
Abstract
Previously we structurally characterized five glucosylated lactose derivatives (F1–F5) with a degree of polymerization (DP) of 3–4 (GL34), products of Lactobacillus reuteri glucansucrases, with lactose and sucrose as substrates. Here, we show that these GL34 compounds are largely resistant to the hydrolytic activities of common carbohydrate-degrading enzymes. Also, the ability of single strains of gut bacteria, bifidobacteria, lactobacilli, and commensal bacteria, to ferment the GL34 compounds was studied. Bifidobacteria clearly grew better on the GL34 mixture than lactobacilli and commensal bacteria. Lactobacilli and the commensal bacteria Escherichia coli Nissle and Bacteroides thetaiotaomicron only degraded the F2 compound α-D-Glcp-(1 → 2)-[β-D-Galp-(1 → 4)-]D-Glcp, constituting around 30% w/w of GL34. Bifidobacteria digested more than one compound from the GL34 mixture, varying with the specific strain tested. Bifidobacterium adolescentis was most effective, completely degrading four of the five GL34 compounds, leaving only one minor constituent. GL34 thus represents a novel oligosaccharide mixture with (potential) synbiotic properties towards B. adolescentis, synthesized from cheap and abundantly available lactose and sucrose.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The human gut microflora has drawn increasing attention in recent years. It constitutes a very interesting ecosystem that varies in density and functionality in the different gut compartments (Ley et al. 2006). These complex ecosystems have a significant impact on host well-being (Flint et al. 2007). Strongest interest is focused on understanding what factors cause variations in microbiota composition and how these gut bacteria modulate host health (Louis et al. 2007). Our work aims to stimulate the growth of health-promoting probiotic gut bacteria by using newly synthesized non-digestible carbohydrates, i.e., prebiotic compounds.
According to the latest definition, a prebiotic is “a substrate that is selectively utilized by host microorganisms conferring a health benefit” (Gibson et al. 2017). Recently, part of the definition was disputed, since selective stimulation of health-promoting species seems not exclusively necessary to confer health benefits (Bindels et al. 2015; Yan et al. 2018). Generally, prebiotics are carbohydrates that are not fully digested by the host. They are fermented by various commensal and health-beneficial gut bacteria, thus promoting their growth and activity which may confer health benefits upon the host (Roberfroid et al. 2010; Callaway et al. 2008). To date, the most well-known prebiotics, supported by good quality data, are human milk oligosaccharides (hMOS) (Bode 2012), β-galacto-oligosaccharides (GOS), β-fructo-oligosaccharides (FOS), inulin, and lactulose (Slavin 2013; Macfarlane et al. 2008). All of these prebiotics are also hydrolyzed by brush border enzymes, but not completely (Ferreira-Lazarte et al. 2017). Isomalto-/malto-polysaccharides (IMMP) (Bai et al. 2016; Rycroft et al. 2001), xylo-oligosaccharides (XOS) (Marquina et al. 2002), resistant starch (Lehmann and Robin 2007), and soy oligosaccharides also are (emerging) prebiotic oligosaccharides (Jaskari et al. 1998), although more data about their effects on gut health are still needed. Each of these prebiotic compounds may exert specific and selective effects on gut bacteria. The search for new and effective prebiotics combined with specific probiotics (synbiotics) is increasing rapidly (Pandey et al. 2015; Rastall and Maitin 2002).
Lactose-derived oligosaccharides attract much attention in view of their prebiotic potential. One example is GOS, which are synthesized from lactose by enzymatic trans-galactosylation using β-galactosidases, achieving a degree of polymerization between 3 and 10 (Van Leeuwen et al. 2016). This prebiotic has been widely studied and shown to stimulate probiotic bacteria to various extents (Macfarlane et al. 2008; Boehm and Moro 2008; Rijnierse et al. 2011). Another commercially available prebiotic in this group is lactosucrose, which is hardly utilized by human digestive enzymes and has stimulatory effects on both lactobacilli and bifidobacteria (Ohkusa et al. 1995; García-Cayuela et al. 2014). Also, the selective bifidogenic effect of 4′-galactosyl-kojibiose, corresponding to compound F2 in our GL34 mixture (Pham et al. 2017), on Bifidobacterium breve 26M2 has been reported (García-Cayuela et al. 2014). These results indicate that there are clear perspectives to further develop and expand this group of lactose-derived prebiotic oligosaccharides.
We recently reported synthesis of a mixture of five novel lactose-derived oligosaccharides (F1–F5) using the Lactobacillus reuteri glucansucrase enzymes Gtf180-ΔN and GtfA-ΔN (Pham et al. 2017). Their structural characterization revealed the presence of various glycosidic linkages, α(1 → 2/3/4), with DP of 3 and 4 (Pham et al. 2017) (Scheme 1). Four out of these five structures were new and only F2 4′-galactosyl-kojibiose had been reported before. In this work, their resistance to degradation by common carbohydrate-degrading enzymes was studied by in vitro incubations. Also, the growth of pure cultures of common gut bacteria, including commensal and probiotic strains, on these novel compounds was evaluated and compared with well-known prebiotic mixtures (GOS and FOS). The GL34 mixture particularly stimulated growth of Bifidobacterium adolescentis. This is also of interest from an industrial perspective, since these new oligosaccharides with very specific prebiotic effects are produced from low-cost lactose and sucrose and may be an option for developing synbiotics.

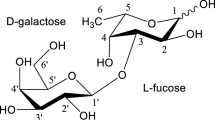
Structures of compounds F1–F5 from the mixture GL34
Materials and methods
Bacterial strains, chemicals, and reagents
Bacteroides thetaiotaomicron VPI-5482, Lactobacillus acidophilus ATCC 4356, B. adolescentis ATCC 15703, and Bifidobacterium longum subsp. infantis ATCC 15697 were purchased from ATCC. B. breve DSM 20213 was obtained from the DSMZ culture collection. L. reuteri strain 121 was obtained from TNO Quality of Life, Zeist, the Netherlands. Lactobacillus casei W56 was provided by Winclove Probiotics B.V. (Amsterdam). All reagents, chemicals, or medium components were purchased from Sigma (Zwijndrecht, Netherlands), or as stated otherwise. The purified GOS mixture TS0903 (lacking glucose, galactose, and lactose) was provided by FrieslandCampina Domo; its detailed GOS composition was published elsewhere (Lammerts van Bueren et al. 2017). The GOS/FOS mixture used is a 90:10 (w/w) mixture of the purified TS0903 GOS and long-chain Inulin (lc-Inulin, Frutafit TEX, provided by SENSUS, Roosendaal, The Netherlands), also serving as a control for the current prebiotic formula added to infant nutrition (Boehm and Moro 2008).
Lactobacillus growth experiments
Lactobacilli were pre-cultured in MRS medium (Oxoid, Basingstoke, UK) anaerobically (or by using the GasPak system (Becton, Dickinson and Company, Sparks, USA)) under an N2 atmosphere for up to 2 days at 37 °C (Daniels and Zeikus 1975). Then, 1-mL samples of the pre-cultures were harvested by centrifugation (2500×g, 2 min). The bacterial pellets were washed twice with sterile 10% NaCl and diluted 25-fold in 2× modified MRS (mMRS; medium that does not contain a carbon source for Lactobacilli) (Watson et al. 2013). In separate tubes, carbohydrates were dissolved with Milli-Q water to 10 mg mL−1 and sterilized by filtration using 0.2-μm cellulose acetate filters (the GL34 mixture) or by autoclaving solutions (glucose). Cultures were inoculated by mixing 1:1 diluted bacterial suspensions with sterilized carbohydrate solutions in sterile microtiter plates to obtain an initial OD600 of 0.01. Inoculation of microtiter plates was carried out in an anaerobic glove box (Sicco, Grünsfeld, Germany) with a constant nitrogen flow, the microtiter plates sealed air tightly and transferred into a plate reader placed under constant N2 flow. Glucose was used as positive control to compare growth of these lactobacilli on the mixture of GL34 compounds. Media without an added carbon source was used as negative control. Bacterial growth was followed at 37 °C by measuring the optical density at 600 nm (OD600 nm) in 5-min intervals. OD values of the negative control samples (no carbohydrate added) were deducted while measuring their corresponding samples and positive controls.
Bifidobacterium and B. thetaiotaomicron growth experiments
The Bifidobacterium strains were sub-cultured (from stocks stored at − 80 °C) in 10 mL of Bifidobacterium medium (BM) supplemented with 1% glucose (1-L BM contained 10 g trypticase peptone, 2.5 g yeast extract, 3 g tryptose, 3 g K2HPO4, 3 g KH2PO4, 2 g triammonium citrate, 0.3 g pyruvic acid, 1 mL Tween 80, 0.574 g MgSO4·7H2O, 0.12 g MnSO4·H2O, and 0.034 g FeSO4·7H2O). After autoclaving, BM was supplemented with 0.05% (w/v) filter-sterilized cysteine-HCl (Ryan et al. 2006). B. thetaiotaomicron was cultured using a carbon-limited minimally defined medium of 100 mM KH2PO4 (pH 7.2), 15 mM NaCl, 8.5 mM (NH4)2SO4, 4 mM L-cysteine, 1.9 M hematin, 200 M L-histidine, 100 nM MgCl2, 1.4 nM FeSO4·7 H2O, 50 M CaCl2, 1 g mL−1 vitamin K3, 5 ng mL−1 vitamin B12, and individual carbon sources (0.5%, w/v) (Koropatkin et al. 2008). Carbohydrates were prepared by dissolving in Milli-Q water to 10 mg mL−1 and sterilized by filtration using 0.2-μm cellulose acetate filters (the GL34 mixture, GOS, FOS) or by autoclaving solutions (lactose). Growth was carried out in fermentation tubes at 37 °C under anaerobic conditions maintained by GasPak EZ anaerobe container system (BD, New Jersey, USA). Cell suspensions from overnight cultures were prepared in 3 mL of BM supplemented with different carbon source in a final concentration of 5 mg mL−1. BM without an added carbon source was used as negative control. OD600 nm measurements of the fermentation tubes were carried out manually at 1-h intervals and data used to generate growth curves. OD values of the negative control samples (no carbohydrate added) were deducted while measuring their corresponding samples and positive controls.
Escherichia coli Nissle growth experiments
Escherichia coli Nissle was cultured using M9 medium as previously described, at 37 °C under aerobic conditions (Sambrook and Russell 2001). OD600 nm measurements of the fermentation tubes were carried out manually at 2-h intervals. OD values of the negative control samples (no carbohydrate added) were deducted while measuring their corresponding samples and positive controls.
Enzyme incubations
The GL34 mixture (1 mg mL−1) was incubated for 24 h with different carbohydrate degrading enzymes (5 U mL−1): α-amylase 1 (porcine pancreas); α-amylase 2 (Aspergillus oryzae); α-glucosidase (yeast); Iso-amylase (Pseudomonas sp.; pullulanase type 1 (Klebsiella planticola); β-galactosidase 1 (A. oryzae) and β-galactosidase 2 (Kluyveromyces lactis) (see Table S1 with detailed information).
Intracellular and extracellular activity essays
After growth with GL34 as their only carbon source, the three tested Bifidobacterium strains were harvested by centrifugation at 10,000×g for 15 min at room temperature. Culture supernatants were sterilized using 0.45-μm filters and concentrated 10 times by Amicon Ultra-4 centrifugal filter units (10,000-Da molecular weight cutoff, Millipore). The harvested cell pellets were washed twice with 0.1 M potassium phosphate buffer (pH 6.6) and then suspended in 1 mL of this buffer into 2.0-mL screw-cap microtubes containing 400 mg of 0.1-mm-diameter glass beads (Biospec Products). Cell disruptions were carried out by homogenization by a mini bead-beater (Biospec Products) at 4200 rpm for six 1-min cycles with 40-s cooling on ice in between. The cytoplasmic extracts were harvested by centrifugation at 10,000×g for 5 min to remove cell wall fragments, and then concentrated to one fifth of the initial volume using Amicon Ultra-4 units as above.
The concentrated cell-free supernatants and cytoplasmic extracts (10 μg protein for each) were incubated separately with 5 mg mL−1 of the GL34 mixture. All reactions were performed in Milli-Q at 37 °C for 24 h. The progress of the reactions was followed by high-performance anion-exchange chromatography (HPAEC).
High-pH anion-exchange chromatography
Samples were analyzed on an ICS-3000 workstation (Dionex, Amsterdam, the Netherlands) equipped with an ICS-3000 pulse amperometric detection (PAD) system and a CarboPac PA-1 column (250 × 2 mm; Dionex). The analytical separation was performed at a flow rate of 0.25 mL min−1 using a complex gradient of eluents A (100 mM NaOH); B (600 mM NaOAc in 100 mM NaOH); C (Milli-Q water); and D 50 mM NaOAc. The gradient started with 10% A, 85% C, and 5% D in 25 min to 40% A, 10% C, and 50% D, followed by a 35-min gradient to 75% A, 25% B, directly followed by 5-min washing with 100% B and reconditioning for 7 min with 10% A, 85% B, and 5% D. External standards of lactose, glucose, and fructose were used for calibration. For the determination of glucosylated lactose compounds with a degree of polymerization (DP) of 3, maltotriose was used as external standard.
Bioinformatic analysis
All protein sequences from B. adolescentis ATCC 15703, B. longum subsp. infantis ATCC 15697, B. breve DSM 20123, and B. breve UCC 2003 used in this study were extracted from the National Center for Biotechnology Information (NCBI) database. Database searches used the non-redundant sequence database accessible at the NCBI website (http://www.ncbi.nlm.nih.gov) using BLASTP and global align search. The BLASTP searches and multiple-sequence alignments were used to find similarity between the characterized glucosidases of B. breve UCC 2003 and annotated glucosidases encoded by the studied bifidobacterial strains. Annotation of carbohydrate-active enzymes encoded by the genome sequences of L. reuteri 121 and L. acidophilus ATCC 4356 was carried out using dbCAN (http://csbl.bmb.uga.edu/dbCAN).
Results
Enzymatic hydrolysis of compounds in the GL34 mixture
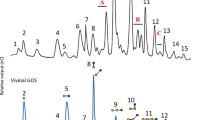
The GL34 mixture of five compounds was synthesized using glucansucrase Gtf180-ΔN, decorating lactose with one or two glucose units from sucrose as donor substrate, also introducing different types of linkages (Pham et al. 2017). GL34 contains three DP3 compounds and two DP4 compounds, i.e., F1 (4′-glc-lac): α-D-Glcp-(1 → 4)-β-D-Galp-(1 → 4)-D-Glcp; F2 (2-glc-lac): α-D-Glcp-(1 → 2)-[β-D-Galp-(1 → 4)-]D-Glcp; F3 (3′-glc-lac): α-D-Glcp-(1 → 3)-β-D-Galp-(1 → 4)-D-Glcp; F4 (4′,2-glc-lac): α-D-Glcp-(1 → 4)-β-D-Galp-(1 → 4)-[α-D-Glcp-(1 → 2)-]D-Glcp; and F5 (3′,2-glc-lac): α-D-Glcp-(1 → 3)-β-D-Galp-(1 → 4)-[α-D-Glcp-(1 → 2)-]D-Glcp (4). Four types of glycosidic linkages thus occur in this mixture, namely α(1 → 2), α(1 → 3), α(1 → 4), and β(1 → 4). Only the F2 2-glc-lac compound had been described before (Díez-Municio et al. 2012). The GL34 mixture also contains glucosyl residues linked α(1 → 3)/α(1 → 4) to the galactosyl residue of the original lactose. In view of the novel composition of this mixture of glucosylated-lactose compounds, we tested their resistance or sensitivity to hydrolysis with several commercially available enzymes. Following incubations with the porcine pancreas and A. oryzae α-amylases (Table S1), the HPAEC profiles at times 0 and 24 h showed no degradation of the GL34 compounds (Fig. 1). Also various malto-oligosaccharide acting enzymes (α-glucosidase, iso-amylase and pullulanase, Table S1) were tested for their ability to hydrolyze GL34 compounds. However, after 24-h incubation, no (monomeric or dimeric) products were detected in the reaction mixtures with these three enzymes (Fig. 1). None of these α-glucose cleaving enzymes thus was active on the GL34 compounds.

HPAEC profiles of oligosaccharides in 1 = the GL34 mixture (1 mg mL−1, blank) and the hydrolysis products after incubation of GL34 with 2 = α-amylase from porcine; 3 = α-amylase from A. oryzae; 4 = α-glucosidase from yeast; 5 = iso-amylase from Pseudomonas sp.; 6 = pullulanase type 1 from K. planticola; 7 = β-galactosidase from A. oryzae and 8 = β-galactosidase from K. lactis
Subsequent incubation of the GL34 mixture with the β-galactosidase enzymes from A. oryzae and K. lactis (Table S1) however, did result in (some) hydrolysis. Fig. 1 shows that galactose and kojibiose (a glucose disaccharide with α(1 → 2)-linkage) were released during incubation with β-galactosidase, especially with the A. oryzae enzyme. Only the peak corresponding to F2 2-glc-lac disappeared, the only GL34 compound with a terminal galactosyl residue. We subsequently studied the utilization of these GL34 compounds for growth by (selected) common intestinal bacteria in more detail.
Growth of human gut bacteria on the GL34 mixture
Fermentation of GL34 compounds by probiotic Lactobacillus strains
In this study, we tested L. casei W56, L. acidophilus ATCC 4356, and L. reuteri 121 and observed that all three strains showed limited growth on media with the GL34 mixture as the only carbon source (5 mg mL−1): the final (relative) OD600 values reached were 3.8%, 10.4%, and 26.5%, respectively, compared to a 100% control grown on glucose (Fig. 2).

Growth of L. casei W56, L. reuteri 121, and L. acidophilus ATCC 4356 on GL34 compounds (5 mg mL−1). Glucose (5 mg mL−1) served as positive control; growth studies were carried out in triplicate
Fermentation of GL34 compounds by probiotic Bifidobacterium strains
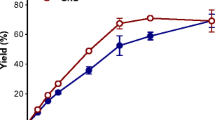
The GL34 mixture (5 mg mL−1) showed different stimulatory effects on the growth of B. breve DSM 20123, B. adolescentis ATCC 15703, and B. longum subsp. infantis ATCC 15697. B. adolescentis grew very well on GL34, its final OD600 value reached 80% of that of a 100% control growing on lactose, the purified TS0903 GOS mixture, and the GOS/FOS mixture (Table 1). However, the final OD600 values for growth of B. breve DSM 20123 and B. longum subsp. infantis ATCC 15697 on GL34 remained below 50% of the values for growth on lactose, the purified TS0903 GOS mixture and the GOS/FOS mixture (Table 1).
The tested bifidobacterial strains displayed two or more growth phases (Fig. 3 and Fig. S1). Some compounds in the GL34 mixture thus are more preferred growth substrates than others. B. breve and B. longum subsp. infantis grew more slowly on GL34 than B. adolescentis. They reached OD values around 0.70 after 24 h incubation compared to 12 h for B. adolescentis. Final maximal OD values were 0.86 for B. breve and 0.73 for B. longum subsp. infantis and 1.60 for B. adolescentis. The latter strain appeared to go through different lag phases, adapting to the different carbon sources in GL34, reaching maximal OD after 36 h of incubation (Fig. S1).

Growth of B. adolescentis ATCC 15703, B. longum subsp. infantis ATCC 15697, B. breve DSM 20123, E. coli Nissle, B. thetaiotaomicron in medium supplemented with 5 mg mL−1 GL34 at different times of incubation (h). Growth experiments were carried out in triplicate
Fermentation of GL34 compounds by commensal gut bacteria
Also, the ability of two selected commensal bacteria to grow on the GL34 mixture was studied. B. thetaiotaomicron is a Gram-negative anaerobic bacterium found dominantly in human distal intestinal microbiota (Xu et al. 2003). E. coli Nissle represents ecologically important inhabitants of the human intestinal tract (Mason and Richardson 1981). Growth of E. coli Nissle on the GL34 mixture, TS0903 GOS and a GOS/FOS mixture (5 mg mL−1), was relatively minor with final OD600 values of 0.19 ÷ 0.35 after 24-h of incubation, compared to growth on lactose and glucose with final OD600 values of 0.66 and 0.74, respectively (Fig. 4, panel 1). The final OD600 value of B. thetaiotaomicron was 0.53 after 72 h of incubation, but its growth displayed a pronounced lag phase (Fig. 4, panel 2). This strain thus may fail to compete with other bacteria which have shorter lag phases of growth with the GL34 mixture, such as the bifidobacteria tested and E. coli Nissle.

1 Growth of E. coli Nissle on the GL34, TS0903 GOS, and GOS/FOS mixtures (5 mg mL−1); lactose and glucose (5 mg mL−1) served as positive controls. 2 Growth of B. thetaiotaomicron on the GL34 compounds (5 mg mL−1); lactose (5 mg mL−1) served as positive control. Growth studies were carried out in triplicate
We subsequently identified the specific GL34 compounds utilized by these strains, and products derived, also aiming to elucidate which hydrolytic enzyme activities are involved, with emphasis on β-galactosidases and α-glucosidases.
Hydrolytic activity of commensal bacteria and lactobacilli on the GL34 mixture
The GL34 compounds can be visualized as individual peaks in HPAEC-PAD chromatograms. The compounds consumed by the tested bacteria were validated by peak disappearance at the corresponding retention time. HPAEC analysis of culture supernatants of the commensal bacteria grown on the GL34 mixture showed that the F2 peak corresponding to 2-glc-lac disappeared completely (Fig. 5), indicating the selective and full utilization of only F2. Most likely this is based on β-galactosidase degradation of F2 2-glc-lac, followed by α-glucosidase degradation of the kojibiose formed, and finally, consumption of the galactose and glucose formed for cellular growth. Only in the case of E. coli Nissle degradation resulted in accumulation of kojibiose. This was not investigated any further. Besides, B. thetaiotaomicron was also able to degrade F4 4′,2-glc-lac partially (Fig. 5).

HPAEC profiles of oligosaccharides in 1 = the GL34 mixture (1 mg mL−1, blank) and 2 = the hydrolysis products of GL34 fermentation by aE. coli Nissle at 40 hand bB. thetaiotaomicron at 72 h
HPAEC profiles of L. casei W56 culture supernatants showed that none of the GL34 compounds were degraded (Fig. 6), in line with the very limited growth observed (Fig. 2). When searching the annotated genome sequence of this strain in the CAZy database (http://www.cazy.org/b7858.html), we did not find any (putative) β-galactosidase. This most likely explains the inability of L. casei W56 to hydrolyze the galactose β(1 → 4) linked to glucosyl residue in F2 2-glc-lac. Many putative α-glucosidases were found encoded in the genome sequence of this strain, however. In view of the results obtained, these enzymes apparently are inactive on the GL34 compounds, or the GL34 mixture is unable to induce their expression. This was not investigated further.

HPAEC profiles of oligosaccharides in 1 = the GL34 mixture (1 mg mL−1, blank) and the hydrolysis products of GL34 fermentation after 16 h of incubation with 2 = L. casei W56; 3 = L. acidophilus ATCC 4356; 4 = L. reuteri 121, and 5 = MRS medium (blank)
Lactobacillus acidophilus ATCC 4356 and L. reuteri 121 strains only showed hydrolytic activity with F2 2-glc-lac in the GL34 mixture. L. reuteri 121 degraded this compound to a higher level than L. acidophilus ATCC 4356 (Fig. 6). Annotation of carbohydrate-active enzymes encoded by the genome sequence of L. reuteri 121 (Gangoiti et al. 2017) was carried out using dbCAN (http://csbl.bmb.uga.edu/dbCAN). One putative β-galactosidase of family GH2 was detected (gene number: BJI45_06415), which may be responsible for hydrolysis of F2. β-Galactosidases of this GH2 family are known to hydrolyze a wide variety of β-(1 → 2, 3, 4, or 6) GOS, including oligosaccharides with a degree of polymerization of 3–6 (Gänzle 2012). Kojibiose released from the F2 compound by this strain remained in the medium without being degraded (completely). Only a single (predicted) extracellular α-glucosidase of family GH31 was encoded in the genome of L. reuteri 121 (gene number: BIJ45_02455) (Gangoiti et al. 2017). This enzyme is apparently unable to effectively degrade kojibiose. Apparently, there is also no transporter expressed that can facilitate the uptake of kojibiose for intracellular degradation.
Lactobacillus acidophilus ATCC 4356 is known to encode a β-galactosidase (LacZ, classified in the GH2 family) (Gänzle 2012; Schwab et al. 2010; Pan et al. 2010).This LacZ enzyme thus may be responsible for degradation of the β(1 → 4)-linkage in F2 and release of kojibiose. Annotation of carbohydrate-active enzymes encoded by the genome sequence of L. acidophilus ATCC 4356 (Palomino et al. 2015) was also carried out by dbCAN (http://csbl.bmb.uga.edu/dbCAN). Ten putative α-glucosidase enzymes (family GH13 and GH31) were annotated in the L. acidophilus ATCC 4356 genome. Agl3 is anα-glucosidase identified in B. breve UCC 2003, with a broad hydrolytic activity towards all possible α-glycosidic linkages, including the α(1 → 2) linkage in sucrose and kojibiose (Kelly et al. 2016). However, a BLAST analysis of these putative glucosidases of L. acidophilus ATCC 4356 and Alg3 showed very low similarity in protein sequence (between 24 and 31%) (Table S2). The observed accumulation of kojibiose in the growth medium of L. acidophilus ATCC 4356 (Fig. 6) thus may be due to lack of an extracellular α-glucosidase active on α(1 → 2) linkages. Furthermore, it is possible that the α-glucosidase enzymes of this strain are only intracellular enzymes and that a suitable transporter for kojibiose is absent.
Hydrolytic activity of the Bifidobacterium strains on the GL34 mixture
Typically, bifidobacteria have extracellular endohydrolases acting on glycosidic bonds of oligo- and polymeric substrates, yielding smaller products which are internalized by carbohydrate-specific (ABC type) transporters. Further utilization may be carried out by cytoplasmic GHs such as α/β-glucosidases and α/β-galactosidases to produce monosaccharides which are used for growth (O’Connell-Motherway et al. 2011; O’Connell et al. 2013). To try and identify the bifidobacterial enzymes responsible for utilization of GL34 compounds, cells grown in modified BM broth containing 5 mg mL−1 GL34 were harvested at 36 h (Fig. 3). Three fractions were prepared, namely growth culture supernatants, concentrated culture supernatants and cytoplasmic extracts (see “Materials and methods”). GL34 was incubated with concentrated culture supernatants and cytoplasmic extracts to verify the presence or absence of extra- and/or intracellular enzyme activities involved in degradation.
Bifidobacterium adolescentis ATCC 15703
Bifidobacterium adolescentis was able to utilize all GL34 compounds, and only 10 and 60% of F4 4′,2-glc-lac and F5 3′,2-glc-lac remained, respectively (Fig. 7, panel 1). This also explains the relatively strong growth of this strain (Fig. 3). None of the GL34 compounds were hydrolyzed by cell-free culture supernatants of this strain. Its cell extracts only hydrolyzed F1 4′-glc-lac, F2 2-glc-lac, and F4 4′,2-glc-lac compounds (Fig. 7, panel 1). Three α-glucosidases are annotated in the B. adolescentis ATCC 15703 genome sequence (http://www.cazy.org), namely AglB; AglA and BAD-0971. AglB exhibited a preference for hydrolyzing α(1 → 4) linkages in maltose (van den Broek et al. 2003), and most likely also is involved in cleavage of the α(1 → 4) linkages present in the F1 and F4 compounds. AglA showed high hydrolytic activity only towards α(1 → 6) linkages in isomaltotriose and minor activity towards α(1 → 1) linkages in trehalose (van den Broek et al. 2003). None of these two enzymes are able to degrade α(1 → 3) linkages occurring in F3 3′-glc-lac and F5 3′,2-glc-lac. It thus remained possible that during growth the F3 and F5 were (partially) degraded by the BAD-0971 enzyme, the third B. adolescentis ATCC 15703 α-glucosidase.

Consumption of GL34 compounds (%) during growth of 1B. adolescentis ATCC 15703; 2B. longum subsp. infantis ATCC 15697; and 3B. breve DSM 20213, and after 24-h incubations of 5 mg mL−1 GL34 with cytoplasmic extracts or cell-free supernatants (obtained after growth of these three strains on the GL34 mixture)
The BAD-0971 protein has been annotated as an α-1,4-glucosidase but has not been characterized yet (Suzuki et al. 2006). A BLASTP analysis was carried out to compare its protein sequence with those of the characterized glucosidases of B. adolescentis ATCC 15703 (AglA and AglB) and B. breve UCC 2003 (Agl1; Agl2 and Agl3). Agl3 from B. breve UCC 2003, as mentioned above, has a broad hydrolytic specificity towards α(1 → 2), α(1 → 3), α(1 → 4), α(1 → 5), and α(1 → 6) in kojibiose, turanose, maltose, leucrose, and palatinose, respectively (McLaughlin et al. 2015). The two other α-glucosidases of this strain (Agl1 and Agl2) are more active in cleaving α(1 → 6) linkages (in panose, isomaltose, isomaltotriose, and palatinose) and less efficient in cleaving α(1 → 3) linkages in turanose and nigerose, α(1 → 4) in maltulose and α(1↔2) in sucrose (Pokusaeva et al. 2009). AglA and Agl3 exhibit 59% and 73% similarity/identity, respectively, to the BAD-0971 protein of B. adolescentis (Table S3). The BAD-0971 enzyme may have a broad hydrolytic specificity similar to Agl3. However F3 3′-glc-lac and F5 3′,2-glc-lac with α(1 → 3) linkages remained undigested in cell extracts. It cannot be excluded that the cell disruption process has weakened activity of BAD-0971 (annotated as an α-1,4-glucosidase), or other hitherto unidentified α-glucosidase enzymes.
Bifidobacterium longum subsp. infantis ATCC 15697
The F2 2-glc-lac and F3 3′-glc-lac compounds were completely utilized during growth of B. longum subsp. infantis, and the F1 4′-glc-lac, F4 4′,2-glc-lac and F5 3′,2-glc-lac compounds only for 20–40% (Fig. 7, panel 2). Cell extracts only hydrolyzed F2 2-glc-lac, apparently involving (an) intracellular enzyme(s) (Fig. 7, panel 2). The F1 4′-glc-lac, F3 3′-glc-lac, F4 4′,2-glc-lacand F5 3′,2-glc-lac structures were not utilized by cell extracts of this strain, suggesting that it lacks α-glucosidase enzymes acting on glucose α(1 → 3)/α(1 → 4) linked to galactosyl residue. The complete use of the F3 3′-glc-lac compound during growth of B. longum subsp. infantis ATCC 15697 thus remained unexplained again. No extracellular activity of B. longum subsp. infantis towards the GL34 mixture was detected (Fig. 7, panel 2). Its limited activity on the GL34 mixture resulted in the lowest growth extent (37%) compared to the other two bifidobacteria (Table 1). Various glucosidases have been annotated in the genome of B. longum subsp. infantis mainly with activity on α(1 → 4) and α(1 → 6) glucosidic linkages, namely α-1,4 glucosidase (locus tag BLIJ-0129); putative amylase (locus tag BLIJ-0286); putative iso-amylase (locus tag BLIJ-0286 and BLIJ-2315); and oligo-1,6-glucosidase (locus tag BLIJ-2526) (www.cazy.org/). However, BLAST analysis showed very low similarity in their protein sequence between the annotated glucosidases from this strain and Alg3 (Table S4). This most likely explains the poor growth of this strain on the GL34 mixture, and its limited intracellular activity on α1 → 3/α1 → 4 glucosidic linkages in F1 4′-glc-lac, F3 3′-glc-lac, F4 4′,2-glc-lac, and F5 3′,2-glc-lac.
Bifidobacterium breve DSM 20213
In case of B. breve, four of the five GL34 compounds (F1 4′-glc-lac, F3 3′-glc-lac, F4 4′,2-glc-lac, and F5 3′,2-glc-lac) (partly) remained unutilized in culture supernatants after growth (Fig. 7, panel 3). Amongst the three bifidobacteria tested, only B. breve possessed extracellular activity on the mixture GL34: the F2 2-glc-lac and F5 3′,2-glc-lac compounds were completely degraded by cell-free supernatants after 24-h incubation (Fig. 7, panel 3). The extracellular activity was also observed previously in different B. breve strains; however, on α(1 → 4) and α(1 → 6) glucosidic linkages present in starch, amylopectin and pullulan (Ryan et al. 2006). Nevertheless, cell extracts fully hydrolyzed F2 2-glc-lac, F3 3′-glc-lac, and F5 3′,2-glc-lac (Fig. 7, panel 3). Most likely B. breve is unable to transport F3 3′-glc-lac and F5 3′,2-glc-lac into the cell. These data suggest that intracellular enzymes of this strain were able to cleave off the glucose units α(1 → 3) linked to the galactosyl residues occurring in F3 3′-glc-lac and F5 3′,2-glc-lac but not the glucose units α(1 → 4) linked to the galactosyl residues in F1 4′-glc-lac and F4 4′,2-glc-lac.
The putative α-glucosidases in the B. breve DSM 20213 genome, however, are annotated to act mainly on α(1 → 4) and α(1 → 6) glucosidic linkages; namely α-1,4 glucosidase (locus tag BBBR-0095); putative amylase (locus tags BBBR-0101; BBBR-0825 and BBBR-0257) and oligo-1,6-glucosidase (locus tags BBBR-0484 and BBBR-1863) (www.cazy.org/). This uncharacterized α-1,4 glucosidase (locus tag BBBR-0095) was found to have 99% similarity with Alg3 of B. breve UCC 2003 (Table S4). This annotated α-1,4 glucosidase thus also may be able to hydrolyze α(1 → 3) linkage in F3 3′-glc-lac and F5 3′,2-glc-lac. Structures of F1 4′-glc-lac and F4 4′,2-glc-lac may be inaccessible to this α-glucosidase, or it is unable to cleave their α(1 → 4) linkages, thus explaining their (very) limited degradation by B. breve DSM 20213.
The annotated β-galactosidases (family GH2) of these three Bifidobacterium strains are active on the F2 2-glc-lac compound, containing a galactosyl-moiety with a β(1 → 4) linkage to kojibiose. Following growth on GL34 as only carbon source, all three strains exhibited clear intracellular activity with F2 2-glc-lac. The data showed that individual Bifidobacterium strains have preference for degradation of glucosylated lactose compounds with α(1 → 3) and α(1 → 4) glucosidic linkages.
Discussion
The GL34 compounds generally consist of a lactose molecule at the reducing end that is elongated with one or more glucose molecules involving different linkage types (Pham et al. 2017). This combination of different monomers, i.e., glucose and galactose, with various linkage types (α(1 → 2), α(1 → 3), α(1 → 4), and β(1 → 4)) and different degrees of polymerization, increases the diversity of lactose-derived oligosaccharides available. The GL34 compounds were resistant to degradation by the α-amylases of porcine pancreas and A. oryzae. These are endo-acting enzymes (EC 3.2.1.1), degrading α(1 → 4) glucosidic linkages in polysaccharides such as starch or glycogen (van der Maarel et al. 2002), mainly yielding glucose and maltose. Also, various MOS-acting enzymes were tested: The α-glucosidase enzyme of yeast used is known as an exo-acting enzyme, hydrolyzing α(1 → 4) glucosidic linkages but only at the terminal non-reducing (1 → 4)-linked α-glucose residues of di- and oligosaccharides to release a single glucose unit (Reese et al. 1968). Iso-amylase from Pseudomonas sp. is an α(1 → 4,6) debranching enzyme (van der Maarel et al. 2002). Pullulanase type I of K. planticola hydrolyzes α(1 → 6) glucosidic linkages in pullulan and starch (Domań-Pytka and Bardowski 2004). Also, these three enzymes failed to cleave any compounds in the GL34 mixture. Incubations with β-galactosidase enzymes however did result in hydrolysis, but only the F2 2-glc-lac molecule disappeared. This is explained by the ability of these enzymes to catalyze hydrolysis of β-glycosidic bonds between galactose and its organic moiety. The combined data thus shows that the GL34 compounds are (largely) resistant to hydrolysis by these common carbohydrate degrading enzymes (Fig. 1).
There is abundant clinical evidence for the important roles of Bifidobacterium and Lactobacillus species in the eco-physiology of the intestinal microbiota (Picard et al. 2005; Sanders and Klaenhammer 2001), and different strains of lactobacilli are marketed as commercial probiotics. Individual bifidobacteria are known to have specific substrate preferences (Degnan and Macfarlane 1991). The GL34 mixture stimulated growth of bifidobacteria, but indeed to different extents. Only utilization of the F2 2-glc-lac compound has been studied previously and was shown to have a limited stimulatory effect on the growth of B. breve 26M2. This F2 compound did not stimulate growth of lactobacilli tested in our present study, as previously shown for L. casei LC-01 (García-Cayuela et al. 2014). However, we observed that F2 2-glc-lac stimulated growth of the probiotic bacteria L. reuteri 121, B. adolescentis ATCC 15703, B. longum subsp. infantis ATCC 15697, and B. breve DSM 20213, and also of two commensal bacteria, E. coli Nissle and B. thetaiotaomicron, albeit to various extents. This F2 compound thus is less selective in comparison with the other compounds in the GL34 mixture. Both F1 4′-glc-lac and F4 4′,2-glc-lac also stimulated growth of all three tested bifidobacteria, again to various extents. The presence of an α(1 → 3) linkage makes F3 3′-glc-lac more selective than F1 and F4: F3 was utilized by only two out of three studied Bifidobacterium strains: B. adolescentis ATCC 15703 and B. breve DSM 20213. Also, F5 3′,2-glc-lac with both α(1 → 2) and α(1 → 3) glucosidic linkages showed similar stimulatory effects on all three Bifidobacterium strains.
In conclusion, the GL34 mixture promotes growth of the tested bacteria to different extents. The bifidobacteria tested generally were better at degrading GL34 compounds than the lactobacilli and commensal bacteria. The stronger metabolic toolset of bifidobacteria in comparison with lactobacilli also has been observed when comparing their growth on human milk oligosaccharides and other prebiotic oligosaccharides as primary carbon source (Watson et al. 2013; Sela et al. 2008; González et al. 2008; McLaughlin et al. 2015). The GL34 mixture thus showed potential to shift microbiota composition by specifically stimulating growth of bifidobacteria, particularly B. adolescentis. Four out of five compounds in this GL34 mixture exerted high and selective growth stimulatory effects towards health-beneficial probiotic bifidobacteria. The combination of monomer composition and linkage type clearly determines the fermentable properties of the GL34 compounds. Individual gut bacteria were able to utilize only specific compounds in the GL34 mixture. Synergistic activities between bacterial species thus are likely to be essential for the utilization of the whole GL34 mixture. In future work, this will be studied in more detail, e.g., by using fecal bacterial cultures. Only B. adolescentis was able to utilize almost all structures, providing a potential synbiotic combination.
References
Bai Y, Böger M, van der Kaaij RM, Woortman AJJ, Pijning T, van Leeuwen SS, van Bueren AL, Dijkhuizen L (2016) Lactobacillus reuteri strains convert starch and maltodextrins into homoexopolysaccharides using an extracellular and cell-associated 4,6-α-glucanotransferase. J Agric Food Chem 64:2941–2952
Bindels LB, Delzenne NM, Cani PD, Walter J (2015) Towards a comprehensive concept for prebiotics. Nat Rev Gastroenterol Hepatol 12:303–310
Bode L (2012) Human milk oligosaccharides: every baby needs a sugar mama. Glycobiology 22:1147–1162
Boehm G, Moro G (2008) Structural and functional aspects of prebiotics used in infant nutrition. J Nutr 138:1818S–1828S
Callaway TR, Edrington TS, Anderson RC, Harvey RB, Genovese KJ, Kennedy CN, Venn DW, Nisbet DJ (2008) Probiotics, prebiotics and competitive exclusion for prophylaxis against bacterial disease. Anim Health Res Rev 9:217–225
Daniels L, Zeikus JG (1975) Improved culture flask for obligate anaerobes. Appl Microbiol 29:710–711
Degnan BA, Macfarlane GT (1991) Comparison of carbohydrate substrate preference in eight species of bifidobacteria. FEMS Microbiol Lett 84:151–156
Díez-Municio M, Montilla A, Luisa Jimeno M, Corzo N, Olano A, Javier Moreno F (2012) Synthesis and characterization of a potential prebiotic trisaccharide from cheese whey permeate and sucrose by Leuconostoc mesenteroides dextransucrase. J Agric Food Chem 60:1945–1953
Domań-Pytka M, Bardowski J (2004) Pullulan degrading enzymes of bacterial origin. Crit Rev Microbiol 30:107–121
Ferreira-Lazarte A, Olano A, Villamiel M, Moreno FJ (2017) Assessment of in vitro digestibility of dietary carbohydrates using rat small intestinal extract. J Agric Food Chem 65:8046–8053
Flint HJ, Duncan SH, Scott KP, Louis P (2007) Interactions and competition within the microbial community of the human colon: links between diet and health, mini review. Environ Microbiol 9:1101–1111
Gangoiti J, Meng X, Lammerts van Bueren A, Dijkhuizen L (2017) Draft genome sequence of Lactobacillus reuteri 121, a source of α-glucan and β-fructan exopolysaccharides. Genome Announc 5:e01691–e01616
Gänzle MG (2012) Review: enzymatic synthesis of galacto-oligosaccharides and other lactose derivatives (hetero-oligosaccharides) from lactose. Int Dairy J 22:116–122
García-Cayuela T, Díez-Municio M, Herrero M, Martínez-Cuesta MC, Peláez C, Requena T, Moreno FJ (2014) Selective fermentation of potential prebiotic lactose-derived oligosaccharides by probiotic bacteria. Int Dairy J 38:11–15
Gibson GR, Hutkins R, Sander ME, Prescott SL, Reimer RA, Salminen SJ, Scott K, Stanton C, Swanson KS, Cani PD, Verbeke K, Reid G (2017) Expert consensus document: the international acientific association for probiotics and prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol 14:491–502
González R, Klaassens ES, Mahnen E, De Vos WM, Vaughan EE (2008) Differential transcriptional response of Bifidobacterium longum to human milk, formula milk, and galactooligosaccharide. Appl Environ Microbiol 74:4686–4694
Jaskari J, Kontula P, Siitonen A, Jousimies-Somer H, Mattila-Sandholm T, Poutanen K (1998) Oat β-glucan and xylan hydrolysates as selective substrates for Bifidobacterium and Lactobacillus strains. Appl Microbiol Biotechnol 49:175–181
Kelly ED, Bottacini F, O'Callaghan J, O’Connell Motherway M, O'Connell KJ, Stanton C, van Sinderen D (2016) Glycoside hydrolase family 13 α-glucosidases encoded by Bifidobacterium breve UCC2003; a comparative analysis of function, structure and phylogeny. Int J Food Microbiol 224:55–65
Koropatkin NM, Martens EC, Gordon JI, Smith TJ (2008) Starch catabolism by a prominent human gut symbiont is directed by the recognition of amylose helices. Structure 16:1105–1115
Lammerts van Bueren A, Mulder M, van Leeuwen SS, Dijkhuizen L (2017) Prebiotic galactooligosaccharides activate mucin and pectic galactan utilization pathways in the human gut symbiont Bacteroides thetaiotaomicron. Sci Rep 7:40478
Lehmann U, Robin F (2007) Slowly digestible starch - its structure and health implications: a review. Trends Food Sci Technol 18:346–355
Ley RE, Peterson DA, Gordon JI (2006) Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 124:837–848. https://doi.org/10.1016/j.cell.2006.02.017
Louis P, Scott KP, Duncan SH, Flint HJ (2007) Understanding the effects of diet on bacterial metabolism in the large intestine. J Appl Microbiol 102:1197–1208
Macfarlane GT, Steed H, Macfarlane S (2008) Bacterial metabolism and health-related effects of galacto-oligosaccharides and other prebiotics. J Appl Microbiol 104:305–344
Marquina D, Santos D, Corpas I, Muñoz J, Zazo J, Peinado JM (2002) Dietary influence of kefir on microbial activities in the mouse bowel. Lett Appl Microbiol 35:136–140
Mason TG, Richardson G (1981) Escherichia coli and the human gut: some ecological considerations. J Appl Bacteriol 51:1–16
McLaughlin HP, Motherway MO, Lakshminarayanan B, Stanton C, Ross RP, Brulc J, Menon R, O'Toole PW, van Sinderen D (2015) Carbohydrate catabolic diversity of bifidobacteria and lactobacilli of human origin. Int J Food Microbiol 203:109–121
O’Connell KJ, O’Connell-Motherway M, O'Callaghan J, Fitzgerald GF, Ross RP, Ventura M, Stanton C, van Sinderen D (2013) Metabolism of four α-glycosidic linkage-containing oligosaccharides by Bifidobacterium breve UCC2003. Appl Environ Microbiol 79:6280–6292
O’Connell-Motherway M, Fitzgerald GF, van Sinderen D (2011) Metabolism of a plant derived galactose-containing polysaccharide by Bifidobacterium breve UCC2003. Microb Biotechnol 4:403–416
Ohkusa T, Ozaki Y, Sato C, Mikuni K, Ikeda H (1995) Long-term ingestion of lactosucrose increases Bifidobacterium sp. in human fecal flora. Digestion 56:415–420
Palomino MM, Allievi MC, Fina Martin J, Waehner PM, Prado AM, Sanchez RC, Ruzal SM (2015) Draft genome sequence of the probiotic strain Lactobacillus acidophilus ATCC 4356. Genome Announc 3:7–8
Pan Q, Zhu J, Liu L, Cong Y, Hu F, Li J, Yu X (2010) Functional identification of a putative β-galactosidase gene in the special lac gene cluster of Lactobacillus acidophilus. Curr Microbiol 60:172–178
Pandey KR, Naik SR, Vakil BV (2015) Prebiotics, probiotcs and synbiotics—a review. J Food Sci Technol 52:7577–7587
Pham HTT, Dijkhuizen L, van Leeuwen SS (2017) Structural characterization of glucosylated lactose derivatives synthesized by the Lactobacillus reuteri GtfA and Gtf180 glucansucrase enzymes. Carbohydr Res 449:59–64
Picard C, Fioramonti J, Francois A, Robinson T, Neant F, Matuchansky C (2005) Review article: bifidobacteria as probiotic agents—physiological effects and clinical benefits. Aliment Pharmacol Ther 22:495–512
Pokusaeva K, O’Connell-Motherway M, Zomer A, Fitzgerald GF, Van Sinderen D (2009) Characterization of two novel α-glucosidases from Bifidobacterium breve UCC2003. Appl Environ Microbiol 75:1135–1143
Rastall RA, Maitin V (2002) Prebiotics and synbiotics: towards the next generation. Curr Opin Biotechnol 13:490–496
Reese ET, Maguire AH, Parrish FW (1968) Glucosidases and exo-glucanases. Can J Biochem 46:25–34
Rijnierse A, Jeurink PV, Van Esch BCAM, Garssen J, Knippels LMJ (2011) Food-derived oligosaccharides exhibit pharmaceutical properties. Eur J Pharmacol 668:S117–S123
Roberfroid M, Gibson GR, Hoyles L, McCartney AL, Rastall R, Rowland I, Wolvers D, Watzl B, Szajewska H, Stahl B, Guarner F, Respondek F, Whelan K, Coxam V, Davicco M-Jeanne, Léotoing L, Wittrant Y, Delzenne NM, Cani PD, Neyrinck AM, Meheust A (2010) Prebiotic effects: metabolic and health benefits. Br J Nutr 104 Suppl: S1–63
Ryan SM, Fitzgeral GF, van Sinderen D (2006) Screening for and identification of starch-, amylopectin-, and pullulan-degrading activities in bifidobacterial strains. Appl Environ Microbiol 72: 5289–96
Rycroft CE, Jones MR, Gibson GR, Rastall RA (2001) A comparative in vitro evaluation of the fermentation properties of prebiotic oligosaccharides. J Appl Microbiol 91:878–887
Sambrook J, Russell DW (2001) Molecular cloning: a laboratory manual. Cold Spring Harb Lab Press Cold Spring Harb NY 999
Sanders ME, Klaenhammer TR (2001) Invited review: the scientific basis of Lactobacillus acidophilus NCFM functionality as a probiotic. J Dairy Sci 84:319–331
Schwab C, Sørensen KI, Gänzle MG (2010) Heterologous expression of glycoside hydrolase family 2 and 42 β-galactosidases of lactic acid bacteria in Lactococcus lactis. Syst Appl Microbiol 33:300–307
Sela DA, Chapman J, Adeuya A, Kim JH, Chen F, Whitehead TR, Lapidus A, Rokhsar DS, Lebrilla CB, German JB, Price NP, Richardson PM, Mills DA (2008) The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc Natl Acad Sci U S A 105:18964–18969
Slavin J (2013) Fiber and prebiotics: mechanisms and health benefits. Nutrients 5:1417–1435
Suzuki T, Tsuda Y, Kanou N, Inoue T, Kumazaki K, Nagano S, Hirai S, Tanaka K, Watanabe K (2006) Bifidobacterium adolescentis complete genome sequence. Publ. Only database
van den Broek L, Struijs K, Verdoes J, Beldman G, Voragen A (2003) Cloning and characterization of two alpha-glucosidases from Bifidobacterium adolescentis DSM20083. Appl Microbiol Biotechnol 61:55–60
van der Maarel MJEC, van der Veen B, Uitdehaag JCM, Leemhuis H, Dijkhuizen L (2002) Properties and applications of starch-converting enzymes of the alpha-amylase family. J Biotechnol 94:137–155
Van Leeuwen SS, Kuipers BJH, Dijkhuizen L, Kamerling JP (2016) Comparative structural characterization of 7 commercial galacto-oligosaccharide (GOS) products. Carbohydr Res 425:48–58
Watson D, O'Connell Motherway M, Schoterman MHC, Joost van Neerven RJ, Nauta A, van Sinderen D (2013) Selective carbohydrate utilization by lactobacilli and bifidobacteria. J Appl Microbiol 114:1132–1146
Xu J, Bjursell MK, Himrod J, Deng S, Carmichael LK, Chiang HC, Hooper LV, Gordon JI (2003) A genomic view of the human-Bacteroides thetaiotaomicron symbiosis. Science 299:2074–2076
Yan YL, Hu Y, Gänzle MG (2018) Prebiotics, FODMAPs and dietary fiber—conflicting concepts in development of functional food products? Curr Opin Food Sci 20:30–37
Acknowledgements
We thank Alicia Brandt for her support for growth of commensal bacteria and for stimulating discussions.
Funding
This work was financially supported by the University of Groningen/Campus Fryslân, and by FrieslandCampina. This research was also performed in the public-private partnership CarboHealth coordinated by the Carbohydrate Competence Center (CCC, www.cccresearch.nl) and financed by participating partners and allowances of the TKI Agri&Food program, Ministry of Economic Affairs.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Ethical approval
This article does not contain any studies with human participants or animals by any of the authors
Electronic supplementary material
ESM 1
(PDF 120 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Pham, H.T.T., Boger, M.C.L., Dijkhuizen, L. et al. Stimulatory effects of novel glucosylated lactose derivatives GL34 on growth of selected gut bacteria. Appl Microbiol Biotechnol 103, 707–718 (2019). https://doi.org/10.1007/s00253-018-9473-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-018-9473-8