
Abstract
We recorded the body axis orientation of free-living black wildebeest relative to incident solar radiation and wind. Observations were made on three consecutive days, on six occasions over the course of 1 year, in a treeless, predominantly cloudless habitat. Frequency of orientation parallel to incident solar radiation increased, and perpendicular to incident solar radiation decreased, as ambient dry-bulb temperature or solar radiation intensity increased, or wind speed decreased. We believe these changes were mediated via their effect on skin temperature. Parallel orientation behavior was more prominent when the wildebeest were standing without feeding than it was when they were feeding. We calculate that a black wildebeest adopting parallel orientation throughout the diurnal period would absorb 30% less radiant heat than the same animal adopting perpendicular orientation. Parallel orientation was reduced at times when water was freely available, possibly reflecting a shift from behavioral to autonomic thermoregulatory mechanisms. The use of orientation behavior by black wildebeest is well developed and forms part of the suite of adaptations that help them to maintain heat balance while living in a shadeless, often hot, environment.







Similar content being viewed by others


References
Acocks JPH (1975) Veld types of South Africa. Mem Bot Surv S Afr 40, 128 pp
Altmann J (1974) Observational study of behaviour: sampling methods. Behavior 49:227–267
Batschelet E (1981) Circular statistics in biology. Academic, London
Ben Shahar R, Fairall N (1987) Comparison of the diurnal activity patterns of blue wildebeest and red hartebeest. S Afr J Wildl Res 17:49–54
Berry HH, Siegfried WR, Crowe TM (1982) Activity patterns in a population of free-ranging wildebeest Connochaetes taurinus at Etosha National Park. Z Tierpsychol 59:229–246
Berry HH, Siegfried WR, Crowe TM (1984) Orientation of wildebeest in relation to sun angle and wind direction. Madoqua 13:297–301
Clapperton JL, Joyce JP, Blaxter KL (1965) Estimates of the contribution of solar radiation to the thermal exchanges of sheep at a latitude of 55° north. J Agric Sci 64:37–49
Clark RG, Ohmart RD (1985) Spread-winged posture of Turkey Vultures: single or multiple function? Condor 87:350–355
David JHM (1973) The behaviour of the bontebok, Damaliscus dorcas dorcas, (Palls 1766), with special reference to territorial behaviour. Z Tierpsychol 33:38–107
Estes RD (1991) The behaviour guide to african mammals. University of California Press Ltd, Oxford, England
Finch VA (1972) Energy exchanges with the environment of two East african antelopes, the eland and the hartebeest. Symp Zool Soc Lond 31:315–326
Frank SM, Raja SN, Bulcao CF, Goldstein DS (1999) Relative contribution of core and cutaneous temperatures to thermal comfort and autonomic responses in humans. J Appl Physiol 86:1588–1593
Hofmeyr MD, Louw GN (1987) Thermoregulation, pelage conductance and renal function in the desert-adapted springbok, Antidorcas marsupialis. J Arid Environ 13:137–151
Hogan AG, Skouby CI (1923) Determination of the surface area of cattle and swine. J Agric Res 25:419–430
Jarman MV, Jarman PJ (1973) Daily activity of impala. East Afr Wildl J 11:75–92
Jessen C, Laburn HP, Knight MH, Kuhnen G, Goelst K, Mitchell D (1994) Blood and brain temperatures of free-ranging black wildebeest in their natural environment. Am J Physiol Regul Integr Comp Physiol 36:R1528–R1536
King JM (1979) Game domestication for animal production in Kenya: field studies of the body-water turnover of game and livestock. J Agric Sci 93:71–79
Lewis JG (1978) Game domestication for animal production in Kenya: shade behaviour and factors affecting the herding of eland, oryx, buffalo and zebu cattle. J Agric Sci 90:587–595
Maloney SK, Dawson TJ (1995) The heat load from solar radiation on a large, diurnally active bird, the emu (Dromaius novaehollandiae). J Therm Biol 20:381–387
Maloney SK, Moss G, Cartmell T, Mitchell D (2005) Alteration in diel activity patterns as a thermoregulatory strategy in black wildebeest (Connochaetes gnou). J Comp Physiol A (in press)
Mitchell A (1977) Preliminary observations on the daytime activity patterns of lesser kudu in Tsavo National Park, Kenya. East Afr Wildl J 15:199–206
Norusis MJ (1992) SPSS for Windows Based System User’s Guide Release 5. SPSS Inc, USA
Rautenberg W, May B, Arabin G (1980) Behavioral and autonomic temperature regulation in competition with food intake and water balance of pigeons. Pflugers Arch Eur J Physiol 384:253–260
Refinetti R, Carlisle HJ (1986) Effects of anterior and posterior hypothalamic temperature changes on thermoregulation in the rat. Physiol Behav 36:1099–1104
Riemerschmid G (1943) Some aspects of solar radiation in its relation to cattle in South Africa and Europe. Onderstepoort J vet Res 18:327–353
Sellers RM (1995) Wing-spreading behaviour of the Cormorant Phalacrocorax carbo. Ardea 83:27–36
Skinner JD, Smithers RHN (1990) The mammals of the southern african subregion. University of Pretoria Press, Pretoria
Torres-Contreras H, Bozinovic F (1997) Food selection in an herbivorous rodent: Balancing nutrition with thermoregulation. Ecology 78:2230–2237
Zar JH (1996) Biostatistical analysis, 3rd edn. Prentice Hall Inc, New Jersey
Acknowledgements
We thank Dr. Mark Berry and DeBeers Consolidated Mines for permission to carry out the study on Benfontein, and Peter, Jennifer, Gregory, and Nicky Gibbs for their friendship and help on site. Tammy Cartmell, Steven Cartmell and Simone Glassom helped with data collection. Petro Vorster from the Kimberley office of the South African Weather Bureau kindly supplied the rainfall data. Professor Phil Withers and an anonymous reviewer helped to improve the manuscript. The study was funded in part by the South African Foundation for Research Development. SKM was in receipt of a University of the Witwatersrand Post Doctoral Fellowship. These experiments comply with the “Principles of animal care” publication number 86–23, revised 1985 of the National Institute of Health, and also the laws of South Africa.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
The heat absorbed from direct solar radiation was calculated for black wildebeest oriented parallel and perpendicular to incident solar radiation for each hour and then summed for the diurnal period for each survey. Heat absorbed was calculated as the product of surface area presented × measured incident solar radiation intensity × absorption coefficient.
-
1.
Solar radiation intensity incident on a horizontal surface was measured at the field site (Fig. 1).
Fig. 8 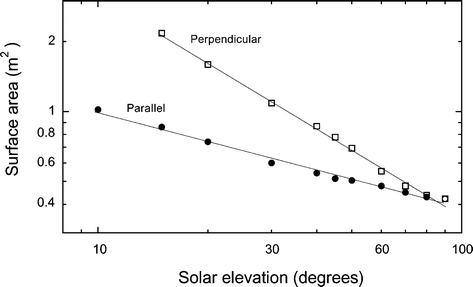
The calculated surface area projected on to a horizontal surface, presented to incident solar radiation by a 130 kg black wildebeest at solar elevation angles between 10° and 90° when oriented perpendicular (squares) or parallel (circles) to incident solar radiation. See Appendix for details of calculations. The best fit for the relationship proved to be a power relationship, as indicated by the linear fit on a log–log plot
-
2.
The absorption coefficient was assumed be 80%, the same as that measured for dark brown cattle (Riemerschmid 1943). The heat load from radiation in dark coats is influenced by wind speed, coat conductance, and the penetrance of radiation into the pelage (Maloney and Dawson 1995). Ignoring these factors would result in an overestimate of the heat load, and so the figures we arrived at would represent the maximal heat load of the orientations. However, the proportional difference between orientations would be independent of these errors.
-
3.
The fraction of the total body surface area presented to incident solar radiation varies with body orientation and solar elevation. Solar elevation was calculated for each hour during each survey for the latitude and longitude of our study site using software available at http://www.susdesign.com/sunangle. The surface area presented at different solar elevations was calculated based on data from Riemerschmid (1943) obtained for an 800 kg bull (total surface area 6.5 m2) oriented either parallel or perpendicular to the solar beam. The mass of an average adult female black wildebeest is 130 kg (Skinner and Smithers 1990). The total surface area of a cow weighing 130 kg is 1.86 m2 (Hogan and Skouby 1923). We assumed a similar geometry for wildebeest and cattle, and so the surface areas presented for cattle was scaled by 1.86/6.5. The best fit was a power equation (Fig. 8). For perpendicular orientation SA (m2)=26.9×solar elevation−0.98 and for parallel orientation SA (m2)=2.54×solar elevation−0.41, with solar elevation in degrees. For each hour (0700 to 1800) during each survey, the surface area presented to incident solar radiation was calculated from these equations for the appropriate solar elevation.
-
4.
Solar radiation absorbed was then calculated as the surface area (m2) × the measured incident solar radiation intensity (W/m2)×0.8. For each day the total absorbed solar energy was calculated by summing the hourly values for over 12 h. These values represent the extremes of the range of orientations, because they assume a fixed orientation for the full day.

Rights and permissions
About this article
Cite this article
Maloney, S.K., Moss, G. & Mitchell, D. Orientation to solar radiation in black wildebeest (Connochaetes gnou). J Comp Physiol A 191, 1065–1077 (2005). https://doi.org/10.1007/s00359-005-0031-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-005-0031-3