
Abstract
Bleaching events impact coral reef functionality and carbonate budget dynamics, which is reflected in reduced reef framework accretion, hindering reef ability to keep pace with sea-level rise projections. Reefs in the Arabian Gulf exist in harsh environmental conditions with seasonal temperatures ranging between 16 and 36 °C. Despite the high thermal thresholds (~ 35 °C) of corals in this region, extensive bleaching and high coral mortality have been reported regionally in 2017. We quantify reef carbonate budgets at a near shore and an offshore reef site in Bahrain pre-bleaching in May 2017 and post-bleaching in May 2018 to assess the impacts of the 2017 severe bleaching event on the budgetary state. Results indicate an overall decrease in hard coral cover from 14.2 ± 5.5 to 8.5 ± 1.4% and a decline in the net carbonate budget state from 3.6 ± 2.2 to 0.3 ± 0.3 kg CaCO3 m−2 year−1 at the shallow nearshore Fasht Al Adhm reef indicating a shift from a positive budgetary state to net neutral, while the deeper offshore Reef Bul Thamah has increased positively from 3.7 ± 1.2 to 4.2 ± 0.6 kg CaCO3 m−2 year−1. We attribute the decline in the nearshore reef to the bleaching event which took place between July and October 2017, resulting in high coral mortality rates and subsequent reduced framework carbonate production. Predicted warming trends present a threat to the structural integrity of shallow Bahraini reefs, compromising their ability to keep pace with future sea-level rise projections.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Coral reefs are experiencing rapid change on a global scale as a result of climate-induced disturbances (e.g., ocean warming and acidification; Anthony et al. 2008; Hughes et al. 2017a, b) coupled with local pressures, such as overfishing and declining water quality due to nutrient runoffs and excessive sedimentation (Hughes et al. 2003; Ove Hoegh-Guldberg 2011; de Bakker et al. 2019). These changes include loss of coral cover resulting in modification of the 3-D framework of reefs which in turn compromises their structural integrity (Perry and Morgan 2017) and loss of architectural complexity such as that described in the Caribbean by Alvarez-Filip et al. (2009). Changes in the functionality of reef associated fish have also been documented and strong negative effects of coral loss were noted on the individual abundance of coral reef fish (Pratchett et al. 2011) with many reefs showing shifts in coral and associated species community composition (Hoey et al. 2016; Ryan et al. 2019).
In Bahrain which is situated in the Arabian Gulf (AG), coral reef areas (locally known as “fashts”) occupy a total area of 850 km2, which is larger than the land area of Bahrain with Fasht Al Adhm itself covering 200 km2 and mostly restricted to the east and north of the main island (Burt et al. 2013). In the 1980s, 30 coral species were recorded in Bahrain with Acropora dominating communities in shallow (< 5 m) habitats, while Poritids and Faviids dominated the deeper (< 20 m) habitats (Alkhuzai et al. 2009); however, prior to this, little information is available on corals reefs in Bahrain (Sheppard 1988; Vousden 1988, 1995; Burt et al. 2013). In 1988, dense coral cover of 50–90% dominated by Acropora was reported on Bahraini reefs (Sheppard 1988; Vousden 1988, 1995); however, just a few years later, reports documented an estimated loss of > 90% of coral cover (Wilkinson 1998; Uwate and Shams 1999). This loss was linked to mass bleaching events in 1996 and 1998 when temperatures reached 37.7 °C and 38 °C, respectively, making Bahrain the worst affected AG country by these bleaching events (Uwate and Shams 1999; Rezai et al. 2004; Al-kuwari 2006).
Reef accretion is essential to reef survival in the face of rising sea levels (Perry et al. 2015, 2018a, b) and there are concerns around the ability of reefs to maintain their 3-D structural framework, which underpins the ecological functioning of coral reefs and the services they provide through continuous accumulation of calcium carbonate (CaCO3) (Graham and Nash 2013; Kennedy et al. 2013; Yanovski et al. 2017; Perry et al. 2018a).
Carbonate budgets measure the balance between CaCO3 production and erosion and allow estimation of reef accretion rates. Detecting changes in budget states can inform reef functionality change and/or stressors; it is therefore important to understand the magnitude of changes in reef carbonate budgets through time, between regions or following disturbances.
CaCO3 is precipitated by primary (hermatypic corals; Stearn and Scoffin 1977; Hubbard 1986; Hubbard et al. 1986; Perry and Hepburn 2008) and secondary reef builders (calcareous encrusters, e.g., coralline algae, Goreau 1963; Choi and Ginsburg 1983; Hepburn et al. 2015), which, together with incorporation of carbonate sediment into the reef framework, lead to carbonate accumulation. Bioerosion is the counter process and is driven by a diverse range of taxa including parrotfish, urchins, bioeroding sponges, and other macro- and micro-endolithic taxa.
Sea surface temperatures (SST) have been documented to increase globally, with records reporting an average rise of 1.0 °C between 1951 and 2015 (i.e., 0.15 °C/decade) in the Indian Ocean (Roxy et al. 2020), while the South Pacific and Western South Atlantic are reported to have experienced a rise in SST of ~ 0.4 °C/decade (Risaro et al. 2022). Short-term temperature anomalies (i.e., marine heat waves) are increasing and have been shown to cause coral bleaching and mortality (Berkelmans and Oliver 1999; Baker et al. 2004; Hughes et al. 2017a, b). An increase in temperature anomaly above a coral’s local tolerance level by 1–2 °C is sufficient to cause bleaching (Hoegh-Guldberg 1999; Purkis and Riegl 2005; Heron et al. 2016; Hoegh-Guldberg et al. 2017), negatively impacting carbonate budgets and reef capacity to sustain framework development and vertical growth (Januchowski-Hartley et al. 2017; Perry and Morgan 2017).
In the AG, corals are subjected to large and unique seasonal variations in temperatures (16–36 °C) (Coles and Riegl 2013; Hume et al. 2013). In the last 2 decades, reefs in this region have been exposed to severe temperature anomalies (~ 2 °C above the average daily temperature in the summer) at a frequent and highly recurring rate (Riegl 2002; Sheppard and Loughland 2002; Burt et al. 2012), similar to those projected (1.5–2 °C above pre-industrial levels) to occur globally by 2100 (IPCC 2014). Due to their extreme tolerances to seasonal temperature changes, coral communities in AG have the highest bleaching threshold (i.e., 35 °C) globally (Rezai et al. 2004; Riegl et al. 2012; Kavousi et al. 2014; Shuail et al. 2016). However, despite their high threshold, they are still susceptible to bleaching and it is reported that 70% of original AG reef cover (i.e., 3800 km2) may be considered to have suffered bleaching-related mortality, and a further 27% threatened or at critical stages of degradation with declines linked to major bleaching events, e.g., 1998 (Wilkinson 2008; van Lavieren et al. 2011). In recent years, one-third of the world’s coral reefs experienced huge losses due to the 2016 heat wave (Schiermeier 2018), whereas the AG reefs experienced a 1-year (2017) delay which resulted in severe mortality (Burt et al. 2019; Paparella et al. 2019). Over the past decades, efforts have been made to calculate reef carbonate budgets (Stearn and Scoffin 1977; Hutchings and Bamber 1985; Holmes et al. 2000; Hepburn 2006; Mallela and Perry 2007; Browne 2011). Bleaching-associated declines in reef carbonate budgets have been found in the central Indian ocean (Perry et al. 2014; Januchowski-Hartley et al. 2017; Perry and Morgan 2017; Lange and Perry 2019); however, no previous attempt has been made to assess reef carbonate budgets in the AG. Therefore, this paper will examine a nearshore and an offshore reef in Bahrain both pre- and post-the 2017-bleaching event with the aim to characterize the benthic composition of carbonate producers and eroders, quantify reef carbonate budgets, and report on the impacts on budgetary states caused by the 2017 warming event.
Materials and methods
Site description
Bahrain is an archipelago of 33 low-lying islands (Fig. 1) and its coral reefs were among the most extensive in the southern basin of the Arabian Gulf (Riegl 2003; Purkis and Riegl 2005; Kavousi et al. 2014). However, in the last 4 decades, these reefs have undergone significant decline in coral cover due to large-scale coastal development in pursuit of socio-economic growth. This has added 13% to the Kingdom’s total land area between 1961 (690 km2) and 2021 (780 km2) (World Bank 2021).

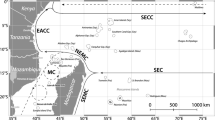
Location of study sites: A location of Arabian Gulf (AG); B location of Bahrain within AG; C location of selected study sites within Bahrain-Fasht Al Adhm (Nearshore) and Reef Bul Thamah (Offshore). Shapefiles were created using google earth, while all maps were created using R
To understand the thermal profile experienced on Bahraini reefs, we analyzed the time series weekly SST mean for the period 1982–2017 and the degree heating weeks (DHW) and thermal stress anomaly (TSA), giving an indication of the delineation between coral bleaching and mortality levels. Data were derived from CoRTAD version 6 database (Fig. 2; refer to supplementary material for method of acquisition and processing in addition to definition and calculation methods of SST thermal history metrics).

A Time series weekly SST mean for the period (1982–2017; n = 1878) illustrating the change in SST across the two study sites in Bahrain. The blue and red dotted lines indicate the minimum and maximum climatology (defined here as the long-term mean of SST conditions). Positive or negative values of SST exceed long-term mean SST (climatology) of a particular area, which indicates whether SST is warmer or cooler than usual (Heron et al. 2016)), respectively, while the red solid line indicates the bleaching threshold. The blue solid line indicates the linear fit with standard error of the regression in dark gray shading. B Degree heating weeks (DHW; in red solid lines) defined as of accumulated daily hotspots over 12 consecutive weeks when the thermal stress anomaly (TSA; in black solid lines) is ≥ 1 °C. The gray dotted lines indicate the DHW values of 4 and 8 °C-weeks, which corresponds to delineation between coral bleaching and mortality levels. Years that correspond to coral bleaching and mortality were highlighted in blue and red fonts, respectively. Data were derived from the CoRTAD version 6 database (Coral Reef Temperature Anomaly Database (CoRTAD) which is a dataset of SST and related thermal stress metrics, developed by NOAA specifically for coral reef ecosystem applications. The CoRTAD Version 6 database used in this study contains global ~ 4 km resolution SST data on a weekly time scale for the period 1982–2017. ftp://ftp.nodc.noaa.gov/pub/data.nodc/cortad/README.html)
In subsequent years following the 1998 bleaching episode, recovery of Bahraini reefs was observed to be limited, (Lough 2000; Uwate et al. 2000) with reports observing the reefs to be dominated by rock (66.9%) and sand (30.6%) with low coral cover (0.6%) in Fasht Al Adhm and no observations of the formerly dominant Acropora (Uwate et al. 2000).
For the purpose of this study, two reefs in Bahrain were investigated in April/May 2017 (pre-bleaching) and 2018 (post-bleaching; Fig. 1c) comparing a nearshore site at Fasht Al Adhm (Site Nearshore, 5–7 m depth, turbid conditions) and an offshore site at Reef Bul Thamah (Site Offshore, 10–12 m, clear conditions). Table 1 provides details of the environmental characteristics of each site. Surveying depths differ due to natural topography of the sites; Site Nearshore is a gently sloping shallow reef, with a maximum depth of 7 m and minimal relief due to the site being relatively homogenous and lacking more typical reef structural complexity. Site Offshore is also relatively homogenous, with a gentle slope starting at 10 m and continuing to 12 m where Pleistocene bedrock continues to a depth of 18 m, after which the substrate becomes patchier with sand and bedrock formations as it slopes off into the deep (> 50 m). Due to limitations related to access to sites, time, and funding constrains, only two sites could be studied over the 2 years.
Results derived from the CORTAD dataset illustrated that over the 35 years (1982–2017), the minimum SST experienced by Site Nearshore and Site Offshore were 15.5 °C and 16.5 °C, respectively, while the maximum SST was 37.0 °C at Site Nearshore and 36.2 °C at Site Offshore. The annual mean temperature at Site Nearshore was 26.0 ± 0.1 °C and Site Offshore was 26.2 ± 0.1 °C. Trends in the accumulated thermal stress [expressed as degree heating weeks (DHW)] shows that prior to the 1998 bleaching event, all DHWs were constantly lower than 4 °C-weeks. However, since 1998, DHWs were recorded to reach ≥ 8 °C-weeks with an exception observed in 1998 and 2017 where DHW reached ≥ 15 °C-weeks (Fig. 2). In addition, the bleaching threshold at Site Nearshore was calculated to be 34.5 °C and 34.1 °C at Site Offshore. It is important to note that these values were calculated based on the thermal stress metrics obtained from satellite data instead of in situ measurements. Given this, the 2017-bleaching event experienced on the reefs in Bahrain is expected to have taken place between July and October 2017 where temperatures exceeded 34 °C for ≥ 15 °C-weeks (Fig. 2). The surveys for this study were undertaken in April/May 2017 (pre-bleaching) and 2018 (post-bleaching).
Quantifying carbonate budget states
Gross carbonate production and erosion rates were calculated to determine net carbonate production (kg CaCO3 m−2 year−1 hereafter expressed as G, while ± represents the SE of all values stated) using the international standardised census-based ReefBudget methodology (Indo-Pacific version) (Perry et al. 2018a, b) hereafter referred to as ReefBudget methodology. The natural reef topography in Bahrain is gently sloping (< 2 m), and thus, all transects were conducted on the bottom flat of the reef at a depth of ~ 5–7 m at Site Nearshore and ~ 10 m at Site Offshore.
At each site, data were collected along 6 × 10 m replicate transects laid parallel to each other with a spacing of 5 m between each transect both in 2017 and 2018. Data were collected from each transect to assess benthic substrate composition, surface rugosity, eroder populations, specifically echinoid, scaridae, and macro- and microbioeroders as per the ReefBudget methodology outlined in Perry et al. (2018a, b) (Table 2). Benthic substrate composition was recorded using the LIT method where each benthic component present directly under the transect line was recorded along with its linear length, while rugosity was calculated from the total surface distance for each linear 1 m of reef using a flexi-tape.
Quantifying carbonate production
Primary carbonate producers
To calculate carbonate production rates, primary producers (Scleractinian coral colonies) when encountered beneath the transect line were: (a) identified to genus level, e.g., Porites spp., (b) measured for colony size [calculated based on linear coverage (length × width) using a flexi-tape and categorized based on morphology]. Although the Reefbudget methodology advises that 3D coral colony measurements should be used for more accurate colony measurements, 2D colony measurements were taken for this study due to field limitations both terms of time and human resources. To date, there are no published growth rates or skeletal densities for corals in the Arabian Gulf, with the exception of two species (Platygyra daedalea and Cyphastrea microphthalma) (Howells et al. 2018). Therefore, coral growth rates and skeletal density data used in this study were those provided in the open source “Indo-Pacific data entry spreadsheets” of the ReefBudget methodology which are derived from published literature. The data were then inserted in the spreadsheets that have been set up with standardized Reefbudget equations that automatically calculate the carbonate production rates for each transect. Datasheets can be downloaded for use through: https://geography.exeter.ac.uk/reefbudget/
Secondary carbonate producers
Secondary carbonate production rates were quantified based on the weight per unit area derived using six PVC pipes which were deployed in September 2017 for a period of 1 year at each study site measuring 35 cm × 16 cm (length x circumference of the pipe; Fig. 3). Following the 1-year period, pipes were photographed underwater, placed in a plastic bag secured with cable ties over the upper part of the pipe for removal. All pipes were successfully retrieved using the ReefBudget methodology from Site Nearshore (n = 6); however, only one out of six pipes was found and retrieved at Site Offshore. Evidence of illegal destructive fishing practices, including trawling was witnessed at the site which could have resulted in the displacement and loss of the other pipes. Once in the lab, pipes were examined and photographed in detail following which they were placed in 10% sodium hypochlorite (bleach) for 36 h. Once removed from the bleach, the pipes were left to dry after which, they were weighed three times. The pipes were then soaked in 10% HCl for another 36 h. Once all the calcium carbonate was dissolved, the pipes were taken out and weighted for a second time (in replicates of three). This allowed for a weight per unit area to be derived. To calculate secondary carbonate production, the weight per unit area previously derived through the 1-year experiment was inserted in the Reefbudget calculation spreadsheets in the growth rates column thereby automatically including it in the overall production budget calculation. The Reefbudget methodology uses locally derived data on the rate of total secondary carbonate production in addition to the data on available substrate for secondary producers (e.g., CCA) recorded by the benthic transects (Perry et al. 2018a, b).

CaCO3 production by secondary carbonate producers over a 1-year period on PVC pipes deployed at Nearshore (Fasht Al Adhm). NB: exposed = high light, cryptic = low light
Quantifying carbonate erosion
For calculating carbonate erosion rates, species, density, and size class measurements were recorded for grazers, such as echinoids (urchins) and Scaridae (parrotfish) along the belt transect area.
-
Echinoids: The area surveyed along each of the six transects was 1 m on either side of the 10 m transect line covering a total of 20 m2 per transect (10 m length × 2 m width). The number of individuals along each transect was counted, identified to species level and recorded in the following size class which is the width of the echinoid without the spines: 0–20 mm, 21–40 mm, 41–60 mm, 61–80 mm, and 81–100 mm. Data were then inputted into the Reefbudget calculation spreadsheets which automatically calculates the carbonate erosion rate of echinoids based on species level.
-
Scaridae: The area surveyed along each of the transects (3 replicates in 2017 and 6 replicates in 2018) was 4 m in width of the 30 m transect line covering a total of 120 m2 per transect (30 m length × 4 m width). The number of individuals encountered along each transect were counted, identified to species level and recorded in the following size class based on total length: 5–10 cm, 11–20 cm, 21–30 cm 31–40 cm, and > 40 cm along with their life phase (i.e., juvenile or adult). Biomass of parrotfish was calculated using published species-specific length–weight relationships as provided by the Reefbudget calculation spreadsheets where the data were inputted for carbonate erosion rate contributed by parrotfish.
-
Macroborers: The Reefbudget (Indo-Pacific version) defines macroborers as those that produce boreholes with diameters of > 1 mm. These include endolithic sponges, bivalves, decapods, polychaete and sipunculid worms, and cirripeds, of these, sponges dominate the macroboring community, comprising 75–90% by proportion of substrate infestation (Perry al. 2018). In general, the macroborer community is less characterized than that of the Caribbean in both the Indian and Pacific Oceans, many bioeroders especially clionaid sponges are cryptic in general making their identification difficult on the field. In addition, macroerosion measurements tend to rely on deploying of experimental substrates (e.g., coral blocks) and/or using X-rays or CT scans to estimate internal rates of bioerosion using cored or slabbed corals (Perry et al. 2018a, b). Therefore, the Reefbudget methodology uses published rates of the total macrobioerosion with data on available substrate for bioerosion (including dead carbonate substrate and live coral colonies) derived from the benthic transects (Perry et al. 2018a, b). Thus, macrobioerosion rates are automatically calculated in the Reefbudget calculation spreadsheets using the benthic substrate data.
-
Microborers: Endolithic microborers such as cyanobacteria, cholorophytes, and fungi are able to exist inside carbonate substrates. Similar to macrobioerosion, the assessment of microbioerosion rates tend to rely on deploying of experimental substrates (Chazottes et al. 1995; Tribollet and Golubic 2005). The Reefbudget methodology acknowledges data scarcity on microbioerosion processes and despite the challenges in measuring microbioerosion on the field, it recognizes that microborers can have the potential to contribute to bioerosion on reefs in a non-negligible amount. Thus, since published rates on microbioerosion are within similar ranges to those of macroborers, the rates are automatically calculated in the Reefbudget calculation spreadsheets using the benthic substrate data.
Data analysis
Data processing, analysis, and visualization were conducted in the software “R” and “RStudio” version 3.5.1 (R Core Team 2018) utilizing the following R packages: ggplot (Hothorn et al. 2008), tidyverse (Wickham et al. 2019), stats package (R Core Team 2021), and multcomp package (Hothorn et al. 2008). Mean ± standard error (SE) is stated throughout along with the median (md) values for primary producers, gross carbonate production, and gross carbonate erosion. Using the glm function in stats package, generalized linear models (GLMs) were used to test the effects of bleaching events on the percentage cover of carbonate producers (i.e., hard coral cover and CCA), rugosity, density of eroders (i.e., Scaridae and echinoids), gross carbonate production, gross carbonate erosion, and net carbonate production at both sites (Supplementary Tables 1 and 2). Likelihood ratio tests (LRT) were conducted to analyze the deviance between null and alternative models to determine the effect of the interaction between bleaching events and sites on the observations using the anova function in the stats package (Supplementary Table 3). The interaction term was included in the model if LRT was found to be significant. Post hoc comparison tests were implemented using the glht function in the multcomp package (Hothorn et al. 2008) with all significant test results reported in the text and additional results presented in Supplementary Table 4.
Results
Site characterization
Carbonate producers
Dominant hard corals recorded at both sites included Platygyra spp., Favia spp., and Porites spp., all of which were either massive, sub-massive, or encrusting with no branching corals observed at any of the study sites either year (with the exception of one colony of Pocillopora sp. seen in 2017 at Site Nearshore). Hard coral cover declined from 13.6 ± 7.0%, pre-bleaching to 1.4 ± 0.6%, post-bleaching at Site Nearshore, while Site Offshore maintained its coral cover (pre-bleaching = 14.8 ± 4.1%, n = 6; post-bleaching = 15.5 ± 2.2%); however, no significant difference was observed between sites in 2017 (Site Nearshore: GLM, z = − 2.1, p > 0.05; Site Offshore: GLM, z = 0.1, p > 0.05) (Figs. 4, 5 and 6a). No branching coral colonies were observed at either study site in 2018 and no change in the coral community was observed as the dominant coral groups remain to be either massive, sub-massive, or encrusting.

Example photographs of reefscapes pre- and post-bleaching at A Fasht Al Adhm (Site Nearshore) and B Reef Bul Thamah (Site Offshore), Bahrain. NB: Photos were taken in September, 2017 while the bleaching period is estimated to have taken place from July to October 2017 based on satellite data derived from the CORTAD version 6 database

Mean percentage cover (%) of the reef benthic community structure pre (2017) and post (2018) bleaching at Site Nearshore and Site Offshore (n = 6), Bahrain

Percentage cover of carbonate producers in 2017 and 2018: A Hard corals (n = 6) and B CCA = crustose coralline algae (n = 6); density (count per unit m2) of carbonate eroders: C Echinoids (n = 6; area = 20 m2 per transect; Nearshore) and D Scaridae (n = 3; area = 120 m2 per transect) at Site Nearshore and Site Offshore, Bahrain
CCA cover was dominant at Site Nearshore pre-bleaching (mean = 42.2 ± 12.9%, md = 38.9); however, this declined significantly to 13.2 ± 3.5%, md = 14.5 (GLM, z = − 2.7, p < 0.05) post-bleaching (Figs. 5 and 6b). CCA was also observed to decline at Site Offshore post-bleaching (pre-bleaching = 10.1 ± 7.0%, md = 4.2; post-bleaching = 1.73 ± 0.4%, md = 1.8) but was statistically non-significant (Fig. 6b—Supplementary Material, Table 4). No change in reef rugosity was noted in Site Offshore (pre-bleaching = 1.8 ± 0.14; post-bleaching = 1.7 ± 0.04), while a decline in rugosity was noted in Site Nearshore (pre-bleaching = 1.7 ± 0.17; post-bleaching = 1.4 ± 0.05). The largest dominating benthic group at Site Offshore was algae, including turf algae (pre-bleaching = 25.5 ± 7.6%, post-bleaching = 34.4 ± 6.6%) and macroalgae (mainly Halimeda sp. and Padina sp.; pre-bleaching = 19.6 ± 3.5%; post-bleaching = 23.4 ± 2.1%) (Fig. 5). At Site Nearshore, macroalgae (mainly Halimeda sp.; pre-bleaching = 2.7 ± 1.5%, post-bleaching = 3.6 ± 1.7%) was seen to be low in comparison to Site Offshore in both years. However, turf algae was higher at Site Nearshore with an increase of 50% post-bleaching (pre-bleaching = 31.5 ± 14.2%, post-bleaching = 63.1 ± 6.0%) (Fig. 5). The remaining benthic substrate for both Site Nearshore and Site Offshore was composed of sand, rubble, sponge, soft coral, and other (e.g., other calcareous encrusters and ascidians) (Fig. 5).
Carbonate eroders
At Site Nearshore, echinoid densities were higher pre-bleaching (25.0 ± 6.4 individuals per 20 m2) than post-bleaching (8.0 ± 2.5 individuals per 20 m2) with a significant reduction of 68% (GLM, z = 3.6, p < 0.01) (Fig. 6c), but echinoids were observed to be larger post-bleaching, i.e., pre-bleaching: size class = 41–60 mm and 61–80 mm; post-bleaching: size class = 61–80 mm and 81–100 mm. Echinometra mathei (short-spined urchin) was the only species of echinoid recorded along the transects at Site Nearshore throughout the study period and no parrotfish were observed either pre- or post-bleaching (Fig. 6d). At Site Offshore, no echinoids were observed; however, parrotfish of the species Chlorurus sordidus, Scarus ghobban, and Scarus persicus were recorded pre-bleaching (17.0 ± 3.8 individuals per 120 m2), which were mainly scrapers in their initial phase of life; contrastingly, very few individuals were observed post-bleaching (1.0 ± 0.5 individuals per 120 m2) (Fig. 6d).
Carbonate budgetary state|pre- and post-bleaching (2017–2018)
Gross carbonate production rates decreased at Site Nearshore from 4.1 ± 2.3 G pre-bleaching to 0.5 ± 0.2 G post-bleaching, whereas Site Offshore maintained its gross carbonate production (pre-bleaching = 4.0 ± 1.1 G; post-bleaching = 4.2 ± 0.6 G) (Fig. 7a). No significant differences in gross carbonate production rates were detected in both Site Nearshore (GLM, z = − 1.9, p > 0.01) and Site Offshore (GLM, z = 0.1, p > 0.01). In addition, the calcification rate for secondary producers derived from the 1-year experiment was 0.027 g cm2 year−1 at Site Nearshore. In terms of gross carbonate erosion, Site Nearshore had a higher rate of gross erosion with 0.5 ± 0.1 G pre-bleaching which was seen to significantly decrease to 0.2 ± 0.1 G post-bleaching (GLM, z = − 3.4, p < 0.01) (Fig. 7 (b)). The majority of erosion at Site Nearshore is dominated by echinoids with negligible contribution by macroborers, while at Site Offshore, erosion was dominated by parrotfish which was observed to be minimal especially post-bleaching (Supplementary Table 5). Site Offshore did experience a decline in gross carbonate erosion from the pre-bleaching value of 0.3 ± 0.1 G to post-bleaching value of 0.01 ± 0.01 G (GLM, z = − 3.0, p < 0.01) (Fig. 7 (b)).

Reef carbonate budget rates (kg CaCO3 m−2 year−1 expressed as G) across study sites Nearshore and Offshore pre- and post-2017 bleaching illustrating: A gross carbonate production; B gross carbonate erosion; C net carbonate production, Bahrain
Overall, a decrease in the net carbonate production rates was recorded on Site Nearshore, while Site Offshore reported an increase. Specifically, Site Nearshore showed a decline in net carbonate production rate following the 2017-bleaching (pre = 3.6 ± 2.2; post = 0.3 ± 0.3 G) indicating a shift from a positive budget state to a nearly a net neutral budget state (Fig. 7c). Contrastingly, Site Offshore documented an increase in its net carbonate production from 3.7 ± 1.2 to 4.2 ± 0.6 G, therefore maintaining its positive budgetary state. Both sites did not experience significant differences in net carbonate production rates (Site Nearshore: GLM, z = − 1.8, p > 0.01); Site Offshore: GLM, z = 0.3, p > 0.01).
Discussion
This study quantified carbonate budgets for two reefs in Bahrain. Here, we report on the impacts observed on the budgetary state of these reefs as a result of the 2017 warming event, which is the most intense bleaching event experienced by these reefs since 1998.
Carbonate production
Hard coral (primary reef producers) cover declined at Site Nearshore between 2017 and 2018. In 2012, it was reported that the live coral coverage at this site averaged 9.8% (Burt et al. 2013). Our results from 2017 indicated that this reef had recovered somewhat between 2010 and 2017, with coral cover at 13.6 ± 6.9% despite facing two bleaching events in that duration. Our observations are in line with what has been reported in the latest Global Coral Reef Monitoring Network (GCRMN) report (2020) which states that live coral in AG has experienced a decline from 30.1 to 18.0% between 1997 and 2002, followed by a period of recovery (2002–2015) when coral cover recovered to 30.2%, almost comparable to the pre-bleaching period. Although there are not much published data which is exclusive or directly focused on the impacts of the bleaching events between 2010 and 2017, consequences of bleaching on reefs in the North of the AG have been reported by Kavousi et al. (2021) where coral mortality between 81 and 100% was experienced due to consecutive bleaching (2014–2017), while the south of the AG experienced a loss of 73% of coral cover in one year (2017–2018) (Burt et al. 2019). Currently, there are no published data on the bleaching rates on Bahraini reefs. Unfortunately, following the 2017-bleaching event, coral cover decreased to 1.4 ± 0.6% at the nearshore site. One of the factors that could have contributed toward this decline could be the shallow depth of the reef (< 7 m) which could subject the reef to warmer waters and higher erosion rates than those occurring at deeper reefs, since bioerosion is dependent on numerous environmental factors including depth and light availability (Chazottes et al. 1995; Mallela 2007). In nearshore reefs that are subjected to higher wave action, turbid conditions are expected especially if the shoreline is low and flat. Wave action influences the sedimentary processes on these reefs, and thus, turbidity induced by the suspended sediment reduces the light level corals are exposed to. This could lead coral communities to be dominated by massive and sub-massive coral groups, thereby transiting the reef from a high accretion reef due to its vertical 3-D framework growth to one that is of low accretion like the reefs in Bahrain. It has been previously mentioned that carbonate erosion and dissolution rates on reefs that exist with low light levels (including mesophotic reefs) exceed carbonate production (Hoegh-Guldberg et al. 2017). In addition, nearshore reefs are shallower and hence are associated with warmer waters. Increased temperatures combined with more acidic conditions have been documented to trigger responses from bioeroding organisms such as microbioeroders which were seen to enhance skeletal dissolution due to increased biomass (Reyes-Nivia et al. 2013) and bioeroding sponge (such as Cliona orientalis) were previously reported to have increased both in biomass and bioerosion (Fang et al. 2013; Hoegh-Guldberg et al. 2017). This coupled with bleaching episodes that reduce calcification rates could subject nearshore reefs to higher erosion reefs. Furthermore, land-based runoff could lead to overproliferation of turf algae and macroalgae, increasing the abundance of echinoids (Kriegisch et al. 2020). Nevertheless, the recovery witnessed prior to the 2017-bleaching event creates a positive outlook for the post-2017-bleaching period as temporal data indicate the reef’s ability for recovery despite encountering multiple bleaching events in previous years (e.g., 2010; Fig. 2). Nevertheless, it is evident that fundamental changes have occurred to the ecology of this shallow reef. Prior to the 2017 warming event, over 50% of the reef was dominated by primary and secondary producers; however, by 2018, these groups covered only 30% of the reef substrate. On the contrary, Site Offshore (> 10 m) demonstrated a 2% increase in hard coral cover reaching 14.8 ± 4.1% in 2018, maintaining its coral cover, which was reported to be 16.3% in 2012 (Burt et al. 2013). It is important to note that in addition to thermal anomalies, reefs in Bahrain are known to be impacted by various anthropogenic activities such as reclamation and dredging which has contributed significantly toward diminishing coral cover at Site Nearshore (Naser 2012; Burt et al. 2013).
It is of particular note that no branching or tabular coral taxa (e.g. Acropora spp.) were observed throughout the study period at either site as this has major ecological implications on both habitat diversity and complexity, as well as overall trends in reef accretion and budgetary dynamics. Branching corals such as Acropora spp. are responsible for the majority of calcification and structural complexity due to their morphology and rapid growth (e.g., Lizard Island, GBR = ~ 7.31 cm year−1; Anderson et al. 2017). Acroporids are, therefore, an important functional genus in terms of structural complexity, which contributes toward habitat provisioning for reef associated species (Alvarez-Filip et al. 2013; Anderson et al. 2017; Roff 2020), as well as contributing to reef accretion and enhancing the ability for reefs to track sea-level rise (SLR). The disappearance of fast growing, branching corals such as Acropora spp. from Bahraini reefs since 1998, is exacerbating reef framework decline and has resulted in shifts from these competitive corals (i.e., fast growing and branching) to opportunistic and stress-tolerant corals (i.e., low growing, thermally tolerant, e.g., Porites spp. and Siderastrea spp.) thereby compromising reef structural integrity. These shifts have also been documented in the Indian Ocean (Perry and Morgan 2017) and Caribbean (Baumann et al. 2016).
In general, the morphological and physiological attributes of reef associated species are closely linked to the coral assemblages and their life-history strategies which is key to the functionality of coral reefs (Darling et al. 2012; Alvarez-Filip et al. 2011). Changes in the functionality of reef associated fish have been documented with links associating coral cover decline to fish diversity decline, thereby impacting various functional groups, e.g., obligate corallivores associated with tabulate Acropora spp. (Pratchett et al. 2011). This was evident on these Bahraini reefs as many functional groups were observed to either be absent, e.g., obligate corallivores (Chaetodon sp.) or represented by only a handful of species. Moreover, many reefs reported shifts in coral and associated species community composition, e.g., shifts toward sponge-dominated reefs in Indonesia (Bell et al. 2013) and shifts toward massive stress-tolerant reefs in the Maldives, e.g., Porites spp., Goniopora spp., and Platygyra spp. (Ryan et al. 2019). This is evident on Bahraini reefs with the community composition shifting from primary carbonate producers associated with massive stress-tolerant corals to algal cover dominated communities. Reefs dominated by massive stress-tolerant corals could impact fish recruitment, diversity, and abundance due to limited resources and lack of structural complexity. This in turn tends toward a decrease in species richness, which could accelerate the collapse of the reef ecosystem. It is an accepted view that ecosystems with high species richness have a higher resilience to environmental change due to increased diversity of functional traits that facilitate successful recruitment of both coral and fish, thereby enabling reef resilience and recovery (Emslie et al. 2014; Hughes et al. 2012; Pratchett et al. 2011; Yachi and Loreau 1999). In addition, the shifts from competitive corals to opportunistic and stress-tolerant corals impact the ability of reefs to function as natural barriers through decreasing their efficiency to absorb wave energy and compromises coastal protection (Hoegh-Guldberg et al. 2007).
Interestingly, CCA percentage cover was extremely high at Site Nearshore pre-bleaching (mean = 42.2 ± 12.9%), perhaps due to the presence of high rubble cover caused by previous dredging activities in the area (Vousden 1988). In addition to being an important carbonate producer, CCA plays a key role in binding the reef through colonizing reef substrate and contributing to cementation and stability (Rasser and Riegl 2002; Langer 2008; Perry and Hepburn 2008; Fujita et al. 2009). This promotes accretion, enhances larval recruitment and maintains wave-resistant reef fronts (Rasser and Riegl 2002; Mallela and Perry 2007). During the field surveys in April/May 2017, extensive rubble binding was witnessed following what was suspected to be a dredging event (c. 10 years), and this could have contributed to the rise in coral cover between the period 2010 and 2017. Following the bleaching event, the reef has experienced a change from CCA domination to turf algae which increased by 50% post-bleaching. This finding aligns with other studies in various locations, such as Hawaii (Foo et al. 2022) and New Caledonia (Tanvet et al. 2022), all of which is attributed to SST warming (Cornwall et al. 2019). Therefore, the high presence of CCA is evidence that this reef was in recovery but has been affected by this warming event which is impacting the recovery process (Teichert et al. 2020; Roth et al. 2018). Most importantly, it has been demonstrated that CCA can match or exceed the contribution of corals to reef carbonate production; however, they are often inaccurately recorded in benthic surveys and sometimes missed in the reef carbonate budget analysis (Cornwall et al. 2023). Finally, the calcification rate (i.e., 0.027 g cm2 year−1) for secondary producers derived from the 1-year experiment was observed to be similar to those recorded in the Indo-Pacific (Perry et al. 2018a, b).
Bioerosion
Erosion rates in Bahrain both pre- and post-bleaching (0.4–0.1 and 0.1–0.1 G, respectively) are generally lower than other regions, e.g., Caribbean = 1.0–3.7 G (Perry et al. 2014); South China Sea = 0.1–0.7 G (Dumont et al. 2013), and the Red Sea = 1.9–2.9 G (Roik et al. 2017). This could be due to lower eroder (fish and urchin) diversity and abundance and could be attributed to lower sedimentation offshore, and other environmental factors (Tribollet and Golubic 2005 at Great Barrier reef; Morris et al. 2022 at Florida Keys.
Similarly, parrotfish were observed only at Site Offshore pre-bleaching, in low numbers, while none were found at Site Nearshore. This could be attributed to extensive overfishing activities at both sites (Morgan 2006) as despite Site Offshore being a marine national park, there is no law enforcement or patrol in the area. In addition, lower erosion rates by parrotfish in the AG could be linked to lower fish biomass (e.g., D’Agostino et al. 2021 and Feary et al. 2010). Erosion rates in Bahrain caused by macroborers (dominated by bioeroding sponges) are negligible and fall well below other regions such as Brazil (0.2–3.0 G) (Reis and Leão 2000), Bonaire (Caribbean) (0.01–0.1 G) (Perry et al. 2012), and Gulf of Aqaba (Red Sea) (0.34 G) (Zundelevich et al. 2007). However, we suggest further examining the contribution of macroborers and microborers to reef erosion, as bioerosion was previously reported to be high in the AG with bioeroders removing 9.2 ± 1.6% of the skeletal surface area of Porites daedalea and 26.4 ± 1.6% in Cyphastrea microphthalma with variation in colony morphology being attributed to the differences between species (Al-Mansoori et al 2019). Bioerosion in the AG appears to be related to environmental stress and past bleaching exposure (Al-Mansoori et al 2019).
Carbonate budgets
Thermal anomalies are known to reduce calcification rates in coral (De’ath et al. 2009) and, therefore, carbonate production and reef accretion rates. This is observed in a 20 year dataset from the Seychelles, which showed a 62.5% difference in reef carbonate budget rates post-bleaching (− 1.5 G (erosive state)) compared to pre-bleaching (~ 4 G (positive state); Januchowski-Hartley et al. 2017). Our results show a similar trend on the shallow nearshore reefs of Bahrain, highlighting a decrease by 88% (pre = 3.6 ± 2.2; post = 0.3 ± 0.3 G), while the deeper offshore reefs reported an increase in budgetary state (3.7 ± 1.2 to 4.2 ± 0.6 G) which could be attributed to the decrease of erosion rate post-bleaching at Site Offshore. The near collapse of the reef budget at Site Nearshore is directly linked to the widespread mortality of corals resulting from the thermal anomaly episode in 2017 (Fig. 2). It is worth noting that massive stress-tolerant corals have been reported to be able to maintain positive carbonate production on reefs, e.g., reefs in the Maldives (Ryan et al. 2019) despite their comparative lack of vertical growth, which was evident at Site Nearshore prior to September 2017; however, following the 2017-bleaching event, we report a shift to nearly a negative state in the reef carbonate budget for this nearshore site. It is important to note that in this study, there may have been an over-estimation of coral growth rates, since rates used for the ReefBudget calculation were not site specific due to unavailable published literature on coral growth rates in the AG, with the exception of growth rates of two species (Platygyra daedalea and Cyphastrea microphthalma; Howells et al. 2018). Similarly, CCA and other secondary producers growth rates could be overestimated as utilizing different experimental substrates could yield different results (Mallela et al. 2017). In addition, it is important to note that the ReefBudget method does not take into account the physical loss and chemical dissolution rates, and hence, this could also be an additional cause for overestimation of the reef budget rates. In general, Bahraini reef budget rates (including pre- and post-bleaching) (range = 0.3–4.2 G) were seen to be in the lower scale, most similar to those in the Caribbean (e.g., Jamaica = 1.1 G (Land 1979) and Barbados = 4.5 G (Scoffin et al. 1980)) and the Indian Ocean (e.g., Maldives = 2.5 ± 2.5 G (Ryan et al. 2019) and Seychelles 4 G (pre-bleaching) and − 1.5 G (post-bleaching) (Januchowski-Hartley et al. 2017)) in comparison to reefs in the Indo-Pacific, where, for example, net carbonate production were reported to be as high as 22.6 G in some areas of the Great Barrier Reef (Brown et al. 2021). Both sites retained low gross erosion rates accompanying the low gross production rates.
In conclusion, to our knowledge, no study has been conducted to quantify carbonate budgets of reefs in Bahrain or AG making this the first attempt. Reefs in AG are unique, particularly in relation to the capability of corals to sustain themselves in such extreme environmental conditions with seasonal temperature fluctuations. However, SST warming events, which are projected to increase in frequency and intensity with climate change, may jeopardize the resilience of these reefs and impacting their ecological functionality. Based on these trends, we predict suppressed budgetary states in AG. This, along with associated IPCC projected sea-level rise, will limit the capacity of Bahraini reefs to perform their natural breakwater function and therefore threaten island stability.
Data availability
The datasets generated and analyzed for this study can be found in the University of Essex Repository (https://dx.doi.org/10.5526/ERDR-00000192).
References
Alkhuzai J, Sheppard C, Abdulqader E, Al-Khuzai S, Loughland R (2009) Subtidal habitats. In: Loughland R, Zainal A (eds) The marine atlas of Bahrain. Miracle Publishing, Manama, Bahrain, pp 113–168
Al-kuwari Z (2006) Bahrain first national report to the convention on biological diversity Manama, Bahrain. https://www.sce.gov.bh/Media/Downloads/reports/pdf/report-1.pdf. Accessed 20 May 2022
Al-Mansoori N, McParland D, Howells E, Bauman A, Burt JA (2019) Coral bioerosion on the marginal reefs of northeastern Arabia. Aquat Ecosyst Health Manag 23(2):187–195. https://doi.org/10.1080/14634988.2019.1677204
Alvarez-Filip L, Dulvy NK, Gill JA, Côté IM, Watkinson AR (2009) Flattening of Caribbean coral reefs: region-wide declines in architectural complexity. Proc R Soc B Biol Sci 276:3019–3025. https://doi.org/10.1098/rspb.2009.0339
Alvarez-Filip L, Dulvy NK, Côté IM, Watkinson AR (2011) Coral identity underpins reef complexity on Caribbean reefs. Ecol Appl 21:2223–2231. https://doi.org/10.1890/10-1563.1
Alvarez-Filip L, Carricart-Ganivet JP, Horta-Puga G, Iglesias-Prieto R (2013) Shifts in coral-assemblage composition do not ensure persistence of reef functionality. Sci Rep 3:3486. https://doi.org/10.1038/srep03486
Anderson KD, Cantin NE, Heron SF, Pisapia C, Pratchett MS (2017) Variation in growth rates of branching corals along Australia’s Great Barrier Reef. Sci Rep 7:2920. https://doi.org/10.1038/s41598-017-03085-1
Anthony KRN, Kline DI, Diaz-Pulido G, Dove S, Hoegh-Guldberg O, Karl DM (2008) Ocean acidification causes bleaching and productivity loss in coral reef builders. Proc Natl Acad Sci USA 105(45):17442–17446. https://doi.org/10.1073/pnas.0804478105
Baker AC, Starger CJ, McClanahan TR, Glynn PW (2004) Coral reefs: corals’ adaptive response to climate change. Nature 430:741. https://doi.org/10.1038/430741a
Baumann JH, Townsend JE, Courtney TA, Aichelman HE, Davies SW, Lima FP, Castillo KD (2016) Temperature regimes impact coral assemblages along environmental gradients on lagoonal reefs in Belize. PLoS ONE 11:1–19. https://doi.org/10.1371/journal.pone.0162098
Bell JJ, Davy SK, Jones T, Taylor MW, Webster NS (2013) Could some coral reefs become sponge reefs as our climate changes? Glob Change Biol 19:2613–2624. https://doi.org/10.1111/gcb.12212
Berkelmans R, Oliver JK (1999) Large-scale bleaching of corals on the Great Barrier Reef, vol 18. Springer, Berlin, pp 55–60
Brown KT, Bender-Champ D, Achlatis M, van der Zande RM, Kubicek A, Martin SB, Castro-Sanguino C, Dove SG, Hoegh-Guldberg O (2021) Habitat-specific biogenic production and erosion influences net framework and sediment coral reef carbonate budgets. Limnol Oceanogr 66:349–365. https://doi.org/10.1002/lno.11609
Browne N (2011) A carbonate and terrigenous sediment budget for inshore turbid reefs on the Great Barrier Reef. PhD thesis, James Cook University
Burt J, Bartholomew A, Feary D (2012) Man-made structures as artificial reefs in the Gulf. In: Riegl BM, Purkis SJ (eds) Coral reefs of the Gulf. Springer, Dordrecht, pp 171–186. https://doi.org/10.1007/978-94-007-3008-3
Burt JA, Al-Khalifa K, Khalaf E, AlShuwaikh B, Abdulwahab A (2013) The continuing decline of coral reefs in Bahrain. Mar Pollut Bull 72:357–363. https://doi.org/10.1016/j.marpolbul.2012.08.022
Burt JA, Paparella F, Al-Mansoori N, Al-Mansoori A, Al-Jailani H (2019) Causes and consequences of the 2017 coral bleaching event in the southern Persian/Arabian Gulf. Coral Reefs 4:567–589. https://doi.org/10.1007/s00338-019-01767-y
Chazottes V, Campion-Alsumard TL, Peyrot-Clausade M (1995) Bioerosion rates on coral reefs: interactions between macroborers, microborers and grazers (Moorea, French Polynesia). Palaeogeogr Palaeoclimatol Palaeoecol 113:189–198. https://doi.org/10.1016/0031-0182(95)00043-L
Choi DR, Ginsburg RN (1983) Distribution of coelobites (cavity-dwellers) in coral rubble across the Florida Reef Tract. Coral Reefs 2:165–172
Coles SL, Riegl BM (2013) Thermal tolerances of reef corals in the Gulf: a review of the potential for increasing coral survival and adaptation to climate change through assisted translocation. Mar Pollut Bull 72:323–332. https://doi.org/10.1016/j.marpolbul.2012.09.006
Cornwall CE, Diaz-Pulido G, Comeau S (2019) Impacts of ocean warming on coralline algal calcification: meta-analysis, knowledge gaps, and key recommendations for future research. Front Mar Sci 6:186. https://doi.org/10.3389/fmars.2019.00186
Cornwall CE, Carlos J, Branson O, Courtney TA, Harvey BP, Perry CT, Andersson AJ, Diaz-Pulido G, Johnson MD, Kennedy E, Krieger EC, Mallela J, McCoy SJ, Nugues MM, Quinter E, Ross CL, Ryan E, Saderne V, Comeau S (2023) Crustose coralline algae can contribute more than corals to coral reef carbonate production. Commun Earth Environ 4(1):105. https://doi.org/10.1038/s43247-023-00766-w
D’Agostino D, Burt JA, Santinelli V, Vaughan GO, Fowler AM, Reader T, Taylor BM, Hoey AS, Cavalcante GH, Bauman AG, Feary DA (2021) Growth impacts in a changing ocean: insights from two coral reef fishes in an extreme environment. Coral Reefs 40:433–446. https://doi.org/10.1007/s00338-021-02061-6
Darling ES, Alvarez-Filip L, Oliver TA, McClanahan TR, Côté IM (2012) Evaluating life-history strategies of reef corals from species traits. Ecol Lett 15:1378–1386. https://doi.org/10.1111/j.1461-0248.2012.01861.x
de Bakker DM, van Duyl FC, Perry CT, Meesters EH (2019) Extreme spatial heterogeneity in carbonate accretion potential on a Caribbean fringing reef linked to local human disturbance gradients. Glob Chang Biol 25:4092–4104. https://doi.org/10.1111/gcb.14800
De’ath G, Lough JM and Fabricius KE, (2009) Declining coral calcification on the Great Barrier Reef. Science 323:116–119. https://doi.org/10.1126/science.1165283
Dumont CP, Lau DCC, Astudillo JC, Fong KF, Chak STC, Qui JW (2013) Coral bioerosion by the sea urchin Diadema setosum in Hong Kong: susceptibility of different coral species. J Exp Mar Biol Ecol 441:71–79. https://doi.org/10.1016/j.jembe.2013.01.018
Emslie MJ, Cheal AJ, Johns KA (2014) Retention of habitat complexity minimizes disassembly of reef fish communities following disturbance: a large-scale natural experiment. PLoS ONE 9(8):e105384. https://doi.org/10.1371/journal.pone.0105384
Fang JKH, Mello-Athayde MA, Schoenberg CHL, Kline DI, Hoegh-Guldberg O, Dove S (2013) Sponge biomass and bioerosion rates increase under ocean warming and acidification. Glob Change Biol 19:3581–3591. https://doi.org/10.1111/gcb.12334
Feary DA, Burt JA, Bauman AG, Usseglio P, Sale PF, Cavalcante GH (2010) Fish communities on the world’s warmest reefs: what can they tell us about the effects of climate change in the future? J Fish Biol 77(8):1931–1947. https://doi.org/10.1111/j.1095-8649.2010.02777.x
Foo SA, Teague CH, Asner GP (2022) Warming alters the relationship between benthic cover and herbivores on Hawaiian reefs. Front Mar Sci 9:787314. https://doi.org/10.3389/fmars.2022.787314
Fujita K, Osawa Y, Kayanne H, Ide Y, Yamano H (2009) Distribution and sediment production of large benthic foraminifers on reef flats of the Majuro Atoll, Marshall Islands. Coral Reefs 28:29–45. https://doi.org/10.1007/s00338-008-0441-0
Goreau TF (1963) Calcium carbonate deposition by coralline algae and corals in relation to their roles as reef builders. Ann N Y Acad Sci 109:127–167
Graham NAJ, Nash KL (2013) The importance of structural complexity in coral reef ecosystems. Coral Reefs 32:315–326. https://doi.org/10.1007/s00338-012-0984-y
Hepburn LJ (2006) Processes of bioerosion and calcareous encrustation on a Mexican Caribbean reef: implications for framework development. PhD thesis, Manchester Metropolitan University, Manchester, United Kingdom
Hepburn LJ, Blanchon P, Murphy G, Cousins L, Perry CT (2015) Community structure and palaeoecological implications of calcareous encrusters on artificial substrates across a Mexican Caribbean reef. Coral Reefs 34:189–200. https://doi.org/10.1007/s00338-014-1227-1
Heron SF, Maynard JA, Van Hooidonk R, Eakin CM (2016) Warming trends and bleaching stress of the world’s coral reefs 1985–2012. Sci Rep 6:1–14. https://doi.org/10.1038/srep38402
Hoegh-Guldberg O (1999) Climate change, coral bleaching and the future of the world’s coral reefs. Mar Freshw Res 50:839–866. https://doi.org/10.1071/MF99078
Hoegh-Guldberg O (2011) Coral reef ecosystems and anthropogenic climate change. Reg Environ Change 11:S215–S227. https://doi.org/10.1007/s10113-010-0189-2
Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ, Caldeira K, Knowlton N, Eakin CM, Iglesias-Prieto R, Muthiga N, Bradbury RH, Dubi A, Hatziolos ME (2007) Coral reefs under rapid climate change and ocean acidification. Science 318:1737–1742
Hoegh-Guldberg O, Poloczanska ES, Skirving W, Dove S (2017) Coral reef ecosystems under climate change and ocean acidification. Front Mar Sci 4:158. https://doi.org/10.3389/fmars.2017.00158
Hoey AS, Howells E, Johansen JL, Hobbs JPA, Messmer V, McCowan DM, Wilson SK, Pratchett MS (2016) Recent advances in understanding the effects of climate change on coral reefs. Diversity 8:12. https://doi.org/10.3390/d8020012
Holmes KE, Edinger EN, Hariyadi, Limmon GV, Risk MJ (2000) Bioerosion of live massive corals and branching coral rubble on Indonesian coral reefs. Mar Pollut Bull 40:606–617. https://doi.org/10.1016/S0025-326X(00)00067-9
Hothorn T, Bretz F, Westfall P (2008) simultaneous inference in general parametric models. Biometr J 50:346–363. https://doi.org/10.1002/bimj.200810425
Howells EJ, Dunshea G, McParland D, Vaughan GO, Heron SF, Pratchett MS, Burt JA, Bauman AG (2018) Species-specific coral calcification responses to the extreme environment of the Southern Persian Gulf. Front Mar Sci 5:56. https://doi.org/10.3389/fmars.2018.00056
Hubbard DK (1986) Sedimentation as a control of reef development: St. Croix, U.S.V.I. Coral Reefs 5:117–125
Hubbard DK, Burke RB, Gill IP (1986) Styles of reef accretion along a steep, shelf-edge reef, St. Croix, U.S. Virgin Islands. J Sediment Petrol 56:848–861
Hughes TP, Baird AH, Bellwood DR, Card M, Connolly SR, Folke C, Grosberg R, Hoegh-Guldberg O, Jackson JB, Kleypas J, Lough JM, Marshall P, Nyström M, Palumbi SR, Pandolfi JM, Rosen B, Roughgarden J (2003) Climate change, human impacts, and the resilience of coral reefs. Science 301:929–933. https://doi.org/10.1126/science.1085046
Hughes TP, Baird AH, Dinsdale EA, Moltschaniwskyj NA, Pratchett MS, Tanner JE, Willis BL (2012) Assembly rules of reef corals are flexible along a steep climatic gradient. Curr Biol 22:736–741. https://doi.org/10.1016/j.cub.2012.02.068
Hughes TP, Kerry JT, Álvarez-Noriega M, Álvarez-Romero JG, Anderson KD, Baird AH, Babcock RC, Beger M, Bellwood DR, Berkelmans R, Bridge TC, Butler IR, Byrne M, Cantin NE, Comeau S, Connolly SR, Cumming GS, Dalton SJ, Diaz-Pulido G, Eakin CM, Figueira WF, Gilmour JP, Harrison HB, Heron SF, Hoey AS, Hobbs JA, Hoogenboom MO, Kennedy EV, Kuo CY, Lough JM, Lowe RJ, Liu G, McCulloch MT, Malcolm HA, McWilliam MJ, Pandolfi JM, Pears RJ, Pratchett MS, Schoepf V, Simpson T, Skirving WJ, Sommer B, Torda G, Wachenfeld DR, Willis BL, Wilson SK (2017a) Global warming and recurrent mass bleaching of corals. Nature 543:373–377. https://doi.org/10.1038/nature21707
Hughes L, Steffen W, Alexander D, Rice M (2017b) Climate change: a deadly threat to coral reefs. Climate Council of Australia Limited, Sydney, p 28
Hume B, D’Angelo C, Burt J, Baker AC, Riegl B, Wiedenmann J (2013) Corals from the Persian/Arabian Gulf as models for thermotolerant reef-builders: prevalence of clade C3 Symbiodinium, host fluorescence and ex situ temperature tolerance. Mar Pollut Bull 72:313–322. https://doi.org/10.1016/j.marpolbul.2012.11.032
Hutchings PA, Bamber L (1985) Variability of bioerosion rates at Lizard Island, Great Barrier Reef: preliminary attemps to explain these rates and their significance. In: Proceedings of the 5th coral reef symposium, Tahiti, pp 333–338
IPCC (2014) Climate change 2014 synthesis report summary chapter for policymakers. IPCC, pp 31
Januchowski-Hartley FA, Graham NAJ, Wilson SK, Jennings S, Perry CT (2017) Drivers and predictions of coral reef carbonate budget trajectories. Proc R Soc B 284:20162533. https://doi.org/10.1098/rspb.2016.2533
Kavousi J, Tavakoli-Kolour P, Mohammadizadeh M, Bahrami A, Barkhordari A (2014) Mass coral bleaching in the northern Persian Gulf, 2012. Sci Mar 78:397–404. https://doi.org/10.3989/scimar.03914.16A
Kavousi J, Tavakoli-Kolour P, Hazraty-Kari S, Goudarzi F (2021) Four consecutive coral bleaching events in the Northern Persian Gulf: 2014–2017. Ann Mar Sci 5(1):007–014. https://doi.org/10.17352/ams.000025
Kennedy EV, Perry CT, Halloran PR, Iglesias-Prieto R, Schönberg CH, Wisshak M, Form AU, Carricart-Ganivet JP, Fine M, Eakin CM, Mumby PJ (2013) Avoiding coral reef functional collapse requires local and global action. Curr Biol 23:912–918. https://doi.org/10.1016/j.cub.2013.04.020
Kriegisch N, Reeves SE, Johnson CR, Ling SD (2020) Sea urchin control of macroalgal communities across a productivity gradient. J Exp Mar Biol Ecol 527:151248. https://doi.org/10.1016/j.jembe.2019.151248
Land LS (1979) The fate of reef-derived sediment on the northern Jamaican island slope. Mar Geol 29:55–71
Lange ID, Perry CT (2019) Bleaching impacts on carbonate production in the Chagos Archipelago: influence of functional coral groups on carbonate budget trajectories. Coral Reefs 38:619–624. https://doi.org/10.1007/s00338-019-01784-x
Langer MR (2008) Assessing the contribution of for aminiferan protists to global ocean carbonate production. J Eukaryot Microbiol 55:163–169. https://doi.org/10.1111/j.1550-7408.2008.00321.x
Long MH, Rheuban JE, Berg P, Zieman JC (2012) A comparison and correction of light intensity loggers to photosynthetically active radiation sensors. Limnol Oceanogr: Methods 10:416–424
Lough JM (2000) 1997–1998: unprecedented thermal stress to coral reefs? Geophys Res Lett 27:3901–3904. https://doi.org/10.1029/2000GL011715
Mallela J (2007) Coral reef encruster communities and carbonate production in cryptic and exposed coral reef habitats along a gradient of terrestrial disturbance. Coral Reefs 26:775–785. https://doi.org/10.1007/s00338-007-0260-8
Mallela J, Perry CT (2007) Calcium carbonate budgets for two coral reefs affected by different terrestrial runoff regimes, Rio Bueno, Jamaica. Coral Reefs 26:129–145. https://doi.org/10.1007/s00338-006-0169-7
Mallela J, Milne BC, Martinez-Escobar D (2017) A comparison of epibenthic reef communities settling on commonly used experimental substrates: PVC versus ceramic tiles. J Exp Mar Biol Ecol 486:290–295. https://doi.org/10.1016/j.jembe.2016.10.028
Morgan G (2006) Country review: Bahrain in review of the state of world marine capture fisheries management: Indian Ocean. In: DeYoung C (ed) FAO fisheries technical paper 488. Rome, pp 187–194
Morris JT, Enochs IC, Besemer N, Viehman TS, Groves SH, Blondeau J, Ames C, Towle EK, Grove LJW, Manzela DP (2022) Low net carbonate accretion characterizes Florida’s coral reef. Sci Rep 12:19582. https://doi.org/10.1038/s41598-022-23394-4
Naser HA (2012) Evaluation of the environmental impact assessment system in Bahrain. J Environ Prot 3:233–239. https://doi.org/10.4236/jep.2012.32029
Paparella F, Xu C, Vaughan GO, Burt JA (2019) Coral bleaching in the Persian/Arabian Gulf is modulated by summer winds. Front Mar Sci 6:1–15. https://doi.org/10.3389/fmars.2019.00205
Perry CT, Hepburn LJ (2008) Syn-depositional alteration of coral reef framework through bioerosion, encrustation and cementation: taphonomic signatures of reef accretion and reef depositional events. Earth Sci Rev 86:106–144. https://doi.org/10.1016/j.earscirev.2007.08.006
Perry CT, Morgan KM (2017) Bleaching drives collapse in reef carbonate budgets and reef growth potential on southern Maldives reefs. Sci Rep 7:40581. https://doi.org/10.1038/srep40581
Perry CT, Edinger EN, Kench P, Murphy GN, Smithers SG, Steneck RS, Mumby PJ (2012) Estimating rates of biologically driven coral reef framework production and erosion: a new census-based carbonate budget methodology and applications to the reefs of Bonaire. Coral Reefs 31:853–868. https://doi.org/10.1007/s00338-012-0901-4
Perry CT, Murphy GN, Kench PS, Edinger EN, Smithers SG, Steneck RS, Mumby PJ (2014) Changing dynamics of Caribbean reef carbonate budgets: emergence of reef bioeroders as critical controls on present and future reef growth potential. Proc R Soc B 281:20142018. https://doi.org/10.1098/rspb.2014.2018
Perry CT, Murphy GN, Graham NAJ, Wilson SK, Januchowski-Hartley FA, East HK (2015) Remote coral reefs can sustain high growth potential and may match future sea-level trends. Sci Rep 5:18289. https://doi.org/10.1038/srep18289
Perry CT, Alvarez-Filip L, Graham NAJ, Mumby PJ, Wilson SK, Kench PS, Manzello DP, Morgan KM, Slangen ABA, Thomson DP, Januchowski-Hartley F, Smithers SG, Steneck RS, Carlton R, Edinger EN, Enochs IC, Estrada-Saldívar N, Haywood MDE, Kolodziej G, Murphy GN, Pérez-Cervantes E, Suchley A, Valentino L, Boenish R, Wilson M, Macdonald C (2018a) Loss of coral reef growth capacity to track future increases in sea level. Nature 558:396–400. https://doi.org/10.1038/s41586-018-0194-z
Perry CT, Lange I, Januchowski-Hartley FA (2018) ReefBudget Indo Pacific: online resource and methodology. https://www.exeter.ac.uk/research/projects/geography/reefbudget/. Accessed 30 July 2023
Pratchett MS, Hoey AS, Wilson SK, Messmer V, Graham NAJ (2011) Changes in biodiversity and functioning of reef fish assemblages following coral bleaching and coral loss. Divers 3:424–452. https://doi.org/10.3390/d3030424
Purkis SJ, Riegl B (2005) Spatial and temporal dynamics of Arabian Gulf coral assemblages quantified from remote-sensing an in situ monitoring data. Mar Ecol Prog Ser 287:99–113. https://doi.org/10.3354/meps287099
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rasser MW, Riegl AB (2002) Holocene coral reef rubble and its binding agents. Coral Reefs 21:57–72. https://doi.org/10.1007/s00338-001-0206-5
Reis MAC, Leão ZMA (2000) Bioerosion rate of the sponge Cliona celata (Grant 1826) from reefs in turbid waters, north Bahia, Brazil. In: Proceedings 9th international coral reef symposium, pp 23–27
Reyes-Nivia C, Diaz-Pulido G, Kline D, Hoegh-Guldberg O, Dove S (2013) Ocean acidification and warming scenarios increase microbioerosion of coral skeletons. Glob Change Biol 19:1919–1929. https://doi.org/10.1111/gcb.12158
Rezai H, Wilson S, Claereboudt M, Riegl B (2004) Coral reef status in the ROPME sea area: Arabian/Persian Gulf, Gulf of Oman and Arabian Sea. In: Wilkinson C (ed) The Status of Coral Reefs of the World, Australian Institute of Marine Science, Townsville, pp 155–169
Riegl B (2002) Effects of the 1996 and 1998 positive sea-surface temperature anomalies on corals, coral diseases and fish in the Arabian Gulf (Dubai, UAE). Mar Biol 140:29–40. https://doi.org/10.1007/s002270100676
Riegl B (2003) Climate change and coral reefs: different effects in two high-latitude areas (Arabian Gulf, South Africa). Coral Reefs 22:433–446. https://doi.org/10.1007/s00338-003-0335-0
Riegl BM, Purkis SJ, Al-Cibahy AS, Al-Harthi S, Grandcourt E, Al-Sulaiti K, Baldwin J, Abdel-Moati AM (2012) Coral bleaching and mortality thresholds in the SE Gulf: highest in the world. In: Riegl BM, Purkis SJ (eds) in Coral reefs of the gulf: adaptation to climatic extremes. Springer, Berlin, pp 95–105
Risaro DB, Chidichimo MP, Piola AR (2022) Interannual variability and trends of sea surface temperature around southern South America. Front Mar Sci 9:829144. https://doi.org/10.3389/fmars.2022.829144
Roff G (2020) Reef accretion and coral growth rates are decoupled in Holocene reef frameworks. Mar Geol 419:106065. https://doi.org/10.1016/j.margeo.2019.106065
Roik A, Röthig T, Pogoreutz C, Voolstra CR (2017) Coral reef carbonate budgets and ecological drivers in the naturally high temperature and high alkalinity environment of the Red Sea. Biogeosciences 1:1–42. https://doi.org/10.5194/bg-15-6277-2018
Roth F, Saalmann F, Thomson T, Coker DJ, Villalobos R, Jones BH, Wild C, Carvalho S (2018) Coral reef degradation affects the potential for reef recovery after disturbance. Mar Environ Res 142:48–58. https://doi.org/10.1016/j.marenvres.2018.09.022
Roxy MK, Gnanaseelan C, Parekh A, Chowdary JS, Singh S, Modi A, Kakatkar R, Mohapatra S, Dhara C, Shenoi SC, Rajeevan M (2020) Indian Ocean warming. In: Krishnan R, Sanjay J, Gnanaseelan C, Mujumdar M, Kulkarni A, Chakraborty S (eds) Assessment of climate change over the Indian Region. Springer, Singapore. https://doi.org/10.1007/978-981-15-4327-2_10
Ryan EJ, Hanmer K, Kench PS (2019) Massive corals maintain a positive carbonate budget of a Maldivian upper reef platform despite major bleaching event. Sci Rep 9:1–11. https://doi.org/10.1038/s41598-019-42985-2
Schiermeier Q (2018) Great barrier reef saw huge losses from 2016 heatwave. Nature 556(7701):281–282. https://doi.org/10.1038/d41586-018-04660-w
Scoffin TP, Stearn CW, Boucher D, Frydl P, Hwakins CM (1980) Calcium carbonate budget of a fringing reef on the west coast of Barbados. Bull Mar Sci 30:475–508
Sheppard C, Loughland R (2002) Coral mortality and recovery in response to increasing temperature in the southern Arabian Gulf. Aquat Ecosyst Health Manag 5:395–402. https://doi.org/10.1080/14634980290002020
Sheppard CR (1988) Similar trends, different causes: responses of corals to stressed environments in Arabian Seas. In: Proceedings of the 6th international coral reef symposium, vol 3. Townsville, Australia, pp 297–302
Shuail D, Wiedenmann J, D’Angelo C, Baird AH, Pratchett MS, Riegl B, Burt JA, Petrov P, Amos C (2016) Local bleaching thresholds established by remote sensing techniques vary among reefs with deviating bleaching patterns during the 2012 event in the Arabian/Persian Gulf. Mar Pollut Bull 105:654–659. https://doi.org/10.1016/j.marpolbul.2016.03.001
Stearn C and Scoffin T. (1977). Carbonate budget of fringing reef, Barbados. In: Proceedings of the 3rd international coral reef symposium, Miami, Florida, pp 471–476
Tanvet C, Benzoni F, Peignon C, Thouzeau G, Rodolfo-Metalpa R (2022) High coral recruitment despite coralline algal loss under extreme environmental conditions. Front Mar Sci 9:837877. https://doi.org/10.3389/fmars.2022.837877
Teichert S, Steinbauer M, Kiessling WA (2020) A possible link between coral reef success, crustose coralline algae and the evolution of herbivory. Sci Rep 10:17748. https://doi.org/10.1038/s41598-020-73900-9
Tribollet A, Golubic S (2005) Cross-shelf differences in the pattern and pace of bioerosion of experimental carbonate substrates exposed for 3 years on the northern Great Barrier Reef, Australia. Coral Reefs 24:422–434
Uwate KR, Shams AJ (1999) Summary of recent coral and coral reef related activities in Bahrain, regional meeting on the assessment of the health of coral reefs. Manama, Bahrain, pp 1–6
Uwate KR, Bader H, Torabi M, Bailey S, Keenan P (2000) Reef check surveys in Bahrain, Arabian Gulf. Manama, Bahrain
Van Lavieren H, Burt J, Feary DA, Cavalcante G, Marquis E, Benedetti L, Trick C, Kjerfve B, Sale PF (2011) Managing the growing impacts of development on fragile coastal and marine ecosystems: lessons from the Gulf. A policy report, UNU-INWEH, Hamilton, ON, Canada
Vousden DH (1988) The Bahrain marine habitat survey: a study of the marine environment of Bahrain using remote sensing as a rapid assessment methodology. In: Proceedings of the ROPME workshop on coastal area development. Regional Seas Reports and Studies No. 90/ROPME. Publication No. GC-5/006, pp 3–34
Vousden DH (1995) Bahrain marine habitats and some environmental effects on seagrass beds: a study of the marine habitats of Bahrain with particular reference to the effects of water temperature, depth and salinity on seagrass biomass and distribution. PhD thesis, University of Wales (Bangor), UK
Wickham H, Averick M, Bryan J, Chang W, McGowan LD, François R, Grolemund G, Hayes A, Henry L, Hester J, Kuhn M, Pedersen TL, Miller E, Bache SM, Müller K, Ooms J, Robinson D, Seidel DP, Spinu V, Takahashi K, Vaughan D, Wilke C, Woo K, Yutani H (2019) Welcome to the tidyverse. J Open Source Softw 4(43):1686. https://doi.org/10.21105/joss.01686
Wilkinson C (1998) The 1997–1998 mass bleaching event around the world. In: Wilkinson C (ed) Status of coral reefs of the world: 1998. Townsville, Australia, pp 15–38
Wilkinson C (2008) Status of coral reefs of the world: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Centre, Townsville, 296
World Bank (2021) Data: land area (sq.km). Bahrain. https://data.worldbank.org/indicator/AG.LND.TOTL.K2?locations=BH. Accessed 20 May 2022
Yachi S, Loreau M (1999) Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. PNAS 96:1463–1468
Yanovski R, Nelson PA, Abelson A (2017) Structural complexity in coral reefs: examination of a novel evaluation tool on different spatial scales. Front Ecol Evol. https://doi.org/10.3389/fevo.2017.00027
Zundelevich A, Lazar B, Ilan M (2007) Chemical versus mechanical Bioerosion of coral reefs by boring sponges—lessons from Phone cf. vastifica. J Exp Biol 210:91–96. https://doi.org/10.1242/jeb.02627
Acknowledgements
The authors thank Hanan Hasan, Hannah Lunt-Bell, Tamera AlHusseini, and Jafar Juma for assistance on the field. Further gratitude goes to the University of Bahrain specifically Dr. Afnan Freije and Dr. Humood Naser for providing lab access. This work would not be possible without the generous funding of Gulf Petrochemical Industries Co. (BSC)-Bahrain and the support of Dr. Abdulrahman Jawahery. Content in this manuscript has been previously submitted as part of a PhD thesis to the University of Essex (AlMealla, 2020).
Funding
Funding for this study was provided by Gulf Petrochemical Industries Co. (BSC) under Grant/Award Number: GPIC/P/D/28/2018. Funding to publish this study in an open-access journal was provided by the University of Essex and Arab Regional Center for World Heritage, Bahrain.
Author information
Authors and Affiliations
Contributions
Study conception and design by LJH and RKA. RKA collected all field data. RKA and BE analyzed the data. RKA and LJH wrote the initial draft of the manuscript. All authors have contributed critically to the drafts and gave the final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Informed consent
This study did not involve any species that were used, captured, and/or experimented on in captivity. All applicable international and national guidelines were followed when handling the species that were encountered along the transect lines of the surveys.
Additional information
Responsible Editor: Saki Harii.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
AlMealla, R.K., Edullantes, B. & Hepburn, L.J. Bleaching threatens positive carbonate budgets on Bahraini reefs. Mar Biol 171, 39 (2024). https://doi.org/10.1007/s00227-023-04351-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-023-04351-9