
Abstract
Reefs formed by scleractinian cold-water corals represent unique biodiversity hot spots in the deep sea, preferring aphotic water depths of 200–1000 m. The distribution of the most prominent reef-building species Lophelia pertusa is controlled by various environmental factors including dissolved oxygen concentrations and temperature. Consequently, the expected ocean deoxygenation and warming triggered by human-induced global change are considered as a serious threat to cold-water coral reefs. Here, we present results on recently discovered reefs in the SE Atlantic, where L. pertusa thrives in hypoxic and rather warm waters. This sheds new light on its capability to adapt to extreme conditions, which is facilitated by high surface ocean productivity, resulting in extensive food supply. Putting our data in an Atlantic-wide perspective clearly demonstrates L. pertusa’s ability to develop population-specific adaptations, which are up to now hardly considered in assessing its present and future distributions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Being ecosystem engineers, framework-forming scleractinian cold-water corals (CWCs) provide habitat for thousands of deep-sea species, revealing equally remarkable levels of biodiversity as found in tropical coral reefs (Henry and Roberts 2017). Lophelia pertusa is the dominant reef-forming CWC in the Atlantic, and based on its distribution correlated with ocean conditions, upper and lower tolerable limits for basic oceanographic parameters were proposed for this species (e.g., Davies and Guinotte 2011). Among them, dissolved oxygen concentrations (DOconc) can exert control on its biogeographic distribution (e.g., Tittensor et al. 2009). However, lowest DOconc inhabited by this species apparently differs between NE Atlantic (~ 3.7 mL L−1; Dullo et al. 2008) and NW Atlantic (~ 2 mL L−1; e.g., Brooke and Ross 2014) reef sites. These observations are corroborated by laboratory experiments, revealing that L. pertusa individuals collected from DOconc of 6 mL L−1 at the Scottish margin, NE Atlantic, were unable to maintain normal aerobic functions at DOconc < 3.2 mL L−1 (Dodds et al. 2007). Moreover, for L. pertusa specimens collected from DOconc of ~ 2.8 mL L−1 in the Gulf of Mexico, 7-day exposure to DOconc of ~ 1.5 mL L−1 proved fatal (Lunden et al. 2014). Furthermore, discoveries of L. pertusa in the oxygen minimum zones (OMZ) of the subtropical eastern Atlantic (Colman et al. 2005; Le Guilloux et al. 2009) hinted to an even wider tolerance of L. pertusa to low DOconc. Nevertheless, the limited capability of L. pertusa to thrive under DOconc (artificially) reduced below those of their natural environment (Dodds et al. 2007; Lunden et al. 2014), questions its ability to cope with the global change-induced ocean deoxygenation expected for the coming century (e.g., Sweetman et al. 2017).
Although hitherto hypoxic settings were regarded as unsuitable habitats for CWC (e.g., Tittensor et al. 2009), here, we present the discovery of L. pertusa-dominated CWC reefs thriving in the hypoxic OMZ off Angola in the SE Atlantic. The regional adaptation of the Angolan CWC to such extreme conditions sheds new light on their potential capability to cope with expected future environmental changes in the ocean.
Methods
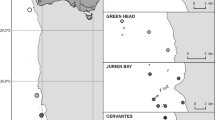
During RV Meteor expedition M122 in January 2016 (Hebbeln et al. 2017), in situ oceanographic parameters such as DOconc and temperature were recorded off Angola (Fig. 1). Data were collected during eight dives with the remotely operated vehicle (ROV) MARUM SQUID (Online Resource 1), three benthic lander deployments (2.5–6.8 days; Online Resource 2) and 17 conventional CTD casts (Online Resource 3). The conventional CTD was additionally equipped with a non-calibrated fluorescence sensor only providing relative values shown as means per water depth averaged from all CTD casts.

Multibeam bathymetry map showing the distribution of cold-water coral reefs off Angola. Locations of CTD casts, benthic lander deployments, and ROV dives are indicated
Results and discussion
Discovery of cold-water coral reefs in the oxygen minimum zone off Angola
ROV video observations revealed the presence of CWC reefs dominated by L. pertusa, which colonize the slopes and summits of up to 100-m-high coral mounds (Fig. 1; Hebbeln et al. 2017). While dispersed CWC colonies were found in a depth range of 250–500 m, large aggregates of healthy colonies were restricted to 330-470 m water depth (Fig. 2). The observation of > 50-cm-high colonies (Fig. 2) clearly evidenced the continuous proliferation of CWC off Angola for many years.

Thriving cold-water corals observed in the oxygen minimum zone (OMZ) off Angola. a, bLophelia pertusa reefs in the center of the OMZ (350 m water depth). c Transported but alive L. pertusa colony in the lower OMZ (500 m depth). dLophelia pertusa colony with many living polyps (439 m depth) (ROV images ©MARUM)
The available oceanographic data revealed water temperatures of 6.8–14.2 °C around the CWC (250–500 m; Fig. 3). The corresponding DOconc of 0.6–1.5 mL L−1 are the lowest ever obtained from waters bathing flourishing L. pertusa colonies (Fig. 3). The ROV–CTD DOconc measurements show the smallest variations. The slightly larger ranges of the lander and conventional CTD data (Fig. 3) likely reflect the impacts of internal waves (Hanz et al. 2019) and the larger geographical coverage, respectively.

Hydrographic setting at the Angolan cold-water coral (CWC) reef site recorded in January 2016. a Water-column temperature (red symbols) and mean relative fluorescence data (green symbols) obtained by conventional CTD and ROV-mounted CTD (temperature only). b Dissolved oxygen concentrations (DOconc) obtained by conventional CTD, ROV-mounted CTD and benthic lander systems (light gray shading indicates entire spread of DOconc data obtained during RV Meteor expedition M122; same in c). c Literature DOconc data spanning from March to September collected between 1995 and 2013 in the same area (Tchipalanga et al. 2018). For all graphs, the depth intervals of either well-developed CWC reefs and/or single living CWC colonies are marked in dark and light yellow, respectively
To gain insight into the seasonal variability of DOconc off Angola, as the M122 data only represent an 8.5-day snapshot from January 2016, we included further 21 CTD casts obtained within the mapped area off Angola (Fig. 1) between 1995 and 2013 (Tchipalanga et al. 2018; Online Resource 4). These data, spanning from March to September, almost completely correspond to the M122 data or reveal even lower DOconc (Fig. 3). Interestingly, even in this hypoxic environment, most prolific CWC reefs are bound to the center of the Angolan OMZ where lowest DOconc prevail (Fig. 3), which coincide with enhanced water-column fluorescence pointing to an increased availability of relatively fresh organic matter (Fig. 3).
Oxygen sensitivity of Lophelia pertusa in the Atlantic Ocean
Based on field observations in the NW and NE Atlantic (Dullo et al. 2008; Freiwald et al. 2009; Brooke and Ross 2014; Georgian et al. 2016), the assumed lower limit of L. pertusa’s oxygen tolerance ranges around DOconc of 2–3.7 mL L−1. This has recently been challenged by very low DOconc of 1.1–1.4 mL L−1 reported from CWC sites off Mauritania (Ramos et al. 2017), which, however, are associated with only sporadic occurrences of small L. pertusa colonies (Wienberg et al. 2018). The new Angolan data documented for the first time L. pertusa’s ability to develop thriving reefs even under DOconc of < 1 mL L−1 (Fig. 3).
In addition, off Angola L. pertusa lives at temperatures of up to 14.2 °C (Fig. 3), which are among the highest temperatures ever observed for this species (13.9–15.2 °C; e.g., Freiwald et al. 2009; Mienis et al. 2014). Thus, off Angola, the partly high temperatures could act as a second stressor since respiration rates of L. pertusa increase with increasing temperature (Dodds et al. 2007).
Stress induced by low DOconc and relatively high temperatures is energetically a challenge for the metabolism of most marine species, but can be compensated by the availability of large quantities of high-quality organic matter (Diaz and Rosenberg 1995). The Angolan and Mauritanian margins belong to highly productive upwelling systems triggering extensive OMZs. Also at many other Atlantic reef sites, L. pertusa is most abundant at depth intervals with highest oxygen depletion (Freiwald 2002; Georgian et al. 2016), most likely linked to highest concentrations of suspended food particles in this layer (e.g., Freiwald 2002) which also applies to Angola (Fig. 3). Comparing ambient DOconc and temperature with site-specific net primary productivity, used as a food supply indicator, for several Atlantic CWC sites, it appears plausible that the negative effects of hypoxia and high temperatures on L. pertusa seemingly could be compensated by significantly enhanced food supply (Fig. 4).

Comparison of environmental parameters (dissolved oxygen concentrations: DOconc; temperature: T; net primary productivity: NPP) obtained for various cold-water coral reef sites in the Atlantic: Gulf of Mexico (GoM; Georgian et al. 2016), Mediterranean Sea (Med; Freiwald et al. 2009), continental margins off Ireland (Dodds et al. 2007; Dullo et al. 2008), Mauritania (Ramos et al. 2017; light yellow: ~ 10 nm east of the CWC reefs highlighting the upwelling impacts), Norway (Dullo et al. 2008), the US east coast (US east; Brooke and Ross 2014) and Angola (this study). Relationship between a DOconc and T and b DOconc and NPP (Behrenfeld and Falkowski 1997)
With respect to L. pertusa preferring regional oxygen minima, ambient DOconc cannot provide any information about its capability to also cope with lower DOconc. However, some information is provided by the aforementioned laboratory experiments. Lophelia pertusa collected in the NE Atlantic and the Gulf of Mexico could not withstand DOconc of less than 40–50% of the ambient values (see above, Dodds et al. 2007; Lunden et al. 2014). Consequently, the range of low DOconc tolerable by L. pertusa—also beyond its natural environment—might depend on the conditions the corals are acclimated to, thus pointing to a possible genotypic adaptive capacity of L. pertusa. Thus, although on a global scale the tolerable DOconc limits for L. pertusa range from < 1 to > 6 mL L−1, smaller ranges define these limits on regional scales.
The future of Lophelia pertusa in a changing ocean
Cold-water coral reefs are vulnerable marine ecosystems that are partly protected within marine protected areas. These can safeguard CWC from destructive human impact (e.g., bottom trawling, hydrocarbon exploration), but offer no sustainable protection against global change-induced threats. In concert with ocean acidification (e.g., Turley et al. 2007) and warming of intermediate waters (e.g., Lunden et al. 2014), also deoxygenation is expected to become a major stressor for CWC (e.g., Sweetman et al. 2017). However, L. pertusa’s general capacity to thrive under well-oxygenated as well as hypoxic bottom waters reveals a rather high oxygen tolerance, although individual L. pertusa populations appear to have limited adaptive capabilities to cope with reductions of 40–50% of ambient DOconc values (Dodds et al. 2007; Lunden et al. 2014). Consequently, the expected decrease in oxygenation of ~ 2% along the Atlantic continental margins until 2100 (Sweetman et al. 2017) by itself might not exert a serious threat to L. pertusa, except for already hypoxic settings like the Angolan margin. However, paleo studies revealed that during the last ~ 20,000 years regional changes in water column structure caused the collapse of L. pertusa-dominated ecosystems due to decreasing DOconc (Wienberg et al. 2018; Tamborrino et al. 2019). Thus, unlike a small overall decrease in DOconc, major regional reductions in DOconc driven by global change-induced changes in ocean circulation have the potential to eradicate regional L. pertusa populations.
Even if smaller decreases in DOconc alone might not pose a serious threat to L. pertusa reefs, these have to be considered in concert with other changing environmental parameters that might form additional stressors (e.g., temperature and pH) with largely unknown consequences for the coral’s biological functions. Moreover, the flux of particulate organic carbon from the surface ocean might decline by ~ 30% until 2100 along the Atlantic margins (Sweetman et al. 2017), resulting in a lower food supply to the CWC and deep-sea organisms in general, thus reducing their capacity to cope with increasing stress.
References
Behrenfeld MJ, Falkowski PG (1997) Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol Oceanogr 42:1–20
Brooke S, Ross SW (2014) First observations of the cold-water coral Lophelia pertusa in tmid-Atlantic canyons of the USA. Deep Sea Res Part 2 Top Stud Oceanogr 104:245–251
Colman JG, Gordon DM, Lane AP, Forde MJ, Fitzpatrick J (2005) Carbonate mounds off Mauritania, Northwest Africa: status of deep-water corals and implications for management of fishing and oil exploration activities. In: Freiwald A, Roberts JM (eds) Cold-water corals and ecosystems. Springer, Heidelberg, pp 417–441
Davies AJ, Guinotte JM (2011) Global habitat suitability for framework-forming cold-water corals. Plos ONE 6:e18483
Diaz RJ, Rosenberg R (1995) Marine benthic hypoxia: a review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanogr Mar Biol Ann Rev 33:245–303
Dodds LA, Roberts JM, Taylor AC, Marubini F (2007) Metabolic tolerance of the cold-water coral Lophelia pertusa (Scleractinia) to temperature and dissolved oxygen change. J Exp Mar Bio Ecol 349:205–214
Dullo W-C, Flögel S, Rüggeberg A (2008) Cold-water coral growth in relation to the hydrography of the Celtic and Nordic European continental margin. Mar Ecol Prog Ser 371:165–176
Freiwald A (2002) Reef-forming cold-water corals. In: Wefer G, Billett D, Hebbeln D, Jorgensen BB, Schlüter M, van Weering TCE (eds) Ocean margin systems. Springer, Berlin, Heidelberg, pp 365–385
Freiwald A, Beuck L, Rüggeberg A, Taviani M, Hebbeln D, R/V Meteor Cruise M70-1 participants (2009) The white coral community in the central Mediterranean Sea revealed by ROV surveys. Oceanography 22:58–74
Georgian SE, DeLeo D, Durkin A, Gomez CE, Kurman M, Lunden JJ, Cordes EE (2016) Oceanographic patterns and carbonate chemistry in the vicinity of cold-water coral reefs in the Gulf of Mexico: Implications for resilience in a changing ocean. Limnol Oceanogr 61:648–665
Hanz U, Wienberg C, Hebbeln D, Duineveld G, Lavaleye M, Juva K, Dullo WC, Freiwald A, Tamborrino L, Reichart GJ, Flögel S, Mienis F (2019) Environmental factors influencing cold-water coral ecosystems in the oxygen minimum zones on the Angolan and Namibian margins. Biogeosciences 16:4337–4356
Hebbeln D, and cruise participants (2017) ANNA Cold-Water Coral Ecosystems off Angola and Namibia - Cruise No. M122 - December 30, 2015 - January 31, 2016 - Walvis Bay (Namibia) - Walvis Bay (Namibia). METEOR-Berichte, M122, DFG-SenatskommissionfürOzeanographie 74 [https://doi.org/10.2312/cr_m122]
Henry L-A, Roberts JM (2017) Global biodiversity in cold-water coral reef ecosystems. In: Rossi S, Bramanti L, Gori A, Orejas Saco del Valle C (eds) Marine animal forests The ecology of benthic biodiversity hotspots. Springer, Cham, pp 235–256
Le Guilloux E, Olu-Le Roy K, Bourillet JF, Savoye B, Iglésias SP, Sibuet M (2009) First observations of deep-sea coral reefs along the Angola margin. Deep Sea Res Part 2 Top Stud Oceanogr 56:2394–2403
Lunden JJ, McNicholl CG, Sears CR, Morrison CL, Cordes EE (2014) Acute survivorship of the deep-sea coral Lophelia pertusa from the Gulf of Mexico under acidification, warming, and deoxygenation. Front Mar Sci. https://doi.org/10.3389/fmars.2014.00078
Mienis F, Duineveld GCA, Davies AJ, Lavaleye MMS, Ross SW, Seim H, Bane J, van Haren H, Bergman MJN, de Haas H, Brooke S, van Weering TCE (2014) Cold-water coral growth under extreme environmental conditions, the Cape Lookout area, NW Atlantic. Biogeosciences 11:2543–2560
Ramos A, Sanz JL, Ramil F, Agudo LM, Presas-Navarro C (2017) The giant cold-water coral mounds barrier off Mauritania. In: Ramos A, Ramil F, Sanz JL (eds) Deep-sea ecosystems off Mauritania: Research of marine biodiversity and habitats in the northwest African margin. Springer, Netherlands, Dordrecht, pp 481–525
Sweetman AK, Thurber AR, Smith CR, Levin LA, Mora C, Wei C-L, Gooday AJ, Jones DOB, Rex M, Yasuhara M, Ingels J, Ruhl HA, Frieder CA, Danovaro R, Würzberg L, Baco A, Grupe BM, Pasulka A, Meyer KS, Dunlop KM, Henry L-A, Roberts JM (2017) Major impacts of climate change on deep-sea benthic ecosystems. Elementa-Sci Anthrop 5:1–23
Tamborrino L, Wienberg C, Titschack J, Wintersteller P, Mienis F, Schröder-Ritzrau A, Freiwald A, Orejas C, Dullo WC, Haberkern J, Hebbeln D (2019) Mid-Holocene extinction of cold-water corals on the Namibian shelf steered by the Benguela oxygen minimum zone. Geology 47:1185–1188
Tchipalanga P, Dengler M, Brandt P, Kopte R, Macuéria M, Coelho P, Ostrowski M, Keenlyside NS (2018) Eastern boundary circulation and hydrography off Angola: building Angolan oceanographic capacities. Bull Am Meteorol Soc 99:1589–1605
Tittensor DP, Baco AR, Brewin PE, Clark MR, Consalvey M, Hall-Spencer J, Rowden AA, Schlacher T, Stocks KI, Rogers AD (2009) Predicting global habitat suitability for stony corals on seamounts. J Biogeogr 36:1111–1128
Turley CM, Roberts JM, Guinotte JM (2007) Corals in deep-water: will the unseen hand of ocean acidification destroy cold-water ecosystems? Coral Reefs 26:445–448
Wienberg C, Titschack J, Freiwald A, Frank N, Lundälv T, Taviani M, Beuck L, Schröder-Ritzrau A, Krengel T, Hebbeln D (2018) The giant Mauritanian cold-water coral mound province: Oxygen control on coral mound formation. Quat Sci Rev 185:135–152
Acknowledgements
Open Access funding provided by Projekt DEAL. We like to thank the nautical and scientific crews for on-board assistance during RV Meteor cruise M122. This research received support from the Deutsche Forschungsgemeinschaft (DFG) through providing ship time and access to the ROV and through the DFG Research Center/Cluster of Excellence “MARUM—The Ocean in the Earth System.” C.O. received a scholarship by the German Academic Exchange Service (DAAD, Grant No. 91723955) supporting her research stay in Bremen, Germany.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Topic Editor Mark R Patterson
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hebbeln, D., Wienberg, C., Dullo, WC. et al. Cold-water coral reefs thriving under hypoxia. Coral Reefs 39, 853–859 (2020). https://doi.org/10.1007/s00338-020-01934-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-020-01934-6