rao MS, Magnus T (2006) Cytoplasmic translocation of Olig2 in adult glial progenitors marks the generation of reactive astrocytes following autoimmune inflammation. Exp Neurol 201:349–358
Cenci di Bello I, Dawson MRL, Levine JM, Reynolds R (1999) Generation of oligodendroglial progenitors in acute inflammatory demyelinating lesions of the rat brain stem is stimulated by demyelination rather than inflammation. J Neurocytol 28:365–381
Chang A, Tourtelotte WW, Rudick RA, Trapp BD (2002) Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N Engl J Med 346:165–200
Charles P, Hernandez MP, Stankoff B, Aigrot MS, Colin C, Rougon G, Zalc B, Lubetzki C (2000) Negative regulation of central nervous system myelination by polysialylated-neural cell adhesion molecule. Proc Natl Acad Sci USA 97:7585–7590
Charles P, Reynolds R, Seilhean D, Rougon G, Aigrot MS, Niezgoda A, Zalc B, Lubetzki C (2002) Re-expression of PSA-NCAM by demyelinated axons: an inhibitor of remyelination in multiple sclerosis? Brain 125:1972–1979
Cheepsunthorn P, Palmer C, Connor JR (1998) Cellular distribution of ferritin subunits in postnatal rat brain. J Comp Neurol 400:73–86
Colello RJ, Pott U, Schwab ME (1994) The role of oligodendrocytes and myelin on axon maturation in the developing rat retinofugal pathway. J Neurosci 14:2594–2605
Connor RR, Menzies SL (1996) Relationship of iron to oligodendrocytes and myelination. Glia 17:83–93
de Castro F, Bribián A (2005) The molecular orchestra of the migration of oligodendrocyte precursors during development. Brain Res Rev 49:227–241
Demerens C, Stankoff B, Logak M, Anglade P, Allinquant B, Couraud F, Zalc B, Lubetzki C (1996) Induction of myelination in the central nervous system by electrical activity. Proc Natl Acad Sci USA 93:9887–9892
Diemel LT, Copelman CA, Cuzner ML (1998) Macrophages in CNS remyelination: friend or foe? Neurochem Res 23:341–347
Domercq M, Sanchez-Gomez MV, Sherwin C, Etxebarria E, Fern R, Matute C (2007) System xc- and glutamate transporter inhibition mediates microglial toxicity to oligodendrocytes. J Immunol 178:6549–6556
Dougherty KD, Dreyfus CF, Black IB (2000) Brain-derived neurotrophic factor in astrocytes, oligodendrocytes, and microglia/macrophages after spinal cord injury. Neurobiol Dis 7:574–585
Du Y, Dreyfus CF (2002) Oligodendrocytes as providers of growth factors. J Neurosci Res 68:647–654
Dubois-Dalcq M, Niedieck B, Buyse M (1970) Action of anti-cerebroside sera on myelinated nervous tissue cultures. Pathol Eur 5:331–347
Dupree JL, Girault JA, Popko B (1999) Axo-glial interactions regulate the localization of axonal paranodal proteins. J Cell Biochem 147:1145–1152
Edgar JM, McLaughlin M, Yool D, Zhang SC, Fowler JH, Montague P, Barrie JA, McCulloch MC, Duncan ID, Garbern J, Nave KA, Griffiths IR (2004) Oligodendroglial modulation of fast axonal transport in a mouse model of hereditary spastic paraplegia. J Cell Biol 166:121–131
Fancy SPJ, Zhao C, Franklin RJM (2004) Increased expression of Nkx2.2 and Olig2 identifies reactive oligodendrocyte progenitor cells responding to demyelination in the adult CNS. Mol Cell Neurosci 27:247–254
Filipovic R, Jakovcevski I, Zecevic N (2003) GRO-alpha and CXCR2 in the human fetal brain and multiple sclerosis lesions. Dev Neurosci 25:279–290
Fitzner D, Schneider A, Kippert A, Möbius W, Willig KI, Hell SW, Bunt G, Gaus K, Simons M (2006) Myelin basic protein-dependent plasma membrane reorganization in the formation of myelin. EMBO J 25:5037–5048
Fogarty M, Richardson WD, Kessaris N (2005) A subset of oligodendrocytes generated from radial glia in the dorsal spinal cord. Development 132:1951–1959
Franklin RJM, Ffrench-Constant C (2008) Remyelination in the CNS: from biology to therapy. Nat Rev Neurosci 9:839–855
Frischer JM, Bramow S, Dal-Bianco A, Lucchinetti C, Rauschka H, Schmidbauer M, Laursen H, Sorensen PS, Lassmann H (2009) The relation between inflammation and neurodegeneration in multiple sclerosis brains. Brain 132:1175–1189
Frost E, Kiernan BW, Faissner A, Ffrench-Constant C (2009) Regulation of oligodendrocyte precursor migration by extracellular matrix: evidence for substrate-specific inhibition of migration by tenascin-C. Dev Neurosci 18:266–273
Garbern JY, Yool DA, Moore GJ, Wilds IB, Faulk MW, Klugmann M, Nave KA, Sistermans EA, van der Knaap MS, Bird TD, Shy ME, Kamholz JA, Griffiths IR (2002) Patients lacking the major CNS myelin protein, proteolipid protein 1, develop length-dependent axonal degeneration in the absence of demyelination and inflammation. Brain 125:551–561
Genain CP, Cannella B, Hauser SL, Raine CS (1999) Identification of autoantibodies associated with myelin damage in multiple sclerosis. Nat Med 5:170–175
Genoud S, Lappe-Siefke C, Goebbels S, Radtke F, Aguet M, Scherer SS, Suter U, Nave KA, Mantei N (2002) Notch1 control of oligodendrocyte differentiation in the spinal cord. J Cell Biol 158:709–718
Gensert JM, Goldman JE (1997) Endogenous progenitors remyelinate demyelinated axons in the adult CNS. Neuron 19:197–203
Glezer I, Lapointe A, Rivest S (2006) Innate immunity triggers oligodendrocyte progenitor reactivity and confines damages to brain injuries. FASEB J 20:750–752
Goldbaum O, Jensen PH, Richter-Landsberg C (2008) The expression of tubulin polymerization promoting protein TPPP/p25alpha is developmentally regulated in cultured rat brain oligodendrocytes and affected by proteolytic stress. Glia 56:1736–1746
Goldschmidt T, Antel J, König FB, Bruck W, Kuhlmann T (2009) Remyelination capacity of the MS brain decreases with disease chronicity. Neurology 72:1914–1921
Griffiths I, Klugmann M, Anderson T, Yool D, Thomson C, Schwab MH, Schneider A, Zimmermann F, McCulloch M, Nadon N, Nave KA (1998) Axonal swellings and degeneration in mice lacking the major proteolipid of myelin. Science 280:1610–1613
Groves AK, Barnett SC, Franklin RJM, Crang AJ, Mayer M, Blakemore WF, Noble M (1993) Repair of demyelinated lesions by transplantation of purified O-2A progenitor cells. Nature 362:453–455
Gyllensten L, Malmfors T (2009) Myelination of the optic nerve, its dependence on visual function: a quantitative investigation in mice. J Embryol Exp Morphol 11:255–256
Hirner A (1969) Elektronenmikroskopische Untersuchungen zur formalen Genese der Balkenläsionen nach experimenteller Cyanvergiftung. Acta Neuropathol (Berl) 13:350–368
Höftberger R, Aboul-Enein F, Bruck W, Lucchinetti C, Rodrigues M, Schmidbauer M, Jellinger K, Lassmann H (2004) Expression of major histocompatibility complex class I molecules on the different cell types in multiple sclerosis lesions. Brain Pathol 14:43–50
Horiuchi M, Itoh A, Pleasure D, Itoh T (2006) MEK-ERK signaling is involved in interferon-gamma-induced death of oligodendroglial progenitor cells. J Biol Chem 281:20095–20106
Huseby ES, Liggitt D, Brabb T, Schnabel B, Öhlen C, Goverman J (2001) A pathogenic role for myelin-specific CD8+ T cells in a model for multiple sclerosis. J Exp Med 194:669–676
Ishibashi T, Dakin KA, Stevens B, Lee PR, Kozlov SV, Stewart CL, Fields RD (2006) Astrocytes promote myelination in response to electrical impulses. Neuron 49:823–832
Itoyama Y, Sternberger NH, Webster HD, Quarles RH, Cohen SR, EP Richardson Jr (1980) Immunocytochemical observations on the distribution of myelin-associated glycoprotein and myelin basic protein in multiple sclerosis lesions. Ann Neurol 7:167–177
Jabs R, Pivneva T, Hüttmann K, Wyczynski A, Nolte C, Kettenmann H, Steinhäuser C (2005) Synaptic transmission onto hippocampla glial cells with hGFAP promoter activity. J Cell Sci 118:3791–3803
Jakovcevski I, Zecevic N (2005) Olig transcription factors are expressed in oligodendrocyte and neuronal cells in human fetal CNS. J Neurosci 25:10064–10073
Jakovcevski I, Zecevic N (2005) Sequence of oligodendrocyte development in the human fetal telencephalon. Glia 49:480–491
Jakovcevski I, Filipovic R, Mo Z, Rakic S, Zecevic N (2009) Oligodendrocyte development and the onset of myelination in the human fetal brain. Front Neuroanat 3:1–15
Jana A, Pahan K (2007) Oxidative stress kills human primary oligodendrocytes via neutral sphingomyelinase: implications for multiple sclerosis. J Neuroimmune Pharmacol 2:184–193
Jarjour AA, Manitt C, Moore SW, Thompson KM, Yuh SJ, Kennedy TE (2003) Netrin-1 is a chemorepellent for oligodendrocyte precursor cells in the embryonic spinal cord. J Neurosci 23:3735–3744
Jurewicz A, Matysiak M, Tybor K, Kilianek L, Raine CS, Selmaj K (2005) Tumour necrosis factor-induced death of adult human oligodendrocytes is mediated by apoptosis inducing factor. Brain 128:2675–2688
Juurlink BH (1997) Response of glial cells to ischemia: roles of reactive oxygen species and glutathione. Neurosci Biobehav Rev 21:151–166
Kalsi AS, Greenwood K, Wilkin G, Butt AM (2004) Kir4.1 expression by astrocytes and oligodendrocytes in CNS white matter: a developmental study in the rat optic nerve. J Anat 204:475–485
Kaplan MR, Meyer-Franke A, Lambert S, Bennett V, Duncan ID, Levinson SR, Barres BA (1997) Induction of sodium channel clustering by oligodendrocytes. Nature 386:724–728
Kaplan MR, Cho MH, Ullian EM, Isom LL, Levinson SR, Barres BA (2001) Differential control of clustering of the sodium channels Na(v)1.2 and Na(v)1.6 at developing CNS nodes of Ranvier. Neuron 30:105–119
Karadottir R, Cavelier P, Bergersen LH, Attwell D (2005) NMDA receptors are expressed in oligodendrocytes and activated in ischaemia. Nature 438:1162–1166
Karadottir R, Hamilton NB, Bakiri Y, Attwell D (2009) Spiking and nonspiking classes of oligodendrocyte precursor glia in CNS white matter. Nat Neurosci 11:450–456
Kassmann CM, Nave KA (2008) Oligodendroglial impact on axonal function and survival—a hypothesis. Curr Opin Neurol 21:235–241
Kassmann CM, Lappe-Siefke C, Baes M, Brügger B, Mildner A, Werner HB, Natt O, Michaelis T, Prinz M, Frahm J, Nave KA (2009) Axonal loss and neuroinflammation caused by preoxisome-deficient oligodendrocytes. Nat Genet 39:969–976
Keirstead HS, Blakemore WF (1997) Identification of post-mitotic oligodendrocytes incapable of remyelination within the demyelinated adult spinal cord. J Neuropathol Exp Neurol 56:1191–1201
Keirstead HS, Levine JM, Blakemore WF (1998) Response of the oligodendrocyte progenitor cell population (defined by NG2 labelling) to demyelination of the adult spinal cord. Glia 22:161–170
Kessaris N, Fogarty M, Iannarelli P, Grist M, Wegner M, Richardson WD (2006) Competing waves ofl oligodendorcytes in the forebrain and postnatal elimination of an embryonic lineage. Nat Neurosci 9:173–179
Kida E, Palminiello S, Golabek AA, Walus M, Wierzba-Bobrowicz T, Rabe A, Albertini G, Wisniewski KE (2006) Carbonic anhydrase II in the developing and adult human brain. J Neuropathol Exp Neurol 65:664–674
Kiernan BW, Gotz B, Faissner A, Ffrench-Constant C (1996) Tenascin-C inhibits oligodendrocyte precursor cell migration by both adhesion-dependent and adhesion-independent mechanisms. Mol Cell Neurosci 7:322–335
Kim JY, Sun Q, Oglesbee M, Yoon SO (2003) The role of ErbB2 signaling in the onset of terminal differentiation of oligodendrocytes in vivo. J Neurosci 23:5561–5571
Kirkpatrick LL, Witt AS, Payne HR, Shine HD, Brady ST (2001) Changes in microtubule stability and density in myelin-deficient shiverer mouse CNS axons. J Neurosci 21:2288–2297
Kiss JZ, Wang C, Olive S, Rougon G, Lang J, Baetens D, Harry D, Pralong W–F (1994) Activity-dependent mobilization of the adhesion molecule polysialic NCAM to the cell surface of neurons and endocrine cells. EMBO J 13:5284–5292
Kotter MR, Setzu A, Sim FJ, Van Rooijen N, Franklin RJM (2001) Macrophage depletion impairs oligodendrocyte remyelination following lysolecithin-induced demyelination. Glia 35:204–212
Kotter MR, Li WW, Zhao C, Franklin RJM (2006) Myelin impairs CNS remyelination by inhibiting oligodendrocyte precursor cell differentiation. J Neurosci 26:328–332
Kuhlmann T, Miron V, Cuo Q, Wegner C, Antel J, Brück W (2008) Differentiation block of oligodendroglial progenitor cells as a cause for remyelination failure in chronic multiple sclerosis. Brain 131:1749–1758
Kukley M, Capetillo-Zarate E, Dietrich D (2007) Vesicular glutamate release from axons in white matter. Nat Neurosci 10:311–320
Kuperman AS, Volpert WA, Okamoto M (1964) Release of adenine nucleotides from nerve axons. Nature 204:1000–1001
Landmesser L, Dahm L, Tang JC, Rutishauser U (1990) Polysialic acid as a regulator of intramuscular nerve branching during embryonic development. Neuron 4:655–667
Lappe-Siefke C, Goebbels S, Gravel M, Nicksch E, Lee J, Braun PE, Griffiths IR, Nave KA (2003) Disruption of CNP1 uncouples oligodendroglial functions in axonal support and myelination. Nat Genet 33:366–374
Lassmann H (1983) Comparative neuropathology of chronic experimental allergic encephalomyelitis and multiple sclerosis. Springer Verlag
Lennon VA, Wingerchuck DN, Kryzer TJ, Pittock SJ, Lucchinetti CF, Fujihara K, Nakashima I, Weinshenker B (2004) A serum autoantibody marker of neuromyelitis potica: distinction from multiple sclerosis. Lancet 264:2106–2112
Lennon VA, Kryzer TJ, Pittock SJ, Verkman AS, Hinson SR (2005) IgG marker of optic-spinal multiple sclerosis binds to the aquaporin-4 water channel. J Exp Med 202:473–477
Levine JM, Reynolds R (1999) Activation and proliferation of endogenous oligodendrocyte precursor cells during ethidium bromide-induced demyelination. Exp Neurol 160:333–347
Li S, Stys PK (2000) Mechanisms of ionotropic glutamate receptor-mediated excitotoxicity in isolated spinal cord white matter. J Neurosci 20:1190–1198
Liedtke W, Edelmann W, Bieri PL, Chiu FC, Cowan NJ, Kucherlapati R, Raine CS (1996) GFAP is necessary for the integrity of CNS white matter architecture and long-term maintenance of myelination. Neuron 17:607–615
Linington C, Bradl M, Lassmann H, Brunner C, Vass K (1988) Augmentation of demyelination in rat acute allergic encephalomyelitis by circulating mouse monoclonal antibodies directed against a myelin/oligodendrocyte glycoprotein. Am J Pathol 130:443–454
Lisak RP, Benjamins JA, Bealmear B, Nedelkoska L, Studzinski D, Retland E, Yao B, Land S (2009) Differential effects of Th1, monocyte/macrophage and Th2 cytokine mixtures on early gene expression for molecules associated with metabolism, signaling and regulation in central nervous system mixed glial cell cultures. J Neuroinflammation 6(4)
Lu QR, Sun T, Zhu Z, Ma N, Garcia M, Stiles CD, Rowitch DH (2002) Common developmental requirement for Olig function indicates a motor neuron/oligodendrocyte lineage connection. Cell 109:75–86
Lucchinetti C, Bruck W, Parisi J, Scheithauer B, Rodrigues M, Lassmann H (1999) A quantitative analysis of oligodendrocytes in multiple sclerosis lesions. A study of 113 cases. Brain 122:2279–2295
Lucchinetti C, Bruck W, Parisi J, Scheithauer B, Rodriguez M, Lassmann H (2000) Heterogeneity of multiple sclerosis lesions: implications for the pathogenesis of demyelination. Ann Neurol 47:707–717
Lucchinetti CF, Mandler RN, McGavern D, Bruck W, Gleich G, Ransohoff RM, Trebst C, Weinshenker B, Wingerchuck D, Parisi J, Lassmann H (2002) A role for humoral mechanisms in the pathogenesis of Devic′s neuromyelitis optica. Brain 125:1450–1461
Ludwin SK (1997) The pathobiology of the oligodendrocyte. J Neuropathol Exp Neurol 56:111–124
Ludwin S, Maitland M (1984) Long-term remyelination fails to reconstitute normal thickness of central myelin sheaths. J Neurol Sci 64:193–198
Mahad D, Lassmann H, Turnbull D (2008) Review: mitochondria and disease progression in multiple sclerosis. Neuropathol Appl Neurobiol 34:577–589
Mahad D, Ziabreva I, Lassmann H, Turnbull D (2009) Mitochondrial defects in acute multiple sclerosis lesions. Brain 131:1722–1735
Maire JC, Medilanski J, Straub RW (1984) Release of adenosine, inosine, hypoxanthine from rabbit non-myelinated nerve fibers at rest and during activity. J Physiol 357:67–77
Mason JL, Langaman C, Morell P, Suzuki K, Matsushima GK (2001) Episodic demyelination and subsequent remyelination within the murine central nervous system: changes in axonal calibre. Neuropathol Appl Neurobiol 27:50–58
Matute C, Sanchez-Gomez MV, Martinez-Millan L, Miledi R (1997) Glutamate receptor-mediated toxicity in optic nerve oligodendrocytes. Proc Natl Acad Sci USA 94:8830–8835
Matute C, Torre I, Pérez-Cerdá F, Pérez-Samartín A, Alberdi E, Etxebarria E, Arranz AM, Ravid R, Rodíguez-Antigüedad A, Sánchez-Gómez M, Domercq M (2007) P2X(7) receptor blockade prevents ATP excitotoxicity in oligodendrocytes and ameliorates experimental autoimmune encephalomyelitis. J Neurosci 27:9525–9533
McDonald JW, Althomsons SP, Hyrc KL, Choi DW, Goldberg MP (1998) Oligodendrocytes from forebrain are highly vulnerable to AMPA/kainate receptor-mediated excitotoxicity. Nat Med 4:291–297
McLaurin JA, Yong VW (1995) Oligodendrocytes and myelin. Neurol Clin 13:23–49
McTigue DM, Tripathi RB (2008) The life, death, and replacement of oligodendrocytes in the adult CNS. J Neurochem 107:1–19
Mi S, Miller RH, Lee X, Scott ML, Shulag-Morskaya S, Shao Z, Chang J, Thill G, Levesque M, Zhang M, Hession C, Sah D, Trapp BD, He Z, Jung V, McCoy JM, Pepinsky RB (2005) LINGO-1 negatively regulates myelination by oligodendrocytes. Nat Neurosci 8:745–751
Micu I, Jiang Q, Coderre E, Ridsdale A, Zhang L, Woulfe J, Yin X, Trapp BD, McRory JE, Rehak R, Zamponi GW, Wang W, Stys PK (2006) NMDA receptors mediate calcium accumulation in myelin during chemical ischaemia. Nature 439:988–992
Mignot C, Boespflug-Tanguy O, Gelot A, Dautigny A, Pham-Dinh D, Rodriguez D (2004) Alexander disease: putative mechanisms of an astrocytic encephalopathy. Cell Mol Life Sci 61:369–385
Milner R, Frost E, Nishimura S, Delcommenne C, Streuli C, Pytela R, Ffrench-Constant C (1997) Expression of alpha vbeta3 and alpha vbeta8 integrins during oligodendrocyte precursor differentiation in the presence and absence of axons. Glia 21:350–360
Minkowsky AE (2009) The myelinogenetic cycles in regional maturation of the brain. In regional development of the brain in early life. Blackwell, Oxford, pp 3–70
Misu T, Fujihara K, Kakita A, Konno H, Nakamura M, Watanabe S, Takahashi T, Nakashima I, Takahashi H, Itoyama Y (2007) Loss of aquaporin-4 in lesions of neuromyelitis optica: distinction from multiple sclerosis. Brain 130:1224–1234
Misu T, Fujihara K, Itoyama Y (2008) Neuromyelitis optica and anti-aquaporin 4 antibody—an overview. Brain Nerve 60:527–537
Murayama S, Saito Y (2007) Neuropathology of progressive multifocal leukoencephalopathy. Brain Nerve 59:119–124
Na SY, Cao Y, Toben C, Nitschke L, Stadelmann C, Gold R, Schimpl A, Hünig T (2008) Naive CD8 T-cells initiate spontaneous autoimmunity to a sequestered model antigen of the central nervous system. Brain 131:2353–2365
Nait-Oumesmar B, Picard-Riera N, Kerninon C, Decker L, Seilhean D, Höglinger GU, Hirsch EC, Reynolds R, Baron-van Evercooren A (2007) Activation of the subventricular zone in multiple sclerosis: evidence for early glial progenitors. Proc Natl Acad Sci USA 104:4694–4699
Neusch C, Rozengurt N, Jacobs RE, Lester HA, Kofuji P (2001) Kir4.1 potassium channel subunit is crucial for oligodendrocyte development and in vivo myelination. J Neurosci 21:5429–5438
Niehaus A, Stegmüller J, Diers-Fenger M, Trotter JL (1999) Cell-surface glycoprotein of oligodendrocyte progenitors involved in migration. J Neurosci 19:4948–4961
Nielsen HH, Ladeby R, Drojdahl N, Peterson AC, Finsen B (2006) Axonal degeneration stimulates the formation of NG2+ cells and oligodendrocytes in the mouse. Glia 54:105–115
Nishiyama A, Komitova M, Suzuki R, Zhu X (2009) Polydendrocytes (NG2 cells): multifunctional cells with lineage plasticity. Nat Rev Neurosci 10:9–22
Nunes MC, Roy NS, Keyoung HM, Goodman RR, McKhann G II, Jiang L, Kang J, Nedergaard M, Goldman SA (2003) Identification and isolation of multipotential neural progenitor cells from the subcortical white matter of the adult human brain. Nat Med 9:439–447
O′Connor KC, McLaughlin KA, De Jager PL, Chtinis T, Bettelli E, Xu C, Robinson WH, Cherry SV, Bar-Or A, Banwell B, Fukaura H, Fukazawa T, Tenembaum S, Wong SJ, Tavakoli NP, Idrissova Z, Viglietta V, Rostasy K, Pohl D, Dale RC, Freedman M, Steinman L, Buckle GJ, Kuchroo VK, Hafler DA, Wucherpfennig KW (2007) Self-antigen tetramers discriminate between myelin autoantibodies to native or denatured protein. Nat Med 13:211–217
Ohya W, Funakoshi H, Kurosawa T, Nakamura T (2007) Hepatocyte growth factor (HGF) promotes oligodendrocyte progenitor cell proliferation and inhibits its differentiation during postnatal development in the rat. Brain Res 1147:51–65
Omlin FX (1997) Optic disc and optic nerve of the blind cape molerat (Georychus capensis): a proposed model for naturally occurring reactive gliosis. Brain Res Bull 44:627–632
Ongur D, Drevets WC, Price JL (1998) Glial reduction in the subgenual prefrontal cortex in mood disorders. Proc Natl Acad Sci USA 95:13290–13295
Park S-K, Miller R, Krane I, Vartanian T (2001) The erbB2 gene is required for the development of terminally differentiated spinal cord oligodendrocytes. J Cell Biol 154:1245–1258
Patani R, Balaratnam M, Vora A, Reynolds R (2007) Remyelination can be extensive in multiple sclerosis despite a long disease course. Neuropathol Appl Neurobiol 33:277–287
Patrikios P, Stadelmann C, Kutzelnigg A, Rauschka H, Schmidbauer M, Laursen H, Sorensen PS, Brück W, Lucchinetti C, Lassmann H (2006) Remyelination is extensive in a subset of multiple sclerosis patients. Brain 129:3165–3172
Prayoonwiwat N, Rodriguez M (1993) The potential for oligodendrocyte proliferation during demyelinating disease. J Neuropathol Exp Neurol 52:55–63
Prestoz L, Chatzopoulo E, Lemkine G, Spassky N, Le Bras B, Kagawa T, Ikenaka K, Zalc B, Thomas JL (2004) Control of axonophilic migration of oligodendrocyte precursor cells by Eph-ephrin interaction. Neuron Glia Biol 1:73–83
Preusser M, Lehotzky A, Budka H, Ovadi J, Kovacs GG (2007) TPPP/p25 in brain tumours: expression in non-neoplastic oligodendrocytes but not in oligodendroglioma cells. Acta Neuropathol 113:213–215
Prineas JW, Kwon EE, Goldenberg PZ, Ilyas AA, Quarles RH, Benjamins JA, Sprinkle TJ (1989) Multiple sclerosis. Oligodendrocyte proliferation and differentiation in fresh lesions. Lab Invest 61:489–503
Prineas JW, Barnard RO, Revesz T, Kwon EE, Sharer LR, Cho ES (1993) Multiple sclerosis. Pathology of recurrent lesions. Brain 116:681–693
Prineas JW, Barnard RO, Kwon EE, Sharer LR, Cho ES (1993) Multiple sclerosis: remyelination of nascent lesions. Ann Neurol 33:137–151
Radtke F, Raj K (2003) The role of notch in tumorigenesis: oncogene or tumour suppressor? Nat Rev Cancer 3:756–767
Raine CS, Scheinberg L, Waltz JM (1981) Multiple sclerosis. Oligodendrocyte survival and proliferation in an active established lesion. Lab Invest 45:534–546
Rakic S, Zecevic N (2003) Early oligodendrocyte progenitor cells in the human fetal telencephalon. Glia 41:117–127
Rando TA (2006) Stem cells, ageing and the quest for immortality. Nature 441:1080–1086
Redwine JM, Armstrong RC (1998) In vivo proliferation of oligodendrocyte progenitors expressing PDGFaR during early remyelination. J Neurobiol 37:413–428
Redwine JM, Blinder KL, Armstrong RC (1997) In situ expression of fibroblast growth factor receptors by oligodendrocyte progenitors and oligodendrocytes in adult mouse central nervous system. J Neurosci Res 50:229–237
Rhodes KE, Raivich G, Fawcett JW (2006) The injury response of oligodendrocyte precursor cells is induced by platelets, macrophages and inflammation-associated cytokines. Neuroscience 140:87–100
Richardson WD, Kessaris N, Pringle N (2006) Oligodendrocyte wars. Nat Rev Neurosci 7:11–18
Roemer SF, Parisi JE, Lennon VA, Mandler R, Weinshenker BG, Benarroch E, Pittock SJ, Wingerchuck DM, Lassmann H, Bruck W, Lucchinetti CF (2007) Distinct pattern of aquaporin-4 expression in neuromyelitis optica lesions. Brain 130:1194–1205
Roy K, Murtie JC, El-Khodor BF, Edgar N, Sardi SP, Hooks BM, Benoit-Marand M, Hen C, Moore H, O`Donnell P, Brunner D, Corfas G (2007) Loss of erbB signaling in oligodendrocytes alters myelin and dopaminergic function, a potential mechanism for neuropsychiatric disorders. Proc Natl Acad Sci USA 104:8131–8136
Salter MG, Fern R (2005) NMDA receptors are expressed in developing oligodendrocyte processes and mediate injury. Nature 438:1167–1171
Sanchez I, Hassinager L, Paskevich PA, Shine HD, Nixon RA (1996) Oligodendroglia regulate the regional expansion of axon caliber and local accumulation of neurofilaments during development independently of myelin formation. J Neurosci 16:5095–5105
Sanchez I, Hassinger L, Sihag RK, Cleveland DW, Mohan P, Nixon RA (2000) Local control of neurofilament accumulation during radial growth of myelinating axons in vivo. J Cell Biol 151:1013–1024
Sanchez-Gomez MV, Matute C (1999) AMPA and kainate receptors each mediate excitotoxicity in oligodendroglial cultures. Neurobiol Dis 6:475–485
Saxena A, Bauer J, Scheikl T, Zappula J, Audebert M, Desbois S, Waisman A, Lassmann H, Liblau R, Mars LT (2008) Cutting edge: multiple sclerosis-like lesions induced by effector CD8 T cells recognizing a sequestered antigen on oligodendrocytes. J Immunol 181:1617–1621
Schenck M, Carpinteiro A, Grassme H, Lang F, Gulbins E (2007) Ceramide: physiological and pathophysiological aspects. Arch Biochem Biophys 462:171–175
Schlesinger H (1909) Zur Frage der akuten Multiplen Sklerose und der encephalomyelitis disseminata im Kindesalter. Arb Neurol Inst (Wien) 17:410–432
Schnadelbach O, Blaschuk OW, Symonds M, Gour BJ, Doherty P, Fawcett JW (2000) N-cadherin influences migration of oligodendrocytes on astrocyte monolayers. Mol Cell Neurosci 15:288–302
Scolding NJ, Rayner PJ, Sussman J, Shaw C, Compston DA (1995) A proliferative adult human oligodendrocyte progenitor. Neuroreport 6:441–445
Shields SA, Gilson JM, Blakemore WF, Franklin RJM (1999) Remyelination occurs as extensively but more slowly in old rats compared to young rats following gliotoxin-induced CNS demyelination. Glia 28:77–83
Sim FJ, Zhao C, Penderis J, Franklin RJM (2002) The age-related decrease in CNS remyelination efficiency is attributable to an impairment of both oligodendrocyte progenitor recruitment and differentiation. J Neurosci 22:2451–2459
Simons M, Trajkovic K (2006) Neuron-glia communication in the control of oligodendrocyte function and myelin biogenesis. J Cell Biochem 119:4381–4389
Singh I, Pahan K, Khan M, Singh AK (1998) Cytokine-mediated induction of ceramide production is redox-sensitive. Implications to proinflammatory cytokine-mediated apoptosis in demyelinating diseases. J Biol Chem 273:20354–20362
Smith KJ, Lassmann H (2002) The role of nitric oxide in multiple sclerosis. Lancet Neurol 1:232–241
Spassky N, Heydon A, Mangatal A, Jankovski A, Olivier C, Queraud-Lesaux F, Goujet-Zalc C, Thomas JL, Zalc B (2001) Sonic hedgehog-dependent emergence of oligodendrocytes in the telencephalon: evidence for a source of oligodendrocytes in the olfactory bulb that is independent of PDGFRa signaling. Development 128:4993–5004
Stadelmann C, Ludwin S, Tabira T, Guseo A, Lucchinetti CF, Leel-Ossy L, Ordinario AT, Bruck W, Lassmann H (2005) Tissue preconditioning may explain concentric lesions in Balo’s type of multiple sclerosis. Brain 128:979–987
Stark AK, Uylings HB, Sanz-Arigita E, Pakkenberg B (2004) Glial cell loss in the anterior cingulate cortex, a subregion of the prefrontal cortex, in subjects with schizophrenia. Am J Psychiatry 161:882–888
Stevens B, Fields RD (2009) Response of Schwann cells to action potentials in development. Science 287:2267–2271
Stevens B, Porta S, Haak LL, Gallo V, Fields RD (2002) Adenosine: a neuron-glial transmitter promoting myelination in the CNS in response to action potentials. Neuron 36:855–868
Stidworthy MF, Genoud S, Li WW, Leone DP, Mantei N, Suter U, Franklin RJM (2004) Notch1 and Jagged 1 are expressed after CNS demyelination but are not a major rate-determining factor during remyelination. Brain 127:1928–1941
Storch M, Stefferl A, Brehm U, Weissert R, Wallstrom E, Kerschensteiner M, Olsson T, Linington C, Lassmann H (1998) Autoimmunity to myelin oligodendrocyte glycoprotein in rats mimics the spectrum of multiple sclerosis pathology. Brain Pathol 8:681–694
Sun T, Pringle NP, Hardy AP, Richardson WD, Smith HK (1998) Pax6 influences the time and site of origin of glial precursors in the ventral neural tube. Mol Cell Neurosci 12:228–239
Suzuki K, Andrews JM, Waltz JM, Terry RD (1969) Ultrastructural studies of multiple sclerosis. Lab Invest 20:444–454
Takebayashi H, Nabeshima Y, Yoshida S, Chisaka O, Ikenaka K, Nabeshima Y (2002) The basic helix-loop-helix factor Olig2 is essential for the development of motoneuron and oligodendrocyte lineages. Curr Biol 12:1157–1163
Tanaka H, Grooms SY, Bennett MV, Zukin RS (2000) The AMPAR subunit GluR2: still front and center-stage. Brain Res 886:190–207
Targett MP, Sussman J, Scolding NJ, O′Leary MT, Compston DA, Blakemore WF (1996) Failure to achieve remyelination of demyelinated rat axons following transplantation of glial cells obtained from the adult human brain. Neuropathol Appl Neurobiol 22:199–206
Tauber H, Waehneldt TV, Neuhoff V (1980) Myelination in rabbit optic nerves is accelerated by artificial eye opening. Neurosci Lett 16:235–238
Taveggia C, Thaker P, Petrylak A, Caporaso GL, Toews A, Falls DL, Einheber S, Salzer JL (2008) Type III neuregulin-1 promotes oligodendrocyte myelination. Glia 56:284–293
Testai FD, Landek MA, Dawson G (2004) Regulation of sphingomyelinases in cells of the oligodendrocyte lineage. J Neurosci Res 75:66–74
Thorburne SK, Juurlink BH (1996) Low glutathione and high iron govern the susceptibility of oligodendroglial precursors to oxidative stress. J Neurochem 67:1014–1022
Tiwari-Woodruff SK, Buznikov AG, Yu Tq, Micevych PE, Chen K, Kornblum HI, Bronstein JM (2001) OSP/claudin-11 forms a complex with a novel member of the tetraspanin super family and beta1 integrin and regulates proliferation and migration of oligodendrocytes. J Cell Biol 153:295–305
Torkildsen O, Brunborg LA, Myhr KM, Bö L (2008) The cuprizone model for demyelination. Acta Neurol Scand Suppl 188:72–76
Trajkovic K, Dhaunchak AS, Goncalves JT, Wenzel D, Schneider A, Bunt G, Nave KA, Simons M (2006) Neuron to glia signaling triggers myelin membrane exocytosis from endosomal storage sites. J Cell Biol 172:937–948
Tsai HH, Tessier-Lavigne M, Miller RH (2003) Netrin-1 mediates spinal cord oligodendrocyte precursor dispersal. Development 130:2095–2105
Uschkureit T, Sporkel O, Stracke J, Bussow H, Stoffel W (2000) Early onset of axonal degeneration in double (plp−/−mag−/−) and hypomyelinosis in triple (plp−/−mbp−/−mag−/−) mutant mice. J Neurosci 20:5225–5233
Vallstedt A, Klos JM, Ericson J (2005) Multiple dorsoventral origins of oligodendrocyte generation in the spinal cord and hindbrain. Neuron 45:55–67
van Heyningen P, Calver AR, Richardson WD (2001) Control of progenitor cell number by mitogen supply and demand. Curr Biol 11:232–241
Vartanian T, Goodearl A, Viehöver A, Fischbach G (1997) Axonal neuregulin signals cells of the oligodendrocyte lineage through activation of HER4 and Schwann cells through HER2 and HER3. J Cell Biol 137:211–220
Vartanian T, Fischbach G, Miller R (1999) Failure of spinal cord oligodendrocyte development in mice lacking neuregulin. Proc Natl Acad Sci USA 96:731–735
Wang C, Rougon G, Kiss JZ (1994) Requirement of polysialic acid for the migration of the O-2A glial progenitor cell from neurohypophyseal explants. J Neurosci 14:4446–4457
Warrington AE, Barbarese E, Pfeiffer SE (1993) Differential myelinogenic capacity of specific developmental stages of the oligodendrocyte lineage upon transplantation into hypomyelinating hosts. J Neurosci Res 34:1–13
Watanabe M, Toyama Y, Nishiyama A (2002) Differentiation of proliferated NG2-positive glial progenitor cells in a remyelinating lesion. J Neurosci Res 69:826–836
Watkins TA, Emery B, Mulinyawe S, Barres BA (2008) Distinct stages of myelination regulated by g-secretase and astrocytes in a rapidly myelinating CNS coculture system. Neuron 60:555–569
Wilkins A, Majed H, Layfield R, Compston A, Chandran S (2003) Oligodendrocytes promote neuronal survival and axonal length by distinct intracellular mechanisms: a novel role for oligodendrocyte-derived glial cell line-derived neurotrophic factor. J Neurosci 23:4967–4974
Williams A, Piaton G, Aigrot MS, Belhadi A, Théaudin M, Petermann F, Thomas J-L, Zalc B, Lubetzki C (2007) Semaphorin 3A and 3F: key players in myelin repair in multiple sclerosis? Brain 130:2554–2565
Wilson HC, Onischke C, Raine CS (2003) Human oligodendrocyte precursor cells in vitro: phenotypic analysis and differential response to growth factors. Glia 44:153–165
Wisniewski HM, Bloom BR (1975) Primary demyelination as a nonspecific consequence of a cell-mediated immune reaction. J Exp Med 141:346–359
Wolswijk G (1998) Chronic stage multiple sclerosis lesions contain a relatively quiescent population of oligodendrocyte precursor cells. J Neurosci 18:601–609
Wolswijk G (2000) Oligodendrocyte survival, loss and birth in lesions of chronic-stage multiple sclerosis. Brain 123:105–115
Wolswijk G, Noble M (1989) Identification of an adult-specific glial progenitor cell. Development 105:387–400
Xin M, Yue T, Ma Z, Wu F-F, Gow A, Lu QR (2005) Myelinogenesis and axonal recognition by oligodendrocytes in brain are uncoupled in olig1-null mice. J Neurosci 25:1354–1365
Yan H, Rivkees SA (2002) Hepatocyte growth factor stimulates the proliferation and migration of oligodendrocyte precursor cells. J Neurosci Res 69:597–606
Zhang SC, Ge B, Duncan ID (1999) Adult brain retains the potential to generate oligodendroglial progenitors with extensive myelination capacity. Proc Natl Acad Sci USA 96:4089–4094
Zhang SC, Ge B, Duncan ID (2000) Tracing human oligodendroglial development in vitro. J Neurosci Res 59:421–429
Zhou Q, Anderson DJ (2002) The bHLH transcription factors Olig2 and Olig1 couple neuronal and glial subtype specification. Cell 109:61–73
Ziskin JL, Nishiyama A, Rubio M, Fukaya M, Bergles DE (2007) Vesicular release of glutamate from unmyelinated axons in white matter. Nat Neurosci 10:321–330
Acknowledgments
The authors were supported by the Fonds zur Förderung der wissenschaftlichen Forschung (FWF, projects P 19854-B02 to H.L. and P 21581-B09 to M.B.).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Appendix 1: Heterogeneity among adult OPCs
Appendix 1: Heterogeneity among adult OPCs
The adult OPCs express the proteoglycan NG2 and the PDGF receptor α [30, 85, 154, 170], and have been given numerous names like NG2-expressing OPCs [50], synantocytes [24], or polydendrocytes [135]. Current experimental evidence suggests that they might be less restricted in their differentiation potential and might not only give rise to oligodendrocytes, but also to neurons in the hippocampus [12] or to astrocytes [5, 29]. They are probably not only heterogenous in terms of the different cell types they can give rise to, but also seem to be heterogenous in function: while some of them become quiescent, others apparently have key physiological roles and are involved in the bi-directional communication between glial cells and neurons since they receive synaptic inputs [14, 69, 80, 95, 215] and are able to generate action potentials [81].
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Bradl, M., Lassmann, H. Oligodendrocytes: biology and pathology. Acta Neuropathol 119, 37–53 (2010). https://doi.org/10.1007/s00401-009-0601-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00401-009-0601-5