
Abstract
Drones, or unoccupied aircraft systems (UAS), can transform the way scientific information on wildlife populations is collected. UAS surveys produce accurate estimates of ground-nesting seabirds and a variety of waterbirds, but few studies have examined the trade-offs of this methodology for counting cliff-nesting seabirds. In this study, we examined how different UAS survey parameters might influence seabird counts for population monitoring and assessed behavioral responses to aerial surveys for three sub-Arctic seabird taxa in the Gulf of Alaska: common murres (Uria aalge), black-legged kittiwakes (Rissa tridactyla), and pelagic and double-crested cormorants (Phalacrocorax pelagicus and Phalacrocorax auritus). We flew two commercially available models of UAS in planned approaches at different speeds and distances from colonies during incubation and chick-rearing periods. We compared counts from UAS-derived images with those from vessel-based photography and assessed video recordings of individual birds’ behaviors for evidence of disturbance during UAS operations and control phases. Count estimates from UAS images were similar to or higher than those from conventional vessel-based images, and UAS were particularly effective at photographing birds at sites with high cliff walls or complex topography. We observed no significant behavioral responses to the UAS by murres or cormorants, but we did observe flushing by black-legged kittiwakes during UAS flights; most of these birds were not incubating or brooding. At both the colony and individual level, we observed slightly greater responses to the smaller UAS platform and closer approaches. These results inform both species specific and general best practices for research and recreational usage of UAS near cliff-nesting seabird colonies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Scientists have embraced the use of drones, or unoccupied aerial systems (UAS), to study wildlife as a potentially less invasive, cost-effective alternative to some conventional survey techniques (Anderson and Gaston 2013; Christie et al. 2016). In the past 10 years, there has been a dramatic increase in the use of UAS to monitor population trends, assess spatial habitat use patterns, and to inform management decisions related to endangered or harvested species (Mustafa et al. 2018; Schofield et al. 2019; Johnston 2019; Ridge and Johnston 2020; Fust and Loos 2020; Corcoran et al. 2021; Larsen et al. 2022). The growing use of UAS in wildlife science has required validation studies to test whether this technology can safely and effectively supplement or replace existing techniques. These include assessments of abundance estimates (Lyons et al. 2019; Hayes et al. 2021; McMahon 2021), as well as assessments of wildlife sensitivity to UAS platforms and best practices (Bennitt et al. 2019; Weston et al. 2020). For many species, UAS represent a new form of aerial disturbance that can elicit as-yet uncharacterized species-specific responses to different types of exposure (Smith et al. 2016). UAS appear to cause less disturbance than conventional aircraft or ground-based surveys for a range of species (Moreland et al. 2015; Borrelle and Fletcher 2017; Sweeney et al. 2015; Krause et al. 2021), but in some cases UAS appear to elicit stress behaviors or physiological responses that may impact breeding success and reduce fitness (Grémillet et al. 2012; Ditmer et al. 2015; McEvoy et al. 2016; Smith et al. 2016; Vas et al. 2015; Weimerskirch et al. 2018; Rush et al. 2018). Furthermore, many incidents of wildlife harassment by recreational UAS users have already been documented (Rebolo-Ifrán et al. 2019).
For a variety of waterbirds and ground-nesting seabirds, studies have documented the use and trade-offs of UAS technology as a survey tool (Barnas et al. 2018; Reintsma et al. 2018; Weimerskirch et al. 2018; Magness et al. 2019; Renner et al. 2021; Krause et al. 2021). For example, comparisons between UAS and ground surveys of large bird colonies suggest that UAS counts are accurate and often more time- or cost-effective (Dunn et al. 2021; Renner et al. 2021). Behavioral responses from ground-nesting bird colonies varied from minimal observed responses (Sardà-Palomera et al. 2017; Rush et al. 2018; Magness et al. 2019; Barr et al. 2020) to increased agitation with closer approaches from the UAS (Rümmler et al. 2016; Valle and Scarton 2021; Krause et al. 2021), with fewer responses among colonial species (Weimerskirch et al. 2018).
Few studies to date have validated this technology as a counting tool for cliff-nesting seabirds (e.g., Brisson-Curadeau et al. 2017). Surveys of cliff-nesting seabirds warrant a few key considerations that distinguish them from waterbirds and ground-nesting bird colonies. For ground-nesting birds and waterfowl, aerial surveys are common and can reliably use nadir camera angles to photograph and count an entire colony, providing productivity estimates, indices for monitoring trends, and total population abundances (Dunn et al. 2021; Hayes et al. 2021). In contrast, conventional surveys of cliff-nesting seabirds have largely been conducted from boats or from overlooking or adjacent cliffs (Bailey 1978; Byrd et al. 2008). Due to the complex topography of these sites and limited vantage points, views of portions of the colony may be impaired using these approaches; however, this is often addressed by counting index plots of “viewable” sections of the cliff, which are assumed to be representative abundance estimates for monitoring trends (Byrd et al. 2008). Aerial surveys, specifically UAS surveys, for cliff-nesting seabirds may increase the viewable portions of the cliff for index counts or enable opportunities to quantify total population abundances from entire census counts. However, in contrast to ground-nesting birds and waterfowl, this may require additional customized flight paths, flight parameters, and UAS model selection to optimize coverage of the complex vertical topography. The effects of these modifications in UAS survey parameters (e.g., UAS size, speed), and variation in responses across different stages of the breeding season (Krause et al. 2021), are still poorly understood for most bird species.
Here, we focused on three taxa of cliff-nesting seabirds that are common in the Gulf of Alaska and Bering Sea and are widely monitored. Black-legged kittiwakes (Rissa tridactyla), common murres (Uria aalge), and pelagic and double-crested cormorants (Phalacrocorax pelagicus and Phalacrocorax auratus) spend the majority of the year at sea, but nest on coastal cliffs in dense colonies during the summer breeding season. We assessed counts of cliff-nesting seabirds derived from UAS images, compared these to estimates from conventional vessel-based photography, and examined how the abundance estimates and behavioral responses varied across UAS platforms, breeding stages, and flight conditions.
Methods
Study site
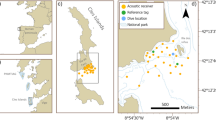
The Alaska Maritime National Wildlife Refuge (Refuge) comprises thousands of sub-Arctic islands that contain breeding habitat for an estimated 40 million seabirds. This study took place in the Gulf of Alaska Unit of the Refuge. We surveyed black-legged kittiwakes and common murres at the Beehive Islands in the Chiswell Island group (Fig. 1a). The Beehive Islands consist of two small dome-shaped islands: Beehive A (59.61° N, −149.61° W) and Beehive B (59.61° N, −149.60° W). Cliff-nesting seabird colonies on these islands are predominantly on the southwest coastlines, at < 10 to > 100 m above sea level (Fig. 1c, d). The Refuge conducts boat-based counts to monitor four distinct cliff segments (A-D) of black-legged kittiwakes on Beehive A and one on Beehive B. Common murres form several large aggregations on Beehive B, three of which were delineated as plots for this study. We also selected a mixed colony of pelagic cormorants and double-crested cormorants (henceforth grouped as “cormorants”) outside of the Refuge, on the eastern coast of Cape Resurrection (59.88° N, −149.26° W, Fig. 1a, b). We surveyed black-legged kittiwakes and common murres on June 15–21, 2021, during their incubation period, and again on July 31–August 6, 2021, during the chick-rearing period. We surveyed cormorants only during the chick-rearing period from July 31 to August 6, 2021.

Locations of study regions and surveys flown. A local map (a) and a regional map (an inset) situate study sites (red, b–d) within the Gulf of Alaska. Drone surveys were flow along pre-programmed flight routes at 30 m (dashed yellow lines) or 60 m (dotted yellow lines) horizontal proximity to cliff-nesting seabirds. Focal nesting sites (light blue lines) consisted of cormorants at Cape Resurrection (b), black-legged kittiwakes at Beehive A (c), or black-legged kittiwakes and common murres at Beehive B (d). Background imagery features an ocean basemap (Esri, GEBCO, DeLorme, NaturalVue, a) or satellite imagery of the sites (Vivid, Maxar, b–d)
Survey design
All observations and surveys were conducted under Alaska Department of Fish and Game scientific research permits 21-106 and 21-106-A1 and the University of Alaska Fairbanks IACUC animal use protocol 1700151-3. Vessel-based photographs of colony plots were taken using a Canon EOS 40D DSLR camera with a 70–300-mm lens. Photographs of each plot were taken twice per survey day: at least 20 min before UAS flights and after completion of all UAS surveys. Images were taken from the boat deck, while the vessel maintained a horizontal distance of approximately 100 m from the center of each plot.
We flew two DJI (Da-Jiang Innovation, Shenzhen, Guangdong, China) models of UAS for the aerial surveys: the larger Inspire 2 (diagonal: 60.5 cm excluding propellers; 3.3 kg) equipped with a gimbal-stabilized Zenmuse X5S camera (M4/3″ CMOS sensor, 20.8 MP) and the smaller Mavic Air (diagonal: 21.3 cm; 0.43 kg) with integrated camera (1/2.3″ CMOS sensor, 12 MP). These quadcopter models were selected to represent large and small commercially available UAS, respectively, and for their ease of operation, high-quality cameras, and affordable prices, which make them accessible to scientists and recreational users. Flight plans were designed by generating waypoints at set distances away from the colony in ArcGIS Pro 2.6.1 and were uploaded to UAS and piloted using DJI Ground Station Pro on a mini tablet. Each flight plan was assigned a maximum speed that was slow (2 m/s) or fast (8 m/s) and a distance from the colony that was close (30 m horizontal buffer, 45 m altitude) or far (60 m horizontal buffer, 75 m altitude). To assess fine-scale behavioral responses and collect abundance data while preventing potential adverse effects on fitness, productivity, or chick-survival, we planned flight routes and schedules within precautionary limits to avoid or minimize flushing breeding birds. Adverse effects among other bird species have been observed when UAS were flown directly over the birds at altitudes ≤ 30 m (Brisson-Curadeau et al. 2017; Barnas et al. 2018; Reintsma et al. 2018; Weimerskirch et al. 2018), so we selected 30 m as our closest horizontal distance from the cliffs. We flew every combination of these three parameters—UAS model, maximum speed, and distance from colony—at each colony in a randomized sequence, for a total of eight surveys at each site during each of the two periods.
UAS were flown under a combination of manual piloting and automated flights along pre-programmed routes with oversight by the UAS pilot. UAS were launched from the boat under manual control after which the pilot initiated the pre-programmed flight plan with the selected parameters of maximum speed (slow or fast) and distance (close or far; Fig. 1). Cameras were programmed to take photographs every 2 s throughout each flight. During the automated survey portion of the flight, the UAS platform remained in motion, accelerating toward its maximum speed between waypoints and decelerating to waypoints, without stopping to hover. The UAS operator actively controlled the camera’s pitch and yaw to try to maximize coverage of focal plots in each photosequence. Camera settings varied between surveys and were recorded in the metadata of each photograph. Following mission completion, the UAS was manually flown back to the vessel. To minimize potential cumulative effects of UAS exposure on bird behavior at each colony, consecutive flights within a day were spaced apart by a minimum of 40 min, with a maximum exposure of 4 flights per colony per day. Additionally, survey days were spaced apart by at least 3 days.
UAS survey assessments consisted of three monitoring phases for analysis: 10 min before take-off (PRE), the duration of each UAS flight, including take-off, approach to the survey route, survey route, return to vessel, and landing (DURING) and 10 min after landing (POST). The duration of the DURING phase varied depending on site and flight parameters (range: 3–15 min). The distance from the vessel to the colony during UAS operations ranged 122–889 m.
Abundance estimates
We evaluated UAS- and vessel-based images of plots, and for each set we selected the image with the greatest clarity for conducting counts (Online Resource 1). Taxa were identified by location, size, shape, and color of individuals. Initially, a representative UAS and boat survey photo from each plot was counted independently by two experienced observers to assess inter-observer variances. In each plot image, observers manually counted all birds, regardless of their incubating/brooding behavior, on a desktop computer using ArcGIS Pro. Individual birds were marked with points to avoid double-counting, and total counts were derived from the shapefile attributes. Additional images along the survey path were occasionally used to assist in resolving cases of uncertainty of a plot’s count due to UAS positioning, image quality, or topography. In cases where agreement between observers varied by > 20%, re-evaluation of the images revealed that the discrepancies were the result of unclear plot boundaries. Following clarification, agreement between observers’ counts was consistent (range: 2–10%), and the remainder of the images were assigned randomly to one of the two observers and counts were conducted as described above.
We used mixed-effects Poisson models (O’Hara and Kotze 2010) to test whether the vessel-based and UAS counts were different. Fixed effects included survey type (boat or UAS) and Plot ID was included as a random effect to account for the spatial arrangement of plots within each study site. Because our cormorant surveys only included one plot, we used a generalized linear model with a Poisson distribution. We used Levene’s tests to assess the equality of variances between the boat and UAS counts. We paired each count from a UAS image with a boat-based image count that was closest in time to calculate the difference (count difference) between methods. If the count difference was positive, there were more birds counted in the UAS images than in the conventional boat counts. If the count difference was negative, there were fewer birds counted from the UAS image than by the conventional boat counts. If count difference was zero or near zero, the counts were similar. We examined three linear mixed-effects models per taxa to test whether the magnitude of the count difference was associated with (1) UAS model, (2) maximum speed of survey, and (3) distance to plot. Plot ID was included as a random effect. Survey design limitations precluded us from exploring interaction terms in these models.
Behavioral responses
At each plot, a group of 12–20 birds were video recorded from the vessel using a Panasonic HC-V180 HD 90 × camera, during each of the survey phases. Groups were selected to be representative of the plot composition: for black-legged kittiwakes we chose 12–20 adjacent nests and for cormorants and common murres, clusters of 12–20 individuals were selected. We also recorded the time of any colony-level responses (> 10 birds flushing), noting whether the birds included attending (e.g., incubating or brooding) or only non-attending individuals, the general location of the flush in the colony relative to the UAS; and duration of the flush. For analysis of individual-level behavioral responses, we generated an ethogram (Table 1) to characterize mutually exclusive and broad behavioral categories, including response behaviors that may indicate disturbance by UAS exposure. A primary observer was trained to identify behaviors using example footage and consultation with experts. Video analysis consisted of instantaneous scan sampling (Altmann 1974) at 10-s intervals, maintaining a consistent order in which each individual bird’s behaviors were recorded and yielding 23–35 min of observation per bird per UAS flight. This interval was selected to maximize our capture of short-duration behaviors while balancing processing times (Altmann 1974). Activity budgets were then calculated from the scan samples (60 scans per bird in PRE, 8–90 scans per bird in DURING, 60 scans per bird in POST) to separately quantify the proportion of time each bird spent in the discrete behavioral categories during each of the three monitoring phases.
To test whether the percentage of time spent in a response behavior changed across UAS monitoring phases, we used non-parametric, repeated measures Friedman tests. Prior to analysis, if more than 50% of the observations in a UAS phase were classified as outside the camera view due to camera shift, boating operations, or other technical issue (Table 1) that animal’s activity budget and all associated data of that individual were removed to avoid bias and meet repeated measures assumptions (Shannon et al. 2008; Challender et al. 2012; Bishop et al. 2015). We ran 40 tests, separately assessing whether activity budgets varied across monitoring phases within each combination of taxon, breeding stage, and combination of flight parameters. For any findings of significant results, we used a post hoc Sign test with Bonferroni corrections to examine differences.
RESULTS
Abundance estimates
UAS-based counts were on average higher than vessel-based counts for cormorants ( Z20 = 2.2, p = 0.02) and black-legged kittiwakes (t103 = 2.9, p = 0.004; Table 2). At one site (Plot D) mean kittiwake counts from UAS imagery were approximately 30% higher than those from boat-based imagery (Table 2). On average, there was no significant difference between vessel- and UAS-based counts of common murres (t77 = 0.29, p = 0.77). Variance in counts from both UAS- and boat-based images were similar for black-legged kittiwakes (F = 1.52, p = 0.22), common murres (F = 0.08, p = 0.78), and cormorants (F = 0.001, p = 0.97).
There were no significant differences between counts from the two UAS models for black-legged kittiwakes (χ2 = −0.68, p = 0.50), common murres (χ2 = −1.17, p = 0.23), or cormorants (F = 1.6, p = 0.22). Additionally, we found no significant differences between counts from flights with different max speeds for black-legged kittiwakes (χ2 = 0.97, p = 0.33), common murres (χ2 = −1.04, p = 0.29), or cormorants (F = 0.07, p = 0.79). Common murre and cormorant counts were highest when the UAS surveyed closer to the colony, resulting in the greatest positive count differences (murres: χ2 = 5.7, p = 0.02; cormorants: F = 3.4, p = 0.04, Fig. 2). Counts did not differ significantly between distances for black-legged kittiwakes (χ2 = 3.0, p = 0.08).

Differences in UAS survey counts relative to boat-based counts (count diff) for common murres (Uria aalge) and pelagic and double-crested cormorants (Phalacrocorax pelagicus and Phalacrocorax auratus; cormorants) at defined nesting plots using photography collected at close distances (light green) and far distances (dark green) of 30 m or 60 m horizontal proximity, respectively. Value of zero indicates counts from UAS and vessels were the same, positive values indicate greater counts from UAS, and negative values indicate greater counts from vessel-based images
Colony-wide behavioral responses
Colony-wide behavioral responses varied by taxa and across breeding periods. We observed no flushes among common murres or cormorants. We observed four cases where > 10 attending black-legged kittiwakes flushed during UAS operations (Table 3). In two of these cases, attending birds flushed near the beginning of the survey flight. During incubation, a large group of attending birds (> 50) approximately 300 m away, at the south end of the Beehive A, flushed 4 min after take-off, when the UAS was at the NW start of the survey route for the small UAS flight on a slow and close flight plan. A bald eagle was spotted near the colony before the subsequent survey (approximately 40 min later), which caused flushing behaviors while the UAS was not in operation. The second flush occurred during chick rearing at Beehive A seconds after the larger UAS was launched from the vessel at 185 m from shore. We paused the flight in mid-air at the vessel location until all birds returned to their nests (< 60 s), and no flushing occurred during the remainder of the flight. The other two cases in which attending birds flushed occurred after the survey route was completed, as the UAS was in route back toward the vessel. First, during incubation, after the smaller UAS finished the fast and close survey route, a small flush of 10–20 attending birds occurred near the south end of the Beehive A. The other instance occurred during chick rearing at Beehive B: approximately 50 attending black-legged kittiwakes flushed after the end of the fast and close survey route using the smaller UAS. All birds returned to their nests within 30 s. We also observed flushes of non-attending black-legged kittiwakes, typically among individuals positioned nearest the waterline within the colony, on six occasions during the incubation period (Table 3). In all cases, most birds returned to the colony within 60 s.
Individual behavioral responses
For individual-level behavioral observations, removing cases where individuals were outside of the camera view for the majority of the scan yielded final sample sizes of 8–16 individuals with repeated observations, depending on the flight (Online Resource 2). Behavioral states of flushing (all taxa) and ‘alert head bobbing’ (common murres only) were not observed during scan samples; so only changes in time spent ‘alert’ and ‘out of sight post-flush’ (OSP) were compared across UAS operation phases.
For black-legged kittiwakes, in 6 of the 16 flights significant differences were detected in the percentage of time spent in response behaviors among PRE, DURING, and POST phases (Online Resource 3); however only two of these flights—one in incubation and one during chick rearing—were associated with predicted responses to UAS operations (Figs. 3 and 4). During incubation, we observed that birds spent more time OSP in the DURING (post hoc Sign test, p = 0.013) and POST phases (post hoc Sign test p = 0.013) compared to the PRE phase for the slow and close, small UAS flight (Fig. 3, Friedman test χ2(2) = 21.88, p = < 0.0001, n = 16). Likewise, during chick rearing, birds spent a greater amount of time OSP in the DURING phase than in the PRE (post hoc Sign test, p = 0.003) or POST phases (post hoc Sign test, p = 0.003) for the fast and close, large UAS flight (Fig. 4).

Individual behavioral responses to the small, slow, and close UAS flight (Mavic Air at 2-m/s maximum speed and 30 m horizontal proximity) during incubation. Attending black-legged kittiwakes (Rissa tridactyla) showed significant differences in the percentage of time spent ‘alert’ (top left) and ‘out of sight post-flush’ (top right) during each phase (PRE, DURING, POST) of the UAS survey. In contrast, common murres (Uria aalge) showed no differences in time spent alert (bottom left) or time spent out of sight post-flush when exposed to the same flight parameters. Cormorants are not considered here because they were not surveyed during incubation

Individual behavioral responses to the large, fast, and close UAS flight (Inspire2, at 8 m/s maximum speed and 30 m horizontal proximity) during chick rearing. Attending black-legged kittiwakes (Rissa tridactyla) had no change in the percentage of time alert (left) across survey phases but did spend a greater percentage of time out of sight post-flush (right) DURING the UAS flight relative to the two control periods (PRE, POST)
The other four flights in which there were significant differences in activity budgets for black-legged kittiwakes were not consistent with predicted responses to UAS exposure (Online Resource 4). In one case, birds were more alert in the PRE phase than in the DURING (post hoc Sign test, p = 0.013) or POST phases (post hoc Sign test p = 0.013) for the slow and close small UAS flight during incubation (Fig. 3, Friedman test χ2(2) = 13.62, p = < 0.0011, n = 16). In the other three cases, we observed significant differences in activity budgets for black-legged kittiwakes when comparing the two control phases (e.g., more alert during the PRE than POST phase; Online Resource 4).
Individual behavioral responses for common murres and cormorants were minimal (Online Resource 3; Fig. 3). For cormorants, birds exhibited an increased amount of time OSP in DURING and POST phases relative to PRE phase for the fast and close, large UAS flight (Friedman Test: χ2 = 7.68, p = 0.02, n = 16), but post hoc Sign tests suggested these differences were not significant (p = 0.375 and p = 0.188). The additional differences detected for cormorants and the single significant difference for common murres were all associated with comparisons between controls (PRE and POST phases) and not with UAS operations (DURING phase; Online Resource 4).
Discussion
Effective conservation and management of wildlife depends on the ability to monitor trends in abundance. Given their ability to access remote or dangerous locations, UAS are valuable tools for resource managers to survey wildlife and accurately count populations. This study demonstrates the utility, precision, and replicability of using UAS for seabird counts at cliff-based colonies.
Abundance estimates
Standardized counts of cliff-nesting seabirds have been conducted on the Alaska Maritime National Wildlife Refuge and elsewhere in Alaska since the early 1970’s at a broad network of monitoring sites. Surveyors conduct in-field counts of adults and, when present, nests at viewable sections of the cliff from vessels or ground-based vantage points. Depending on the species, trends are monitored from sub-sampling the colony at predetermined index plots or from a census by full circumnavigation of islands. Our study demonstrates that at sites with high cliff walls or complex topography, UAS-based and vessel-based photography provide comparable assessments of cliff-nesting seabird abundances and in some cases increased viewability. Relative to the images from the vessel, the multicopter UASs maximized available positions and lines of sight to capture photos of colonies in scenarios when viewsheds would otherwise be fully or partially obstructed from views at sea level. For example, Plot D from the black-legged kittiwake surveys comprises complex topography, and counts from UAS images consistently identified more birds at this site than did those from vessel-based photography. For sites near the waterline with overhanging rocks that obscure views of birds from above, vessel-based counts may perform better than UAS counts. Notably, photography collected from UAS provides records of the survey with ancillary data (e.g., nesting habitat, structure, and positions) that is not typically obtained by conventional vessel-based images or counts. Our approach to post hoc processing of images was time consuming, but promising advances in machine learning and computer vision may expedite such processes (McClelland et al. 2016; Rush et al. 2018; Hayes et al. 2021).
The flight parameters of maximum speed and UAS model had minimal impacts on the abundance estimates; however, flights close to colonies (30 m horizontal proximity) yielded significantly higher counts of common murres and cormorants. Compared to the black-legged kittiwakes, these two taxa do not contrast as highly with the background cliff faces (Online Resource 1); therefore, the higher counts may be due to the closer images providing greater details to differentiate individual birds from shadows. These results confirm previous findings that small inexpensive multicopter UAS could accurately survey the number of breeding birds and that flights at closer distances yielded higher abundance counts (Brisson-Curadeau et al. 2017).
We also detected both nests and chicks of black-legged kittiwakes and cormorants in the UAS-derived images. For common murres, we could not confirm absence or presence of chicks due to body positioning. We did not include these data in our analysis because it was difficult to obtain similar images from our vessel-based surveys for statistical comparisons. However, we do believe that this technology can facilitate assessments of nesting success.
Behavioral responses
Behavioral responses to the UAS flights were species and context specific. We found little to no evidence of colony or individual behavioral responses to UAS surveys for common murres and cormorants; however, black-legged kittiwakes exhibited both colony-wide flushing behaviors and individual variation in activity budgets during UAS flights relative to control phases. Kittiwake flushing events occurred most commonly among non-incubating birds that perched close to the waterline. Incubating adults’ fidelity to their nests constrains their behavioral responses (Gilchrist 1999), which likely explains this pattern of non-incubating adults being comparatively more responsive. This result aligns with past studies of reproductive state and wildlife responses to UAS flights (Brisson-Curadeau et al. 2017; Mulero-Pázmány et al. 2017; Weimerskirch et al. 2018). While behaviorally constrained, nesting adults may still exhibit physiological responses to UAS exposure (Weimerskirch et al. 2018; Krause et al. 2021). The population consequences of flushing can be quantified relatively easily, for example, as the number of chicks or eggs lost (Brisson-Curadeau et al. 2017), however, the impacts of non-lethal physiological or fine-scale changes in behaviors, such as time spent alert, still warrants further research.
The Alaska Maritime National Wildlife Refuge protocol specifies that counts are timed to coincide with the period of the nesting season spanning mid-incubation to early chick rearing, when attendance is least variable (Byrd 2006). In a study examining the response of chinstrap penguins (Pygoscelis antarcticus) to UAS flights, nest defense behavior increased as the breeding season progressed and adults actively guarded their chicks from aerial predators (Krause et al. 2021). Similarly, in the present study, birds exhibited less flight behavior during the chick-rearing period. Bald eagles, an aerial predator of black-legged kittiwakes, were not visibly present during our UAS flights, but we observed one agitating and disturbing the black-legged kittiwake colony at Beehive A during the first day of surveys in the incubation period, which may have contributed to the disturbance of non-attending birds during the early surveys. The presence of aerial predators, colony size, and conspecific proximity has all been suggested as potential contributing factors that affect seabird responses to UAS (Mulero-Pázmány et al. 2017; Brisson-Curadeau et al. 2017; Weimerskirch et al. 2018). Together, these studies and our findings emphasize the importance of considering the contextual factors associated with behavioral responses to UAS when developing protocols, timing of surveys, guidelines, and recommendations.
In addition to intrinsic factors impacting birds’ responses to UAS surveys, we explored how disturbance was linked to three flight parameters of interest. Nesting black-legged kittiwakes appeared to flush only during closer surveys (30 m horizontal proximity). However, we are cautious in interpreting this UAS distance as a causal factor because in one case the flushes occurred just after take-off when the UAS was still > 100 m from the island, and in two cases the flushes occurred as the UAS was returning to the vessel. Still, prior work has found that distance of aircraft strongly determines flush responses across a range of species, with higher incidence of response for approaches closer than 30 m (Rümmler et al. 2016; Fuller et al. 2018; Krause et al. 2021; Valle and Scarton 2021) and for vertical approaches relative to horizontal ones (Vas et al. 2015, Rümmler et al. 2016). We selected conservative treatments of flight distance to avoid mass flushing events and based our closest approach on these prior studies as a threshold that minimized disturbances (Barnas et al. 2018; Brisson-Curadeau et al. 2017; Reintsma et al. 2018; Weimerskirch et al. 2018). Therefore, it is unsurprising that we observed minimal effect of flight distance on disturbances for this species.
Our study suggests that the smaller UAS (Mavic Air) elicited a stronger behavioral response than that of the larger (Inspire) model. The Mavic was in flight during 4 of the 6 observed cases of flushing by non-incubating birds and in 1 of the 2 flushes of incubating birds that were consistent with a clear response to UAS. Interestingly, reviews of the use of UAS surveys in wildlife monitoring have suggested that larger UAS models produce greater responses, likely as a function of both perceived risk and detection from visual stimuli (Stankowich and Blumstein 2005; Mulero-Pázmány et al. 2017). UAS noise relative to ambient soundscape may also indicate likely disturbance (Arona et al. 2018), with responses expected only from sounds louder than background noise. However, larger models may allow birds to visually or acoustically detect the UAS from farther away, while smaller platforms like the Mavic Air may approach closer before detection and thus elicit stronger startle effects. This disturbance response is commonly observed for hauled out harbor seals (Phoca vitulina), who respond more strongly to non-motorized vessels than to motorized vessels due to seals being taken by ‘surprise’ by the slow, quiet profiles (Henry & Hammill 2001; Cates and Acevedo-Gutiérrez 2017). Further work testing seabird responses to varying acoustic and visual UAS profiles may help identify what signals may be driving observed behaviors and how to best mitigate any adverse responses.
All of our flights were conducted between 0800 and 1400. There was a slight indication of greater responses to the last two flights of the day. This pattern may be due to diel influence on behaviors and reactivity (Daunt et al. 2002), other environmental factors (e.g., wind direction and speed), or it could indicate a cumulative impact of multiple, consecutive flights. We provided a minimum of 40 min between surveys at a colony, but habituation or sensitivity to repeated UAS exposures requires further investigation. Colonies surveyed in this study were southwest facing, so early morning surveys facilitated photography with less glare, fewer shadows, and lower variation in light levels across the survey region; therefore, while the time-of-day effect on bird behavior remains unclear, there are benefits to planning surveys to maximize optimal light conditions for photography.
Conclusion
We observed minimal disturbance in response to UAS exposure, but highlight variability in bird responses and count estimates depending on species and breeding stage, which managers should consider when designing future UAS flights. For the species investigated in this study, our findings on the UAS models, approach distances, and speeds that maximize counts and maintain little disturbance can be used by managers to develop protocols and robust survey designs that complement or replace conventional vessel-based surveys or enable novel explorations. Specifically, we found that both the Mavic Air and the Inspire 2 provided similar assessments of counts across a range of flight parameters, but the smaller, cheaper Mavic Air was associated with more behavioral responses from the black-legged kittiwakes. Additionally, while closer flights marginally improved counts for common murres and cormorants, they were also associated with more disturbance among black-legged kittiwakes. Flushing can result in egg loss and population-level impacts, so the variability in responses observed here and in other studies suggests that managers considering this technology for novel species or contexts (e.g., colonies with differing sizes or composition) should validate choices of UAS model and flight conditions to achieve high data quality and low animal disturbance before adopting a surveillance protocol. Operators in both recreational and research scenarios should give special attention to in situ conditions, particularly the presence of predators and overall colony behaviors, and should adapt protocols or refrain from surveying when agitation levels are likely elevated.
Finally, additional studies are needed to understand possible cumulative effects of repeated UAS exposure on seabird colonies and other wildlife. Our approach incorporated within- and between-day breaks amid consecutive UAS flights at each colony, but the efficacy of these thresholds at reducing individual- and population-level consequences is unknown. If, in a practical scenario, a single day and flight is used to survey a colony, then cumulative impacts are unlikely to produce population consequences. However, even low-disturbance non-lethal activities, if persistent, can have cumulative effects on some wildlife species (Cecchetti et al. 2018; Mandl 2020; Burnham et al. 2021).
Data availability
The data that support these findings will be publicly available in the DataOne repository following a 2-year embargo (2024). Prior to that time, data are available on request from the corresponding author, AB.
References
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49:227–266
Anderson K, Gaston KJ (2013) Lightweight unmanned aerial vehicles will revolutionize spatial ecology. Front Ecol Environ 11:138–146
Arona L, Dale J, Heaslip SG, Hammill MO, Johnston DW (2018) Assessing the disturbance potential of small unoccupied aircraft systems (UAS) on gray seals (Halichoerus grypus) at breeding colonies in Nova Scotia. Canada Peerj 6:e4467
Bailey EP (1978) Breeding seabird distribution and abundance in the Shumagin Islands, Alaska. Murrelet 59:82–91
Barnas AF, Felege CJ, Rockwell RF, Ellis-Felege SN (2018) A pilot (less) study on the use of an unmanned aircraft system for studying polar bears (Ursus maritimus). Polar Biol 41:1055–1062
Barr JR, Green MC, DeMaso SJ, Hardy TB (2020) Drone surveys do not increase colony-wide flight behaviour at waterbird nesting sites, but sensitivity varies among species. Sci Rep-UK 10:1–10
Bennitt E, Bartlam-Brooks HL, Hubel TY, Wilson AM (2019) Terrestrial mammalian wildlife responses to Unmanned Aerial Systems approaches. Sci Rep-UK 9:1–10
Bishop A, Pomeroy P, Twiss SD (2015) Breeding male grey seals exhibit similar activity budgets across varying exposures to human activity. Mar Ecol Prog Ser 527:247–259
Borrelle SB, Fletcher AT (2017) Will drones reduce investigator disturbance to surface-nesting seabirds? Mar Ornithol 45:89–94
Brisson-Curadeau É, Bird D, Burke C, Fifield DA, Pace P, Sherley RB, Elliott KH (2017) Seabird species vary in behavioural response to drone census. Sci Rep-UK 7:1–9
Burnham RE, Duffus DA, Malcolm CD (2021) Towards an enhanced management of recreational whale watching: The use of ecological and behavioural data to support evidence-based management actions. Biol Conserv 255:109009
Byrd GV (2006) Seabird monitoring on Alaska maritime national wildlife refuge. In: Community-based coastal observing in Alaska: Aleutian life forum, pp. 39–45
Byrd GV, Schmutz JA, Renner HM (2008) Contrasting population trends of piscivorous seabirds in the Pribilof Islands: a 30-year perspective. Deep Sea Res Part II Top Stud Oceanogr 55:1846–1855
Cates K, Acevedo-Gutiérrez A (2017) Harbor seal (Phoca vitulina) tolerance to vessels under different levels of boat traffic. Aquat Mamm 43:193
Cecchetti A, Stockin KA, Gordon J, Azevedo JM (2018) Short-term effects of tourism on the behaviour of common dolphins (Delphinus delphis) in the Azores. J Mar Biol Assoc UK 98:1187–1196
Challender DW, Thai NV, Jones M, May L (2012) Time-budgets and activity patterns of captive sunda pangolins (Manis javanica). Zoo Biol 31:206–218
Christie KS, Gilbert SL, Brown CL, Hatfield M, Hanson L (2016) Unmanned aircraft systems in wildlife research: current and future applications of a transformative technology. Front Ecol Environ. https://doi.org/10.1002/fee.1281
Corcoran E, Winsen M, Sudholz A, Hamilton G (2021) Automated detection of wildlife using drones: synthesis, opportunities and constraints. Methods Ecol Evol 12:1103–1114
Daunt F, Benvenuti S, Harris MP, Dall Antonia L, Elston DA, Wanless S (2002) Foraging strategies of the black-legged kittiwake Rissa tridactyla at a North Sea colony: evidence for a maximum foraging range. Mar Ecol Prog Ser 18:239–247
Ditmer MA, Vincent JB, Werden LK, Tanner JC, Laske TG, Iaizzo PA et al (2015) Bears show a physiological but limited behavioral response to unmanned aerial vehicles. Curr Biol 25:2278–2283
Dunn MJ, Adlard S, Taylor AP, Wood AG, Trathan PN, Ratcliffe N (2021) Un-crewed aerial vehicle population survey of three sympatrically breeding seabird species at Signy Island, South Orkney Islands. Polar Biol 44:717–727
Fuller AR, McChesney GJ, Golightly RT (2018) Aircraft disturbance to Common Murres (Uria aalge) at a breeding colony in central California, USA. Waterbirds 41:257–267
Fust P, Loos J (2020) Development perspectives for the application of autonomous, unmanned aerial systems (UASs) in wildlife conservation. Biol Conserv 241:108380
Grémillet D, Puech W, Garçon V, Boulinier T, Le Maho Y (2012) Robots in ecology: welcome to the machine. Open J Ecol 2:49–57
Gilchrist HG (1999) Declining thick-billed murre Uria lomvia colonies experience higher gull predation rates: an inter-colony comparison. Biol Conserv 87:21–29
Hayes MC, Gray PC, Harris G, Sedgwick WC, Crawford VD, Chazal N, Crofts S, Johnston DW (2021) Drones and deep learning produce accurate and efficient monitoring of large-scale seabird colonies. The Condor. https://doi.org/10.1093/ornithapp/duab022
Henry E, Hammill MO (2001) Impact of small boats Literature Cited on the haul-out activity of harbor seals (Phoca vitulina) in Metis Bay, Saint Lawrence Estuary, Quebec, Canada. Aquat Mamm 27:140–148
Johnston DW (2019) Unoccupied aircraft systems in marine science and conservation. Annu Rev Mar Sci 11:439–463
Krause DJ, Hinke JT, Goebel ME, Perryman WL (2021) Drones minimize antarctic predator responses relative to ground survey methods: an appeal for context in policy advice. Front Mar Sci 8:152
Larsen GD, Seymour AC, Richmond EL, Divine LM, Moreland EE, Newton E, London JM, Johnston DW (2022) Drones reveal spatial patterning of sympatric Alaskan pinniped species and drivers of their local distributions. Drone Sys Appl 10:235–255
Lyons MB, Brandis KJ, Murray NJ, Wilshire JH, McCann JA, Kingsford RT, Callaghan CT (2019) Monitoring large and complex wildlife aggregations with drones. Methods Ecol Evol 10:1024–1035. https://doi.org/10.1111/2041-210X.13194
Magness DR, Eskelin T, Lake M, Renner HM (2019) Evaluation of small unmanned aerial systems as a census tool for Aleutian Tern Onychoprion aleuticus colonies. Mar Ornith 47:11–16
Mandl C (2020) Cumulative Exposure of Sperm Whales to Whale Watching Boats using Spatially-Explicit Capture-Recapture Models. Dissertation, Universität Hamburg
McClelland GT, Bond AL, Sardana A, Glass T (2016) Rapid population estimate of a surface-nesting seabird on a remote island using a low-cost unmanned aerial vehicle. Mar Ornith 44:215–220
McEvoy JF, Hall GP, McDonald PG (2016) Evaluation of unmanned aerial vehicle shape, flight path and camera type for waterfowl surveys: disturbance effects and species recognition. PeerJ 4:e1831. https://doi.org/10.7717/peerj.1831
McMahon MC (2021) Applying unmanned aerial systems (UAS) and thermal infrared technology for the detection and surveying of wild ungulates. Dissertation, University of Minnesota
Moreland EE, Cameron MF, Angliss RP, Boveng PL (2015) Evaluation of a ship-based unoccupied aircraft system (UAS) for surveys of spotted and ribbon seals in the Bering Sea pack ice. J Unmanned Veh Syst 3:114–122
Mulero-Pázmány M, Jenni-Eiermann S, Strebel N, Sattler T, Negro JJ, Tablado Z (2017) Unmanned aircraft systems as a new source of disturbance for wildlife: a systematic review. PLoS ONE 12:e0178448. https://doi.org/10.1371/journal.pone.0178448
Mustafa O, Barbosa A, Krause DJ, Peter HU, Vieira G, Rümmler MC (2018) State of knowledge: Antarctic wildlife response to unmanned aerial systems. Polar Biol 41:2387–2398
O’hara RB, Kotze DJ (2010) Do not log-transform count data. Methods Ecol Evol 1:18–122
Rebolo-Ifrán N, Grilli MG, Lambertucci SA (2019) Drones as a threat to wildlife: YouTube complements science in providing evidence about their effect. Environ Conserv 46:205–210
Reintsma KM, McGowan PC, Callahan C, Collier T, Gray D, Sullivan JD, Prosser DJ (2018) Preliminary evaluation of behavioral response of nesting waterbirds to small unmanned aircraft flight. Waterbirds 41:326–331
Renner H, Renner M, Lyons D, Zykov V, Revyakina Z, Oehlers S (2021) Aleutian Tern Onychoprion aleuticus abundance estimates at four globally significant colonies. Bird Conserv Int. https://doi.org/10.1017/S095927092100037X
Ridge JT, Johnston DW (2020) Unoccupied aircraft systems (UAS) for marine ecosystem restoration. Front Mar Sci 7:438
Rümmler MC, Mustafa O, Maercker J, Peter HU, Esefeld J (2016) Measuring the influence of unmanned aerial vehicles on Adélie penguins. Polar Biol 39:1329–1334
Rush GP, Clarke LE, Stone M, Wood MJ (2018) Can drones count gulls? Minimal disturbance and semiautomated image processing with an unmanned aerial vehicle for colony-nesting seabirds. Ecol Evol 8:12322–12334. https://doi.org/10.1002/ece3.4495
Sardà-Palomera F, Bota G, Padilla N, Brotons L, Sardà F (2017) Unmanned aircraft systems to unravel spatial and temporal factors affecting dynamics of colony formation and nesting success in birds. J Avian Biol 48:1273–1280
Schofield G, Esteban N, Katselidis KA, Hays GC (2019) Drones for research on sea turtles and other marine vertebrates–a review. Biol Conserv 238:108214
Shannon G, Page BR, Mackey RL, Duffy KJ, Slotow R (2008) Activity budgets and sexual segregation in African elephants (Loxodonta africana). J Mammal 89:467–476
Smith CE, Sykora-Bodie ST, Bloodworth B, Pack SM, Spradlin TR, LeBoeuf NR (2016) Assessment of known impacts of unmanned aerial systems (UAS) on marine mammals: data gaps and recommendations for researchers in the United States. J Unmanned Veh Syst 4:31–44
Stankowich T, Blumstein DT (2005) Fear in animals: a meta-analysis and review of risk assessment. Proc Roy Soc B-Biol Sci 272:2627–2634
Sweeney KL, Helker VT, Perryman WL, LeRoi DJ, Fritz LW, Gelatt TS, Angliss RP (2015) Flying beneath the clouds at the edge of the world: using a hexacopter to supplement abundance surveys of Steller sea lions (Eumetopias jubatus) in Alaska. J Unmanned Veh Syst 4:70–81
Vas E, Lescroel A, Duriez O (2015) Approaching birds with drones: first experiments and ethical guidelines. Biol Lett 11:20140754. https://doi.org/10.1098/rsbl.2014.0754
Valle RG, Scarton F (2021) Drone-conducted counts as a tool for the rapid assessment of productivity of Sandwich Terns (Thalasseus sandvicensis). J Ornith 162:621–628
Weimerskirch H, Prudor A, Schull Q (2018) Flights of drones over sub-Antarctic seabirds show species-and status-specific behavioural and physiological responses. Polar Biol 41:259–326
Weston MA, O’Brien C, Kostoglou KN, Symonds MR (2020) Escape responses of terrestrial and aquatic birds to drones: towards a code of practice to minimize disturbance. J Appl Ecol 57:777–785
Acknowledgements
Funding for this project was provided by the North Pacific Research Board (R1818). We thank B. Merkel and an anonymous reviewer for their suggestions and comments, which improved this manuscript. We would like to thank the Seward Marine Center staff and R/V Nanuq Captain Brian Mullaly for their logistical support for fieldwork. Special thanks to J. Dale, D. Johnston, A. Reed, C. Robison, and K. Colson who assisted with or participated in field operations. All activities conducted under ADFG research permit 21-106 and UAF IACUC Research Protocol [1700151-3]. The authors declare there are no competing interests. The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the U.S. Fish and Wildlife Service.
Author information
Authors and Affiliations
Contributions
AB, CB, KC, AK, and HR contributed to project conceptualization. AB, CB, and KC designed the study and acquired funding. AB, CB, GL, and LY participated in data collection; AB and CB led statistical analyses. All authors contributed to writing, reviewing, and editing of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bishop, A.M., Brown, C.L., Christie, K.S. et al. Surveying cliff-nesting seabirds with unoccupied aircraft systems in the Gulf of Alaska. Polar Biol 45, 1703–1714 (2022). https://doi.org/10.1007/s00300-022-03101-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-022-03101-9