
Abstract
Sediments from a small lake, Loch Beag, in Barrees, Beara peninsula, south-western Ireland, provide a record of vegetation history that spans most of the Lateglacial and Holocene (14–0.43 ka). The detailed part of the record relates to the Holocene, which is the focus of this paper. The investigations carried out include pollen and macrofossil analyses, loss-on-ignition measurements and 14C dating. As well as providing records for tree and shrub taxa, and especially Pinus, Quercus, Ulmus, Fraxinus and Taxus, other taxa mainly with Atlantic and/or Lusitanian distributions are also considered. These include Arbutus unedo (strawberry tree) and fern species such as Osmunda regalis, and the filmy ferns Hymenophyllum tunbrigense, H. wilsonii and Trichomanes speciosum (Killarney fern). Detailed records for a suite of bog/heath taxa, including ericoid species (Erica tetralix and E. cinerea), Narthecium ossifragum, Hypericum elodes and Myrica gale, are presented. The results of 14C dating of bog-pines from western Ireland are summarised. Results of other investigations, including key Holocene pollen diagrams, and charcoal and fossil-wood records from archaeological excavations that relate mainly to the Bronze Age in the wider west Cork/Kerry region, are reviewed. Vegetation dynamics, human impact and plant biogeography are discussed in the light of the considerable data now available for the region.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
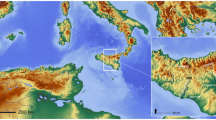
South-western Ireland, consisting of a series of peninsulas, is a distinctive region as regards climate, geomorphology, floristics and vegetation (Fig. 1). There is also substantial evidence for a human presence starting in the Mesolithic and particularly from the Neolithic/Bronze Age transition onwards (O’Brien 2009; Aalen et al. 2011). The region has a pronounced oceanic climate with mild winters, warm summers, infrequent frosts, and high precipitation distributed evenly throughout the year (see Study area). The geology is dominated by Devonian sandstone, often referred to as ‘Old Red Sandstone’ (Pracht 1997; Pracht and Sleeman 2002). There is a considerable exposure of Carboniferous limestone in the Killarney area including the Muckross peninsula, where the karstic limestone supports a Taxus (yew) woodland of some 25 ha (Kelly 1981). The upland character and north-east/south-west grain of the landscape was created by the Variscan (Armorican) orogeny towards the end of the Carboniferous period. This folding resulted in the creation of a metalliferous zone of copper, manganese and barites, that became the basis for Bronze Age and later copper mining in west Cork and Kerry (Kinnaird and Nex 2004). During the Weichselian, i.e. the last Ice Age (Midlandian in Ireland), the whole area was subject to severe glaciation that shaped and scoured the uplands, and resulted in considerable glacial deposits especially in the Killarney and Bantry areas (Ballantyne et al. 2011). The region is rich in periglacial features, and in the Beara peninsula solifluction lobes and involutions, of possible Younger Dryas age, are recognized (Coxon 1997; Harrison et al. 2010).

A Map of Ireland. County boundaries and the study region shown in B are indicated. B The study region, i.e. part of Counties Kerry and Cork. Locations of the main pollen diagrams (including profiles by Vokes 1966; V1: Muckross Bog; V2: Ladies View) discussed in the text and selected prehistoric copper mines (Ross Island, Crumpane and Mount Gabriel; also Arderrawinny and Derrycarhoon) are indicated. Various geographical features, including roads, lakes and rivers, are shown. C Aerial photograph (detail of the Beara peninsula centred on Barrees; source: Google Earth, accessed November 2023) showing location of the pollen profiles, and archaeological sites for which charcoal data are available (sites are referred to by the letter given in parentheses). FF fulacht fiadh (burnt mound)
As regards vegetation, the region is renowned for its Atlantic, temperate rainforests, including the yew woodlands at Muckross (see above) and especially the Quercus petraea-dominated woodlands that find best expression on sandstone in Killarney National Park, Ireland’s first and premier National Park (Kelly 1981; Cross 2006; Fig. 1). Pockets of fern and bryophyte-rich oak woodlands exist, such as at Glengarriff, and also in other parts of the Beara peninsula including that recently created as a result of the rewilding project by conservationist, Eoghan Daltun (2022). Bog, and especially blanket bog, forms extensive tracts throughout the region. Indeed, blanket bog and heath dominate most of the uplands.
As regards floristics, the region is noted for its eu-Atlantic and Lusitanian elements (Webb 1983). Particularly noteworthy are the filmy ferns, Hymenophyllum tunbrigense, H. wilsonii and Trichomanes speciosum, i.e. the so-called Killarney fern (Richards and Evans 1972; Ratcliffe et al. 1993). Lusitanian species include Arbutus unedo (strawberry tree) which occurs in many of the oak (Quercus) and yew (Taxus) woodlands (e.g. Killarney, Glengarriff and elsewhere; Sheehy Skeffington and Scott 2021; Scott and Sheehy Skeffington 2022), Euphorbia hiberna, Saxifraga spathularis and Pinguicula grandiflora which are common, and the rare Simethis planifolia (S. mattiazzii) which, in Ireland and Britain, is known only from the Iveragh and Beara peninsulas (Lupton and Sheehy Skeffington 2020; Stroh et al. 2023). A recent discovery is the tropical fern, Stenogrammitis myosuroides (Sw.) Labiak, in Killarney National Park, the spores of which have probably been wind-transported from the Carribean (Hodd and Rumsey 2020).
The main focus of this paper is the Beara peninsula which straddles the county boundaries of Cork and Kerry (Fig. 1). Nearby areas that share many of the same archaeological, geographical and vegetational features are also considered, and especially Iveragh peninsula to the north which includes the Killarney National Park, and Mizen peninsula to the south, a mainly upland area noted for its Bronze Age copper mines including those associated with Mount Gabriel (MG) (O’Brien 1994, 1999, 2003, 2004, 2009, 2015, 2022).
The Beara peninsula is some 60 km long with a backbone consisting of uplands that seldom exceed 650 m altitude (a.s.l.). The peninsula is noted for its rich prehistoric archaeology and especially that relating to the Bronze Age which includes megaliths (wedge tombs, stone circles, stone rows and single standing stones), fulachta fiadh (burnt mounds) and copper mines (four MG-type mines). Ancient stonewalls, many of which are pre-bog and may have served as field and/or enclosure boundaries, are frequent. Details of some 200 such sites are provided by Murphy (2009) for Beara while O’Brien (2009, p 50) reports 77 locations for ancient field walls. These are frequently overlain by peat up to a metre thick and so, presumably, are of considerable age.
Barrees townland is situated between the villages of Ardgroom and Eyeries (Fig. 1). The part that formed the main study area (referred to here as Barrees) is low-lying (it extends from about 100 to 200 m a.s.l.) and is relatively sheltered from the prevailing south-westerlies by a low ridge. Today it supports pastoral farming on improved agricultural land, and rough grazing on heath and blanket bog at higher elevation. Small patches of Atlantic oak woodland survive at low elevation, while at higher elevation but outside the main study area there has been considerable coniferous planting in recent decades.
Barrees is particularly rich in archaeological evidence for past human activity that is now summarised (for a detailed account see O’Brien 2009). Several stretches of pre-bog stonewalls are present. These define fields (36 have been identified) that are mostly small, i.e. < 0.1–0.7 ha, with the largest extending to 2.8 ha. These appear to have been laid out and modified at various times in later prehistory and the early historical period. There is also substantial evidence for human activity that relates specifically to prehistory and especially the Bronze Age. This includes standing stones, fulachta fiadh (burnt mounds) and a small copper (Cu) mine (Crumpane; MG-type mine) in the uplands. Archaeological excavations have shown that the Iron Age is also well represented; e.g. Site A (Fig. 1C), which is a mid- to late Iron Age enclosure (ca. 1st millennium ce) the use of which extended into early Medieval times. Glass beads of various colours recovered during excavation serve to strengthen the dating and indicate distant connections with other Iron Age communities (O’Brien 2009, pp 248–249). These and other finds are of particular significance given the paucity of evidence for Iron Age activity in Ireland generally and specifically in Beara (Comber 2009).
Bearing in mind the rich archaeology, Barrees was an obvious choice for detailed archaeological survey and excavation (O’Brien 2009), while the peat deposits and organic-rich soils provided ample opportunity for detailed palaeoecological investigations to be carried out at, or in close association with, the excavated sites (short pollen profiles BAR-1 to BAR-5). A small enclosed, peat-filled basin within the main area of archaeological interest yielded a pollen profile of local character that spanned the last ca. 4,500 years, i.e. from the late Neolithic/Bronze Age transition onwards (profile BAR-L1). The results of these investigations are discussed in a contribution by Overland and O’Connell (2009) to the O’Brien (2009) monograph and in a more detailed account elsewhere (Overland and O’Connell 2008).
The investigations reported on here relate mainly to a small lake, Loch Beag, in a relatively sheltered, north-west-facing depression within the study area (Fig. 1). The lake is unnamed in the Ordnance Survey of Ireland (OSI) maps so it has been given the name Loch Beag which in Gaelic means ‘small lake’. The resulting pollen profile, BEG1, spans most of the Holocene and includes a Late-glacial sequence at the base. A preliminary description of this profile with focus on the upper part of profile, i.e. the last 4,500 years, is available in Overland and O’Connell (2008, 2009). Here a comprehensive account of the investigations is provided and the implications of the results for long-term vegetation and landscape developments, and the effects of human impact and other factors including climate change is discussed, as well as the implications of the new findings for plant biogeography and particularly for the Lusitanian elements of the flora. Though the lake basin at Loch Beag is small (see Study area), the pollen profile is more regional in character than BAR-L1 and, given its detail, provides an important long-term Holocene record for the Beara peninsula and south-west Ireland generally. In the Discussion, results from other palaeoecological investigations in the wider region are compiled, evaluated and discussed towards providing an improved regional picture of woodland and land-use history, and long-term environmental change in the Kerry/West Cork region of south-western Ireland.
Study area
The lake, Loch Beag, lies in a closed basin on Devonian sandstone (Fig. 1, ESM 1 Fig. S1; 103 m a.s.l.). When cored (23/10/2004), the open-water body extended to about 50 × 30 m; prior to recent rapid infilling the lake size was about 100 × 80 m. The catchment size is small and consists mainly of elevated ground and, to a less extent, peat-covered flat ground to the south-east of the lake. The lake has no obvious natural out-flowing or in-flowing streams. Common bog plants about the lake include Erica tetralix, Myrica gale, Narthecium ossifragum and Rhynchospora alba. Mosses include Hylocomium splendens, Sphagnum cuspidatum, S. papillosum and S. subnitens. In rough and partly reclaimed pastures, rushes are common including Juncus effusus, J. acutiflorus, J. articulatus and J. squarrosus. Fragments of Atlantic oak woodland with abundant holly (Ilex aquifolium) and ivy (Hedera helix), are present in the lowlands (up to 100 m a.s.l.) to the north of the lake and include typical moss and fern species. Here H. tunbrigense is frequent on shaded boulders together with eu-Atlantic leafy liverworts such as Saccogyna viticulosa and Scapania spp. H. wilsonii was not recorded in the woodland but fertile specimens were noted on north-facing boulders in upland (up to 150 m a.s.l.), heathy parts. Plant nomenclature follows Parnell and Curtis (2012) for vascular plants and Atherton et al. (2010) for bryophytes.
The climate is characterised by high but seasonally evenly distributed rainfall (1,143 mm y−1; 249 and 193 days with precipitation ≥ 0.2 mm and ≥ 1 mm, respectively), and mild winters (January: 8.2 and 3.1 °C) and rather cool summers (August: 17.2 and 11.1 °C) (mean daily maximum and minimum monthly temperatures, respectively, are given). Frost and snow are infrequent (mean number of days with snow lying on the ground at 9:00 h UTC: 2.3). Winds are predominantly south-westerly and wind speeds are high (mean annual wind speed 6.7 m sec−1; mean number of days with gales, i.e. wind speeds > 17 m sec−1: 30.5). The statistics relate to Valencia Observatory, i.e. the nearest synoptic weather station 35 km to the north-west, and refer to the period 1961–1990 (https://data.gov.ie/dataset/met-eireann-1961-1990-climate-averages; accessed 13 October 2023).
Material and methods
Fieldwork
Trial cores were taken at four points that were relatively easily accessible on the lake margin with a gouge (Dutch) corer, with a view to finding the deepest location and most suitable sediment. Late-glacial sediments were found between 7 and 9 m depth in these trial cores. The location with thickest sediments was chosen for coring. This lay at the south-eastern side, 4 m distant from open water. Two parallel cores, BEG1 and BEG2 (51.71235N, 9.92578W) were retrieved from beneath a floating mat of vegetation using an 80 mm diameter, Usinger piston corer (Fig. 1, ESM 1 Fig. S1). The cores were taken 50 cm apart in 1 m long drives, the drives starting 20 cm higher in the case of BEG2. As sediment from BEG1 has mainly been used for detailed analyses, the pollen profile is here referred to as BEG1. The uppermost 1.8 m of deposit, which was fibrous and so difficult to collect, was not retrieved. The cores were extruded on site, cut in two longitudinally onto 1 m long halved plastic pipe, and wrapped in a strong plastic sleeve. The cores were stored horizontally in a cool location so as to maintain, as far as possible, their original condition.
Laboratory methods
The cores were photographed a few days after being taken (ESM 1 Fig. S1) and stratigraphic features were described following the Troels-Smith (1955) system (updated in Aaby and Berglund 1986). Magnetic susceptibility (κ) was measured using ‘split-core logging’ as described by Nowaczyk (2001) using a Bartington MS2E spot-reading sensor (details in Overland 2007). Cross correlation of the parallel cores was carried out mainly on the basis of stratigraphy and to some degree the magnetic susceptibility results. Over critical intervals, i.e. where substantial changes in the pollen curves were recorded at/close to core segment ends, correlation between the two cores was confirmed by carrying out additional analyses on the duplicate core.
Pollen and macrofossil samples of 1 cm3 were taken at mainly 4 cm intervals and, in places of particular interest, at closer intervals. For pollen analysis, standard procedures as implemented at the Palaeoenvironmental Research Unit (PRU), University of Galway were followed (Overland and O’Connell 2008). This included initial sieving using a 100-µm mesh sieve, treatments with KOH, HCl and HF followed by acetolysis, and sieving the final pellet using a 5 µm mesh sieve in an ultra-sonicator. A calibrated suspension of Lycopodium spores (exotic) was added to each sample at the beginning of the preparation procedure to facilitate estimation of pollen concentration. The pollen samples were mounted in glycerol, and counted under phase contrast using a Leica DM LB2 microscope fitted with × 12.5 oculars and a phase contrast Planapo 63/1.4 objective. Approximately 1,000 pollen were counted per sample. Pollen and spore identification followed mainly Faegri and Iversen (1989). Other authorities consulted included Moore et al. (1991), Beug (2004) and Reille (1992, 1995). For all critical determinations, the modern reference collections in the PRU were consulted. Selected non-pollen palynomorphs (NPPs), including fungal spores, epidermal fragments of E. tetralix seed, and microscopic charcoal (≤ 37 µm) were also counted. NPPs were identified using Van Geel (1978) and other publications. Pollen and spore nomenclature mainly follows Moore et al. (1991). Type numbers preceded by ‘HdV-’, in addition to the names that are in common use, are shown for NPPs in the pollen diagrams (cf. Shumilovskikh et al. 2022, 2023). Pinus sylvestris (Scots pine) stomata were identified by reference to modern material and the descriptions by Trautmann (1953).
For calculation of percentage pollen diagrams a total terrestrial pollen sum (TTP) that included bog taxa was used. Taxa excluded from the pollen sum include Sphagnum spores, aquatic taxa, corroded/unknown grains (generally low), charcoal and NPPs. The percentage representation of these taxa was calculated relative to TTP and the sum of taxa pertaining to the component in question. Pollen concentration and accumulation (referred to here as pollen influx) rates were calculated in the usual way; in calculating pollen influx rates, sediment accumulation rates derived from the age/depth curve were used.
Zonation of the pollen diagram was carried out by careful inspection of the percentage pollen curves. Superzones were recognised in the Holocene sequence, which in turn were subdivided to reflect less pronounced changes in one or more curves, the changes in question having been deemed to be of significance in terms of vegetation and/or land-use history.
The material retained in the 100-µm mesh sieves, after KOH treatment during pollen preparation, was scanned for macrofossils and other entities using a Wild M8 binocular stereoscope that gave magnifications ranging from × 9 to × 75. Quantitative estimates of macrofossils, macro-charcoal and mineral matter were recorded using the scale ‘rare’ ( +), ‘occasional’ (1), ‘frequent’ (2) and ‘abundant’ (3). ‘ + ’ values were subsequently converted to 0.2% to facilitate plotting. Identification of Juncus seeds followed Körber-Grohne (1964), whereas other seeds were identified using Schoch et al. (1988) and Beijerinck (1947). Smith (1978) and Watson (1981) were used for moss identification.
Ash dry-weight as a percentage of sample dry-weight was determined for levels corresponding to where sampling was carried out for pollen analysis. Samples of up to 10 g were dried for 24 h at 90 °C and burned for six h at 550 °C.
Terrestrial macrofossils for accelerator mass spectrometry (AMS) 14C dating were obtained by sieving lake sediments (sample thickness mainly 2 cm; sample volume up to 35 cm3) through a 125-µm sieve and scanning for macrofossils using a Leica MZ125 stereo-microscope. A bulk sample of lake sediments, relating to the early Holocene, was also AMS 14C dated. 14C dates were calibrated using OxCal v. 4.4.4 (Bronk Ramsey 2009; Reimer et al. 2020) and an age-depth curve was constructed using Clam v. 2.3.8 (Blaauw 2010).
Results and interpretation
Sediment stratigraphy, magnetic susceptibility, ashing
Lithological features described here are based mainly on core BEG1, i.e. the core used for most of the detailed sampling (stratigraphy/lithology is shown in Figs. 2, 3 and also in the pollen plots; for photographs of the cores see ESM 1 Fig. S1; stratigraphical details are in ESM 1 Tables S1a–c; core correlation is depicted schematically in ESM 1 Fig. S3). To facilitate the descriptions of lithology, pollen and other data, reference is occasionally made to the classical pollen zones as recognised in Ireland (Lateglacial after Walker et al. 1994; Holocene after Mitchell 1956) and also to cultural/historical periods in Ireland (Waddell 2022 for prehistory generally; O’Brien 2012 and Roberts et al. 2013 for the Chalcolithic and Bronze Age, respectively; Aalen et al. 2011 for historical times).

A Ash as a percentage of dry weight. B Magnetic susceptibility values. For ash and magnetic susceptibility, individual values and weighted averages (WA; weightings are given in parentheses) are shown using thin and thick lines, respectively. Other details shown include the stratigraphy, the PAZs and the ages for the PAZ boundaries (rounded to the nearest 10 y). C Sediment accumulation rates and times derived from the age/depth model as in Fig. 3. Numbered stars are used to highlight elevated accumulation rates (see text). Keys to the lithology are in Figs. 4, 5

Age/depth curve (smooth spline curve; smoothing = 0.3) for profile BEG1 generated by Clam. Details of 14C dates, and lithology and pollen zonation are also shown
The basal sediments (870–900 cm from the surface) consist of highly minerogenic sediments that were deposited presumably towards the end of the Pleniglacial. After a short transition, gyttja, i.e. an organic-rich lake sediment sometimes with conspicuous silt, is present between 836.5 and 868 cm; this is followed by a 10 cm thick silty/clay sediment. These two units can be ascribed, on the basis of lithology and pollen evidence, to the Bølling/Allerød and Younger Dryas periods, respectively. The transition to more organic-rich sediment (corresponding to the Lateglacial/Holocene transition) occurs over a short interval (826–828.5 cm), above which the sediment consists of typical gyttja that was generally dark, lake-derived and organic-rich. Starting at about 309 cm, the sediment became more fibrous, and pronouncedly so from at/above 261.5 cm. The increased fibrosity is probably the result of infilling that led to terrestrialisation and the local development of a floating mat of vegetation at the lake’s margin.
Highest ash values relate to the minerogenic Younger Dryas sediments (98%; Fig. 2A); the basal Pleniglacial sediments, also highly minerogenic, were not ashed. Between 808 and 372 cm, the values are generally in the range 20–60%. There are distinctly low ash values at or about 772 cm (9.3 ka), 732 cm (8.3 ka) and 710 cm (7.85 ka). Between 628 and 604 cm (5.7–5.4 ka) and 516–500 cm (3.9–3.76 ka), ash values remain low. At 372 cm (1.3 ka), the ash value declines sharply to about 6%, and values continue to be low until 276 cm (0.81 ka) after which point values increase and a generally increasing trend is then maintained to the top of the core.
As expected given the highly minerogenic sediments, the Younger Dryas and Pleniglacial sediments have exceptionally high κ values (Fig. 2B). In much of the core, however, κ values are inversely related to the ash values, which is not as expected. This suggests a complex relationship between soil erosion in the catchment and the magnetic and other properties of the sediment.
Chronology and sedimentation patterns
The results of 14C dating and data relating to construction of the age/depth curve are given in ESM 1 Table S2 and are shown in Fig. 3. Four 14C dates relating to the upper part of the core were regarded as unrealistic. The uppermost two of these dates are too old, due probably to inwash of organics (peat) as a result of increased land-use in the catchment (14C dates affected by ‘old’ carbon are frequently encountered in the upper parts of cores from Irish and British lakes; Edwards and Whittington 2001). Accepting the two lowermost dates would require major changes in sedimentation rates for which there is little supporting evidence. In the case of the 14C assay from near the base of the core—the only bulk-sample 14C dated—dates for alkali-soluble and non-soluble fractions were returned by the 14C laboratory. The difference in age is small (< 100 14C y) and so both dates have been used within CLAM for construction of the age/depth curve. To better constrain the upper and lower parts of the age/depth curve, the top of the core (1 cm) is assigned the date 2000 ce and the lower parts are constrained by the generally accepted dates for the beginning and end of the Younger Dryas and the end of the Pleniglacial (Lowe et al. 2008; Walker et al. 2019; Fig. 3).
The most acceptable age/depth model is considered to be that represented by a smooth spline curve (smoothing = 0.2) (Fig. 3). The main trends in accumulation rates are as follows (Fig. 2C). Until ca. 10 ka (8 m), rates are slow (0.14 to 0.24 mm y−1); after ca. 8.8 ka (7.5 m) rates of > 0.4 mm y−1 are attained (1 in Fig. 2C) but decline to about 0.3 mm y−1 between ca. 7 and 5.8 ka. Elevated accumulation rates are again recorded at ca. 5.3 and 3.75 ka (2 and 3 in Fig. 2C). The upper part of the sequence is characterised by high accumulation rates (1 mm y−1) that increase to 4 mm y−1, i.e. > 3 times the average rate. Note that the patterns as described are greatly influenced by points for which 14C dates are available and so, like the age/depth model itself, are in reality best approximations.
Pollen, macrofossil and other data
Percentage pollen data for the lower part of the sequence, i.e. the Lateglacial and early Holocene, are plotted in Fig. 4 (this plot includes spectrum 808 cm which is also included in Fig. 5). Pollen curves for the mid- and top parts of the profile (most of the Holocene) are shown in Fig. 5. Macrofossils and other entities for the whole sequence are plotted in Fig. 6. Percentage pollen records for some minor taxa are not plotted in the interests of legibility of the plots. Summary percentage, concentration and influx (pollen accumulation rate) data are shown in ESM 1 Fig. S4. All data will be made available in PANGAEA. The dates cited below and elsewhere are derived from the age/depth curve (Fig. 3). In the descriptions that follow, all references are to the percentage curves unless otherwise indicated.

Percentage pollen profile BEG1 plotted to a depth scale. The lowermost spectra, i.e. Lateglacial and early Holocene, and ash values are shown. A TTP pollen sum is used. Dashed curves indicate × 10 exaggeration



Pollen percentage profile BEG1 plotted to a depth scale (in three parts, A–C). The conventions followed are similar to those in Fig. 4, where the lowermost spectra, i.e. spectra relating to the Lateglacial and early Holocene, are plotted. Two x-axis scales are used, i.e. a standard scale and, in the case of taxa with exceptionally low values, the standard scale is exaggerated × 2. NAP1 and NAP2 refer to NAP indicative of grassland and disturbed/arable habitats, respectively. Cultural/historical periods are indicated in B

Macro-remains and other entities recorded in sievings of pollen and 14C samples, profile BEG1 plotted to a depth scale. The x-axes are abundance scales (see text). Plant parts are as follows: ca caryopsis (Poaceae), lf leaf, fr fruit, o oospores (Characeae), s seed, ut utricle
Lateglacial and earliest Holocene (zones BA, YD and A1; 860–812 cm; 14–10.9 ka; Fig. 4). Slow accumulation rate at the base of the sequence results in the Lateglacial being represented in ∼30 cm of sediment. This, and the small number of samples analysed (10), result in a record that provides only an outline of vegetation change during this dynamic period. Subzone BA1 is regarded as equivalent to the Bølling and is here characterised by rather high Juniperus and Rumex-type values. The very high Juniperus values that characterise the early Bølling in many Irish Late-glacial sequences (e.g. Craig 1978; Mitchell and Cooney 2019) are not recorded possibly because of slow sedimentation and low-resolution sampling. The available record suggests open shrubby vegetation in which juniper is the main shrub. Empetrum nigrum is also present and probably also Betula nana. Separation of dwarf-shrub from tree birch pollen was not attempted; it is assumed that the Betula pollen derives mainly from the former as tree birch (most likely B. pubescens) is rare in the Irish Lateglacial (Mitchell 1981; Andrieu et al. 1993). Rumex-type is the main NAP taxon; Oxyria digyna was probably an important contributor to this pollen taxon as well as various Rumex species.
In subzone BA2, which is regarded as equivalent to the Allerød, Poaceae is the main pollen curve and Juniperus is well represented but it has decreased. This, and the high diversity of NAP taxa, indicate a shift towards herb-dominated communities at the expense of juniper. High Pediastrum values (also high Tetraëdron at the base of the zone) suggest increased inwash as a result of higher solifluction rates. These changes are indicative of a decline in temperature for which there is evidence from various proxies at several sites in Britain and Ireland (Walker et al. 1993; Watson et al. 2010). Elevated Juniperus, a minor peak in Filipendula and a decrease in ash content (less solifluction) in the uppermost spectrum suggest a minor but perceptible climatic upturn towards the end of the zone (cf. O’Connell et al. 1999).
Zone YD, regarded as representing the Younger Dryas, has four pollen spectra from the most mineral-rich part of the sequence (Fig. 4). Fern spores, i.e. mainly spores without perine, and Polypodium, are well represented. This may be due to differential preservation of these decay-resistant spores. Juniperus values are so low that it can be concluded that juniper is no longer locally present or, if it is, its pollen production is exceptionally low. These changes, as well as low pollen concentration and influx values (ESM 1 Fig. S4), suggest substantially lower productivity as a result of a drop in temperature of several degrees compared with the preceding Allerød (O’Connell et al. 1999; Watson et al. 2010; Van Asch et al. 2012).
The four spectra that constitute zone A1 record the post-glacial expansion firstly of Juniperus, then Betula (presumably tree birch), and finally the expansion of Corylus (hazel). Herbaceous communities were replaced by woody vegetation (initially juniper, then birch and finally hazel). Willows were also important as is indicated by Salix values > 10%. These developments, and the progressive and the more or less complete suppression of the pioneering tall shrubby plants and especially juniper, reflect changes that were facilitated by a rapid though probably non-linear rise in temperature (cf. Molloy and O’Connell 2014; Krüger et al. 2020) and soil development at the beginning of the Holocene.
Low values for Pinus and Quercus (also Ulmus) are probably best attributed to long-distance transport. Populus and Viburnum pollen (mostly V. opulus; the pollen were not distinguished to species level in all instances) are recorded which is noteworthy especially given that there are relatively few such records available for Ireland (Watts 1984; Kollmann and Grubb 2002; Molloy and O’Connell 2014).
Early to mid-Holocene (zones A2–D2; 808–584 cm; 10.9–5 ka; Figs. 5, 6). During this long interval, AP generally constitutes 70–90% of TTP from which it can be inferred that woodland was dominant. Changes in the contribution of individual pollen taxa, not only AP but also NAP and other microfossils, and also macrofossils, enable fine-scale subdivisions to be made as follows.
Zone A2 (808–772 cm; 10.9–9.26 ka, 8950–7305 bce). Corylus values are high (∼50%), Quercus increases sharply towards the top of the zone to achieve 25%, and Ulmus and Pinus show only minor increases and fail to achieve 2%. Hazel was dominant throughout this period, but towards the end oak was making a substantial contribution. Both Quercus petraea and Q. robur may have been present but it is more likely the former on the basis of the greater tolerance of Q. petraea to non-calcareous soils and its present-day, distinct western distribution in Ireland (Stroh et al. 2023; updated, online version at uhttps://plantatlas2020.org/atlas, accessed 1 September 2023). If elm (presumably Ulmus glabra) were present, then it was of minor importance. In view of the high pollen production and dispersal of pine, the Pinus pollen records (probably P. sylvestris) are best attributed to long-distance transport. Populus persists as does Viburnum, Sorbus is well represented (peak: 2.2%) which is regarded as indicative of local presence (probably S. aucuparia; Sorbus spp. generally are severely under-represented in pollen records; Boyd and Dickson 1987), and Hedera is consistently recorded in the upper part of the zone, starting at 9.9 ka. Osmunda (royal fern) is recorded in zone A1 (also occasional earlier records but given the strong solifluction during at least part of the Lateglacial, reworked spores cannot be ruled out), and achieves high values (max. 7.4%) at the base of zone A2 which suggests local presence. A more or less continuous record for Pteridium also commences in zone A2. NAP (mainly Poaceae and Rumex-type) decline to low values which lends weight to the idea of the development of a dominant, closed-canopy woodland.
Zone B1 (768–740 cm; 9.3–8.4 ka, 7305–6465 bce). Within this zone, contributions by the various AP taxa change substantially. Pinus increases initially rather slowly and then rapidly to achieve 31%. A consistent Pinus stomatal record begins at 756 cm (8.84 ka) which signals local presence of Scots pine (P. sylvestris). As Pinus increases, Corylus declines and Quercus also decreases but to a lesser extent. These changes show that pine has become the dominant tree, a role it maintains for at least four millennia (until D2/E1 transition at ca. 5 ka). Betula values are not particularly high (∼12%) but macrofossil evidence suggests local presence of tree birch (B. pubescens) during this and succeeding zones until at least zone E1 (ca. 4.5 ka). The ferns, Polypodium and Pteridium, show small increases (also Poaceae) which suggest rather open woodland structure. Changes in local wetland vegetation are indicated by a decline in Osmunda and several E. tetralix pollen records (the first record – but only a single occurrence – of epidermal fragments of E. tetralix seed is from near the base of the zone), and Calluna pollen is consistently recorded. This indicates that bog and heath communities have a long history locally, even if expansion came much later (mid and late Holocene; cf. Hjelle et al. 2010 for history of heathland in mid-western Norway). Noteworthy also are the substantial values for Nymphaea and Isoëtes. The Isoëtes records may be referred to I. echinospora, based on the small size of the microspores (cf. Birks 1973). This contrasts with the records from Mám Éan in upland Connemara where macrospores only of I. lacustris were recorded and the micro-spores were, by and large, also referable to that species on the basis of size (O’Connell 2021). Today, I. echinospora is the less common quillwort species in Britain and Ireland. There are several records from south-western Ireland but not from the Beara peninsula (Stroh et al. 2023) which may be due partly to under-recording.
Macroremains of mosses indicative of local woodland include Thuidium tamariscinum and Hypnum cf. cupressiforme (Fig. 6). In sample 748 cm (8.6 ka) leaves of Bryum pseudotriquetrum, a fen moss, were recorded and in sample 753 cm (8.8 ka) leaves of Antitrichia curtipendula were noted. There are few present-day and fossil records for A. curtipendula from Ireland (Lockhart et al. 2012; https://www.britishbryologicalsociety.org.uk/ (accessed 19 October 2023)). In Britain it is more common where, today, it has a distinctly northern distribution (mainly Scotland) which is as expected given that it is an European Boreo-temperate moss (Holyoak 2003; Blockeel et al. 2014), though in Europe as a whole an oceanic tendency is apparent (Dickson 1973, p 122). As a moss of open or somewhat shaded places including open woodland and especially pine woodland, in Barrees it was probably a constituent of the bryophyte flora of the local woodlands. There are present-day records from the Beara peninsula (Blockeel et al. 2014) so survival through the Holocene is a distinct possibility.
Zone B2 (736–692 cm; 8.4–7.5 ka, 6465–5505 bce). In this zone, which includes 16 spectra, AP composition maintains the overall pattern established at the end of the previous zone but representation of AP taxa varies within the zone and there are also considerable differences between this and zone B1. There is high and generally increasing Pinus but now with a strong stomatal record and also Pinus leaves are recorded; Figs. 5A, 6). Corylus declines sharply to ∼12% near the base of the profile while Betula expands – B. pubescens macro-remains are frequent – and peaks to 25% at 732 cm. According to the age/depth model, this spectrum dates to 8.3 ka which raises the possibility that these changes reflect the 8.2 ka climate oscillation (Thomas et al. 2007; Bondevik et al. 2012) that favoured cold-tolerant trees such as birch and pine at the expense of thermophilous species such as hazel (Ghilardi and O’Connell 2013; Molloy and O’Connell 2014; Fletcher et al. 2024a). In the mid- and upper parts of the zone, there are also considerable shifts in AP values that involve a temporary but pronounced dip in Pinus that coincides with a peak in Betula (31% at 708 cm; 7.8 ka) and a distinct rise in Quercus that peaks at 26% in the subsequent sample (706 cm). These changes may also be climate driven but differ from what is expected from an 8.2 ka-type oscillation and the age/depth model suggests that the feature is younger. The record of a B. nana catkin bract at 708 cm (ESM 1 Fig. S2 E) is noteworthy insofar as it suggests survival of dwarf birch at this lowland location well into the Holocene (B. nana has not been recorded in the present-day Irish flora; in Britain it is known only from the Cairngorm Mountains, central Scotland; Stroh et al. 2023).
There are also interesting records for minor woody taxa. Alnus is consistently recorded starting from near the top of the previous zone. The values (∼0.2%) are such – also given the high pollen productivity and dispersal capacity of Alnus – that local presence is unlikely. Fraxinus is rather consistently recorded (also in B1 but less frequently) but again the values are very low which argues against local presence. As regards low shrubs, there are several records for Lonicera and Ulex. As under-represented taxa (both are insect pollinated; Lonicera pollen is rather large and heavy; both taxa are recorded in zone A2; Fig. 5A), local presence is likely, quite possibly from ca. 10.7 ka onwards.
Changes in pollen taxa other than AP are small so it seems that woodland composition varied without involving changes in structure and/or degree of openness. Ash content, however, varies considerably with two troughs coinciding with the peaks in Betula at 732 and 708 cm. This is not easily explained particularly in the case of the lower trough, insofar as a climatic downturn, such as the 8.2 ka event, would be expected to result in increased mineral erosion (O’Connell et al. 2014).
Zone C1 (688–664 cm; 7.5–6.8 ka, 5505–4825 bce). In this zone Pinus continues to increase and attains 52% at the top of the zone, Corylus has low representation (it falls to 4.5%), and there are small increases in Alnus and Fraxinus. NAP increases and especially Poaceae which attains close to 7%. There is the beginning of interrupted curves for Plantago. Both P. lanceolata and P. maritima are recorded which points to the presence of both plantain species in grasslands within the region at this time and probably also earlier (occasional pollen of the P. lanceolata were recorded in lower B1). Ericoids, including Calluna, E. tetralix and E. cinerea, are better represented than heretofore. Pediastrum (also Botryococcus, and there is a peak in Isoëtes) increases as do ash values which points to increased erosion.
The pollen data indicate a continued increase in the contribution of pine at the expense of most other trees. Alder and ash may be present but, if so, do not make a substantial contribution. Tilia pollen (as an insect-pollinated tree it is under-represented in pollen records) is recorded in three of the seven spectra (there are also two records in B2) which raises the possibility of the presence of lime in the region. However, such records—mainly single grains; in the present instances < 0.1%, apart from a single spectrum with 0.17%—are rather common in early/mid-Holocene Irish pollen records but, given that there is apparently no supporting macrofossil evidence for the presence of lime in Ireland prior to introduction in recent centuries, it seems best to attribute these early pollen records to long-distance pollen transport from Britain and/or continental Europe (for an opposite view see Stolze and Monecke 2017).
Zone C2 (660–628 cm; 6.8–5.8 ka, 4825–3865 bce). This zone is similar in many respects to C1. The increasing trend in Pinus values, however, is halted until the top of the zone (pine needles abundant; ESM 1 Fig. S2 A–D) where increasing values lead to a peak of 57% at the base of the next zone, i.e. D1. Alnus increases slowly so presumably alder is present if not locally, at least in the west Cork/south Kerry region. A curve for Ilex is initiated which, given the under-representation of Ilex, suggests local presence of holly from ca. 6.1 ka onwards.
NAP (especially Poaceae; also Filipendula and bog taxa) are relatively well represented but decline towards the top of the zone coinciding with a pronounced rise in AP. Interestingly, P. lanceolata is consistently recorded in all spectra at 0.2–0.4% and this before the start of Neolithic farming. Ash content falls to low values (∼30%) towards the top of the zone which suggests low levels of mineral erosion in the catchment. The green algae, Pediastrum and Tetraëdron, are, however, well represented which points to some mineral input to the lake (cf. Bakker and Van Smeerdijk 1982).
Zone D1 (624–604 cm; 5.8–5.37 ka, 3865–3420 bce). In this zone there is a substantial shift in the contribution by various AP taxa. Pinus declines from a sharp peak at the base of the zone, and Betula, Quercus and Ulmus also decline especially in the lower part of the zone. The rather substantial records for Hymenophyllum (mainly H. wilsonii but also H. tunbrigense) in this part of the profile are noteworthy (for a detailed account of the filmy fern record at Barrees see Overland and O’Connell 2008). There is no major change in NAP nor in bog taxa apart from a small increase in P. lanceolata, and ash values remain low. The 14C dating suggests that the decline in Ulmus corresponds with what is normally regarded as the classical mid-Holocene Elm Decline (usually ascribed to ca. 5.8 ka; Parker et al. 2002). Given the low NAP, the decline in Pteridium and the low ash values (but κ values are elevated), the changes recorded are probably the result of factors other than human activity and specifically farming, even though Neolithic culture had spread to Ireland by this time and, in many places, results in substantial and long-lasting Landnam phases (O’Connell et al. 2020).
Zone D2 (600–584 cm; 5.37–5.05 ka, 3420–3095 bce). Pinus continues to decline but most other AP taxa show an increasing trend, and especially Quercus, Corylus and Betula, with the net result that AP is close to its maximum for the profile (Fig. 5). NAP and other indicators of openness, including Pteridium, have particularly low values. Sphagnum (spores; also leaves) is well represented and this continues to be the case over the next several zones. Ash values increase substantially which points to increased mineral erosion. Isoëtes and κ values, however, decline which would normally be regarded as indicating less erosion. Human impact was low during this time so any elevated erosion may have been due to increased runoff, the result of fluctuations in climate and especially rainfall.
Zone E1 (580–536 cm; 5.05–4.2 ka, 3095–2225 bce). This zone, consisting of 12 spectra, records considerable change. Pinus representation is halved to almost 10%, Corylus initially increases but then decidedly declines. A minor Taxus curve with a small peak (6%) occurs mid-zone (548 cm; 4.4 ka). Immediately before the Taxus peak there are two spectra with elevated Pinus values that may be indicative of a ‘pine flush’, i.e. establishment of pine on blanket bog which was widespread in western Ireland at about this time (though usually somewhat earlier, i.e. at ca. 5 ka; O’Connell et al. 2020, 2021; O’Connell 2021; also Discussion).
Towards the top of the zone there is a distinct increase in Pteridium, and NAP also increases including P. lanceolata, and the first cereal-type pollen is recorded. Ericoids also increase which is regarded as reflecting an expansion of bog vegetation and an opening-up of the landscape. There is a large peak in Pediastrum that may have been induced by mineral erosion, a view supported by elevated ash values. The changes in general were probably driven by a variety of factors including climate and pedogenesis, but also increased farming especially towards the top of the zone. The chronology suggests that the farming activity relates to the late Neolithic/Chalcolithic (Fig. 5B).
Zone E2 (532–500 cm; 4.2–3.7 ka, 2225–1785 bce). The trends in the pollen curves established in the previous zone are maintained. AP continues to decrease but the rate of decline in Pinus slows. The continuous stomatal record for Pinus ceases at the top of the zone which suggests that pine may no longer be present in the vicinity of the lake. NAP and especially Poaceae (Molinia caerulea, which is characteristic of blanket bog, may be contributing substantially to Poaceae) increase and also micro- and macro-charcoal values. Characeae oospores are plentiful. Ericoids and Cyperaceae values increase, as do values for other taxa indicative of expansion of bog including Narthecium, Pedicularis, Hypericum elodes-type, Anagallis tenella-type and Sphagnum (spores and leaves). Ash content declines considerably, which suggests low levels of mineral erosion. Overall, there appears to be only moderate levels of human impact.
Zone E3 (496–436 cm; 3.7–2.7 ka, 1785–785 bce). More or less all AP decline and particularly Pinus (stomata recorded in a single sample only). Pinus, however, is still at > 1% as the zone ends which suggests at least regional presence. Fagus (beech) is recorded in the basal spectrum and in three spectra near the top and is more frequent in subsequent zones (to G2/base of G3). As in the case of Tilia, these records are probably best ascribed to long-distance transport. NAP increases substantially including P. lanceolata, Asteraceae (Liguliflorae) and Urtica. Pteridium is well represented and records for Juncus (rush) seeds begin. Cereal-type pollen is recorded in 5 of the 15 spectra and ruderals such as Chenopodiaceae and Brassicaceae are also recorded. Micro-charcoal has distinctly elevated values towards the top of the zone. All in all, the evidence points to less woodland cover, increased openness including an expansion of bog, and farming that was probably mainly pastoral (substantial rise in P. lanceolata) but with an arable component. High ash content, and also elevated κ values, and also increased values for Isoëtes, Botryococcus and Pediastrum suggest active mineral erosion in the catchment.
Zone E4 (432–400 cm; 2.7–1.83 ka, 785 bce–120 ce). AP values increase somewhat but Pinus hovers at ∼1.3%, which suggests it is now very rare in the region and possibly extinct locally. NAP values show little change but P. lanceolata decreases (also ash content) so that farming has probably declined compared to that pertaining in E3. As regards bog/heath taxa, the main change is the substantial increase in Myrica, bog myrtle (it plateaus at > 13% in zone F), which begins above mid-zone, i.e. at ca. 2.3 ka. Preceding this, i.e. at the base of zone E4, there are substantial changes in aquatic taxa (Nymphaea, Isoëtes and Botryococcus; there are peaks in these taxa near the base of the zone) so that climate change – possibly involving drier/wetter shifts – may be the main factor driving change at this time (Overland and O’Connell 2008; Plunkett et al. 2020; for British contexts see, for example, Dark 2006).
Zone F (396–364 cm; 1.83–1.27 ka, 120–685 ce). This distinctive zone is characterised by elevated AP (but declining towards the top) and low values for P. lanceolata and Pteridium. Corylus, Fraxinus, Taxus and especially Betula values increase. Tall shrubs such as Ilex are also better represented. These changes seem to be in response to a decline in farming as signalled especially by the low values for P. lanceolata and also NAP, and especially NAP2 a group that includes mainly pollen of ruderal plants. Zone F is regarded as corresponding to the Late Iron Age Lull (LIAL), a feature of many Irish pollen diagrams during which there is a short-lived regeneration of woody vegetation, and especially trees and shrubs that regenerate early in successional sequences, e.g. Corylus, Betula; also Fraxinus and Taxus (Molloy and O’Connell 2004; O’Connell et al. 2021).
Zone G1 (360–348 cm; 1.27–1.11 ka, 685–845 ce). Zone G1 sets a pattern that is continued to the top of the profile, i.e. to G5. AP declines (more or less all taxa), and NAP, including P. lanceolata, and bog taxa increase. These changes signal increased human activity, and especially pastoral farming, in early Medieval times (for an overview of the relevant archaeology see Comber 2009).
Zone G2 (344–300 cm; 1.11–0.77 ka, 845–1180 ce). NAP increase steadily and there is now a modest NAP2 (ruderal and arable-indicator taxa) complement that includes consistent records of cereal-type pollen. This indicates that farming, now with an arable component, is of increased importance. The first and only record for Secale in G2 is from the base of the zone; in the subsequent zones, there is only the occasional record so rye was never an important crop. In the upper part of the zone, there is increased representation of aquatic and littoral taxa such as Hypericum elodes-type, Typha/Sparganium-type, Nymphaea and Isoëtes which points to lowering of the watertable probably as a consequence of inwash that had a considerable organic component (ash values are < 10%) and autogenic infilling of the lake. The inwash has resulted in reversal in 14C dates and substantially higher sediment accumulation rates (beginning shortly after the bce/ce transition; cf. Fig. 3).
Zone G3 (296–268 cm; 0.77–0.65 ka, 1180–1305 ce). This zone features particularly low AP, distinctly elevated NAP (both NAP1 and NAP2), bog/heath taxa and micro- and macro-charcoal, and much reduced values for aquatic taxa. These changes indicate substantially increased human activity—including pastoral and arable farming—the impact of which was also felt at and within the lake. Interesting also are the records for Juncus (also in G2) which probably derive from wet rushy pastures beside the lake. Zone G3 corresponds to the early Norman period. The arrival of the Normans in eastern Ireland (1169 ce) from which they quickly expanded and the resultant increase in farming and other activities seems to have also been felt in this remote part of south-western Ireland (MacCotter 2004; Breen 2005).
Zone G4 (264–200 cm; 0.65–0.49 ka, 1305–1465 ce). AP increases (also the influx values; ESM 1 Fig. S4), NAP remains steady (but NAP2 increases; ruderals rather than cereal-type pollen are here the main contributors to NAP2) and bog taxa decline. Nymphaea, Isoëtes, Botryococcus and Pediastrum values are greatly elevated, and near the top of the zone Potamogeton peaks and Typha/Sparganium-type and Menyanthes are well represented. This signals substantial changes in the aquatic environment, which, in part at least, is the result of erosional inputs from the catchment. Farming activity continues but probably at reduced intensity so that there is some regeneration of woody species, at least regionally.
Zone G5 (196–184 cm; 0.49–0.45 ka, 1465–1500 ce). In the final four spectra, AP declines and NAP increases and especially NAP1, i.e. pastoral indicators. This suggests increased human activity at the transition from the late Medieval to the early modern period.
Discussion
Given the long and detailed Holocene record that is now available from Barrees we here focus on Holocene rather than Lateglacial environmental change. From Barrees there is not only profile BEG1 from Loch Beag that spans the Holocene, but also several short profiles associated with excavated pre-bog walls of late Iron Age and early Medieval age (BAR-1 to 5), and also a 264 cm long profile BAR-L1 (spans 4.43–0.09 ka, i.e. 2480 bce–1860 ce) from a small peat-filled basin. This basin lies 600 m south-east of BEG1 and usefully complements the information provided by the lake profile BEG1 (cf. Overland and Hjelle (2009) for a similar comparative study from Norway).
Profiles BEG1 and BAR-L1 are summarised in Fig. 7 where a revised age/depth curve for BAR-L1 is given (Fig. 7C). The BAR profiles are presented and discussed in detail in Overland and O’Connell (2008); for archaeological contexts see O’Brien (2009). In the former publication, the records for the filmy ferns and M. gale (bog myrtle) in the BAR profiles and BEG1 are discussed. Here the emphasis is on Holocene environmental change as reflected in BEG1 and other long profiles from the wider region. Our survey includes pollen profiles and also charcoal data from an area stretching from Killarney to the Mizen peninsula, a distance of about 60 km, with the Beara peninsula centrally located (Fig. 1).

Summary percentage pollen diagrams plotted to an age scale: A Profile BEG1 and B Profile BAR-L1. C Age/depth curve for BAR-L1
There are several Holocene pollen profiles from the Killarney area, some of which relate to the early days of pollen analysis (e.g. Erdtman 1924; Jessen 1949; Welten 1952), but there are also several from more recent times. Many of the latter are profiles from small hollows in the Killarney woodlands (e.g. Mitchell 1988, 1990) which, while giving much information on local woodland development and tree population dynamics, are not directly comparable to profile BEG1 from Barrees. Long profiles, however, are available including those by Vokes (1966; see below) and also a long profile from Sheheree Bog (SH) (Mitchell and Cooney 2004). As the pollen data for profile SH are available in Neotoma (Mitchell and Cooney 2019) and also given the detailed analyses, this profile is taken as indicative of overall vegetation development in the Killarney area. Unfortunately, the 14C dates obtained for profile SH proved to be unreliable but the chronology as proposed here (similar to that in Mitchell and Cooney 2004) (ESM 1 Fig. S5 B) fits with what is known generally about vegetation development in the region and so is considered acceptable and appropriate for present purposes.
The profiles described below are from lowland situations (< 90 m except Cashelkeelty at 132 m and Derrycarhoon at 157 m a.s.l.) and, apart from Sheheree Bog which is on Carboniferous limestone with substantial drift cover, Devonian sandstone with little or no drift cover predominates. The sites on sandstone generally support blanket bog and/or heathy vegetation, and are decidedly less fertile which, presumably, was also so in earlier times. Additional information re. site details and the pollen profiles described below are given in ESM 1 Fig. S4.
Sheheree Bog (profile SH), Killarney
Profile SH (Mitchell and Cooney 2004; ESM 1 Fig. S5) features high Juniperus values in the early Holocene. There follows expansions, in turn, of birch (presumably B. pubescens), Corylus and finally Quercus. Oak contributes substantially to the hazel-dominated woodlands for several centuries (probably till ca. 9.2 ka), when pine and also elm, but to a much lesser degree, begin to expand. From then on, oak and pine, with minor contributions by elm and birch, are the main trees of a woodland that includes the filmy fern, H. tunbrigense (there are no records for H. wilsonii; Osmunda has a more or less continuous record from ca. 9.2 ka to 1.3 ka). Shortly after 8 ka, Alnus slowly expands but does not contribute substantially until the later Holocene. An Elm Decline can be distinguished but evidence for a Neolithic Landnam is weak. Neither is there a pine flush, i.e. a short-lived peak in Pinus, at the expected time (ca. 5 ka). Indeed, at this time Pinus values have declined to ∼4% and subsequently fail to make any major recovery until recent times and afforestation. The Pinus stomatal record ceases at 1.3 ka, which suggests survival of a sparse but local population of pine up to that point, i.e. the early Medieval period. Taxus has a more or less continuous curve from ca. 5.3 ka. Evidence for human impact/farming is first obvious shortly after 5 ka (late Neolithic/Chalcolithic), becomes more pronounced at 4 ka (early Bronze Age) and cereal cultivation assumes importance at ca. 2.7 ka but is interrupted in the period (ca. 1.7–1.3 ka) when there is reduced human activity that may be ascribed to the LIAL. Afterwards there is a distinct upturn in human impact that relates to the early Medieval period.
Arbutus pollen (single records in each case) are recorded in three spectra near the top of the diagram (modern times). Nearby on Ross Island, Arbutus was not recorded in a small-hollow profile that spans ca. 4.3 ka to recent times (though Arbutus grows locally and is a common feature of the woods (especially at woodland margins) of the Killarney National Park; Cooney 1996; Mitchell and Cooney 2004). Long pollen profiles from Muckross Bog (Muckross peninsula) and the contrasting upland blanket-bog site, Ladies View, by Vokes 1966 (summarised in Mitchell 1988; pollen data on Neotoma) show that Pinus dominated for most of the Holocene at both sites but it is only in the Muckross profile that there is a Taxus curve that expands at ca. 5 ka (there are no 14C dates so this date is an estimate based on the pollen curves and what is now known about Taxus in other Irish pollen diagrams; cf. Molloy and O’Connell 2014; O’Connell and Molloy 2017). Single grains of Arbutus pollen were recorded in two spectra in both the Muckross and Ladies View profiles; the records can be assigned to the later and later/mid Holocene, respectively, based on the pollen stratigraphy.
L. Inchiquin: Uragh Wood and lake-island pollen profiles, Beara peninsula
Uragh Wood (UW) and L. Inchiquin (LI) pollen profiles derive from small-hollow, peaty deposits near the north-western and south-western ends of L. Inchiquin, respectively (Mitchell 1993; Little et al. 1996). Both relate to the later Holocene and extend to recent times. The UW profile is 14C dated and begins at ca. 3.3 ka (Little et al. 1996). This profile has a substantial Pinus curve until ca. 1.9 ka, with the final demise of pine occurring possibly as late as 1.4 ka, i.e. in early Medieval times. There are consistent but very low Taxus values and Arbutus is not recorded. On the small island, 600 m distant, where core LI was taken, Arbutus dominates and its pollen is substantially represented throughout the LI profile and especially towards the top where it reaches > 10%. Mitchell (1993) suggested, mainly based on the Pinus record from Cashelkeelty (CK-I; see below), that the base of the profile may date to 3.3 ka. Making reference to a much younger date for the final decline of Pinus in the UW profile, Little et al. (1996) suggest that the base of the profile may date to as late as the early Medieval period (1,650 bp). Sheehy Skeffington and Scott (2021, 2022), relying on the latter paper, suggest that the base of the profile and the start of the Arbutus record in LI date to ca. 2,000 bp. Given that the base of the profile has no independent evidence of age and there are no 14C dates for the profile, the dating evidence is weak and especially that suggested for the base of the profile.
Cashelkeelty (profile CK-I), Beara peninsula
Cashelkeelty (Lynch 1981; ESM 1 Fig. S6) is situated at ∼125 m a.s.l., mid-way between L. Inchiquin and Barrees, on the Beara peninsula. The locally flat terrain is largely covered by shallow blanket bog. The site, which includes stone circles, a stone alignment, a large fulacht fiadh (recorded by C. Murphy; see O’Brien 2009, p 50), and pre-bog stone walls, was the main focus of investigations carried out by Lynch (1981) which included archaeological excavations and pollen analytical studies (three short pollen profiles and a 3 m long profile CK-I). Profile CK-I derives from blanket-bog peat at < 200 m from the stone circles and the fulacht fiadh. A consistent set of 14C dates (seven, disregarding the lowermost date which is clearly too young; ESM 1 Fig. S6 B), high pollen counts, identification of critical pollen taxa including separation of Corylus and Myrica (doubtful grains assigned to Corylus), and differentiation of cereal-type pollen have made this a key reference pollen site, especially in archaeological contexts.
Profile CK-I opens shortly after 7 ka. An Elm Decline can be distinguished at the zone CK-I-1/2 transition but an associated opening-up of the pine- and oak-dominated woodlands is not evident in the pollen data. Lynch (1981) interpreted the lower part of the profile (mid- and upper part of zone CK-1, which is equivalent to CK-I-2 as plotted here; it relates to the end fifth and early fourth millennium bce) as indicative of early Neolithic impact that involved woodland clearance and cereal cultivation. This suggests a pre-Elm Decline Landnam which is unexpected within a British/Irish context (cf. Innes et al. 2003 re. possible pre-Elm Decline impact and its manifestation in British/Irish pollen records). The three cereal-type pollen recorded are regarded as derived from cereals, which, given what we know now, is unlikely (O’Connell 1987; Innes et al. 2003). We therefore suggest that it is incorrect to imply that there is evidence for a Neolithic Landnam at Cashelkeelty as has often been done up to now (cf. O’Brien 2009, p 326). There are, however, substantial fluctuations in the pollen curves in both of the first two zones (Lynch 1981; ESM 1 Fig. S6 A), but indicators of human impact such as P. lanceolata do not expand as would be expected if there had been a Landnam event.
By 4.9 ka Pinus has fallen to ∼5% but peaks again to almost 10% at 4.6 ka which is probably indicative of a local pine flush. Pine declines shortly afterwards and is locally extinct by 2.6 ka and possibly earlier (as much as 600 years). Substantial evidence for human activity (mainly P. lanceolata; also cereal-type pollen) is present from ca. 3.2 ka, i.e. late mid-Bronze Age, and is interrupted at the transition to the Medieval period (CK-1–6; probably a LIAL). Myrica is frequent from ca. 3 ka. Taxus, however, is not recorded in this or the short profiles from Cashelkeely, and neither are filmy fern spores or Arbutus.
Palaeoecological investigations on Mizen peninsula
Mount Gabriel (MG4) (Mighall et al. 2008) (ESM 1 Fig. S7 A). The peat-derived profile MG4 spans the Holocene until ca. 3.8 ka (uppermost peat has been cutaway). It is 14C dated but three of the dates are closely spaced (at ca. 7 ka) and pollen-based age criteria are lacking in the profile; e.g. there is no obvious Elm Decline (Ulmus values are very low) nor is there a Neolithic Landnam. The weak chronological control, however, pertains mainly to the top and bottom of the profile and so is not of serious concern in the present context.
Pinus dominates most of the early and mid-parts of the profile until ca. 4.3 ka when it declines from > 20% to ca. 4%, a value that persists to the top of the profile. The first pronounced increase in human impact occurs in the Chalcolithic/early Bronze Age at shortly after 4.3 ka. It follows shortly after the decline in Pinus which begins at/shortly after 4.5 ka and so the pine decline cannot be linked directly to human impact. Taxus pollen is not recorded. Macro-fossil analyses yielded bud scales of Populus (but there are no pollen records) and fruit stones of Viburnum opulus (there are pollen records for Viburnum) (Mighall et al. 2008). The macrofossil records relate to ca. 9.7–8.5 ka. Microspores of Pilularia globulifera are recorded in the early and late part of the profile (centred on 9 and 3.5 ka; ESM 1 Fig. S7 A). A species of wet habitats, this small fern is rare in Ireland where its present-day centre of distribution is Connemara. In south-western Ireland there are records from the Iveragh peninsula but it has not been recorded on Beara or Mizen peninsulas (Stroh et al. 2023). There are interglacial records from Ireland and early to mid-Holocene records from Britain as well as a Lateglacial record (Godwin 1975).
Mighall et al. (2008) emphasise a series of disturbances in the early part of the Holocene (see D1–D4 on the pollen profile, ESM 1 Fig. S7 A). These are reflected in fluctuations in the AP curves, increases in NAP and especially Filipendula, and increased representation of micro- and macro-charcoal. The authors suggest that these perturbations may be attributable to Mesolithic activity but concede that factors other than human impact cannot be excluded. Also in the lower part of the profile, there are major shifts in the Betula and Corylus curves (increase and decrease, respectively; see CO1 and CO2, ESM 1 Fig. S7 A). These may be an expression of climatic oscillations referable to the 9.2 and 8.2 ka climatic oscillations. In CO2, Pinus also increases substantially which may be a response to increased continentality and lower winter temperatures as is known from Ireland during the 8.2 ka event (Ghilardi and O’Connell 2013; Holmes et al. 2016).
Cadogan’s Bog (CB) (Mighall et al. 2004) (ESM 1 Fig. S7 B). Cadogan’s Bog is an extensive, largely cut-away blanket bog, situated about 4 km west of Mount Gabriel. From this bog there is a 14C-dated pollen profile that spans the last ca. 7,000 years, and fossil pine timbers from the bog have been 14C dated and dendrochronologically investigated. The pollen profile is broadly similar to MG4, in that Pinus dominates in the Atlantic period, there is little Alnus and Ulmus (so no Elm Decline) and Taxus is not recorded. Pinus values are elevated between ca. 5–4.4 ka (zone CB-c; ESM 1 Fig. S7 B) which is probably indicative of a pine flush. After this, Pinus declines to ∼5% at 4.2 ka and after 3 ka is generally < 1% which suggests local extinction of pine.
The main results from the investigations of the pine timbers from Cadogan’s Bog are as follows (Mighall et al. 2004; ESM 1 Fig. S7 C). The oldest two timbers (CRO16 and CRO17), which showed a weak dendrochronological match, lay on mineral soil beneath the bog. These have been 14C dated to ca. 7.4 ka (start of life) which, taken in conjunction with the age of the base of profile CB, confirms local peat initiation being underway at ca. 7 ka. Two of the younger dates are from the inner and outer rings of a pine tree (CRO21) with an exceptionally long life-span of 344 y. These dates indicate that this tree began life at ca. 5.3 ka and died shortly before ca. 4.9 ka, i.e. coinciding with the beginning of elevated Pinus pollen values (zone CB-b3). The youngest pine timber dated (4170 ± 50; ca. 4.8 ka; rings 109–139) is probably from the end phase of bog-pine at the site (cf. zone CB-c). However, there was a difference of about 200 y in the age of a peat sample from the core and a pine timber from the same level in the core, the timber giving the older 14C date (Mighall et al. 2004; see also O’Connell et al. 2020 where similar chronological discrepancies are discussed). Also, it is of interest that pine stomata were not recorded during pollen analysis. This has been remarked upon by Mighall et al. (2004), and regarded as unexpected and not easily explained.
In general, the results from Cadogan’s Bog confirm the general picture of bog-pine having a long history in blanket bog contexts in western Ireland. The available 14C dates (calibrated) of pine timbers from western Ireland are plotted in Fig. 8 and details are provided in ESM 2. The dates relate mainly to Mayo and west Galway, and also Clare, Kerry, west Cork and Donegal. In addition to the 14C dates considered in O’Connell et al. (2020, 2021), the new summary includes dates from Co. Clare (3 dates; O’Connell 2022) and Cadogan’s Bog (6 dates; pine timber CRO21 has two dates; only the younger (end of life), i.e. 4,355 ± 55 bp, is included). Also included is the date 3,360 ± 30 bp from a pine timber at Shanvallycahill, Co. Mayo (B. McConnell, pers. comm. 2022). The last mentioned is noteworthy in that it is an outlier with respect to other dates from pine timbers at Shanvallycahill (younger by about a millennium). Similar outliers have been recorded 7 km to the south at Ballydoo Bog, Co. Galway (ca. 3.6 ka; O’Connell et al. 2021, Fig. 14) and also from Gortbrack North, west Mayo (ca. 3.3 ka; Caulfield et al. 1998). There are a few still younger 14C dates from pine timbers available but these have been omitted here as they are regarded as unreliable (see O’Connell 2021 and ESM 2).

Histograms showing frequency of the modal value (µ) of the calibrated age range, classified in 200-y intervals, of 14C-dated timbers from counties/regions (listed north to south) in western Ireland. The number of dates are given in brackets; n = total dates. Further details, including data and sources, are available in ESM 2
That there are outliers as regards the younger end of the age range is not surprising given the wide ecological tolerance of P. sylvestris especially on poor soils, its good seed-dispersal capacity and the inevitable persistence of locally favourable conditions on blanket bog which, by that time, was widespread in western Ireland. There is also the pollen analytical evidence for survival of pine at least in south-west Ireland (see above) into the 1st millennium ce and probably into the early Medieval period.
Arderrawinny (ARD), south-west of Mount Gabriel, near Toormore (Kearney 2019). The small peat-filled basin, Arderrawinny, which was investigated pollen analytically, is located between two nearby megaliths, i.e. a portal tomb (early Neolithic) to the north-east and a large wedge tomb (late Neolithic/early Bronze Age). The 14C-dated, detailed profile ARD spans ca. 10–2 ka. As the profile opens, oak is the dominant tree, pine starts to expand at ca. 8.2 ka (the 8.2 ka oscillation may be reflected by the small peak in Pinus and corresponding decrease in Corylus) and is dominant during the mid-Holocene. Alnus, Ulmus and Fraxinus are poorly represented throughout and there are only a few records of Taxus pollen. Pinus declines in three stages, at ca. 5, 4.4 and 3.9 ka (from > 50% to ca. 10% and then the curve ceases). A feature of the record is the evidence for woodland disturbance (increased NAP and especially P. lanceolata) in pre-Elm Decline spectra, i.e. prior to the start of the Neolithic in Ireland. It is suggested that ‘pastures/meadows’ were already present at the site during the late Mesolithic (Kearney 2019, p 8). Despite the proximity of megaliths, human impact is not pronounced at any point in the profile perhaps on account of the local character of the record (as expected given the small basin size). High Osmunda values, achieving > 40% post Elm Decline, also serve to emphasise the local character of the profile.
Derrycarhoon (DER), north-east of Mount Gabriel. Kearney and O’Brien (2021) present the results of palynological analyses from near a small prehistoric mine in the Mizen peninsula. The mining activity has been dated to 1400–1000 bce, i.e. the later Bronze Age, and so represents an end-phase of prehistoric MB-type mining activity that is typical of the Mizen and Beara peninsulas.
Core DER is from a bog at 160 m a.s.l. and 35 m south-west of a small Cu-mine. The pollen profile is < 1.9 m long and spans most of the Holocene (ca. 11–0.23 ka). It is quite similar to the ARD profile which is from near sea level at the other end of the Mizen peninsula. Pinus is dominant (achieves 55%), Quercus values vary between 10 and 25%, but other AP taxa (even Betula) are represented by interrupted curves and invariable by very low values. Pinus declines from high values between ca. 3.7–3.5 ka, and at ca. 1.9 ka a continuous curve more or less ceases which presumably reflects local extinction of pine. Human impact, mainly seen in elevated P. lanceolata and lower Quercus values, registers relatively strongly in the intervals ca. 4.3–3.9 ka (Chalcolithic/early Bronze Age) and again at ca. 2.9–2 ka (late Bronze Age to early and mid-Iron Age). Fraxinus and Taxus have very low values. Wooden artefacts made of yew wood have been recovered from the site (Briggs 1984) and a tube-like yew artefact has been 14C dated to the early 2nd millennium ce (Kearney and O’Brien 2021, p 62).
Remarkably, there is little or no impact registering at the time when Bronze Age mining was taking place. In these particular mines, fire-setting, which requires substantial wood, was not used which may account for the apparent lack of impact in the pollen record (Kearney and O’Brien 2021). However, a distinct decline in the organic component of the core that coincides with the mining may be indicative of increased inwash arising from mining (Kearney and O’Brien 2021). On the basis of the palynological results from this and other prehistoric mining sites elsewhere in Europe, the authors conclude that the scale and intensity of prehistoric mining was such that its environmental impact was much smaller than that associated with mining in historical times (see also O’Brien 2015).
Charcoal analyses from Barrees and other Cu-mining sites in south-western Ireland
In this section, results of published charcoal data from archaeological sites in south-western Ireland, and in particular, from areas of greatest interest namely, Barrees (Van Rijn and Vorst 2009), Mount Gabriel (MG; McKeown 1994) and Ross Island (Van Rijn 2004), are discussed. The results of analyses of waterlogged wood from Mine 3 at MG are also considered. The data from these three areas provide insights into local environments, at a local scale, that most usefully complement the more general information provided by pollen analyses. Furthermore, they provide firm evidence for local presence, which is important in the case of trees and shrubs that are under-represented or indeed silent in pollen records, or are so rare that they are often not captured at all in pollen data.
Barrees (Fig. 9A). Results of charcoal analyses of Barrees material are available from eight contexts relating to six sites that include the Cu mine at Crumpane (J), two fulachta fiadh (B and C), a house site (D), a hut site (F) and a charcoal-making pit (G) (capitals letters designate archaeologically investigated sites; Van Rijn and Vorst 2009; Fig. 1). In all 744 fragments were identified, representative of at least nine different species (Van Rijn and Vorst 2009). The date range represented in the dataset extends from the early mid-Bronze Age to the late Medieval (details in Fig. 9A). The oldest and youngest sites (Cu-mine and charcoal-making pit; J and G) have the least variety, the latter being dominated by oak and the former consisting exclusively of birch. According to Van Rijn and Vorst (2009) this probably indicates selective use of these trees rather than local frequency. These sites are also from the ends of an altitude range (250 vs 115 m). Birch was probably locally frequent at the higher altitude (Crumpane) on account of its high regenerative capacity and wide ecological tolerances that enabled it to rapidly exploit clearances in the immediate vicinity of the mine, even under unfavourable soil and climatic conditions. The absence of pine charcoal is noteworthy. It is not recorded even at the mine site, which contrasts with the MG and Ross Island charcoal data (see below). Failure to record pine charcoal supports the stomatal evidence from profile BEG1, Loch Beag, that pine was extinct locally by ca. 3.7 ka (Fig. 5). On the other hand, the records for Ulex (furze) serve to confirm local presence during prehistory (mid-Bronze Age and early Iron Age) and thus support the pollen evidence that suggests local presence for most of the Holocene and particularly in the historical period (Fig. 5A). As regards the species present, the two likely species, U. europaeus and U. gallii (both common locally today) have not been distinguished in the wood or pollen records. Both species are generally regarded as native (Parnell and Curtis 2012). Elm was not recorded in the charcoal data, which is not surprising given the very low Ulmus pollen values. Fraxinus charcoal is only recorded at the hut site (J; Medieval) and Taxus charcoal goes unrecorded. The pollen record, however, suggests low but consistent presence of both ash and yew, particularly during prehistory.

A Plot of charcoal data from excavated sites in Barrees (data source: Van Rijn and Vorst 2009). For each site, the percentage frequency of fragments of the respective taxa is indicated. Abbreviations used to indicate site type: Cu, copper mine; FF, fulacht fiadh; H, house; F, hut site; G, charcoal pit. B Charcoal quantities (g) recovered from Levels 3–6 in spoil heaps relating to Mines 3 and 4, and charcoal recorded within Mine 3 at Mount Gabriel (data source: McKeown 1994)
Mount Gabriel. At MG, charcoal and waterlogged wood data are available from Mines 3 and 4 (832 fragments in all; McKeown 1994). Mine 4 probably came into operation immediately after operations at Mine 3 ceased and so the charcoal and wood data relate to approximately the same time, i.e. ca. 1700–1500 bce (ca. 3.6 ka; Brindley and Lanting 1994; see also O’Brien 2003). Most of the waterlogged wood (from Mine 3), mainly arising from the abandonment of Mine 3, consisted mostly of roundwood with bark. Corylus and Quercus predominated, Fraxinus was important, and Salix, Alnus, Betula and Ilex were recorded in small quantities.
Charcoal data (weights) relating to spoil heaps from Mines 3 and 4 (layers 3–6) and those relating to fire setting within Mine 3 are presented in Fig. 9B. These data show the relative importance of various trees and shrubs during the operation of Mines 3 and 4. Quercus and Salix are, by far, the main taxa represented. It is noteworthy that willow was not at all as frequent in the waterlogged samples or indeed in the charcoal from within Mine 3. The low but consistent presence of Pinus is regarded by McKeown (1994) as reflecting use limited to chips for lighting in the coarse of mining and this, in turn, suggests local scarcity of pine as indicated by the pollen data from the vicinity of MG (Mighall et al. 2004; ESM 1 Fig. S7 A) and from the Mizen peninsula generally (Mighall et al. 2008; Kearney 2019; Kearney and O’Brien 2021). The records for Fraxinus fit in with the expansion of ash (though Fraxinus representation is rather weak in the pollen record) from the late Neolithic onwards. Taxus is not recorded which again fits with the evidence, i.e. none or few Taxus records in pollen profiles from the Mizen peninsula (see above). The scarcity of Corylus (also Alnus, except in Level 6 where it is well represented) in the charcoal records is difficult to explain, especially given the high frequency of Corylus in the waterlogged samples and the role of Corylus as a pioneer shrub.
Ross Island. At Ross Island, charcoal samples were analysed from six archaeological contexts distinguished during the course of excavations at the mining site (3,643 fragments; 16 taxa identified). A summary of the results pertaining to each cultural period, the date range of the samples, the main taxa recorded and a short commentary follow (after Van Rijn 2004).
Periods 1 and 3. Period 1 relates to the mid-Mesolithic, i.e. ca. 7.5–7.3 ka. The analyses are limited to three samples; only oak was recorded in two samples while in a third it was mainly Alnus, some Quercus and two fragments of Fraxinus. The Fraxinus record is interesting in that it confirms the presence of ash in the Mesolithic and supports a pre-Elm Decline presence as indicated by low percentage values in many detailed pollen records from Ireland, including relatively substantial values in profile BEG1. The absence of pine is surprising given its importance as reflected in pollen diagrams and also its importance in samples of Bronze Age date at Ross Island. Period 3 relates to renewed Cu working in the early Medieval. One sample consisted only of oak while two samples included oak and yew, the latter being less frequent.
Period 2: Beaker and early Bronze Age 4.35–3.9 ka. Most of the samples (43 samples in all) and the fragments identified (3,150) relate to this period. The highest taxon diversity is also recorded in this period (average eight taxa per sample; in all, 16 different taxa). Several shrubs were recorded such as Ilex aquifolium (holly; 65%), apple/pear/hawthorn (30%), Arbutus unedo (strawberry tree; 12%), Euonymus europaeus (spindle; 7%) and Prunus avium (wild cherry; 7%) (percentages are based on presence in the 43 samples relating to Period 2). The taxon ‘apple/pear/hawthorn’ potentially includes crab apple (Malus sylvestris), pear (Pyrus communis) and hawthorn (Crataegus spp.). P. communis is unlikely to have been present in the charcoal records given that it is rare in Ireland and is generally regarded as an introduction (Stroh et al. 2023). Prunus spinosa (blackthorn) was not recorded which is surprising given its present-day status as a widespread, native species in Ireland.
These shrub records are indicative of opening-up of the woodlands presumably as a result of human impact. Unsurprisingly, in view of the low values of Ulmus in pollen diagrams from the region, elm is recorded in only 7% of the samples. This is in sharp contrast to the dominance of Quercus (84%), Alnus (65%), and also Pinus and Taxus (60%). The high values for the last mentioned trees may, however, be due to selective use rather than high frequency in the local woodlands at this time.
Based on the 14C dates and also the archaeological contexts, Van Rijn (2004) (also O’Brien 2004, p 168) suggests the following subdivision of period 2: sub-period 2A (4.35–4.2 ka) in which Quercus dominated, Pinus and Alnus were recorded in substantial quantities, and Fraxinus, Corylus, Taxus and Ilex being rare; and sub-period 2B (4.15–3.9 ka), in which Alnus was the most important wood, followed by Quercus and Fraxinus, and then Pinus, Corylus, Taxus and Ilex, with the last three mentioned taxa making greater contributions than in sub-period 2A. These shifts in composition are probably best attributed in large measure to human impact involving woodland clearance but natural processes, associated with regeneration and competition, were undoubtedly also important.
From a biogeographical perspective, the most interesting records are undoubtedly those of Arbutus unedo. Charcoal assignable to this tall shrub/tree was recorded in small quantities in four samples and possibly in a fifth (included in the percentage value given above) but definitive identification proved impossible in that instance. The samples come from three different contexts relating to the Bronze Age mining operation. Three of the samples have been assigned to period 2B; none have been assigned to period 2A (Van Rijn 2004). There are no 14C dates deriving directly from Arbutus but the records are well constrained chronologically by their archaeological contexts and associated 14C dating (W. O’Brien, pers. comm. 2023). Van Rijn (2004, p 397) indicates that “The Arbutus charcoal from excavation C.03 and C.440 at Ross Island is firmly dated to the Period 2 (Beaker/Early Bronze Age) …. [i.e.] broadly between 2400 and 1900 bc (3,900–3,600 bp)”, i.e. the records in question date to ca. 3.9 ka at the earliest. Based on this, Van Rijn 2004, p 398 concluded, “This provides the earliest secure evidence for the native character of the species [A. unedo]”. Mitchell and Cooney (2004, p 490; i.e. in the same volume) concur with this view: “This would appear to confirm the native status of this tree in Killarney”. These views contrast with those of Sheehy Skeffington and Scott (2021, p 411), who conclude that A. unedo “is probably not a native species to Ireland but is an ancient archaeophyte introduced more than 4,000 years ago to beside Lough Leane Co. Kerry from northern Iberia by miners that came there to extract the copper”. Given the charcoal evidence, this linkage with early miners can certainly be advanced with some justification for Ross Island but many questions remain, not least proving that an introduction took place rather than the tree/shrub responding positively, like many of the other light-demanding trees and shrubs, to the clearances effected by the early Bronze Age miners. It is noteworthy that Arbutus does not appear to be associated with later Bronze Age mining in the Kerry/west Cork region. At least there are no pollen or charcoal/wood records for Arbutus from the Mizen peninsula (the centre of Bronze Age mining of the MG type; see above), nor indeed from Barrees which has a MG-type mine (with associated charcoal analyses) and where large quantities of pollen have been counted in long and short profiles with particular attention being paid to identifying ericoid pollen to genus/species level. So it seems that the spread of Arbutus, at least within Ireland and specifically in the Kerry/west Cork region, is not particularly associated with prehistoric Cu-mining, as has been intimated by Sheehy Skeffington and Scott (2021). If there were an introduction from northern Spain to Ross Island by the first miners on account of the cultural and other importance of Arbutus to these peoples as suggested by Sheehy Skeffington and Scott (2021), it seems strange that the process of introduction (now much easier as there was a readily available source in the Killarney area) was not repeated by Bronze Age miners in adjacent parts of Kerry and west Cork during later centuries. It should also be noted that the archaeological record from Ross Island, where linkage between Beaker culture and the mining activity is a key feature, does not provide support for contacts with Spain at this time (O’Brien 2015; W. O’Brien, pers. comm. 2023). Phylogeographic studies of Arbutus and other Lusitanian species, however, indicate close genetic relationships between the Iberian and Irish populations but these fall far short of providing evidence for introductions by people at specific times and locations (Beatty et al. 2015; Santiso et al. 2016; Fagúndez and Díaz-Tapia 2023).
Conclusions
The conclusions as summarised here relate to the lake profile BEG1 from Barrees, Beara peninsula, unless otherwise indicated (Figs. 4, 5). The Younger Dryas stadial resulted in a tabula rasa as regards tall shrubby vegetation which began to spread before/at the Younger Dryas/Holocene transition (11.7 ka; dates cited here and below are best approximations based on the age/depth curve in Fig. 3). In the early Holocene, tall shrub/tree succession involved initially Juniperus, then Salix and Betula, and finally Corylus.
Quercus is the first tall-canopy tree to spread and expand (before 10 ka and 9.6 ka, respectively; the dates from 10 ka onwards derive from that part of the age/depth model that is based on 14C AMS dates; Fig. 3), followed by Pinus for which local presence is confirmed by stomatal records beginning at 8.64 ka and ceasing at 3.7 ka (outlying oldest and youngest records date to 8.84 and 3.5 ka, respectively). Ulmus may have been present locally prior to the expansion of Pinus but this is uncertain given its low pollen values in the early Holocene and throughout the profile (and also in profile, BAR-L1, from the nearby small bog).
The first records for Osmunda pre-date the expansion of Corylus (there are consistent records from 11 ka). The Hymenophyllum wilsonii records begin later (9.2 ka), are at first very sporadic, then more or less continuous (from 5.7 ka), and end at ca. 0.7 ka. H. tunbrigense is fairly consistently recorded in the mid-Holocene (5.5–3.3 ka) and thereafter sporadically to near the top of the profile (0.46 ka). As regards Trichomanes speciosum (ESM 1 Fig. S2 F), its fossil spores have not been recorded in profile BEG1 but there are records from other profiles in Barrees including BAR-L1 in which single spores were recorded in three spectra, the earliest dating to 3.1 ka (Overland and O’Connell 2008).
Other tree and tall shrub taxa that are seldom recorded in Holocene pollen diagrams and for which are substantial early Holocene records include Populus, Viburnum (cf. opulus), Sorbus and Ulex. There are also early records (from 9 ka onwards) for several ericoid species, including Erica tetralix and E. cinerea; these have elevated values beginning at 5 ka (Fig. 5C). More or less continuous curves for other bog/heath taxa begin at about 5 ka, e.g. Narthecium ossifragum, or afterwards in the case of Hypericum elodes, Pedicularis palustris and Anagallis tenella-type which expand after about 3.7 ka, and Myrica gale which has a consistent record that begins at 3.2 ka and expands greatly at 2 ka. Noteworthy early Holocene macrofossil records include leaves of the moss, Antitrichia curtipendula (8.8 ka), and a catkin bract of the dwarf birch, Betula nana (7.8 ka) (Fig. 6).
As in many detailed pollen diagrams from Ireland, a curve for P. lanceolata commences before the Elm Decline (start date in BEG1 is 6.8 ka; sporadic records of P. lanceolata and P. maritima begin at 9.1 and 10.4 ka, respectively). Given the early dates and scarcity of evidence for Mesolithic activity in the region and especially on Beara (O’Brien 2009, p 326), these records can hardly be attributed to human activity; rather perturbations in woodland cover due to natural causes seem to have created conditions favourable for Plantago spp already in Atlantic times (there is considerable evidence to suggest that the mid-Holocene was a time of considerable climate instability; cf. Bonsall et al. 2002; Wanner et al. 2011; Achterberg et al. 2018; Plunkett et al. 2020; Fletcher et al. 2024b). An Elm Decline at 5.82 ka is recorded but there is little evidence for early Neolithic impact which accords with the archaeological evidence (O’Brien 2009). Human impact, as reflected especially in the NAP component, begins to register at about 5 ka (mid-Neolithic), increases substantially shortly before 4 ka (Chalcolithic/early Bronze Age) and with a further increase commencing at ca. 3 ka (late Bronze Age). Thus the Late Neolithic and especially the Bronze Age emerge as a time of major human impact, a view strongly supported by the archaeological evidence from Barrees and the Beara peninsula generally (O’Brien 2009).
A lull in human activity and woodland regeneration, i.e. a Late Iron Age Lull, is recorded between ca. 1.83 and 1.26 ka. In BAR-L1 a woodland regeneration phase is also recorded but it is of much longer duration. Interestingly, there is considerable archaeological evidence for prolonged human activity at higher elevation in the late Iron Age (especially at site A; O’Brien 2009) so that the lull should not be equated with absence of human impact; rather activity seems to have been centred at higher elevation (favoured by distinct climatic amelioration at about this time or earlier; cf. Lamb 1995; McCormick et al. 2012) and hence goes largely unrecorded in the relatively distant BEG1 profile.
Shortly after 1.1 ka, the final woodland clearances commence in the context of greatly increased human activity in the Medieval period. By about 1 ka, peat development had commenced on a wide scale in the archaeologically investigated area (Overland and O’Connell 2008), including that associated with the pre-bog walls that have been shown to relate to late prehistory/early historical period (late Iron Age to early Medieval period; O’Brien 2009).
Pinus steadily declines from shortly after the Elm Decline until ca. 5 ka, then more slowly until ca. 3.7 ka. It undergoes final decline between ca. 3.7 and 1.7 ka when pine probably became regionally extinct. A compilation of 14C dates from bog-pine timbers in western Ireland shows that, while there is a large range of dates (8.8–3.3 ka), the frequency peaks in the interval 5.2–4.6 ka (57.4% of dates lie within this interval) after which the frequency rapidly declines (Fig. 8). These changes are assumed to be mainly climate driven (cf. Plunkett et al. 2020).
There is a more or less continuous Taxus curve between 5.1 and 1.3 ka, but the values are small (generally 1–2%). Later, Taxus values are generally < 0.5%. It seems that yew, and also pine growing on mineral soils, were adversely affected in Barrees and in the wider region by several factors, often operating over a long period, including climate change, human impact, pedogenesis and successional processes.
A review of Holocene vegetation dynamics and human impact based on key published pollen diagrams from the wider region, charcoal and macrofossil records from archaeological excavations, and bog-pine 14C dates provide an added spatial, indeed regional, dimension to the detailed records from Barrees. There is a consistent sequence of tree expansion in the early Holocene, namely birch and hazel (in that order; hazel particularly pronounced), then oak which is followed by pine. Pine goes on to achieve dominance or co-dominance with oak at most sites. Elm, which probably arrives after oak, never makes a major contribution. At Sheheree, however, where soils derive from limestone (bedrock and drift) and hence are fertile, oak remains dominant and is better represented than at other sites. Edaphic factors have probably been decisive in determining the outcome of the long-term competition between these tall-canopy trees. Alder, which had spread into the region by ca. 7 ka, does not achieve major importance and, when expansion occurs, it is rather late (e.g. in BEG1 it is shortly after 6 ka). This is surprising in that wet habitats are common. Ash also fails to expand greatly. Once considered to be a tree that registered in Irish pollen records almost exclusively in post-Elm Decline contexts, it is clear that it had achieved some importance much earlier in our region. At Loch Beag, for instance, it was probably present before alder (at ca. 7.5 ka). Its early presence (Mesolithic; pre-Elm Decline) at Ross Island is confirmed by charcoal records (Van Rijn 2004).
Yew, which shows a major expansion in some Irish pollen diagrams at about 5 ka or later and sometimes less pronounced during the LIAL (this is the pattern in BEG1 though Taxus representation remains low), never achieves importance in the region. Indeed, several of the pollen profiles have no records. This rather difficult-to-recognise pollen (especially if present in small quantities) may have gone unrecognised or largely unrecognised, and so is probably under-recorded. Yew has not been recorded in the charcoal records from Barrees or Mount Gabriel but it is important at Ross Island in samples that relate to the early Bronze Age (ca. 4 ka). This is not surprising given the major role Taxus plays at sites in the nearby Muckross peninsula in post-Elm Decline contexts (Mitchell 1988, 1990). That yew survived in the region into historical times is indicated by the yew-wood artefacts of Medieval age that have been recorded at Derrycarhoon (see above).
The later Holocene is characterised by woodland demise, expansion of blanket bog and varying, but generally strong, human impact. Unusually for Ireland, the evidence for early and mid-Neolithic impact is low, even where there is strong archaeological evidence for the presence of early farmers (e.g. Arderrawinny, Mizen peninsula; Kearney 2019). This is not altogether surprising, however, given that the archaeology of the region is predominantly Bronze Age in character. In agreement, the pollen data reflects strong human impact beginning before 4 ka, i.e. at the late Neolithic/Chalcolithic transition (dates, etc. relate mainly to the BEG1 profile and the investigations at Barrees). Prehistoric copper mining, however, does not appear to have had major environmental impact.
Human impact is at its highest for the prehistoric period in the interval ca. 3.8–2 ka, i.e. early Bronze Age to mid-Iron Age presumably due to population increase and pressures arising from farming. During this time, pine declines and probably becomes extinct at Barrees and the region generally. Strong human impact was probably important but other factors—increased wetness; spread of bog, etc.—undoubtedly also contributed. The farming economy during this time was mainly, but not exclusively, pastoral based. At Barrees, the archaeological record derived from extensive excavations, shows the late Iron Age to be of particular importance in upland parts (O’Brien 2009). The pollen record, however, points to a lull in human impact leading to woodland regeneration in the lowlands so there appears to have been small-scale spatial variation in land-use at this time.
The present-day landscape, with extensive blanket bog and trees limited to small copses mainly in the lowlands, has its origins in Medieval times and especially from the late 1st millennium ce onwards. It is the product of human activity acting in conjunction with a wet, cool climate and largely acidic soils. For further consideration of the relationships between climate, farming economies and human impact generally during the Medieval and later in upland areas, including Barrees, see Costello et al. 2023.
The biogeography of various floristic elements of the regional flora is considered in some detail. Particular attention is paid to the records of ericaceous species including Arbutus unedo which has present-day Irish centres of distribution in the Killarney National Park and the Beara peninsula. On the basis of the fossil and other evidence, it is argued that the recent suggestion that A. unedo is an archaeophyte brought from Iberia to Killarney by early copper miners (Sheehy Skeffington and Scott 2021) remains unproven and probably over-simplifies the history of this distinctive Lusitanian element in the Irish flora.
Data availability
The pollen data are being prepared for submission to PANGAEA. Other data are made available in the ESM files.
References
Aaby B, Berglund BE (1986) Characterization of peat and lake deposits. In: Berglund BE (ed) Handbook of Holocene palaeoecology and palaeohydrology. Wiley, Chichester, pp 231–246
Aalen FHA, Whelan K, Stout M (eds) (2011) Atlas of the Irish rural landscape, 2nd edn. Cork University Press, Cork
Achterberg IEM, Eckstein J, Birkholz B, Bauerochse A, Leuschner HH (2018) Dendrochronologically dated pine stumps document phase-wise bog expansion at a northwest German site between ca. 6700 and ca. 3400 BC. Clim past 14:85–100
Andrieu V, Huang CC, O’Connell M, Paus A (1993) Lateglacial vegetation and environment in Ireland: first results from four western sites. Quat Sci Rev 12:681–705
Atherton I, Bosanquet S, Lawley M (2010) Mosses and liverworts of Britain and Ireland. A field guide. British Bryological Society, Plymouth
Bakker M, Van Smeerdijk DG (1982) A palaeoecological study of a late Holocene section from “Het Ilperveld”, western Netherlands. Rev Palaeobot Palynol 36:95–163
Ballantyne CK, McCarroll D, Stone JO (2011) Periglacial trimlines and the extent of the Kerry-Cork Ice Cap, SW Ireland. Quat Sci Rev 30:3,834–3,845
Beatty GE, Lennon JJ, O’Sullivan CJ, Provan J (2015) The not-so-Irish spurge: Euphorbia hyberna (Euphorbiaceae) and the Littletonian plant ‘steeplechase.’ Biol J Linn Soc 114:249–259
Beijerinck W (1947) Zadenatlas der Nederlandsche flora. Ten behoeve van de botanie, palaeontologie, bodemcultuur en warenkennis. Veenman en Zonen, Wageningen
Beug H-J (2004) Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete. Pfeil, München
Birks HJB (1973) Past and present vegetation of the Isle of Skye: A palaeoecological study. Cambridge University Press, Cambridge
Blaauw M (2010) Methods and code for ‘classical’ age-modelling of radiocarbon sequences. Quat Geochronol 5:512–518
Blockeel TL, Bosanquet SDS, Hill MO, Preston CD (eds) (2014) Atlas of British & Irish bryophytes, Vols 1 & 2. NatureBureau (Pisces Publications for the British Bryological Society), Newbury
Bondevik S, Stormo SK, Skjerdal G (2012) Green mosses date the Storegga tsunami to the chilliest decades of the 8.2 ka cold event. Quat Sci Rev 45:1–6
Bonsall C, Anderson DE, Macklin MG (2002) The Mesolithic-Neolithic transition in western Scotland and its European context. Doc Praehist 29:1–19
Boyd WE, Dickson JH (1987) A post-glacial pollen sequence from Loch a’Mhuilinn, north Arran: a record of vegetation history with special reference to the history of endemic Sorbus species. New Phytol 107:221–244
Breen C (2005) The Gaelic Lordship of the O’Sullivan Beare. A landscape cultural history. Four Courts Press, Dublin
Briggs S (1984) The discovery and description of trench mines at Derricarhoon TD, Co. Cork, in 1846. J Ir Archaeol 2:33–39
Brindley A, Lanting JN (1994) The radiocarbon chronology of mines 3 and 4, Mount Gabriel. In: O’Brien W (ed) Mount Gabriel. Bronze Age copper mining in Ireland. Bronze Age Studies 3. Galway University Press, Galway, pp 281–287
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360
Caulfield S, O’Donnell RG, Mitchell PI (1998) 14C dating of a Neolithic field system at Céide Fields, County Mayo, Ireland. Radiocarbon 40:629–640
Comber M (2009) Beara in the first millennium AD. In: O’Brien W (ed) Local worlds. Early settlement landscapes and upland farming in south-west Ireland. Collins Press, Cork, pp 58–68
Cooney T (1996) Vegetation changes associated with late-Neolithic copper mining in Killarney. In: Delaney C, Coxon P (eds) Central Kerry. Field Guide No. 20. Irish Association for Quaternary Studies, Dublin, pp 28–32
Costello E, Kearney K, Gearey B (2023) Adapting to the Little Ice Age in pastoral regions: an interdisciplinary approach to climate history in north-west Europe. Hist Methods: J Quant Interdiscip Hist 56:77–96. https://doi.org/10.1080/01615440.2022.2156958
Coxon P (1997) Pleistocene climate change: the evidence from Irish sequences. In: Sweeney J (ed) Global change and the Irish environment. Royal Irish Academy, Dublin, pp 17–35
Craig AJ (1978) Pollen percentage and influx analyses in south-east Ireland: a contribution to the ecological history of the Late-glacial period. J Ecol 66:297–324
Cross JR (2006) The potential natural vegetation of Ireland. Biol Environ (Proc R Ir Acad, Ser B) 106B:65–116
Daltun E (2022) An Irish Atlantic rainforest. A personal journey into the magic of rewilding. Hachette Books Ireland, Castleknock (Dublin)
Dark P (2006) Climate deterioration and land-use change in the first millennium BC: perspectives from the British palynological record. J Archaeol Sci 33:1381–1395
Dickson JH (1973) Bryophytes of the Pleistocene. The British record and its chorological and ecological implications. Cambridge University Press, Cambridge
Edwards KJ, Whittington G (2001) Lake sediments, erosion and landscape change during the Holocene in Britain and Ireland. Catena 42:143–173
Erdtman OGE (1924) Mitteilungen über einige irische Moore. Sven Bot Tidskr 18:451–459
Faegri K, Iversen J (1989) Textbook of pollen analysis. 4th edn. by Faegri K, Kaland PE, Krzywinski K. Wiley, New York
Fagúndez J, Díaz-Tapia P (2023) Comparative phylogeography of a restricted and a widespread heather: genetic evidence of multiple independent introductions of Erica mackayana into Ireland from northern Spain. Bot J Linn Soc 201:329–340
Fletcher WJ, Sánchez Goñi MF, Naughton F, Seppä H (2024a) Greenlandian Stage (Early Holocene, 11.7–8.2 ka). In: Palacios D, Hughes PD, Jomelli V, Tanarro LM (eds) European glacial landscapes. The Holocene, Elsevier, Amsterdam, pp 73–87
Fletcher W, Sánchez Goñi MF, Naughton F, Seppä H (2024b) Northgrippian Stage (Middle Holocene, 8.2–4.2 ka). In: Palacios D, Hughes PD, Jomelli V, Tanarro LM (eds) European glacial landscapes. The Holocene. Elsevier, Amsterdam, pp 89–104
Ghilardi B, O’Connell M (2013) Early Holocene vegetation and climate dynamics with particular reference to the 8.2 ka event: pollen and macrofossil evidence from a small lake in western Ireland. Veget Hist Archaeobot 22:99–114
Godwin H (1975) The history of the British flora: a factual basis for phytogeography, 2nd edn. Cambridge University Press, Cambridge
Harrison S, Glasser N, Anderson E, Ivy-Ochs S, Kubik PW (2010) Late Pleistocene mountain glacier response to North Atlantic climate change in southwest Ireland. Quat Sci Rev 29:3948–3955
Hjelle KL, Halvorsen LS, Overland A (2010) Heathland development and relationship between humans and environment along the coast of western Norway through time. Quat Int 220:133–146
Hodd RL, Rumsey F (2020) Stenogrammitis myosuroides (Polypodiaceae): a new addition to the European flora from southwest Ireland. Brit Irish Bot 2:158–168
Holmes JA, Tindall J, Roberts N et al. (2016) Lake isotope records of the 8200-year cooling event in western Ireland: comparison with model simulations. Quat Sci Rev 131:341–349
Holyoak DT (2003) The distribution of bryophytes in Ireland. An annotated review of the occurrence of liverworts and mosses in the Irish vice-counties, based mainly on the records of the British Bryological Society. Broadleaf Books, Dinas Powys (Wales)
Innes JB, Blackford JJ, Davey PJ (2003) Dating the introduction of cereal cultivation to the British Isles: early palaeoecological evidence from the Isle of Man. J Quat Sci 18:603–613
Jessen K (1949) Studies in Late Quaternary deposits and flora-history of Ireland. Proc R Ir Acad, Sect B 52:85–290
Kearney K (2019) Vegetation impacts and early Neolithic monumentality: a palaeoenvironmental case study from south-west Ireland. J Archaeol Sci Rep 27:101940
Kearney K, O’Brien W (2021) Palynological investigation of a later Bronze Age copper mine at Derrycarhoon in south-west Ireland. Proc R Ir Acad, Sect C 121:55–90
Kelly DL (1981) The native forest vegetation of Killarney, south-west Ireland: an ecological account. J Ecol 69:437–472
Kinnaird JA, Nex PAM (2004) The geological setting. In: O’Brien W (ed) Ross Island. Mining, metal and society in early Ireland. Bronze Age Studies 6. Department of Archaeology, National University of Ireland (NUI), Galway, pp 23–63
Kollmann J, Grubb PJ (2002) Viburnum lantana L. and Viburnum opulus L. (V. lobatum Lam., Opulus vulgaris Borkh.). J Ecol 90:1044–1070
Körber-Grohne U (1964) Bestimmungsschlüssel für subfossile Juncus-Samen und Gramineen-Früchte. Probleme der Küstenforschung im südlichen Nordseegebiet 7. Lax, Hildesheim
Krüger S, Fischer Mortensen M, Dörfler W (2020) Sequence completed—palynological investigations on Lateglacial/Early Holocene environmental changes recorded in sequentially laminated lacustrine sediments of the Nahe palaeolake in Schleswig-Holstein, Germany. Rev Palaeobot Palynol 280:104271
Lamb HH (1995) Climate, history and the modern world, 2nd edn. Routledge, London
Little DJ, Mitchell FJG, von Engelbrechten S, Farrell EP (1996) Assessment of the impact of past disturbance and prehistoric Pinus sylvestris on vegetation dynamics and soil development in Uragh Wood, SW Ireland. Holocene 6:90–99
Lockhart N, Hodgetts N, Holyoak DT (2012) Rare and threatened bryophytes of Ireland. National Museums Northern Ireland, Holywood (Co. Down)
Lowe JJ, Rasmussen SO, Björck S, Hoek WZ, Steffensen JP, Walker MJC, Yu ZC (2008) Synchronisation of palaeoenvironmental events in the North Atlantic region during the Last Termination: a revised protocol recommended by the INTIMATE group. Quat Sci Rev 27:6–17
Lupton D, Sheehy Skeffington M (2020) A review of the ecology and status of the Kerry Lily Simethis mattiazzii (S. planifolia) Asphodelaceae in Ireland. Brit Irish Bot 2:309–334
Lynch A (1981) Man and environment in south-west Ireland, 4000 B.C.–A.D. 800. A study of man’s impact on the development of soil and vegetation. BAR British Series 85. British Archaeological Reports, Oxford
MacCotter P (2004) Lordship and colony in Anglo-Norman Kerry, 1177–1400. J Kerry Archaeol Hist Soc 4:39–85
McCormick M, Büntgen U, Cane MA et al. (2012) Climate change during and after the Roman Empire: reconstructing the past from scientific and historical evidence. J Interdiscipl Hist 43:169–220
McKeown SA (1994) The analysis of wood remains from Mine 3, Mount Gabriel. In: O’Brien W (ed) Mount Gabriel. Bronze Age copper mining in Ireland. (Bronze Age Studies 3).
Mighall TM, Lageard JGA, Chambers FM, Field MH, Mahi P (2004) Mineral deficiency and the presence of Pinus sylvestris on mires during the mid- to late Holocene: palaeoecological data from Cadogan’s Bog, Mizen Peninsula, Co. Cork, southwest Ireland. Holocene 14:95–109
Mighall TM, Timpany S, Blackford JJ, Innes JB, O’Brien CE, O’Brien W, Harrison S (2008) Vegetation change during the Mesolithic and Neolithic on the Mizen Peninsula, Co. Cork, south-west Ireland. Veget Hist Archaeobot 17:617–628
Mitchell GF (1956) Post-Boreal pollen-diagrams from Irish raised-bogs. Proc R Ir Acad, Sect B 57:185–251
Mitchell GF (1981) Littleton Bog, Tipperary: an Irish Lateglacial birch copse. Striae 14:76–78
Mitchell FJG (1988) The vegetational history of the Killarney oakwoods, SW Ireland: evidence from fine spatial resolution pollen analysis. J Ecol 76:415–436
Mitchell FJG (1990) The history and vegetation dynamics of a yew wood (Taxus baccata L.) in S.W. Ireland. New Phytol 115:573–577
Mitchell FJG (1993) The biogeographical implications of the distribution and history of the strawberry tree, Arbutus unedo, in Ireland. In: Costello MJ, Kelly KS (eds) Biogeography of Ireland: past, present and future. Irish Biogeographical Society. Occasional Publication 2. The Irish Biogeographical Society, Dublin, pp 35–44
Mitchell FJG, Cooney T (2004) Vegetation history in the Killarney valley. In: O’Brien W (ed) Ross Island. Mining, metal and society in early Ireland. (Bronze Age Studies 6). Department of Archaeology, National University of Ireland (NUI), Galway, pp 481–493
Mitchell FJG, Cooney T (2019) Sheheree Bog pollen dataset (Version 1.0) . Neotoma Paleoecological Database. https://doi.org/10.21233/PS1X-ES22. Accessed 1 May 2023
Molloy K, O’Connell M (2004) Holocene vegetation and land-use dynamics in the karstic environment of Inis Oírr, Aran Islands, western Ireland: pollen analytical evidence evaluated in light of the archaeological record. Quat Int 113:41–64
Molloy K, O’Connell M (2014) Post-glaciation plant colonisation of Ireland: fresh insights from An Loch Mór, Inis Oírr, western Ireland. In: Sleeman DP, Carlsson J, Carlsson JEL (eds) Mind the Gap II: new insights into the Irish postglacial. Irish Naturalists’ Journal, Belfast, pp 66–88
Moore PD, Webb JA, Collinson ME (1991) An illustrated guide to pollen analysis, 2nd edn. Blackwell Scientific, London
Murphy C (2009) Ancient field walls in the Beara peninsula. In: O’Brien W (ed) Local worlds. Early settlement landscapes and upland farming in south-west Ireland. Collins Press, Cork, pp 371–392
Nowaczyk NR (2001) Logging of magnetic susceptibility. In: Last WM, Smol JP (eds) Tracking environmental changes using lake sediments, volume 1. Basin analysis, coring, and chronological techniques. Springer, Dordrecht, pp 155–170
O’Brien W (1994) Mount Gabriel. Bronze Age copper mining in Ireland. (Bronze Age Studies 3). Galway University Press, Galway
O’Brien W (1999) Sacred ground. Megalithic tombs in coastal south-west Ireland. (Bronze Age Studies 4). Department of Archaeology, National University of Ireland (NUI), Galway
O’Brien W (2003) The Bronze Age copper mines of the Goleen area, Co. Cork. Proc R Ir Acad, Sect C 103:13–59
O’Brien W (2004) Ross Island. Mining, metal and society in early Ireland. (Bronze Age Studies 6). Department of Archaeology, National University of Ireland (NUI), Galway
O’Brien W (2009) Local worlds. Early settlement landscapes and upland farming in south-west Ireland. The Collins Press, Cork
O’Brien W (2012) The Chalcolithic in Ireland: a chronological and cultural framework. In: Allen MJ, Gardiner J, Sheridan A (eds) Is there a British Chalcolithic? People, place and polity in the later 3rd millennium. The Prehistoric Society and Oxbow Books, Oxford, pp 211–225
O’Brien W (2015) Prehistoric copper mining in Europe: 5500–500 BC. Oxford University Press, Oxford
O’Brien W (2022) Derrycarhoon. A later Bronze Age copper mine in south-west Ireland. BAR International Series S3059. BAR Publishing, Oxford
O’Connell M (1987) Early cereal-type pollen records from Connemara, western Ireland and their possible significance. Pollen Spores 29:207–223
O’Connell M (2021) Post-glacial vegetation and landscape change in upland Ireland with particular reference to Mám Éan, Connemara. Rev Palaeobot Palynol 290:104377
O’Connell M (2022) Bog-deal in Co. Clare, with particular reference to bog-pine and its significance. The Other Clare (J Shannon Archaeol Soc) 46:9–17
O’Connell M, Molloy K (2017) Mid- and late-Holocene environmental change in western Ireland: new evidence from coastal peats and fossil timbers with particular reference to relative sea-level change. Holocene 27:1825–1845
O’Connell M, Huang CC, Eicher U (1999) Multidisciplinary investigations, including stable-isotope studies, of thick Late-glacial sediments from Tory Hill, Co., Limerick, western Ireland. Palaeogeogr Palaeoclimatol Palaeoecol 147:169–208
O’Connell M, Ghilardi B, Morrison L (2014) A 7000-year record of environmental change, including early farming impact, based on lake-sediment geochemistry and pollen data from County Sligo, western Ireland. Quat Res 81:35–49
O’Connell M, Molloy K, Jennings E (2020) Long-term human impact and environmental change in mid-western Ireland, with particular reference to Céide Fields – an overview. E&G Quat Sci J 69:1–32
O’Connell M, Jennings E, Molloy K (2021) Holocene vegetation dynamics, landscape change and human impact in western Ireland as revealed by multidisciplinary, palaeoecological investigations of peat deposits and bog-pine in lowland Connemara. Geographies 1:251–291
Overland A, Hjelle KL (2009) From forest to open pastures and fields: cultural landscape development in western Norway inferred from two pollen records representing different spatial scales of vegetation. Veget Hist Archaeobot 18:459–476
Overland A, O’Connell M (2008) Fine-spatial paleoecological investigations towards reconstructing late Holocene environmental change, landscape evolution and farming activity in Barrees, Beara Peninsula, southwestern Ireland. J North Atlantic 1:37–73
Overland A, O’Connell M (2009) Palaeoecological investigations in the Barrees valley. In: O’Brien W (ed) Local worlds. Early settlement landscapes and upland farming in south-west Ireland. Collins Press, Cork, pp 285–322
Overland A (2007) Palaeoecological investigations of lake sediments and peats towards reconstruction of long-term environmental change at Barrees, Beara Peninsula, south-west Cork. Ph.D. thesis (unpublished), Palaeoenvironmental Research Unit, Department of Botany, NUI Galway, Galway
Parker AG, Goudie AS, Anderson DE, Robinson MA, Bonsall C (2002) A review of the mid-Holocene elm decline in the British Isles. Prog Phys Geogr 26:1–45
Parnell J, Curtis T (2012) Webb’s an Irish flora, 8th edn. Cork University Press, Cork
Plunkett G, Brown DM, Swindles GT (2020) Siccitas magna ultra modum: examining the occurrence and societal impact of droughts in prehistoric Ireland. Proc R Ir Acad, Sect C 120:83–104
Pracht (1997) Geology of Kerry–Cork [booklet to accompany GSI Sheet 21, Kerry–Cork geology map]. Geological Survey of Ireland, Dublin
Pracht M, Sleeman AG (2002) Geology of west Cork [booklet to accompany GSI Sheet 24, west Cork geology map]. Geological Survey of Ireland, Dublin
Ratcliffe DA, Birks HJB, Birks HH (1993) The ecology and conservation of the Killarney fern Trichomanes speciosum Willd. in Britain and Ireland. Biol Conserv 66:231–247
Reille M (1992) Pollen et spores d’Europe et d’Afrique du nord. Laboratoire de Botanique Historique et Palynologie, Faculté des Sciences et Techniques Saint-Jérôme, Marseille
Reille M (1995) Pollen et spores d’Europe et d’Afrique du nord, Suppl 1. Laboratoire de Botanique Historique et Palynologie, Faculté des Sciences et Techniques Saint-Jérôme, Marseille
Reimer PJ, Austin WEN, Bard E et al. (2020) The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP) Radiocarbon 62:725–757
Richards PW, Evans GB (1972) Biological flora of the British Isles. Hymenophyllum. J Ecol 44:245–268
Roberts BW, Uckelmann M, Brandherm D (2013) Old father time: the Bronze Age chronology of western Europe. In: Fokkens H, Harding A (eds) The Oxford handbook of the European Bronze Age. Oxford University Press, Oxford, pp 18–46
Santiso X, Lopez L, Retuerto R, Barreiro R (2016) Phylogeography of a widespread species: pre-glacial vicariance, refugia, occasional blocking straits and long-distance migrations. AoB PLANTS 8:plw003
Schoch WH, Pawlik B, Schweingruber FH (1988) Botanical macro-remains. Paul Haupt, Berne
Scott N, Sheehy Skeffington M (2022) The strawberry tree and how it may have reached Ireland. Brit Wildlife 34:85–93
Sheehy Skeffington M, Scott N (2021) Is the Strawberry Tree, Arbutus unedo (Ericaceae), native to Ireland, or was it brought by the first copper miners? Brit Ir Bot 3:385–418
Sheehy Skeffington M, Scott N (2022) The strawberry tree in Ireland. Could botanical research help explain how Bronze Age culture came to Ireland? Archaeol Ireland 36:24–29
Shumilovskikh LS, Shumilovskikh ES, Schlütz F, van Geel B (2022) NPP-ID: Non-Pollen Palynomorph Image Database as a research and educational platform. Veget Hist Archaeobot 31:323–328
Shumilovskikh LS, Shumilovskikh ES, Schlütz F, van Geel B (2023) Correction: NPP-ID: Non-Pollen Palynomorph Image Database as a research and educational platform. Veget Hist Archaeobot 32:109
Smith AJE (1978) The moss flora of Britain and Ireland, 1st edn. Cambridge University Press, Cambridge
Stolze S, Monecke T (2017) Holocene history of ‘non-native’ trees in Ireland. Rev Palaeobot Palynol 244:347–355
Stroh PA, Walker KJ, Humphrey TA, Pescott OL, Burkmar RJ (2023) Plant atlas 2020: mapping changes in the distribution of the British and Irish flora. Princeton University Press/Botanical Society of Britain and Ireland (BSBI), Princeton, New Jersey
Thomas ER, Wolff EW, Mulvaney R et al. (2007) The 8.2ka event from Greenland ice cores. Quat Sci Rev 26:70–81
Trautmann W (1953) Zur Unterscheidung fossiler Spaltöffnungen der mitteleuropäischen Coniferen. Flora 140:523–533
Troels-Smith J (1955) Karakterisering af løse jordarter (Characterization of unconsolidated sediments). Danmarks Geologiske Undersøgelse, IV Række, bd 3, nr 10. Reitzel, København
Van Asch N, Lutz AF, Duijkers MCH, Heiri O, Brooks SJ, Hoek WZ (2012) Rapid climate change during the Weichselian Lateglacial in Ireland: chironomid-inferred summer temperatures from Fiddaun, Co. Galway. Palaeogeogr Palaeoclimatol Palaeoecol 315–316:1–11
Van Geel B (1978) A palaeoecological study of Holocene peat bog sections in Germany and The Netherlands, based on the analysis of pollen, spores and macro- and microscopic remains of fungi, algae, cormophytes and animals. Rev Palaeobot Palynol 25:1–120
Van Rijn P (2004) Analysis of charcoal from Ross island. In: O’Brien W (ed) Ross Island. Mining, Metal and Society in Early Ireland. (Bronze Age Studies 6). Department of Archaeology, National University of Ireland (NUI), Galway, pp 386–401
Van Rijn P, Vorst Y (2009) Charcoal from archaeological excavations in the Barrees valley. In: O’Brien W (ed) Local worlds. Early settlement landscapes and upland farming in south-west Ireland. Collins Press, Cork, pp 367–370
Vokes E (1966) Late-glacial and post-glacial vegetation of Killarney, Co. Kerry. M.Sc. thesis (unpublished), University of Dublin (TCD), Dublin
Waddell J (2022) The prehistoric archaeology of Ireland, 4th edn. Wordwell, Dublin
Walker MJC, Coope GR, Lowe JJ (1993) The Devensian (Weichselian) Lateglacial palaeoenvironmental record from Gransmoor, East Yorkshire, England. Quat Sci Rev 12:659–680
Walker MJC, Bohncke SJP, Coope GR, O’Connell M, Usinger H, Verbruggen C (1994) The Devensian/Weichselian Late-glacial in northwest Europe (Ireland, Britain, north Belgium, The Netherlands, northwest Germany). J Quat Sci 9:109–118
Walker M, Head MJ, Lowe J et al. (2019) Subdividing the Holocene Series/Epoch: formalization of stages/ages and subseries/subepochs, and designation of GSSPs and auxiliary stratotypes. J Quat Sci 34:173–186
Wanner H, Solomina O, Grosjean M, Ritz SP, Jetel M (2011) Structure and origin of Holocene cold events. Quat Sci Rev 30:3109–3123
Watson EV (1981) British mosses and liverworts, 3rd edn. Cambridge University Press, Cambridge
Watson JE, Brooks SJ, Whitehouse NJ, Reimer PJ, Birks HJB, Turney C (2010) Chironomid-inferred late-glacial summer air temperatures from Lough Nadourcan, Co. Donegal, Ireland. J Quat Sci 25:1200-1210
Watts WA (1984) The Holocene vegetation of the Burren, western Ireland. In: Haworth EY, Lund JWG (eds) Lake sediments and environmental history. Leicester University Press, Leicester, pp 359–376
Webb DA (1983) The flora of Ireland in its European context (The Boyle Medal Discourse, 1982). J Life Sci (R Dublin Soc) 4:143–160
Welten M (1952) Pollenanalytische Stichproben über die subrezente Vegetationsentwicklung im Bergland von Kerry (Irland). In: Lüdi W (ed) Die Pflanzenwelt Irlands. Hans Huber, Bern, pp 85–99
Acknowledgements
Patrick O’Rafferty, Ann Bingham and Marion Camy-Palau (Ecole Nationale d'Ingénieurs des Travaux Agricoles de Clermont-Ferrand) assisted with coring Loch Beag. Local landowners, Jerome and Rose Ann Harrington, and John Joe Sullivan, granted access to their lands. Áine McCann carried out the magnetic susceptibility measurements on cores BEG1 and BEG2. A. Overland was financially supported by a Postgraduate Fellowship from NUI Galway, now the University of Galway. T.M. Mighall (Geosciences, University of Aberdeen) provided raw pollen counts for profiles CD and MG4 (Cadogan’s Bog and Mount Gabriel). F.J.G. Mitchell and T. Cooney (Botany, TCD) made the raw counts for Sheheree bog available in Neotoma. B. McConnell (GSI) provided details of a 14C date from a pine stump at Shanvallycahill, Co. Mayo. Two anonymous referees gave helpful comments on the manuscript. We thank the above for the help and especially those who made pollen data available to us. We also gratefully acknowledge the financial support that we received towards 14C dates and fieldwork from The Heritage Council (2005 Archaeology Grant Scheme). Finally, we thank W. O’Brien (Archaeology, NUI Galway; now Professor of Archaeology, UCC) for introducing us to Barrees, and for many helpful discussions during fieldwork and subsequently.
Funding
Open Access funding provided by the IReL Consortium. This research was funded by a University of Galway, Postgraduate Fellowship to A. Overland, and a Heritage Council grant (2005 Archaeological Grant Scheme) towards 14C dates to A. Overland and M. O’Connell
Author information
Authors and Affiliations
Contributions
MO’C and AO planned the research and carried out coring with assistance from others at Loch Beag. AO carried out the analyses of the lake cores and wrote-up the results as part of her PhD research at NUI Galway. The present manuscript was initially drafted by MO’C; subsequently, AO made inputs to all part of the manuscript. Both authors have read, and given their approval to, the final version.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of, or views expressed in, this article.
Additional information
Communicated by F. Bittmann.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
O’Connell, M., Overland, A. Holocene vegetation and flora dynamics of the west Cork/Kerry region, south-western Ireland. Veget Hist Archaeobot (2024). https://doi.org/10.1007/s00334-024-00998-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00334-024-00998-6