
Abstract
Thermogravimetric analysis (TGA) was performed on five softwood and five hardwood thin wood samples in the longitudinal (L) and radial (R) directions. Dimensional changes were monitored using a charge-coupled device camera under a nitrogen flow. A comparison of the TG and derivative TG (DTG) curves revealed that shrinkage in the R direction began when the weight was reduced to 79–92% at 305–330 °C and 87–96% at 275–290 °C for softwoods and hardwoods, respectively. Hemicellulose is mainly degraded in this temperature range. In contrast, shrinkage in the L direction started at temperatures close to the DTG peaks, i.e., 360–380 °C and 345–370 °C, respectively, at which temperatures cellulose is mainly degraded. In general, the R/L shrinkage anisotropy was greater for hardwoods than for softwoods, but the species variation was large and the magnitude was directly related to the difference in the shrinkage onset temperatures between the R and L directions, regardless of the wood species. Therefore, shrinkage anisotropy can be attributed to the relative reactivity of hemicellulose and cellulose in wood cell walls. The shrinkage mechanism during carbonization is discussed in terms of the cell wall ultrastructure, in which cellulose microfibrils are covered by a hemicellulose–lignin matrix, and the orientation of the cells in the L and R directions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Global warming caused by the emission of anthropogenic CO2 into the atmosphere is an urgent issue to be solved if the goal of carbon neutrality by 2050 is to be met (Ozawa et al. 2022). Therefore, negative emission technologies that remove CO2 from the atmosphere are attracting attention (Minx et al. 2017), and biochar technology is one such technology (Zimmerman et al. 2011). Atmospheric CO2 can be stored in the soil as biochar, which is produced by carbonizing biomass, and the amount of CO2 assimilated from the atmosphere by terrestrial plants is as high as approximately 15% per year (Amthor 1995).
Wood pyrolysis, which is defined as the thermal degradation of wood in a limited amount of oxygen, is the fundamental basis of thermochemical conversion technologies, including carbonization, gasification, and liquefaction (Zhang et al. 2010). The properties of biochar are related to the pyrolytic degradation of the wood components cellulose, hemicellulose, and lignin within the cell wall nanocomposite structures, which are based on crystalline cellulose microfibrils covered by a hemicellulose–lignin matrix (Wang et al. 2020). Charcoal for fuel has long been produced by carbonization, but the required properties of biochar are different for fuels and soil amendments (i.e., soil additives) (Weber and Quicker 2018). The size and distribution of pores and functional groups are more important for soil amendments because they confer properties such as water retention, an environment for microbial growth, and the ability to chelate fertilizer elements (Mukherjee and Lal 2013). Therefore, research is needed to understand the mechanisms by which these properties develop. This would inform the improvement of carbonization methods for the production of biochar for use as a soil amendment and for non-fuel applications such as adsorbents (Samsudin et al. 2022), electrodes (Yao et al. 2023), and electromagnetic shielding materials (Dai et al. 2023). Wood shrinks during carbonization without melting. This affects the properties of biochar, where the high surface area and porous structure have a wide range of applications in many fields such as carbon capture, carbon sequestration and pollutant removal (Quan et al. 2020). Therefore, the present article focuses on the shrinkage behavior of wood during carbonization.
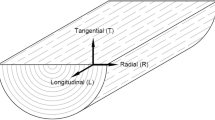
Wood shrinks when it is carbonized, even though its anatomical structure is retained (Baraúna et al. 2021; Byrne and Nagle 1997a; Ehrburger et al. 1982; Vafaeenezhad et al. 2013). The shrinkage is highly anisotropic and its magnitude follows the order: tangential (T) > radial (R) > longitudinal (L) directions (Byrne and Nagle 1997b; Ehrburger et al. 1982; McGinnes et al. 1971; Moore et al. 1974; Paris et al. 2005; Pulido Novicio et al. 2001; Slocum et al. 1978). The information from the literature is summarized in Table 1. The thickness of the cell wall decreases and cell lumen deformation occurs owing to shrinkage anisotropy (Ehrburger et al. 1982). The characteristics of various types of wood cells, including the noticeable shrinkage of vessels (Ehrburger et al. 1982; Kwon et al. 2009; McGinnes et al. Jr 1971) and the lower shrinkage of broad rays (Slocum et al. 1978), have also been investigated by scanning electron microscopy (SEM).
The anisotropy of wood cell walls, which results from the arrangement of cellulose microfibrils, reportedly changes to isotropy at temperatures above 300–400 °C during wood carbonization, as demonstrated by nanoindentation (Brandt et al. 2010), acoustic velocity investigations (Byrne and Nagle 1997b), and SEM observations (Cutter et al. 1980). Brandt et al. (2010) reported that elastic anisotropy disappeared at temperatures above 325 °C. Using scanning thermal microscopy, Xu et al. (2017) reported that thermal conductivity became isotropic at > 325 °C. Byrne and Nagle (1997b) reported that shrinkage continued up to 1000 °C, but the shrinkage anisotropy disappeared and the cell walls became isotropic in terms of acoustic velocity at temperatures above 400 °C. The degradation of cellulose microfibrils in this temperature range has also been reported by researchers using small-angle X-ray scattering (Paris et al. 2005) and transmission electron microscopy (TEM)–Fourier-transform techniques (Jakob et al. 1995; Zollfrank and Fromm 2009). The observations described above suggest that the thermal degradation behavior of wood components (cellulose, hemicellulose, and lignin) in the cell wall ultrastructure is very important for an understanding of the anisotropic shrinkage of wood in the T, R, and L directions. However, this has not been well discussed. Moreover, there have been few reports on the differences between softwoods and hardwoods during carbonization, even though the chemical structures of hemicellulose and lignin, and their coagulation in the cell wall ultrastructure, differ between these two types of wood (Wang et al. 2021).
In the present study, the shrinkage behaviors of wood slices in the R and L directions were investigated using thermogravimetric analysis (TGA) with a charge-coupled device (CCD) camera. The wood slices were taken from five softwoods (Japanese cedar, Japanese cypress, western red cedar, Douglas fir, and larch) and five hardwoods (Japanese beech, Japanese zelkova, walnut, ash, and paulownia). This system can be used to determine shrinkage rates in the R and L directions during TGA, providing insight into the thermal degradation of hemicellulose and cellulose.
Materials and methods
Materials
Veneer sheets (approximately 0.2 mm thick) from five softwoods (Japanese cedar (Cryptomeria japonica), Japanese cypress (Chamaecyparis obtusa), western red cedar (Thuja plicata), Douglas fir (Pseudotsuga menziesii), and Japanese larch (Larix leptolepis)) and five hardwoods (Japanese beech (Fagus crenata), Japanese zelkova (Zelkova serrata), walnut (Juglans), ash (Fraxinus), and paulownia (Paulownia tomentosa)) were purchased from Daiwa Tsukiita Sangyo Corp., Hiroshima, Japan. The sapwood fraction was separated from each veneer sheet and dried in an oven at 105 °C to constant weight. A sample of approximately 4 × 4 mm was then cut in the longitudinal (L) and radial (R) directions for subsequent analytical testing.
Analytical method
Thermogravimetric (TG) analysis was performed on each veneer sample using a thermal analyzer equipped with a CCD camera (NEXTA STA200RV, Hitachi High-Tech Corp., Tokyo, Japan). Each sample was placed in an aluminum pan, the weight loss was recorded from room temperature to 550 °C (10 °C/min) under a dry nitrogen flow (100 mL/min), and the resulting TG and derivative TG (DTG) curves are reported.
Based on the images of the veneer sample taken during the TG analysis, the dimensions in the L and R directions were measured using an ad hoc software program, and reported as a percentage of the dimensions at 150 °C, defined by the following equations:
where L is the relative length in the L direction; and Li and L150 are the lengths at a given temperature and 150 °C in the L direction, respectively; the same definitions apply for the R direction.
Results and discussion
Shrinkage behavior
Figure 1 presents the results of the TGA of western red cedar and images obtained using the CCD camera as an example. The lengths in the R and L directions were recorded with the TG/DTG curves. The onset temperature (Onset T) values at which shrinkage began in the R or L directions, i.e., Onset T (R) or Onset T (L), were determined from the length (%) curves. The corresponding weights, i.e., Onset W (R) or Onset W (L), were determined by comparison with the TG curve. These parameters will be used in the following discussion. The measurements obtained for the other softwood and hardwood species are available in the Electronic Supplementary Information (ESI). The basic parameters obtained in the present paper are summarized in Table 2.

Results of TGA of western red cedar and images obtained using a CCD camera. Onset T (R or L): temperature at which shrinkage began in the R or L direction; Onset W (R or L): corresponding weight for Onset T (R or L). (TGA = thermogravimetric analysis; CCD = charge-coupled device)
Figure 2 (a), (b), and (c) present the R length (%), L length (%), and the R/L ratio defined by R length (%)/L length (%) during TGA, respectively. The R/L ratio indicates the shrinkage anisotropy in the R and L directions. Softwood and hardwood are represented by blue and red lines, respectively, and the ranges of Onset T (R) and Onset T (L) are indicated by the colored boxes (blue: softwood, red: hardwood). The Onset T (L) for Japanese cedar was excluded as an exception because it was in the very low temperature range.

Change in R length (a), L length (b), and R/L ratio (c) according to the TGA results for softwoods and hardwoods. 1: Japanese cedar; 2: Japanese cypress; 3: western red cedar; 4: Douglas fir; 5: larch; 6: Japanese beech; 7: Japanese zelkova; 8: walnut; 9: ash; 10: paulownia. blue: softwoods; red: hardwoods. (TGA = thermogravimetric analysis)
Three temperature zones, Zone I (Onset T 350 °C), Zone II (350–400 °C), and Zone III (400–450 °C), are shown in the figures. They were obtained based on the following characteristic of R-direction shrinkage in Fig. 2 (a): in the Zone I temperature rage, there was steady shrinkage in the R direction, but it was not significant; the shrinkage rates became significant in the Zone II temperature range, then slowed down and continued in the Zone III temperature range.
The ranges of Onset T (R) differ markedly between the hardwoods (275–290 °C) and softwoods (305–330 °C), indicating that hardwoods tend to shrink at lower temperatures than softwoods. Similar trends were observed for Onset T (L) (hardwood: 325–360 °C, softwood: 365–380 °C), except in the case of Japanese cedar, which began to shrink at the lower temperature of 336.7 °C. TGA revealed that up to the Zone I temperature range, shrinkage occurred in the R direction but was very limited in the L direction. Therefore, the R/L ratio curves showing the shrinkage anisotropy are very similar to those of the R length (%).
As the temperature was increased to Zone II, the shrinkage rate in the R direction became significant, and shrinkage began in the L direction. There was significant shrinkage in both the R and L directions in this temperature range, in which thermal degradation of the cellulose microfibrils occurs, as discussed below. There was greater shrinkage in the R direction than in the L direction, so the R/L ratio decreased to less than 1.0, except in the case of the Japanese cedar (close to 1.0, i.e., isotropic).
In the Zone III temperature range, the shrinkage rates became similar for all the species, and in both directions, resulting in constant R/L ratios over the temperature range 400–450 °C, regardless of continuous shrinkage. Therefore, the biochar shrunk isotropically, suggesting that the cell walls became homogeneous at temperatures above 400 °C. In the present study, the R/L ratio at 450 °C was used to compare the intrinsic shrinkage anisotropy of the wood samples. Similar results are reported for wood block carbonization, but at lower temperatures of approximately 300–350 °C (Brandt et al. 2010; Kwon et al. 2009; Xu et al. 2017; Zollfrank and Fromm 2009). The different temperature range is thought to be owing to the different carbonization conditions including heating rate, heating time, and nitrogen flow rate. It should also be noted that the shrinkage anisotropy, which varies with wood species, is determined by the thermal degradation of the wood components up to the Zone II temperature range (below 400 °C).
The R/L ratio tended to be lower for hardwoods than for softwoods, but the variation among species was large (R/L ratio at 450 °C: 0.76–0.90 for hardwoods, 0.78–0.97 for softwoods). Ash (hardwood) had a relatively high R/L ratio of 0.90, and Douglas fir (softwood) had a relatively low R/L ratio of 0.78. Because the variation in L-direction shrinkage rates in the Zone II temperature range was comparatively small, except for Japanese cedar, the difference in the R/L ratio can be attributed to the variation in R-direction shrinkage. Japanese cedar was the exception, with a relatively small shrinkage anisotropy (an R/L ratio at 450 °C of 0.97, i.e., close to 1.0). This was because it shrank more in the L direction than the other species.
Comparison with data from a TGA
The TG and DTG curves measured for five softwoods (blue lines) and five hardwoods (red lines) are illustrated in Fig. 3 (a) and (b), respectively. The colored boxes indicate the ranges of Onset T, and the Zone I–III temperature ranges in Fig. 2 have been inserted to facilitate comparison between the shrinkage behavior and the TG/DTG curves.

TG (a) and DTG (b) curves of softwoods (blue lines) and hardwoods (red lines). 1: Japanese cedar; 2: Japanese cypress; 3: western red cedar; 4: Douglas fir; 5: larch; 6: Japanese beech; 7: Japanese zelkova; 8: walnut; 9: ash; 10: paulownia. (TG = thermogravimetric; DTG = derivative TG)
The main weight loss of the wood specimens occurred in the temperature range 250–400 °C in both softwoods and hardwoods, but the red and blue TG/DTG curves are clearly separated, indicating that the thermal degradation activity of the softwoods and hardwoods differed. Low-temperature weight loss was more significant in the hardwoods than in the softwoods, although the difference in the high-temperature weight loss observed around the DTG peak temperatures was small. These results indicate that the hemicellulose in hardwoods is more thermally reactive than that in softwoods, although the degradation of hemicellulose and cellulose in the wood cell walls cannot be clearly separated by the temperature range according to our ongoing investigations.
The DTG curves of the hardwoods tended to feature a small peak or a clear shoulder in the temperature range 250–330 °C, but the DTG curves of the softwoods, except for larch, did not. The exceptional results for larch can be attributed to a different hemicellulose component from that in the other softwoods. Larch contains arabinogalactan (Timell 1967). This difference between softwoods and hardwoods is believed to result from the difference in the reactivities of xylan and glucomannan: isolated xylan, which is the major hemicellulose of hardwoods and contains uronic acid moieties, has a higher reactivity than isolated glucomannan. However, Wang et al. (2020) used Japanese cedar and Japanese beech to demonstrate that the xylan in wood is not as reactive, and that its reactivity is similar to that of glucomannan. They also reported that such a difference in the DTG curves is owing to the reactivity of cellulose; cellulose and hemicellulose tend to degrade together in Japanese cedar, but independently in Japanese beech. Similar studies are underway for the eight other wood species investigated in the present study, and the results will be published in a separate paper. Nevertheless, it is clear that the thermal degradation of bulk cellulose occurs around the DTG peak temperature.
The Zone II temperature range (350–400 °C) overlaps with the DTG peak temperatures, except for in the case of ash (DTG peak: 345.0 °C). Significant ash shrinkage was observed at temperatures below those of Zone II. Therefore, the cellulose microfibrils of ash cell walls must be exceptionally reactive, which explains the progression of shrinkage in both the R and L directions at such low temperatures. This confirmed that significant shrinkage occurs in both directions when cellulose microfibrils are degraded.
Figure 4 shows the plots of Onset T (L) versus DTG peak temperature. The black dashed line indicates the case in which both temperatures were exactly the same. The DTG peak temperatures overlap for softwoods and hardwoods, except in the case of ash as described above. A linear relationship close to parallel to the black dashed line is observed in hardwoods with an R2 of 0.8757. Therefore, the Onset T (L) of the hardwoods investigated in the present study tended to be 10–20 °C lower than the DTG peak temperature. In contrast, the Onset T (L) of the softwoods was near or above the DTG peak temperature, except in the case of Japanese cedar. This is the reason the Onset T (L) of hardwoods tends to be lower than that of softwoods. The Onset T (L) of Japanese cedar was approximately 20 °C lower than the DTG peak. In contrast, the Onset T (L) of Douglas fir was approximately 20 °C higher than the DTG peak. In hardwoods, the MFA tends to be smaller, which correlates with higher stiffness and density. This could explain why the Onset T (L), or the temperature at which thermal degradation begins, is lower for hardwoods. The tighter alignment of microfibrils in hardwoods may lead to a more rapid initiation of degradation due to the denser structure (Tamer and Fauziah Abdul 2013). Softwoods, on the other hand, generally have a larger MFA, especially in juvenile wood, which results in lower stiffness. This could contribute to the Onset T (L) being near or above the DTG peak temperature, as the less dense structure with a higher MFA may require more energy to initiate degradation (Sultana and Rahman 2014). It’s important to note that while MFA is a contributing factor, other aspects of the cell wall ultrastructure, such as lignin content and distribution, also play a role in the thermal behavior of wood. The differences in Onset T (L) between softwoods and hardwoods, and even among species within these categories, can be complex and influenced by multiple factors.

Relationship between Onset T (L) and DTG peak temperature. Blue: softwoods; red: hardwoods. 1: Japanese cedar; 2: Japanese cypress; 3: western red cedar; 4: Douglas fir; 5: larch; 6: Japanese beech; 7: Japanese zelkova; 8: walnut; 9: ash; 10: paulownia. Onset T (L): temperature at which the onset of shrinkage occurs in the L direction
Figure 5 presents plots of Onset T versus Onset W. The relationships were linear, with high R2 values of 0.9826 (softwood) and 0.9911 (hardwood). This may be attributed to the similar shapes of the TG curves for all the species. The different slopes for the softwoods and hardwoods is owing to the Onset T and Onset W values, which are different for softwoods and hardwoods. The Onset T (hardwood: 273–286 °C, softwood: 305–327 °C) and Onset W (hardwood: 87–96%, softwood: 79–92%) are within relatively narrow ranges in the R direction, but vary widely in the L direction (Onset T: hardwood: 323–359 °C, softwood: 336–380 °C, Onset W: hardwood: 51–70%, softwood: 32–71%). In both directions, more weight loss is required for the onset of shrinkage in softwoods than in hardwoods, especially in the R direction. Together with the higher hemicellulose reactivity described above, this increases the Onset T (R) difference between softwoods and hardwoods, although the variation within softwoods and hardwoods is also large, especially in the L direction.

Relationships between Onset T and Onset W for softwoods (blue) and hardwoods (red) in the R and L directions. Open circles: R direction; solid circles: L direction. 1: Japanese cedar; 2: Japanese cypress; 3: western red cedar; 4: Douglas fir; 5: larch; 6: Japanese beech; 7: Japanese zelkova; 8: walnut; 9: ash; 10: paulownia. Onset T: temperature at which shrinkage began; Onset W: corresponding weight for Onset T
The R/L ratios at 450 °C, which indicate the intrinsic shrinkage anisotropy of wood, are plotted against the difference (ΔT) between Onset T (L) and Onset T (R) in Fig. 6. A clear linear relationship is observed with R2 values of 0.9008. In the case of softwood, the R2 values reach to 0.9438. This is very interesting because the cell composition of these two wood types is very different: softwoods consist of tracheid and parenchyma cells, whereas hardwoods contain vessels with large lumens along with fiber and parenchyma cells. SEM investigations have revealed the preferential shrinkage of vessels (Kwon et al. 2009). However, the present results show that the R/L shrinkage anisotropy can only be determined from the difference between the hemicellulose reactivity (Onset T (R) and the cellulose reactivity (Onset T (L), which can be modified by the nano-assemblies of cellulose microfibrils and the hemicellulose–lignin matrix. The various nano-assemblies of softwood and hardwood have been discussed in the literature (Åkerholm and Salmén 2001; Dammström et al. 2009; Kumagai and Endo 2018; Salmén and Burgert 2009; Simmons et al. 2016; Terashima et al. 2009). We have discussed the influences on the reactivity of cellulose and hemicellulose previously (Wang et al. 2020). Importantly, the results suggest that R/L shrinkage anisotropy is related to cell wall shrinkage rather than wood cell type.

Relationship between ΔT and R/L ratio at 450 °C. 1: Japanese cedar; 2: Japanese cypress; 3: Western red cedar; 4: Douglas fir; 5: larch; 6: Japanese beech; 7: Japanese zelkova; 8: walnut; 9: ash; 10: paulownia. Blue: softwoods; Red: Hardwoods. ΔT: Onset T (L) – Onset T (R). R/L ratio (450 °C): ratio of the lengths in the R and L directions at 450 °C
Mechanism of shrinkage anisotropy
The cell wall ultrastructure and cell orientation must be considered when considering shrinkage anisotropy. Cellulose microfibrils are oriented at an angle of 0–30° (the microfibril angle) in the longitudinal direction of the cell, and amorphous hemicellulose and lignin are present between the cellulose microfibrils. Cell wall shrinkage is caused by the thermal degradation of these nanocomposites.
A proposed degradation and shrinkage behavior of the cell wall ultrastructure is shown in Fig. 7. Hemicellulose and lignin begin to degrade before cellulose in nanocomposites; cellulose is more stable because it is a crystalline substance. This causes shrinkage in the equatorial side of the nanocomposite. However, by investigating the carbonization of wood by in situ X-ray diffraction analysis, Kim et al. (2001) and Zickler et al. (2007) have demonstrated the heterogeneous thermal degradation of cellulose microfibrils in wood. The thermal degradation of the cellulose microfibrils takes place in a direction that is orthogonal to the axis. Therefore, the longitudinal shrinkage of cellulose microfibrils is expected to occur at a very late stage of carbonization. This difference in the timing of the degradation of the hemicellulose–lignin matrix and cellulose can cause shrinkage anisotropy in the cell walls of wood. At temperatures above 400 °C, the cellulose microfibrils disappear completely, and the wood cell walls are converted into an isotropic substance. Therefore, in this temperature range, the shrinkage of the wood cell walls becomes isotropic.

A putative cell wall ultrastructure shrinkage mechanism that explains the shrinkage anisotropy of wood cell walls
As shown in Fig. S2, most of the wood cells are oriented along the L direction, resulting in the suppression of L-direction shrinkage. Therefore, shrinkage preferentially occurs in the R direction in the early stage of carbonization at Zone I temperatures, but not extensively owing to suppression by R-direction-oriented parenchyma cells in the rays. This was corroborated by the observation that the R-direction shrinkage rate increased in the Zone II temperature range, in which cellulose is degraded.
As described above, the intrinsic R/L shrinkage anisotropy of wood is directly determined by the ΔT, i.e., Onset T (L) − Onset T (R), regardless of the wood species. Therefore, the R/L shrinkage anisotropy may be determined by the difference in reactivity between hemicellulose (the matrix) and cellulose. The greater the difference in reactivity, the greater the matrix degradation. Consequently, shrinkage occurs in the R-direction before it occurs in the L-direction, resulting in greater R/L shrinkage anisotropy.
Conclusion
In the present study, we investigated the weight loss behaviors, and shrinkage behaviors in the radial (R) and longitudinal (L) directions of five softwoods and five hardwoods by TGA using a CCD camera. Hardwood TG curves were observed in the lower temperature range owing to the low temperature degradation of hardwood hemicellulose, and indicated that hardwood hemicellulose is more reactive than softwood hemicellulose. The hardwoods tended to exhibit greater R/L shrinkage anisotropy than the softwoods, owing to the greater tendency for shrinkage in the R direction, although species variation was also very large.
The samples shrank in the R direction as the hemicellulose degraded, whereas in the L-direction, shrinkage occurred around the DTG peak temperature, at which cellulose primarily degrades. Therefore, the onset temperature (Onset T) in the R direction was lower for hardwoods than for softwoods. In addition, the corresponding weight (%, based on initial weight) (Onset W), which indicates the weight loss required to initiate shrinkage, was higher for hardwoods than for softwoods. This explains the lower R-direction shrinkage onset temperature in hardwoods compared with that in softwoods.
In contrast, the DTG peak temperature ranges did not differ significantly between the softwoods and the hardwoods, although the species variation was large. In the hardwoods, the onset temperature in the L direction was 10–20 °C lower than the DTG peak temperature, but was close to the DTG peak temperatures for softwoods, except in the case of Japanese cedar, resulting in a lower Onset T and a higher Onset W for hardwoods than for softwoods in the L direction.
Interestingly, there was a linear relationship between R/L shrinkage anisotropy and ΔT, i.e., Onset T (L) − Onset T (R), in both softwoods and hardwoods. This suggests a very interesting conclusion: R/L shrinkage anisotropy is directly determined by the difference in reactivity between hemicellulose and cellulose in wood, regardless of the wood type and cell composition. Therefore, the tendency for greater R/L anisotropy in hardwood compared with softwood can be explained by the greater difference in reactivity between hemicellulose and cellulose in hardwood, and the lower weight reduction required to initiate shrinkage.
Therefore, in the present paper we present a mechanism for R/L shrinkage anisotropy that focuses on the cell wall ultrastructure, which consists of crystalline cellulose microfibrils covered by an amorphous hemicellulose–lignin matrix.
Data availability
No datasets were generated or analysed during the current study.
References
Åkerholm M, Salmén L (2001) Interactions between wood polymers studied by dynamic FT-IR spectroscopy. Polymer 42(3):963–969. https://doi.org/10.1016/S0032-3861(00)00434-1
Amthor JS (1995) Terrestrial higher-plant response to increasing atmospheric [CO2] in relation to the global carbon cycle. Glob Change Biol 1(4):243–274. https://doi.org/10.1111/j.1365-2486.1995.tb00025.x
Baraúna EEP, Stallbaun PH, Monteiro TC, Silva TC, Baldin T, Colen F, Arantes MDC (2021) The effect of carbonization on the wood anatomy of sclerolobium paniculatum vogel. BioResources 16(4):7846. https://doi.org/10.15376/biores.16.4.7846-7854
Beall FC (1974) Carbonized wood: physical properties and uses as an SEM preparation. Wood Sci 6:212–219. https://api.semanticscholar.org/CorpusID:136022915
Brandt B, Zollfrank C, Franke O, Fromm J, Göken M, Durst K (2010) Micromechanics and ultrastructure of pyrolysed softwood cell walls. Acta Biomater 6(11):4345–4351. https://doi.org/10.1016/j.actbio.2010.05.026
Byrne CE, Nagle DC (1997a) Carbonization of wood for advanced materials applications. Carbon 35(2):259–266. https://doi.org/10.1016/S0008-6223(96)00136-4
Byrne CE, Nagle DC (1997b) Carbonized wood monoliths—characterization. Carbon 35(2):267–273. https://doi.org/10.1016/S0008-6223(96)00135-2
Cutter BE, Cumbie B, McGinnes E (1980) SEM and shrinkage analyses of southern pine wood following pyrolysis. Wood Sci Technol 14:115–130. https://doi.org/10.1007/BF00584041
Dai ZH, Wei YY, Hu CS, Wang ZQ, Wei AZ, Zhang WW, Lin XY (2023) Effects of pore structure on wide-frequency electromagnetic interference shielding performance of carbonized wood. Compos Commun 38:101501. https://doi.org/10.1016/j.coco.2023.101501
Dammström S, Salmén L, Gatenholm P (2009) On the interactions between cellulose and xylan, a biomimetic simulation of the hardwood cell wall. BioResources 4(1):3–14
Ehrburger P, Lahaye J, Wozniak E (1982) Effect of carbonization on the porosity of beechwood. Carbon 20(5):433–439. https://doi.org/10.1016/0008-6223(82)90044-6
Jakob HF, Fengel D, Tschegg SE, Fratzl P (1995) The elementary cellulose fibril in picea abies: comparison of transmission electron microscopy, small-angle X-ray scattering, and wide-angle X-ray scattering results. Macromolecules 28(26):8782–8787. https://doi.org/10.1021/ma00130a010
Kim DY, Nishiyama Y, Wada M, Kuga S, Okano T (2001) Thermal decomposition of cellulose crystallites in wood. Holz 55(5):521–524. https://doi.org/10.1515/HF.2001.084
Kumagai A, Endo T (2018) Comparison of the surface constitutions of hemicelluloses on lignocellulosic nanofibers prepared from softwood and hardwood. Cellu 25(7):3885–3897. https://doi.org/10.1007/s10570-018-1861-9
Kwon SM, Kim NH, Cha DS (2009) An investigation on the transition characteristics of the wood cell walls during carbonization. Wood Sci Technol 43(5):487–498. https://doi.org/10.1007/s00226-009-0245-6
McGinnes EA Jr, Kandeel S, Szopa PS (1971) Some structural changes observed in the transformation of wood into charcoal. Wood Fiber Sci :77–83
Minx JC, Lamb WF, Callaghan MW, Bornmann L, Fuss S (2017) Fast growing research on negative emissions. Environ Res Lett 12(3):035007. https://doi.org/10.1088/1748-9326/aa5ee5
Moore G, Blankenhorn P, Beall F, Kline D (1974) Some physical properties of birch carbonized in a nitrogen atmosphere. Wood Fiber Sci :193–199
Mukherjee A, Lal R (2013) Biochar impacts on soil physical properties and greenhouse gas emissions. Agronomy 3(2):313–339. https://doi.org/10.3390/agronomy3020313
Ozawa A, Tsani T, Kudoh Y (2022) Japan’s pathways to achieve carbon neutrality by 2050 – scenario analysis using an energy modeling methodology. Renew Sustain Energy Rev 169:112943. https://doi.org/10.1016/j.rser.2022.112943
Paris O, Zollfrank C, Zickler GA (2005) Decomposition and carbonisation of wood biopolymers—a microstructural study of softwood pyrolysis. Carbon 43(1):53–66. https://doi.org/10.1016/j.carbon.2004.08.034
Pulido Novicio L, Hata T, Kurimoto Y, Doi S, Ishihara S, Imamura Y (2001) Adsorption capacities and related characteristics of wood charcoals carbonized using a one-step or two-step process. J Wood Sci 47:48–57. https://doi.org/10.1007/BF00776645
Quan C, Jia XY, Gao NB (2020) Nitrogen-doping activated biomass carbon from tea seed shell for CO2 capture and supercapacitor. IJER 44(2):1218–1232. https://doi.org/10.1002/er.5017
Salmén L, Burgert I (2009) Cell wall features with regard to mechanical performance. A review COST action E35 2004–2008: Wood machining–micromechanics and fracture. https://doi.org/10.1515/HF.2009.011
Samsudin MH, Hassan MA, Mohd Yusoff MZ, Idris J, Ahmad Farid MA, Lawal ABA, Norrrahim MNF, Shirai Y (2022) Production of nanopore structure bio-adsorbent from wood waste through a self-sustained carbonization process for landfill leachate treatment. Biochem Eng J 189:108740. https://doi.org/10.1016/j.bej.2022.108740
Simmons TJ, Mortimer JC, Bernardinelli OD, Pöppler A-C, Brown SP, Deazevedo ER, Dupree R, Dupree P (2016) Folding of xylan onto cellulose fibrils in plant cell walls revealed by solid-state NMR. Nat Commun 7(1):13902. https://doi.org/10.1038/ncomms13902
Slocum D, McGinnes E Jr, McKown D (1978) Elemental analysis of oak and hickory charcoal using neutron activation analysis. Wood Fiber Sci :200–209
Sultana R, Rahman M (2014) An overview of microfibril angle in fiber of tension wood. Eur J Biophys 2(2):7–12. https://doi.org/10.11648/j.ejb.20140202.11
Tamer AT, Fauziah Abdul A (2013) Cellulose microfibril angle in wood and its dynamic mechanical significance, in: Theo van de, V., Louis, G. (Eds.), Cellulose. IntechOpen, Rijeka, p. Ch. 5
Terashima N, Kitano K, Kojima M, Yoshida M, Yamamoto H, Westermark U (2009) Nanostructural assembly of cellulose, hemicellulose, and lignin in the middle layer of secondary wall of ginkgo tracheid. J Wood Sci 55:409–416. https://doi.org/10.1007/s10086-009-1049-x
Timell T (1967) Recent progress in the chemistry of wood hemicelluloses. Wood Sci Technol 1(1):45–70
Vafaeenezhad H, Zebarjad SM, Khaki JV (2013) Intelligent modeling using fuzzy rule-based technique for evaluating wood carbonization process parameters. Int J Adv Manuf Technol 68:1471–1478. https://doi.org/10.1007/s00170-013-4935-8
Wang JW, Minami E, Kawamoto H (2020) Thermal reactivity of hemicellulose and cellulose in cedar and beech wood cell walls. J Wood Sci 66(1):41. https://doi.org/10.1186/s10086-020-01888-x
Wang JW, Minami E, Asmadi M, Kawamoto H (2021) Thermal degradation of hemicellulose and cellulose in ball-milled cedar and beech wood. J Wood Sci 67:1–14. https://doi.org/10.1186/s10086-021-01962-y
Weber K, Quicker P (2018) Properties of biochar. Fuel 217:240–261. https://doi.org/10.1016/j.fuel.2017.12.054
Xu DL, Ding T, Li YJ, Zhang Y, Zhou DG, Wang SQ (2017) Transition characteristics of a carbonized wood cell wall investigated by scanning thermal microscopy (SThM). Wood Sci Technol 51(4):831–843. https://doi.org/10.1007/s00226-017-0919-4
Yao WR, Zheng DY, Li ZH, Wang YN, Tan HY, Zhang YH (2023) MXene@ carbonized wood monolithic electrode with hierarchical porous framework for high-performance supercapacitors. Appl Surf Sci 638:158130. https://doi.org/10.1016/j.apsusc.2023.158130
Zhang LH, Xu CB, Champagne P (2010) Overview of recent advances in thermo-chemical conversion of biomass. Energy Convers Manage 51(5):969–982. https://doi.org/10.1016/j.enconman.2009.11.038
Zickler GA, Wagermaier W, Funari SS, Burghammer M, Paris O (2007) In situ X-ray diffraction investigation of thermal decomposition of wood cellulose. J Anal Appl Pyrol 80(1):134–140. https://doi.org/10.1016/j.jaap.2007.01.011
Zimmerman AR, Gao B, Ahn M-Y (2011) Positive and negative carbon mineralization priming effects among a variety of biochar-amended soils. Soil Biol Biochem 43(6):1169–1179. https://doi.org/10.1016/j.soilbio.2011.02.005
Zollfrank C, Fromm J (2009) Ultrastructural development of the softwood cell wall during pyrolysis. Holz 63:248–253. https://doi.org/10.1515/HF.2009.031
Acknowledgements
We thank the China Scholarship Council (CSC) for supporting Yu Wang (202204910132) to conduct the present research at Kyoto University. We also thank Frank Kitching, MSc., from Edanz (https://jp.edanz.com/ac) for editing a draft of this manuscript.
Funding
The present work was supported by the Japan Society for the Promotion of Science (grant number JP22H00381) and the JST-Mirai Program, Japan (grant number JPMJMI20E3).
Author information
Authors and Affiliations
Contributions
Yu Wang conducted the experiments, data analysis and wrote the manuscript. Takashi Nomura involved in experiment guidance and data analysis and Ramadan Eljamal was involved in sample characterization and data analysis. All work was carried out under the supervision of Haruo Kawamoto and Eiji Minami. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, Y., Nomura, T., Eljamal, R. et al. Thermogravimetric investigation of anisotropy of dimensional shrinkage of softwood and hardwood during carbonization. Wood Sci Technol 58, 959–974 (2024). https://doi.org/10.1007/s00226-024-01560-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00226-024-01560-3