
Abstract
Recent research activities have provided new insights in vitamin D metabolism in various conditions. Furthermore, substantial progress has been made in the analysis of vitamin D metabolites and related biomarkers, such as vitamin D binding protein. Liquid chromatography tandem mass spectrometric (LC–MS/MS) methods are capable of accurately measuring multiple vitamin D metabolites in parallel. Nevertheless, only 25(OH)D and the biologically active form 1,25(OH)2D are routinely measured in clinical practice. While 25(OH)D remains the analyte of choice for the diagnosis of vitamin D deficiency, 1,25(OH)2D is only recommended in a few conditions with a dysregulated D metabolism. 24,25(OH)2D, free and bioavailable 25(OH)D, and the vitamin D metabolite ratio (VMR) have shown promising results, but technical pitfalls in their quantification, limited clinical data and the lack of reference values, impede their use in clinical practice. LC–MS/MS is the preferred method for the measurement of all vitamin D related analytes as it offers high sensitivity and specificity. In particular, 25(OH)D and 24,25(OH)2D can accurately be measured with this technology. When interpreted together, they seem to provide a functional measure of vitamin D metabolism beyond the analysis of 25(OH)D alone. The determination of VDBP, free and bioavailable 25(OH)D is compromised by unresolved analytical issues, lacking reference intervals and insufficient clinical data. Therefore, future research activities should focus on analytical standardization and exploration of their clinical value. This review provides an overview on established and new vitamin D related biomarkers including their pathophysiological role, preanalytical and analytical aspects, expected values, indications and influencing conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Vitamin D deficiency has developed into a global health issue that affects males and females of all age groups. The lack of sun exposure and a limited availability of vitamin D from natural food sources are the main causes of vitamin D deficiency in modern societies. Other risk factors that can cause or exacerbate vitamin D deficiency are dark skin pigmentation, pregnancy, chronic inflammatory bowel disease with malabsorption, obesity, and advanced age [1]. Acknowledging the global dimension of vitamin D deficiency has sparked an exponential rise in vitamin D testing [2,3,4] and research activities that have resulted in new insights in vitamin D metabolism in various conditions [5,6,7,8,9,10]. Today, it is well established that vitamin D has pleiotropic effects that go far beyond calcium and phosphate metabolism [6]. For example, vitamin D modulates innate and adaptive immunity, cell growth and differentiation, cardiovascular function and hormonal actions [6, 11]. Furthermore, vitamin D deficiency has been linked to a broad range of clinical conditions including cardiovascular disease, malignancies, autoimmune diseases, neuropsychiatric diseases and endocrinopathies [6, 11]. Substantial progress has also been made in the analysis of vitamin D metabolites and related biomarkers, such as vitamin D binding protein (VDBP). Mass spectrometric methods are capable of measuring multiple vitamin D metabolites in parallel [12,13,14]. Recently, Jenkinson et al. have developed a method that can measure 13 different vitamin D metabolites, including 25(OH)D, 24,25(OH)2D and 1,25(OH)2D [13]. While this method is rather complex, the parallel measurement of 25(OH)D and 24,25(OH)2D can easily be performed with liquid-chromatography tandem mass spectrometry using standard instrumentation. Several groups have suggested that the parallel measurement of these two metabolites provides valuable information beyond 25(OH)D alone [15,16,17,18]. This development has recently been recognized by DEQAS, an external quality assessment program for vitamin D. However, clinical guidelines still unanimously recommend assessing vitamin D status by the measurement of 25(OH)D. Furthermore, 1,25(OH)2D is only indicated in a very few clinical conditions, such as severe chronic kidney disease, hereditary phosphate-losing disorders, oncogenic osteomalacia, pseudovitamin D-deficiency rickets, vitamin D-resistant rickets, as well as chronic granuloma forming disorders such as sarcoidosis and some lymphomas [19, 20]. However, recent evidence from experimental and clinical studies merits a closer look into the utility and the analytical standards of traditional and emerging biomarkers of vitamin D metabolism. The aim of this review is to provide structured information on 25(OH)D, 1,25(OH)2D, 24,25(OH)2D, VDBP and the derived parameters vitamin D metabolite ratio (VMR), free (free-25(OH)D) and bioavailable (bio-25(OH)D) 25(OH)D. Particular focus will be put on the clinical utility, preanalytical, analytical and postanalytical aspects, and interfering factors.
Vitamin D Metabolism
Vitamin D is not a single compound, but refers to a group of over 50 metabolites, which are derived from cholesterol via a complex cascade of enzymatic and non-enzymatic reactions [19]. Chemically, they are secosteroids that are characterized by a broken bond in one of the steroid rings. The individual metabolites vary greatly in their plasma concentrations and biological activities [21]. Cholecalciferol (vitaminD3) and ergocalciferol (vitamin D2) are the two main forms of vitamin D. The difference between the two is an additional double bond between carbons 22 and 23 and a methyl group on carbon 24 in the side chain of vitamin D2 [22]. Vitamin D3 is primarily synthesised in human skin under the effect of sunlight. In contrast, vitamin D2 is exclusively obtained from exogenous sources. Many food items, such as fatty fish, liver oil, and egg yolk, contain vitamin D2 and D3 so that the diet contributes 10–20% to the vitamin D supply of humans [23,24,25]. Higher amounts of vitamin D can be obtained from fortified foods, such as milk and margarine, or vitamin supplements [26].
Circulating vitamin D2 and D3 are activated by two hydroxylation reactions that occur in the liver and the kidneys. The hepatic cytochromes P450 CYP2R1 (microsomal) and CYP27A1 (mitochondrial) hydroxylate vitamin D2 and D3 at carbon 25 resulting in the production of 25(OH)D, the most abundant vitamin D metabolite in blood, which is still inactive. Renal CYP27B1 attaches the second hydroxygroup in position 1 forming active 1,25-(OH)2D [27]. In addition to the kidneys, CYP27B1 is also expressed in many other cell types so that 1,25-(OH)2D can be produced by most extra-renal tissues, where it has primarily autocrine or paracrine function. However, this extra-renal 1,25-(OH)2D synthesis contributes little to the circulating concentration of this metabolite [28]. Vitamin D degradation is predominantly driven by CYP24A1, which metabolises 25(OH)D to 24,25-dihydroxy-vitamin D [24,25(OH2D)] and 1,25(OH)2D to 1,24,25-trihydroxy-vitamin D [1,24,25(OH)3D][27]. In the circulation, all vitamin D metabolites are bound to vitamin D binding protein (DBP), albumin and lipoproteins.
The C3 hydroxyl group of 25(OH)D2 and 25(OH)D3 can be converted to beta orientation resulting in the formation of 3-epi-25(OH)D3, 3-epi-25(OH)D2. It is believed that this irreversible modification is catalysed by the enzyme 3-epimerase (C-3 epimerisation pathway), which also epimerizes 1α,25(OH)2D3 and 1 α,25(OH)2D2. Similar to the non-epimerized metabolites, also 3-epi-25(OH)D3, and 3-epi-25(OH)D2 can be further hydroxylized by 1α-hydroxylase forming 3-epi-1 α,25(OH)2D3 and 3-epi-1 α,25(OH)2D2. So far, 3-epimerase has been identified in the endoplasmatic reticulum of liver, bone and skin [29, 30]. In serum from adult humans, the concentration of 3-epi-25(OH)D3 varies between 1 and 25% [31]. Children appear to have markedly higher proportion of 3-epi-25(OH)D3 reaching up to 60% [32]. With the methods that are commonly used in medical laboratories, the concentrations of all other 3-epimers are too low to be detected. In a study from Shah et al. the mean concentrations of 3-epi-25(OH)D3, and 3-epi-25(OH)D2 were 6.1 nmol/L and 1.1 nmol/L, respectively. The role of vitamin D 3-epimers in health and disease is still a matter of debate as most of our knowledge is derived from in vitro studies. Apparently, 3-epi-25(OH)D has low affinity for VDBP (36–46% compared with 25(OH)D) and VDR (2–3%), which explains at least partly the low serum concentration. At present it is believed that 3-epi-25(OH)D does not adequately reflect vitamin D status and thus its separate measurement is not recommended. Existing knowledge indicates that 1 α,25(OH)2D3 harbours a lower biological activity and has a markedly lower affinity to the VDR than 1α,25(OH)2D3. Also, its ability to stimulate intestinal calcium absorption is significantly reduced [33,34,35]. PTH suppression, however, was detected at similar rates as 1,25(OH)2D3 (reviewed in [32]).
25(OH)D
Role of the Marker
The inactive prohormone 25(OH)D represents the main reservoir and transport form of vitamin D. The generation of 25(OH)D from pro-vitamin D3 is catalyzed by CYP27A, a 25-hydroxylase encoded by the CYP2R1 gene [36]. As the most abundant vitamin D metabolite in blood, 25(OH)D includes the two forms 25(OH) D2 and 25(OH)D3 [37]. Hydroxylation of 25(OH)D in position 1 is mediated by CYP27B1 and results in the formation of active 1,25(OH)2D. However, this alpha-hydroxylation occurs on demand, primarily in the kidneys, under the control of parathyroid hormone (PTH). Consequently, the serum concentration of 1,25(OH)2D is kept within the reference range over a wide concentration range of 25(OH)D and does not reflect vitamin D stores. In fact, individuals with a low serum 25(OH)D concentration often have a high-normal or raised 1,25(OH)2D concentration due to a compensatory induction of CYP27B1. Therefore, current guidelines unanimously recommend 25(OH)D as the preferred indicator of the body’s vitamin D stores.
People with dark skin have 30–40% lower 25(OH)D serum concentrations than Caucasians but comparable or higher bone mineral density (BMD) and lower fracture risk [38]. Furthermore, lower serum 25(OH)D concentrations in blacks are not associated with higher PTH concentrations, lower BMD or increased fracture risk [39,40,41,42]. This conundrum is at least partly explained by a high prevalence (> 90%) of the GC1F haplotype of VDBP (rs7041-T/rs4588-C) in blacks, whereas the GC1S haplotype of VDBP (rs7041-G/rs4588-C) is dominant in Caucasians [43]. While carriers of the GC1F and the GC1S haplotypes show comparable VDBP concentrations [43] they differ in their affinity for most vitamin D species, which contributes to the different 25(OH)D concentrations. However, the availability of free vitamin D for metabolism is not affected. Berg et al. have shown that blacks and whites with comparable serum PTH concentrations also have a comparable VMR despite substantially lower 25(OH)D levels [44]. Other factors that contribute to lower 25(OH)D concentrations in blacks include adiposity and skin pigmentation. A recent report from an international expert panel stated that no one factor alone could fully explain the vitamin D paradox in Black Americans. In addition, blacks have no skeletal benefits from high doses of vitamin D supplementation [45].
Pre-analytical Considerations
The concentration of 25(OH)D can be assessed in both, serum and plasma, with similar results [46]. A low biological variability [47, 48] in combination with rather high concentrations in the nM range and a long in vitro half-life make 25(OH)D a robust biomarker that can reliably be measured by clinical laboratories [14, 19, 20]. A recent study from Denmark has shown that the in vivo half life depends on the individual 25(OH)D concentration and the vitamin D receptor (VDR) gene polymorphism rs2228570 [49]. In individuals with a 25(OH)D start concentration between 68 and 213 nmol/L, mean half-life was 89 days, whereas lower concentrations were associated with a half-life between 149 and 199 days. The longer half-live at lower 25(OH)D concentrations may be caused by storage mobilisation, changed catabolism or increased intestinal absorption. In addition, the VDR gene polymorphism rs2228570 may explain up to 88% of the observed variation with the genotype GG having a 120 days shorter half-life than the genotype AA/AG. Stability studies have shown that 25(OH)D is stable in whole blood for up to 24 h when kept at room temperature (RT) [50]. Also, storage of serum for up to 72 h at RT or refrigerated at 6 °C has negligible effects on 25(OH)D. Good short term stability of serum 25(OH)D at RT has also been reported by Zelzer S et al. [17]. Long-term stability at − 20 °C and − 80 °C is generally good. In an own study, we were able to demonstrate that changes do not exceed 15% after 2 months [17]. Recently, Cavalier et al. reported only minor changes of 25(OH)D after 5 years of storage at − 80 °C, which are most likely the result of sample concentration rather than degradation [51]. Long-term storage appears to have the least effect on 25(OH)D measured by LC–MS/MS, whereas samples from renal and pregnant patients may show substantial differences when analysed by immunoassay. Importantly, freezing and thawing of plasma and serum samples does not affect the 25(OH)D concentration [52]. Also, centrifugation temperature has no substantial effect on the 25(OH)D concentration [53, 54]. Importantly, serum and plasma samples for the measurement of 25(OH)D should always be stored in the dark. While short term UV-irradiation has little impact on the 25(OH)D concentration, prolonged sun exposure reduces 25(OH)D by more than 50% [54].
Analytical Considerations
25(OH)D is a challenging analyte due to its strong binding to VDBP and other carriers, the need of measuring 25(OH)D2 and 25(OH)D3 in an equimolar fashion, the coexistence of multiple chemically related molecules that may cross-react, and common matrix effects, such as heterophilic antibodies or changes in protein composition [19, 20]. Medical laboratories employ different methods for the measurement of 25(OH)D in serum and plasma with automated immunoassays, ELISAs and LC–MS/MS being the most frequently used. Radioimmunoassays (RIA) and high-performance liquid chromatography (HPLC) have been widely used in the past, but due to a broad range of alternatives they play a marginal role in today’s practice. From an analytical point of view, the available methods can be divided in those with and without complete removal of proteins and lipids by strong organic solvents prior to analysis. The first group includes LC–MS/MS, HPLC and, RIAs whereas the second group comprises automated immunoassays. As automated immunoassay-analysers cannot use such organic solvents, they employ alternative strategies to release 25(OH)D from its carriers. However, they have an inferior dissociation efficacy. The strategies that are used by immunoassay manufacturers to separate 25(OH)D from its carriers are optimized for the expected matrix composition. In situations where the matrix is altered, such as in pregnant women, patients with chronic kidney disease or individuals with a polymorphic variant of VDBP, these approaches may be less efficient and thus introduce analytical bias. In contrast, the organic solvents used in LC–MS/MS are strong enough to precipitate all proteins and detach all vitamin D metabolites from their carriers [55, 56]. Consequently, automated immunoassays are characterized by a variable analytical performance. In external quality assurance programs, the means of commonly used automated immunoassays show differences of up to 20 nmol/L in the clinically relevant range. Standardization of the 25(OH)D measurement has been a major step forward. With the help of the standard reference materials (SRM) 972 and 972a, developed by the National Institute of Standardization (NIST), and validated reference measurement procedures (RMP) from the University of Gent [57] and the Centre of Disease Control [58] it is possible to accurately determine the 25(OH)D concentration in serum and plasma samples. As these RMPs are not suitable for clinical laboratories, the Vitamin D Standardization and Certification Program (VDSCP) has been developed with the aim to align the results of different methods to this reference system [59]. Today, 38 methods are listed at the CDC homepage as being VDSCP certified (https://www.cdc.gov/labstandards/vdscp_participants.html). However, the main weakness of VDSCP is that certification depends on the mean bias obtained on a standard set of samples, regardless of the scatter that these samples produce around the target values. In other words, a wide scatter can produce a similar mean bias than a narrow scatter, as long as the individual values are distributed equally around the target values. Therefore, Wise et al. have proposed the percentage of samples with a bias ≤ 10% as a better criterion for accuracy [60]. While the different standardization efforts have clearly improved the comparability of 25(OH)D results obtained with automated immunoassays, external quality assurance programs still show unsatisfactory variability. Numerous laboratories have acknowledged the intrinsic variability of automated immunoassays and switched to LC–MS/MS with electrospray ionisation [61] as the gold standard for the quantification of 25(OH)D. This technology is suitable for the analysis of serum, heparin and EDTA samples [62]. It can accurately measure 25(OH)D2 and 25(OH)D3. Currently, 20% of all participants of the DEQAS program use LC–MS/MS for the measurement of 25(OH)D. This group is consistently well aligned to the target values and copes best with interferences from other vitamin D metabolites and matrix effects.
All LC–MS/MS methods require a more or less complex pre-analytical sample preparation that cleans the sample and extracts vitamin D metabolites through protein precipitation, liquid–liquid extraction and sometimes derivatisation [40]. Regardless of the pre-analytical sample preparation, thoroughly validated LC–MS/MS methods are consistently capable of producing accurate results [17, 62]. For example, Zhang et al. validated a highly specific LC–MS/MS method for the simultaneous quantification of 25(OH)D2 and 25(OH)D3 in serum and plasma [62]. Using a combination of methanol precipitation and liquid–liquid extraction by heptane, they were able to obtain a 72% recovery with a coefficient of variation (CV) < 7.05%. The results were well aligned to the Vitamin D External Quality Assessment Scheme (DEQAS) LC–MS/MS method mean values, but about 9% higher than the commonly used DiaSorin Liasson assay. However, it should be mentioned that LC–MS/MS methods for the measurement of 25(OH)D also have disadvantages, such as a (semi)manual sample preparation, and the need of expensive instrumentation and experienced staff. In addition, they are not yet suitable for high throughput. Nevertheless, first fully automated solutions are already available.
From the analytical point of view, 3-epi-25(OH)D3 co-elutes and has identical mass as 25(OH)D3, and they can only be separated by high resolution chromatography [63]. Considering the ongoing controversy about the biological relevance of vitamin D 3-epimers in health and disease the separate measurement of 3-epi-25(OH)D3 is not recommended. In addition, most 25(OH)D automated immunoassays do not cross-react with 3-epi-25(OH)D so that they do not represent a relevant issue in clinical practice. Only in children, the higher 3-epi-25(OH)D concentrations may eventually result in a misclassification of vitamin D status, when measured with one of the few 25(OH)D methods that are interfered by C3-epimers, such as protein-binding assay.
Reference Values
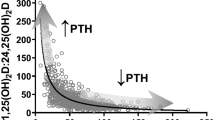
Typically, clinical laboratories provide their results with a reference range that comprises the values between the 2.5th and the 97.5th percentile of a healthy reference cohort. For several reasons, 25(OH)D results should not be interpreted on the basis of such a reference range. The circulating 25(OH)D concentration is subject to pronounced seasonal variation of 20–30% with the highest values in summer and autumn [64]. In 74,235 serum samples from Northern Italy that were collected over a two-year period and analysed by a validated LC–MS/MS method, the 2.5th and 97.5th percentile spanned a rather wide range from 12 to 159 nmol/L. Amongst 2000 healthy Austrian blood donors the traditional reference interval was 28.5–136 nmol/L (unpublished data). However, within the traditional reference interval of 25(OH)D, higher concentrations of this metabolite were associated with improved blood biomarker concentrations of calcium homeostasis and bone turnover. In addition, within the reference range, higher 25(OH)D concentrations are also associated with a lower risk of bone fractures [65,66,67]. Valcour et al. have shown that PTH continuously decreases with increasing concentrations of 25(OH)D and that there is no threshold above which this relationship plateaus [68]. The same applies to other conditions, such as stroke, cardiovascular disease, diabetes mellitus and autoimmune disease [69,70,71,72,73]. Considering that the risk for numerous clinical conditions substantially varies across the reference interval, clinical cut-offs are recommended for the interpretation of serum 25(OH)D results. Several scientific societies have developed practice guidelines that propose 25(OH)D cut-offs on the basis of clinical risk [74]. Most guidelines distinguish between sufficiency, insufficiency and deficiency [37, 75, 76]. Some guidelines also provide cut-offs from severe deficiency to toxicity as summarised in Table 1 [74, 77,78,79,80,81,82]. These cut-offs apply to bone health, osteoporosis, fracture risk and general health. The First International conference on Controversies in Vitamin D established that levels of 25(OH)D below 12 ng/mL (30 nmol/L) were associated with an increased risk of rickets and osteomalacia, while concentrations between 50 and 125 nmol/L are sufficient to maintain bone health [83]. However, commonly used 25(OH)D cut-off values apply primarily to Caucasians and Asians. People with dark skin have 30–40% lower 25(OH)D serum concentrations than Caucasians but comparable or higher bone mineral density (BMD) and lower fracture risk [38].
Indications
Measurement of serum 25(OH)D is recommended for the diagnosis of vitamin D deficiency [37, 78, 81]. Individuals at increased risk of vitamin D deficiency are those with fragility fractures, chronic kidney disease, malabsorption, and abnormalities of calcium and phosphate metabolism [84]. Determining serum 25(OH)D is also useful for the differential diagnosis of rickets, osteomalacia, and the monitoring of vitamin D supplementation. Finally, 25(OH)D should be determined in patients with suspected hypervitaminosis D and intoxication. When considering measuring vitamin D in the context of various non-bone related disease, it should be kept in mind that reverse causality and residual confounding cannot be excluded. Rather than being an actionable causal factor for chronic diseases, vitamin D deficiency could simply be the result of an underlying condition that alters vitamin D metabolism, or a lifestyle that is associated with poor health and micronutrient deficiencies. Therefore, the value of measuring 25(OH)D for the assessment and management of non-bone-related diseases is a matter of ongoing debate and not yet recommended.
Influence of 25(OH)D in Pathological Conditions
Most 25(OH)D in serum is bound to VDBP (85%) and albumin (15%) [85]. Variations in the serum concentrations of these carriers lead to corresponding changes of 25(OH)D. The hepatic 25-hydroxylation of vitamin D was long believed to be unregulated [36]. However, recent studies suggest that both age and metabolism can modulate CYP27A activity. Aged humans and animals are characterized by a lower expression of CYP2R1 than their young counterparts and thus they are less responsive to vitamin D supplementation [86]. As vitamin D can be sequestered in adipose tissue [87], it is not surprising that nutrition and body composition also modulate CYP2R activity [87, 88].
Pregnancy causes a small increase in 25(OH)D which is at least partly driven by an increase in VDBP [89]. However, not all studies found a significant changes of serum 25(OH)D in pregnant women [90, 91]. Figueiredo et al. showed that the serum 25(OH)D concentration during pregnancy is influenced by seasonal effects [92]. Women starting pregnancy during winter showed rising 25(OH)D concentrations, whereas women who fell pregnant in summer exhibited no increase. Interestingly, the placenta possesses the principal enzymes of vitamin D metabolism including CYP2R1, CYP27B1 and CYP24A1 and may thus be involved in the regulation of circulating 25(OH)D levels [93]. Maternal 25(OH)D can crosses the placenta and is the exclusive vitamin D source of the foetus reaching cord blood levels between 75 and100% of the maternal value [94].
Anti-epileptic drugs, such as phenobarbital, carbamazepine, phenytoin and valproate, are known to accelerate vitamin D catabolism and thus lower the serum 25(OH)D concentration [82]. Binding of these drugs to the pregnane X receptor increases 25(OH)D catabolism by an induction of CYP3A4 and CYP24A1. The preganane X receptor is a nuclear receptor that shares homologies with the VDR. In order to mitigate the risk of vitamin D deficiency, reduced BMD and increased fracture risk, patients on long-term treatment with anti-epileptic drugs should receive a prophylactic vitamin D supplementation and have their serum 25(OH)D concentration monitored.
Patients with CKD at stages 3 and 4 usually present deficiency of vitamin D, a marker of poor prognosis [95]. Similarly, 60–80% of pre-dialysis children with CKD also have low levels of 25(OH)D [96]. In addition to low serum 25(OH)D levels, CKD patients are also characterized by elevated phosphate and FGF-23 concentrations. The adverse impact of CKD on calcium and phosphate metabolism is associated with secondary hyperparathyroidism. However, the management of vitamin D status in nephrology is still a matter of ongoing debate. In any case, CKD patients with 25(OH)D deficiency should always be treated, regardless of PTH [95]. Vitamin D deficiency in Stages 1 to 3a follow general population recommendations. Later stages are treated with calcitriol or vitamin D analogues, depending on patient characteristics [97].
Obese and bariatric surgery patients frequently present with vitamin D deficiency. Fat soluble vitamin D can be sequestered in adipose tissue resulting in reduced circulating 25(OH)D concentrations. Instead, bariatric surgery compromises the absorption of dietary vitamin D [98, 99]. To prevent adverse skeletal effects, vitamin D deficiency should be diagnosed and corrected prior to bariatric surgery [100]. However, it may be difficult to normalize 25(OH)D and PTH in these patients.
Vitamin D levels have also been investigated in cancer and cardiovascular disease (CVD). Observational studies identified an increased CVD mortality only in vitamin D deficient subjects, but not individuals with normal 25(OH)D serum concentrations. Large randomized intervention studies, such as the VITAL study (25,000 individuals aged 50 years or older treated with 2000 IU/day of vitamin D and 1 g/day of omega-3 fatty acid for over 5 years) [101] and the VIDA study (5000 individuals over 50 years of age treated with an initial dose of 200,000 IU and then 100,000 IU/month of vitamin D for 3.3 years) [102] failed to show any beneficial effect of vitamin D supplementation on the incidence of invasive cancer or the appearance of CVD. The lacking effect of vitamin D supplementation on CVD and cancer incidence is further supported by meta-analyses that pooled randomised clinical trials (reviewed in [103]). However, in line with the observational studies mentioned before, a reduced risk was found in vitamin D deficient subjects. Although existing intervention studies failed to show reductions in CVD and cancer risk, the results may be biased due to some pitfalls, such as the lack of appropriate controls that are completely naive to oral vitamin D intake via the food or de-novo synthesis by the skin [103]. Based on existing observational and intervention studies, measurement of serum 25(OH) is primarily useful to identify vitamin D deficient individuals, where supplementation appears to have beneficial effects on CVD and cancer risk. It should be kept in mind that the interpretation of 25(OH)D results in CVD and cancer patients could be complicated by residual confounding and reverse causality, as vitamin D insufficiency could be the result of poor health and disease specific alterations of vitamin D metabolism [104]. Therefore, the quantification of additional vitamin D metabolites could aid the assessment of vitamin D status in these patients.
Due to its immune-modulatory function, vitamin D has also been investigated in COVID-19 patients. Observational studies have shown that individuals with vitamin D deficiency have an increased risk to develop severe COVID-19 (as reviewed in [105]). However, a retrospective study by Mangge et al. failed to find any association between 25(OH)D3, 25(OH)D2, 24,25(OH)2D3 and 25,26(OH)2D3) and clinical outcome measures, such as recovery rate, death or the need of respiratory support [106]. In the absence of COVID-19-specific prevention studies, some information can be derived from vitamin D supplementation studies that measured the incidence of other infectious respiratory diseases, especially in children. Again, while these studies did not show significant effects in individuals with adequate 25(OH)D concentrations, those with vitamin D deficiency benefitted from vitamin D supplementation. As these studies used primarily historical 25(OH)D analyses, their results may be biased. Based on the limited evidence available, the role of vitamin D in SARS-CoV2-infected patients remains controversial. However, there is some support for a correction of vitamin D deficiency in SARS-CoV2-infected patients to prevent severe disease and adverse outcome.
Measurement of 1,25(OH)2D
Role of the Marker
1,25(OH)2D is the active form of vitamin D that is produced through 1α-hydroxylation of 25(OH)D. This hydroxylation step is catalysed by CYP27B1 (1α-hydroxylase) and occurs mainly in the kidneys. Consequently, the circulating 1,25(OH)2D concentration primarily reflects renal production. Meanwhile, it is well established that many other cells and tissues, such as monocytes, macrophages, dendritic cells, osteoblasts, and keratinocytes, also express 1α-hydroxylase and thus are cable of producing 1,25(OH)2D locally [107]. However, 1,25(OH)2D that is produced by extra-renal tissues has mainly auto- and paracrine functions and adds little to the serum level [108, 109]. The biological activity of 1,25(OH)2D is mediated by binding to the VDR [5], a nuclear receptor that binds specific DNA sequences located at different genomic regions (vitamin D response elements), to subsequently modulate the expression of the respective target genes [110].
With concentrations in the lower pmol/L range, 1,25(OH)2D circulates at much lower levels than 25(OH)D. The half-life of 1,25(OH)2D is only 4–6 h, which makes serum levels rather volatile. Both metabolites have similar physical and chemical properties [111,112,113], but unlike 25(OH)D, 1,25(OH)2D does not reflect the storage levels of vitamin D in the body [114]. Supplementation of vitamin D increases serum 25(OH)D, but has no measurable effect on serum 1,25(OH)2D [115, 116]. Some studies suggest that there is no relationship between both metabolites [117, 118]. In fact, it appears that 1,25(OH)2D is produced on demand depending on the cellular need, whereas 25(OH)D reflects the stores that are available for utilization.
The main role of 1,25(OH)2D is regulating blood calcium levels through intestinal absorption, renal reabsorption and release from bone stores [119]. When the calcium intake is low, PTH increases the synthesis of 1,25(OH)2D, which subsequently activates the transport of calcium from the intestinal lumen to the blood by upregulating TRPV6 and other genes, such as S100G, ATP2B1, and CLDN2, in the intestine [120]. When calcium intake increases, PTH secretion is reduced leading to a decrease in 1,25(OH)2D and activation of a paracellular route, a non-saturable and passive transport through the intercellular space between adjacent cells, which is also modulated by 1,25(OH)2D (reviewed in [121]). Therefore, 1,25(OH)2D is a key regulator of bone mineralisation [122]. The release of calcium from bone is accompanied by an increased secretion of fibroblast growth factor 23 (FGF23) by osteocytes, which, upon binding to Klotho, inhibits 1-alpha hydroxylase and ultimately reduces the renal synthesis of 1,25(OH)2D [123].
In addition to the regulation of calcium homeostasis, 1,25(OH)2D also supports bone formation by increasing the osteoblastic expression of alkaline phosphatase, osteocalcin and ostepontin [124]. Furthermore, it stimulates bone resorption by increasing osteoclastogenesis and osteoclast activity [125]. 1,25(OH)2D is also involved in the regulation of cell proliferation, differentiation and apoptosis [126]. Experimental evidence from 1α-hydrolase knockout mice suggests that low levels of 1,25(OH)2D increase age-related bone loss by activating senescence pathways, like p16/p19 [127]. An adequate supply with 1,25(OH)2D could thus preserve bone health by activating an anti-ageing mechanism [128].
In addition to its role in bone, 1,25(OH)2D also attenuates the inflammatory response of monocytes and macrophages and increases the expression of anti-inflammatory compounds like IL-10. In contrast, the secretion of pro-inflammatory cytokines like IL-1b, IL-6, TNFα, RANKL and COX2 is reduced by 1,25(OH)2D [129]. These immunomodulatory effects may explain the 1,25(OH)2D-mediated risk reduction of autoimmune diseases like multiple sclerosis [130]. Blood pressure, insulin secretion and insulin sensitivity are also modified by calcitriol [131,132,133]. These associations could, at least partially, be due to the regulatory role of 1,25(OH)2D in calcium homeostasis. Also, the renin-angiotensin axis is influenced by 1,25(OH)2D, which may explain the association with hypertension (reviewed in [134]).
Recent evidence suggests that 1,25(OH)2D is also linked to the gut microbiome. High serum concentrations of 1,25(OH)2D, but not 25(OH)D, are strongly correlated with alpha and beta diversity, especially with butyrate-producing bacteria, like Firmicutes. Based on these results, it has been hypothesized that butyrate-producing bacteria could stimulate the local production of 1,25(OH)2D by dendritic cells in the colon [135]. In this context, a previous animal model lacking VDR and CYP27B1 also showed impaired levels of 1,25(OH)2D and reduced number of Firmicutes [136].
Pre-analytical Considerations
1,25(OH)2D has similar physical and chemical properties as 25(OH)D; however, it is more hydrophilic, has shorter half-life (around 6 h), and it is found in blood at very low concentrations (in the range of picomol/L) [137]. Altogether, it presents similar challenges as 25(OH)D for analytical detection, but requires a substantially higher sensitivity [46]. Because of these characteristics, results are more volatile.
Analytical Considerations
Measurement of 1,25(OH)2D in serum or plasma is considerably more challenging than 25(OH)D due to is very low concentration. In addition, a standard reference material and a reference measurement procedure are lacking. The first assay for the measurement of 1,25(OH)2D was a radioreceptor binding assay, where 1,25(OH)2D in the sample displaced tritiated hormone from a cytosol-chromatin receptor preparation isolated from chick small intestine [138]. With this assay, Brumbaugh et al. were able to determine a human plasma concentration of 60 pg/ml in patients with renal disease. In the following years competitive protein binding assays [139], RIAs [140], enzyme immune assays (EIAs) [141], HPLC [142,143,144], gas chromatography-mass spectrometry (GC–MS) [145] and LC–MS/MS [146] methods were developed. A comprehensive overview of available methods and their characteristics is provided by several review articles [147, 148]. Until recently, commercial RIAs were widely used to determine 1,25(OH)2D in clinical laboratories. With the development of fully automated immunoassays [141, 149] this picture has changed. In 2017, 75% of the participants in the DEQAS program used automated immunoassays, 15% manual immunoassays and 9% LC–MS/MS. However, the comparability of automated immunoassays with LC–MS/MS methods is not ideal [150]. Even within the individual method groups substantial variability exists. In this context, different calibration procedures are a common source of variability [151]. LC–MS/MS methods adopt different strategies to cope with the very low analyte concentration. Vitamin D metabolite profiling, and in particular measurement of 1,25(OH)2D, typically requires a dual column system, where the first column serves for analyte enrichment and the second one for separation. Other methods include a preanalytical immunopurification step [149, 152, 153] whereas others do not [154]. The antibody used for immunopurification is supposed to enrich the target analyte 1α-25-dihydroxyvitamin D and thus reduces interferences from isobaric compounds, such as 1β-25-dihydroxyvitamin D [84]. Another way to improve analytical sensitivity is the derivatization of 1,25(OH)2D [46]. PTAD (4-phenyl-1,2,4-triazoline-3,5-dione) is the most frequently used derivatization agent for this purpose. Also the ionization mode impacts the analytical performance of LC–MS/MS methods [63]. While derivatization methods typically use electrospray ionization (ESI), atmospheric-pressure chemical ionisation (APCI) is a valid alternative that achieves good results without the need of derivatization [63]. However, in the absence of standardization it is not possible to conclude that one method is more accurate than another.
Reference Values
In contrast to 25(OH)D, where results are interpreted on the basis of clinical cut-offs, method specific reference intervals are recommended for 1,25(OH)2D. In healthy adults, the reference interval for the IDS RIA is 43–168 pmol/L, whereas the automated IDS iSYS immunoassay has a reference interval of 63–228 pmol/L [141]. A different reference interval of 59 -159 pmol/L has been reported for LC–MS/MS. However, these methods typically measure only 1,25(OH)2D3. For a separate quantitation of 1,25(OH)2D2 most methods are not sensitive enough as the circulating level has been reported to be < 17 pmol/L [155]. In addition, immunoassays may be interfered by chemically related vitamin D metabolites that cross-react.
Blood levels of 1,25(OH)2D in children are higher than in adults. The following paediatric reference intervals have been established by Higgins et al. on the CALIPER cohort using the DiaSorin Liason XL method: 77—471 pmol/L between 0 and 1 year, 113–363 pmol/L between 1 and 3 years, and 108 – 246 pmol/L for children older than 3 years. The CALIPER cohort includes 377 Canadian children and adolescents [156]. For other methods, paediatric reference intervals have not been reported.
Indications
To date, the clinical utility of 1,25(OH)2D is insufficiently understood, which limits its clinical use. Serum levels of 1,25(OH)2D have little or no relationship to vitamin D stores but rather are regulated by PTH. Measurement of 1,25(OH)2D is helpful in the investigation of patients with unexplained hypercalcaemia, sarcoidosis, granulomatous disorders, pseudo-vitamin D deficiency, rickets, tumour-induced osteomalacia, hyperparatiroidism, and CYP24A1 deficiency [20]. Moreover, measurement of this vitamin D metabolite can be helpful to differentiate between FGF23-dependent and –independent phosphopenic rickets [157]. Levels of 1,25(OH)2D are usually low in CKD, but its measurement has only been recommended when patients present severe and progressive hyperparathyroidism [158].
Influence of 1,25(OH)2D in Pathological Conditions
Low serum levels of 1,25(OH)2D are observed in CKD and hypoparathyroidism [159]. In contrast, elevated serum concentrations occur in patients with sarcoidosis and tuberculosis. Antimycotic drugs, such as ketoconazole reduce the circulating 1,25(OH)2D concentration and have successfully been used to treat tuberculosis-associated hypercalcaemia [160, 161]. Pregnancy increases the serum 1,25(OH)2D concentration due to an induction of renal 1α-hydroxylase activity [162]. In addition, 1α-hydroxylase and the VDR are both expressed in the placenta. However, the function of placental 1,25(OH)2D synthesis is poorly understood. Serum 1,25(OH)2D levels are also higher in African individuals of all age-groups [163, 164], which may be due to increased PTH levels in this population [165]. However, the differences in the metabolism of vitamin, calcium and phosphate are insufficiently understood. Mutations of genes involved in 1,25(OH)2D metabolism also influence circulating 1,25(OH)2D levels (reviewed in [121]). These mutations cause rare hereditary metabolic bone conditions, like hereditary vitamin D-resistant rickets (VDR), vitamin-D-dependent rickets type 1A (CYP27B1), type 1B (CYP2R1) or idiopathic infantile hypercalcemia (CYP24A1). Mutations of the cell surface metalloproteinase PHEX cause X-linked hypophosphataemia (XLH), which is characterized by low-normal 1,25(OH)2D concentrations [166].
Measurement of 24,25(OH)2D and Calculation of the 24,25(OH)2D/25(OH)D Ratio
Clinical Utility
24,25(OH)2D is the principal catabolite of 25(OH)D that is formed through the action of CYP24A1 when sufficient amounts of 25(OH)D are available. Under physiological conditions the serum concentration of 24,25(OH)2D ranges at approximately 10% of the 25(OH)D concentration. Normally, 25(OH)D and 24,25(OH)2 are highly correlated [167, 168]. Therefore, the isolated measurement of 24,25(OH)2 does not provide superior information when compared to 25(OH)D. However, the simultaneous determination of both metabolites has been found useful to identify patients with reduced CYP24A1 activity. Affected individuals are characterized by a reduced ratio between 24,25(OH)2D and 25(OH)D [17]. This ratio is also known as vitamin D metabolite ratio (VMR). CYP24A1 loss-of-function mutations cause infantile hypercalcemia (IIH) and represent a genetic risk factor for serious adverse effects in response to vitamin D supplementation [169]. In hypercalcaemic patients, the parallel measurement of both vitamin D metabolites and calculation of the VMR allows distinguishing CYP24A1 deficiency from other causes of hypercalcaemia, such as vitamin D intoxication, granulomatous disease, Williams-Beuren syndrome or mutations of the SLC34A1 gene [18, 170]. Therefore, 24,25(OH)2D and 25(OH)D can be helpful in guiding genetic testing in patients with inherited hypercalcaemia.
In addition to the rather rare conditions mentioned before, Cavalier et al. have proposed the simultaneous quantitation of 24,25(OH)2D and 25(OH)D as a dynamic measure of vitamin D metabolism that aids the identification of individuals with functional vitamin D deficiency [15]. In a cohort of 1200 infants, children, adolescent and young adults, the vast majority of individuals with low 25(OH)D had 24,25(OH)2D concentrations below the lower level of quantitation. With increasing 25(OH)D levels, the percentage of individuals such a low 24,25(OH)2D concentration decreased and was negligible when serum 25(OH)D exceeded 21 ng/ml (52,5 nmol/L). Based on these results, the authors concluded that this cut-off corresponds to biochemical vitamin D sufficiency. However, Cavallier et al. have also shown that different individuals with the same 25(OH)D concentration can have detectable or undetectable 24,25(OH)2D levels, suggesting that they are not equally vitamin D sufficient. This observation questions the use of a fixed 25(OH)D cut-off and suggests that an individual evaluation of the vitamin D status based on a simultaneous analysis of 24,25(OH)2D and 25(OH)D might be more appropriate. Also, in black individuals, the parallel determination of both metabolites does better reflect the individual vitamin D status than the measurement of 25(OH)D alone. On average, blacks have a 40% lower 25(OH)D serum concentration, which is at least partially due to ethnic differences in the prevalence of common genetic VDBP polymorphisms [38]. Recent evidence from a cohort study of elderly community-dwelling individuals has shown that the VMR was associated with the risk of hip fracture whereas 25(OH)D was not [171]. In addition, higher 24,25(OH)2D, but not VMR, was associated with a higher bone mineral density (BMD). The management of renal patients may also benefit from the measurement of 24,25(OH)2D. In addition to a decreased production of 1,25(OH)2D, also vitamin D catabolism is impaired in patients with chronic kidney disease [172]. The Seattle Kidney Study has shown that patients with a 24,25(OH)2D serum concentration below the cohort median of 2.4 ng/ml had a significantly lower eGFR and an increased risk of mortality. Moreover, the inverse correlation of 24,25(OH)2D with PTH was stronger than that of 25(OH)D or 1,25(OH)2D.
Last, but not least, the VMR has also been proposed as an interesting tool to guide vitamin D supplementation at an individual level [173]. However, in existing vitamin D supplementation studies, neither 24,25(OH)2D nor VMR predicted the increase in serum 25(OH)D better than the baseline 25(OH) concentration [118, 174, 175]. In addition, neither 24,25(OH)2D nor VMR predicted the biological effect of vitamin D supplementation on bone and mineral metabolism better than 25(OH)D [176]. However, as most participants had detectable 24,25(OH)2D in serum, it cannot be excluded that they were vitamin D sufficient, which may have limited the diagnostic potential of this analyte. So far, no study compared the response of vitamin D supplementation in individuals with and without measureable concentrations of 24,25(OH)2D, but comparable serum 25(OH)D concentrations.
Current guidelines do not yet recommend the measurement of 24,25(OH)2D in clinical practice. However, existing studies suggest that this practice should be changed. The determination of 24,25(OH)2D and calculation of the VMR are helpful in the differential diagnosis of patients with hypercalcaemia and seem to improve risk prediction in conditions where vitamin D catabolism is impaired. In addition, the simultaneous measurement of 25(OH)D and 24,25(OH)2D may help to individualize the assessment of patients’ vitamin D status. In contrast, neither 24,25(OH)2D nor VMR can predict the response to vitamin D supplementation better than 25(OH)D.
Preanalytical Considerations
24,25(OH)2D and 25(OH)D share similar chemical properties. Both compounds are very stable in serum and can thus be kept for up to 2 months at temperatures between + 25 °C and − 80 °C [17, 177]. For long-term storage samples should be frozen. More than 2 freeze-and-thaw cycles should be avoided as the serum 24,25(OH)2D concentration decreases by more than 20%. So far, the effect of light exposure and sample matrix on the 24,25(OH)2D serum concentration has not been studied systematically. However, it is prudent to protect samples from light exposure. The influence of patient related preanalytical factors, such as food intake, menstrual cycle or sunlight exposure has not been studied systematically. However, it can be expected that in line with 25(OH)D, vitamin D supplementation and sunlight exposure will increase the synthesis of 24,25(OH)2D.
Analytical Considerations
Measurement of 24,25(OH)2D is typically performed by LC–MS/MS. Although this technology is considered the gold standard for the measurement of vitamin D and its metabolites, an interlaboratory assay comparison by Wise et al. has shown substantial systemic differences of up to 36% [178]. In order to improve the comparability of 24,25(OH)2D results, a standard reference material [179, 180] and a reference method [181] have been developed. In addition, first EQA programs have included this analyte in their portfolio [182]. The efficacy of these measures has been demonstrated by a recent comparison study of two in-house LC–MS/MS methods [16]. Both methods were properly validated and underwent regular proficiency testing. So far, immunoassays or other analytical methods for the measurement of this lowly concentrated analyte have yet not been developed.
Reference Values
So far, two studies have determined references ranges for the serum concentration of 24,25(OH)2D and the VMR using validated LC–MS/MS methods [183, 184]. In 1996, Tang et al. found an average 24,25(OH)2D serum concentration of 5.7 ± 3.4 nmol/L and calculated a reference interval (2.5th–97.5th percentile) of 1.1.-13.5 nmol/L in healthy young army recruits (504 females and 1492 males) with a mean age of 23 years [184]. The diagnostic cut-off for vitamin D sufficiency (serum 25(OH)D > 50 nmol/L) was > 4.2 nmol/L. Dirks et al. established a serum 24,25(OH)2D reference interval in 92 middle-aged adults that ranged from 0.4 to 8.9 nmol/L [183]. Both studies calculated the VMR as 25(OH)D/24,25(OH)2D and reported reference intervals of 10–33 [184] and 7–23 [183], respectively. In line with these results, an earlier study from Ketha et al. reported a VMR reference interval of 7–35, which was obtained in 91 adults with an age range of 28–86 years [185]. It should be mentioned that not all laboratories calculate the VMR in the same way, while some use the formula 25(OH)D/24,25(OH)2D, others do it the other way round 24,25(OH)2D/25(OH)D, which provides totally different figures. Moreover, the VMR is further limited by the fact the two vastly different numbers are divided by each other and thus small changes in 24,25(OH)2D result in a great change in VMR. Furthermore, imprecision of this ratio is affected by the imprecision of the two methods.
Measurement of Vitamin D Binding Protein (VDBP) and Calculation of Free and Bioavailable 25(OH)D
Role of the Marker
VDBP is a highly polymorphic protein with over 100 isoforms that is encoded by the VDBP gene on chromosome 4q12-q13. Structurally, the protein is closely related to albumin and alpha-fetoprotein [85]. It is mainly produced in the liver, but animal studies suggest that it can also be expressed by other tissues, such as kidney, testis, and adipose tissue [186]. The expression of VDBP is regulated by estrogens, which show higher concentrations during pregnancy and upon oral contraception [85].
As mentioned above, VDBP binds approximately 85% and albumin 15% of all circulating vitamin D metabolites [85]. Less than 0.1% of all 25(OH)D in the plasma of normal individuals is free. Similar to other steroid hormones, it is believed that only the free fraction can enter the cells to bind the cytosolic VDR. The free hormone hypothesis is supported by the observation that lacking VDBP does not necessarily cause functional vitamin D deficiency despite very low serum concentrations of 25(OH)D and 1,25(OH)2D [187,188,189]. In addition to its transport function of vitamin D metabolites, VDBP has also other functions including the renal megalin/cubilin-mediated reuptake of 25(OH)D in the kidneys,the scavenging of actin after muscular damage, neutrophil recruitment, complement 5a-mediated chemotaxis, fatty acid binding and formation of VDBP-Macrophage Activating Factor V (DBP-MAF) [85, 187]. For example, significant amounts of actin are released from damaged cells in response to trauma, sepsis, liver trauma, acute lung injury, preeclampsia, surgery, and burns. Upon polymerization, filamentous F-actin in combination with coagulation factor Va can promote disseminated intravascular coagulation and multiorgan failure. Through binding to VDBP, actin is rapidly cleared by the liver, lungs and spleen, thus preventing polymerization.
Based on the free hormone hypothesis [190] is has been proposed that free 25(OH)D might be a better marker than total 25(OH)D to reflect a patient’s current vitamin D status [38, 191]. For example, patients with liver cirrhosis have a 100% higher median of free 25(OH)D than unaffected controls, despite lower total 25(OH)D concentrations. Interestingly, these patients develop osteoporosis rather than osteomalacia, as it would be expected for vitamin D deficient patients with a low total 25(OH)D concentration [192]. In contrast, pregnant women have lower free 25(OH)D concentrations than non-pregnant controls [193]. Due to existing inconsistencies, the applicability of the free hormone hypothesis to vitamin D is still a matter of ongoing debate [153]. In contrast to other steroid hormones, the active metabolite 1,25(OH)2D is feedback regulated and the concentration of free hormone is so low that it may be insufficient for passive diffusion across the cell membrane and effective receptor binding [153]. In addition, it cannot be excluded that the megalin-dependent uptake of the VDBP-25(OH)D complex into cells is limited to the proximal renal tubule. For example, a vitamin D-dependent expression megalin and cubilin has been found in several other epithelia including placenta, parathyroid glands, pneumocytes, and epididymal epithelial cells [93, 194,195,196].
Considering the much lower affinity of 25(OH)D to albumin (Ka = 7 × 108/M) than to VDBP (Ka = 6 × 105/M), it is believed that albumin-bound 25(OH)D can easily dissociate from the protein and is thus available for metabolism. This concept has led to the term bioavailable 25(OH)D, which refers to all the circulating 25(OH)D that is not tightly bound to VDBP [197]. Several studies have used bioavailable 25(OH)D rather than free 25(OH)D [38, 198,199,200].
Free [201] and bioavailable [38] 25(OH)D are calculated from total 25(OH)D, albumin and VDBP:
For accuracy, the measurements of 25(OH)D and VDBP are best performed by LC–MS/MS. The polymorphic nature of VDBP results in numerous variants of the protein with Gc1f and Gc1s and Gc2 being most common. These variants are believed to differ in their affinity for 25(OH)D [202, 203]. The lowest kVDBP has been reported for the 1f/1f variant with 3.6 × 108/M, whereas 2/2 has supposedly the highest kVDBP with 11.2 × 108/M. However, different affinities of VDBP variants to 25(OH)D are a matter of ongoing scientific debate [85]. Nevertheless, VDBP haplotypes impact the concentrations of total 25(OH)D, free 25(OH)D, and VDBP. The 2/2 haplotype is associated with the lowest total and free 25(OH)D concentrations. The lowest free percentage was seen with the 1s/1s haplotype and the highest one with the 1f/1f haplotype. Furthermore, the different Gc alleles affect the response to vitamin D supplementation [204]. The clinical significance of these VDBP variants is insufficiently understood. Existing studies do not show differences in fracture rate [44, 205]. However, significant associations with numerous chronic disease, such as type 1 and 2 diabetes, osteoporosis, chronic obstructive lung disease, inflammatory bowel disease, some malignancies and tuberculosis, have been reported [206, 207].
Preanalytical Considerations
VDBP is relatively stable. Storing serum for 48 h at room temperature or refrigerated introduces only a small positive bias of < 10% [43]. Also, up to 2 freeze–thaw-cycles have no major impact on VDBP. Regarding LC–MS/MS methods, autosampler stability is also acceptable with < 16% loss during 24 h in a 96-well plate. In order to minimize the impact of preanalytical factors, samples should be processed rapidly after collection. The distribution of VDBP variants differs substantially between black and white individuals, which has important analytical implications (see below) and impacts free 25(OH)D levels. Haemolysis and lipaemia have also been reported to interfere with the measurement [43]. In addition, free 25(OH)D is less stable than protein-bound 25(OH)D and thus requires some precautions for direct measurement. Samples are light-sensitive and should be protected from UV-irradiation. Long term storage should be at − 70 °C [208]. To mitigate the impact of preanalytical factors on free 25(OH)D, measurement in fresh samples is preferred.
Analytical Considerations
The measurement of VDBP is challenging and LC–MS/MS is the preferred method for this analyte due to superior sensitivity and specificity [43]. For example, only LC–MS/MS can detect VDBP isoforms. However, LC–MS/MS analyses require experienced staff and expensive equipment, which are not widely available [208]. Immunoassays have been developed as easy-to-use alternative that allow a relatively high throughput. However, the use of immunoassays for the quantitation of a highly polymorphic protein, such as VDBP, is problematic, especially when the target epitope(s) are located near the polymorphic region [43]. For example, Henderson CM et al. have shown that assays with a monoclonal capture antibody may react differently with common variants, and thus deliver misleading results [43]. With a monoclonal immunoassay, blacks, which carry predominantly the Gc1f variant, have been reported to have substantially lower VDBP concentrations than whites [38, 209]. When using a validated LC–MS/MS method that can distinguish between VDBP variants, no ethnic differences in VDBP concentration were found. Although polyclonal assays are less influenced by isoforms and other genetic variations, they still deliver largely discrepant results when compared to LC–MS/MS with a median bias of + 50% [210]. Such differences have great impact on the calculation of free and bioavailable 25(OH)D [209, 211]. Therefore, free and bioavailable 25(OH)D concentrations should be determined on the basis of accurate VDBP and total 25(OH)D results obtained with validated LC–MS/MS methods.
The direct quantification of free 25(OH)D constitutes an analytical challenge, due to its very low concentration (0.02% to 0.09% of total 25(OH)D concentration) [193]. Even sensitive LC–MS/MS systems are not sufficient to accurately measure such low concentrations. Therefore, most studies calculate the free 25(OH)D concentration from measurements of total 25(OH)D, VDBP and albumin. However, the result is strongly dependent on the accuracy of the methods used for total 25(OH)D and VDBP. A comparison of calculated free 25(OH)D results obtained with four different VDBP immunoassays has shown differences of up to 50%. Direct methods for the measurement of free 25(OH)D have also been developed [212, 213]. Centrifugal ultrafiltration has been described in the 1980s and appears reasonably accurate [212]. However, this method is highly complex and expensive. A recently marketed ELISA from DiaSource showed good agreement with a dialysis-based reference method [213], but low concordance with indirect methods [211, 214]. Amongst 173 healthy women, Deming's regression showed a slope of 0.3 ± 0.03, and a correlation coefficient of 0.6 ± 0.06.
Expected Values
To date, reliable reference values for free 25(OH)D and VDBP have not yet been established, and several ranges have been proposed. Due to the lack of standardization, reference intervals have to be method-specific. In addition, differences between black and white individuals have been reported for some methods, but not for others. When measuring free 25(OH)D by direct immunoassay in 279 healthy controls with a median age of 36.6 years, 95% of the results ranged between 0.5 and 8.1 pg/ml, with VDBP concentrations measured by a polyclonal immunodiffusion assay between 190.8 and 395.2 µg/ml [193]. Amongst 2085 participants of the Healthy Aging in Neighbourhoods of Diversity across the Life Span cohort, blacks and whites showed mean ± standard error concentrations of 168 ± 3 µg/ml and 337 ± 5 µg/ml, respectively [38]. Bioavailable 25(OH)D-was 2.9 ± 0.1 ng/ml in blacks and 3.1 ± 0.1 ng/ml in whites. However, VDBP was measured by a monoclonal immunoassay that suffers from variable detection of different VDBP variants. In 173 healthy women, Peris et al. reported free 25(OH)D concentrations between 2 and 15 pg/ml using an indirect method whereas directly measured results ranged between 2 and 8 pg/ml [214]. Zeng et al. determined normal free 25(OH)D results on the basis of sufficient total 25(OH)D concentrations and suggested a range between 8.5 and 28.3 pg/ml (equivalent to 30 and 100 ng/ml of total 25(OH)D [74]. In our opinion, this range is rather high. In healthy children, Lopez-Molina et al. calculated that directly measured free 25(OH)D results above 9.8 pmol/L (equal to 3.9 pg/ml) correlate best with a total 25(OH)D concentration > 50 nmol/L [215].
Indications
Currently, routine measurement of VDBP, free and bioavailable 25(OH)D are not recommended due to unresolved analytical issues and the lack clinical evidence that supports an additional value beyond the measurement of total 25(OH)D. Potential areas of interest for the use of these markers are patients with liver or renal disease, which cause a decreased synthesis or loss of VDBP. Moreover, determining free 25(OH)D could also be useful for pregnant women or women on hormone therapy, because estrogens can upregulate VDBP levels. In the same line, elderly people could also benefit from assessing this marker since VDBP levels can be altered in this group due to a multifactorial effect [216, 217]. However, future clinical studies have to clarify the potential value of VDBP, free and bioavailable 25(OH)D in such patients.
Influence of VDBP in Pathological Conditions
Different variants of VDBP with different affinities for 25(OH)D influence the measurement of VDBP as well as the determination of free 25(OH)D. As the distribution of VDBP variants changes between blacks and whites, ethnicity needs to be considered when assessing these markers [38]. Based on the method used, differences in VDBP and free 25(OH)D have been found or not [38, 43, 211]. Changes in estrogen levels, such as in pregnancy, alter the expression of VDBP. Between the second and third trimester, VDBP levels increase up to two-fold. However, these variations are not linked to changes in free 25(OH)D, which remains stable or slightly decreases [85, 218, 219].
Critically ill patients with organ dysfunction also show substantial variation in serum VDBP when compared with normal individuals [220, 221]. Liver diseases, especially at late stages, often cause impaired protein synthesis, resulting in decreasing levels of VDBP and albumin. In turn, free 25(OH)D is increased in these patients compared to controls [85]. These changes are most pronounced in cirrhotic patients. Renal disease also decreases VDBP levels, via proteinuria and saturation of the maximal transport capacity of the megalin/cubilin system. Consistently, nephrotic patients present low total and free 25(OH)D levels [222]. However, in dialysis patients VDBP levels are comparable with healthy individuals [218]. Despite normal VDBP levels, immunoassays and LC–MS/MS methods vary in the quantitation of 25(OH)D in dialysis patients. Potential differences may be due to increased urea concentrations that interfere with the release of 25(OH)D from VDBP, the binding equilibrium or the reagents of immunoassays. In contrast, LC–MS/MS methods are not affected from such interferences.
Conclusion
The existence of multiple vitamin D metabolites with different biological activities and their binding to VDBP and other carriers makes the assessment of vitamin D status a challenging task. Despite promising results for alternative markers, such as 24,25(OH)D, VMR, free and bioavailable 25(OH)D, 25(OH)D remains the preferred analyte for this purpose. The assessment of 25(OH)D is recommended for patients with established metabolic bone disease or individuals with an increased risk of developing these conditions, and the monitoring of vitamin D supplementation. For non-bone-related diseases, the determination of 25(OH)D is not recommended. Despite recent standardization, the accurate measurement of 25(OH)D remains a critical issue in clinical practice. Properly validated LC–MS/MS methods are preferable due to their high sensitivity, specificity, and robustness for interferences from matrix effects. 1,25(OH)2D, 24,25(OH) 2D and the VMR provide functional information on vitamin D metabolism and are thus useful for the investigation of rare enzyme defects (24-hydroxylase deficiency) and some other diseases (e.g. sarcoidosis, tuberculosis). The determination of VDBP, free and bioavailable 25(OH)D is compromised by unresolved analytical issues, the lack of reference intervals and insufficient clinical evidence that justifies their use. Future research should address analytical standardization and explore the clinical value of novel markers of vitamin D metabolism.
Data availability
Availability of data and material is not applicable.
References
Holick M (2017) The vitamin D deficiency pandemic: approaches for diagnosis, treatment and prevention. Rev Endocr Metab Disord 18:153–165. https://doi.org/10.1007/S11154-017-9424-1
Crowe FL, Jolly K, Macarthur C et al (2019) Trends in the incidence of testing for vitamin D deficiency in primary care in the UK: a retrospective analysis of the Health Improvement Network (THIN), 2005–2015. BMJ Open. https://doi.org/10.1136/bmjopen-2018-028355
Zhao S, Gardner K, Taylor W et al (2015) Vitamin D assessment in primary care: changing patterns of testing. London J Prim Care (Abingdon) 7:15–22. https://doi.org/10.1080/17571472.2015.11493430
Schumacher LD, Jäger L, Meier R et al (2020) Trends and between-physician variation in laboratory testing: a retrospective longitudinal study in general practice. J Clin Med 9:1–11. https://doi.org/10.3390/jcm9061787
Pike JW, Christakos S (2017) Biology and mechanisms of action of the vitamin D hormone. Endocrinol Metab Clin North Am 46:815–843
Christakos S, Dhawan P, Verstuyf A et al (2016) Vitamin D: metabolism, molecular mechanism of action, and pleiotropic effects. Physiol Rev 96:365–408. https://doi.org/10.1152/PHYSREV.00014.2015
Girgis CM, Baldock PA, Downes M (2015) Vitamin D, muscle and bone: integrating effects in development, aging and injury. Mol Cell Endocrinol 410:3–10
Charoenngam N, Holick MF (2020) Immunologic effects of vitamin d on human health and disease. Nutrients 12:1–28
Charoenngam N, Shirvani A, Holick MF (2019) Vitamin D for skeletal and non-skeletal health: what we should know. J Clin Orthop Trauma 10:1082–1093
Christakos S, Seth T, Hirsch J et al (2013) Vitamin D biology revealed through the study of knockout and transgenic mouse models. Annu Rev Nutr 33:71–85. https://doi.org/10.1146/ANNUREV-NUTR-071812-161249
Muscogiuri G, Mitri J, Mathieu C et al (2014) Mechanisms in endocrinology: vitamin D as a potential contributor in endocrine health and disease. Eur J Endocrinol 171:R101–R110
Tuckey RC, Cheng CYS, Slominski AT (2019) The serum vitamin D metabolome: what we know and what is still to discover. J Steroid Biochem Mol Biol 186:4–21
Jenkinson C, Desai R, Slominski AT et al (2021) Simultaneous measurement of 13 circulating vitamin D3 and D2 mono and dihydroxy metabolites using liquid chromatography mass spectrometry. Clin Chem Lab Med 59:1642–1652. https://doi.org/10.1515/cclm-2021-0441
Makris K, Sempos C, Cavalier E (2020) The measurement of vitamin D metabolites: part I-metabolism of vitamin D and the measurement of 25-hydroxyvitamin D. Hormones (Athens) 19:81–96. https://doi.org/10.1007/S42000-019-00169-7
Cavalier E, Huyghebaert L, Rousselle O et al (2020) Simultaneous measurement of 25(OH)-vitamin D and 24,25(OH)2-vitamin D to define cut-offs for CYP24A1 mutation and vitamin D deficiency in a population of 1200 young subjects. Clin Chem Lab Med. https://doi.org/10.1515/CCLM-2019-0996
Zelzer S, Le Goff C, Peeters S et al (2021) Comparison of two LC-MS/MS methods for the quantification of 24,25-dihydroxyvitamin D3 in patients and external quality assurance samples. Clin Chem Lab Med. https://doi.org/10.1515/cclm-2021-0792
Zelzer S, Meinitzer A, Enko D et al (2020) Simultaneous determination of 24,25- and 25,26-dihydroxyvitamin D3 in serum samples with liquid-chromatography mass spectrometry—a useful tool for the assessment of vitamin D metabolism. J Chromatogr B Anal Technol Biomed Life Sci. https://doi.org/10.1016/j.jchromb.2020.122394
Kaufmann M, Schlingmann KP, Berezin L et al (2021) Differential diagnosis of vitamin D-related hypercalcemia using serum vitamin D metabolite profiling. J Bone Miner Res 36:1340–1350. https://doi.org/10.1002/JBMR.4306
Herrmann M, Farrell CJL, Pusceddu I et al (2017) Assessment of vitamin D status—a changing landscape. Clin Chem Lab Med 55:3–26
Altieri B, Cavalier E, Bhattoa HP et al (2020) Vitamin D testing: advantages and limits of the current assays. Eur J Clin Nutr 74:231–247
Zerwekh JE (2008) Blood biomarkers of vitamin D status. Am J Clin Nutr. https://doi.org/10.1093/ajcn/87.4.1087s
Horst RL, Reinhardt TA, Reddy GS (2005) Vitamin D metabolism. In: Feldman D, Pike JWGF (eds) Vitamin D, 2nd edn. Elsevier Academic Press, London, pp 15–36
Ross AC, Taylor CL, Yaktine AL, Del Valle HB (2011) Institute of Medicine (US) Committee to review dietary reference intakes for vitamin D and calcium. In: Dietary reference intakes for calcium and vitamin D. National Academies Press, US. https://doi.org/10.17226/13050
Holick MF (2002) Vitamin D: the underappreciated D-lightful hormone that is important for skeletal and cellular health. Curr Opin Endocrinol Diabetes 9:87–98
Willett AM (2005) Vitamin D status and its relationship with parathyroid hormone and bone mineral status in older adolescents. Proc Nutr Soc 64:193–203. https://doi.org/10.1079/pns2005420
National Institutes of Health. Office of dietary supplements (2014) Vitamin D. https://ods.od.nih.gov/factsheets/VitaminD-HealthProfessional/
Jones G, Prosser DE, Kaufmann M (2014) Cytochrome P450-mediated metabolism of vitamin D. J Lipid Res 55:13–31
Prentice A, Goldberg GR, Schoenmakers I (2008) Vitamin D across the lifecycle: physiology and biomarkers. Am J Clin Nutr. https://doi.org/10.1093/ajcn/88.2.500S
Kamao M, Hatakeyama S, Sakaki T et al (2005) Measurement and characterization of C-3 epimerization activity toward vitamin D3. Arch Biochem Biophys 436:196–205. https://doi.org/10.1016/j.abb.2005.01.017
Astecker N, Satyanarayana Reddy G, Herzig G et al (2000) 1α,25-Dihydroxy-3-epi-vitamin D3 a physiological metabolite of 1α,25-dihydroxyvitamin D3: its production and metabolism in primary human keratinocytes. Mol Cell Endocrinol 170:91–101. https://doi.org/10.1016/S0303-7207(00)00330-0
Lensmeyer G, Poquette M, Wiebe D, Binkley N (2012) The C-3 epimer of 25-hydroxyvitamin D 3 is present in adult serum. J Clin Endocrinol Metab 97:163–168. https://doi.org/10.1210/jc.2011-0584
Al-Zohily B, Al-Menhali A, Gariballa S et al (2020) Epimers of vitamin d: a review. Int J Mol Sci 21:470. https://doi.org/10.3390/ijms21020470
Masuda S, Kamao M, Schroeder NJ et al (2000) Characterization of 3-epi-1 α,25-dihydroxyvitamin d3 involved in 1 α,25-dihydroxyvitamin d3 metabolic pathway in cultured cell lines. Biol Pharm Bull 23:133–139. https://doi.org/10.1248/bpb.23.133
Norman AW, Bouillon R, Farach-Carson MC et al (1993) Demonstration that 1β,25-dihydroxyvitamin D3 is an antagonist of the nongenomic but not genomic biological responses and biological profile of the three A-ring diastereomers of 1α,25-dihydroxyvitamin D3. J Biol Chem 268:20022–20030. https://doi.org/10.1016/s0021-9258(20)80689-9
Nakagawa K, Sowa Y, Kurobe M et al (2001) Differential activities of 1α,25-dihydroxy-16-ene-vitamin D3 analogs and their 3-epimers on human promyelocytic leukemia (HL-60) cell differentiation and apoptosis. Steroids 66:327–337
Cheng JB, Motola DL, Mangelsdorf DJ, Russell DW (2003) De-orphanization of cytochrome P450 2R1: a microsomal vitamin D 25-hydroxylase. J Biol Chem 278:38084–38093. https://doi.org/10.1074/jbc.M307028200
Holick MF, Binkley NC, Bischoff-Ferrari HA et al (2011) Evaluation, treatment, and prevention of vitamin D deficiency: an endocrine society clinical practice guideline. J Clin Endocrinol Metab 96:1911–1930
Powe CE, Evans MK, Wenger J et al (2013) Vitamin D-binding protein and vitamin D status of black Americans and white Americans. N Engl J Med 369:1991–2000. https://doi.org/10.1056/nejmoa1306357
Hannan MT, Litman HJ, Araujo AB et al (2008) Serum 25-hydroxyvitamin D and bone mineral density in a racially and ethnically diverse group of men. J Clin Endocrinol Metab 93:40–46. https://doi.org/10.1210/jc.2007-1217
Bischoff-Ferrari HA, Dietrich T, Orav EJ, Dawson-Hughes B (2004) Positive association between 25-hydroxy vitamin D levels and bone mineral density: a population-based study of younger and older adults. Am J Med 116:634–639. https://doi.org/10.1016/j.amjmed.2003.12.029
Cauley JA, Lui LY, Ensrud KE et al (2005) Bone mineral density and the risk of incident nonspinal fractures in black and white women. J Am Med Assoc 293:2102–2108. https://doi.org/10.1001/jama.293.17.2102
Cauley JA, Danielson ME, Boudreau R et al (2011) Serum 25-hydroxyvitamin D and clinical fracture risk in a multiethnic cohort of women: the women’s health initiative (WHI). J Bone Miner Res 26:2378–2388. https://doi.org/10.1002/jbmr.449
Henderson CM, Lutsey PL, Misialek JR et al (2016) Measurement by a novel LC-MS/MS methodology reveals similar serum concentrations of Vitamin D-binding protein in blacks and whites. Clin Chem 62:179–187. https://doi.org/10.1373/clinchem.2015.244541
Leong A, Rehman W, Dastani Z et al (2014) The causal effect of vitamin D binding protein (DBP) levels on calcemic and cardiometabolic diseases: a Mendelian randomization study. PLoS Med. https://doi.org/10.1371/JOURNAL.PMED.1001751
Brown LL, Cohen B, Tabor D et al (2018) The vitamin D paradox in Black Americans: a systems-based approach to investigating clinical practice, research, and public health—expert panel meeting report. BMC Proc. https://doi.org/10.1186/s12919-018-0102-4
Zelzer S, Goessler W, Herrmann M (2018) Measurement of vitamin D metabolites by mass spectrometry, an analytical challenge. J Lab Precis Med 3:99–99. https://doi.org/10.21037/jlpm.2018.11.06
Klingberg E, Oleröd G, Konar J et al (2015) Seasonal variations in serum 25-hydroxy vitamin D levels in a Swedish cohort. Endocrine 49:800–808. https://doi.org/10.1007/S12020-015-0548-3
Cavalier E, Fraser CG, Bhattoa HP et al (2021) Analytical performance specifications for 25-hydroxyvitamin D examinations. Nutrients 13:431 13-43431. https://doi.org/10.3390/NU13020431
Datta P, Philipsen PA, Olsen P et al (2017) The half-life of 25(OH)D after UVB exposure depends on gender and vitamin D receptor polymorphism but mainly on the start level. Photochem Photobiol Sci 16:985–995. https://doi.org/10.1039/C6PP00258G
Wielders JPM, Wijnberg FA (2009) Preanalytical stability of 25(OH)-vitamin D3 in human blood or serum at room temperature: solid as a rock. Clin Chem 55:1584–1585. https://doi.org/10.1373/CLINCHEM.2008.117366
Cavalier E (2021) Long-term stability of 25-hydroxyvitamin D: Importance of the analytical method and of the patient matrix. Clin Chem Lab Med 59:E389–E391
Antoniucci DM, Black DM, Sellmeyer DE (2005) Serum 25-hydroxyvitamin D is unaffected by multiple freeze-thaw cycles. Clin Chem 51:258–261. https://doi.org/10.1373/CLINCHEM.2004.041954
Colak A, Toprak B, Dogan N, Ustuner F (2013) Effect of sample type, centrifugation and storage conditions on vitamin D concentration. Biochem Medica 23:321–325. https://doi.org/10.11613/BM.2013.039
Lewis JG, Elder PA (2008) Serum 25-OH vitamin D2 and D3 are stable under exaggerated conditions. Clin Chem 54:1931–1932
Cavalier E, Lukas P, Bekaert AC et al (2016) Analytical and clinical evaluation of the new Fujirebio Lumipulse®G non-competitive assay for 25(OH)-vitamin D and three immunoassays for 25(OH)D in healthy subjects, osteoporotic patients, third trimester pregnant women, healthy African subjects, hemodia. Clin Chem Lab Med 54:1347–1355. https://doi.org/10.1515/CCLM-2015-0923
Moreau E, Bächer S, Mery S et al (2016) Performance characteristics of the VIDAS® 25-OH vitamin D total assay—comparison with four immunoassays and two liquid chromatography-tandem mass spectrometry methods in a multicentric study. Clin Chem Lab Med 54:45–53. https://doi.org/10.1515/CCLM-2014-1249/PDF
Tsugawa N, Shiraki M, Suhara Y et al (2008) Low plasma phylloquinone concentration is associated with high incidence of vertebral fracture in Japanese women. J Bone Miner Metab 26:79–85. https://doi.org/10.1007/s00774-007-0790-8
Tai SSC, Bedner M, Phinney KW (2010) Development of a candidate reference measurement procedure for the determination of 25-hydroxyvitamin D3 and 25-hydroxyvitamin D 2 in human serum using isotope-dilution liquid chromatography tandem mass spectrometry. Anal Chem 82:1942–1948. https://doi.org/10.1021/ac9026862
Binkley N, Dawson-Hughes B, Durazo-Arvizu R et al (2017) Vitamin D measurement standardization: the way out of the chaos. J Steroid Biochem Mol Biol 173:117–121. https://doi.org/10.1016/J.JSBMB.2016.12.002
Wise SA, Camara JE, Sempos CT et al (2021) Vitamin D standardization program (VDSP) intralaboratory study for the assessment of 25-hydroxyvitamin D assay variability and bias. J Steroid Biochem Mol Biol. https://doi.org/10.1016/J.JSBMB.2021.105917
Volmer DA, Mendes LRBC, Stokes CS (2015) Analysis of vitamin D metabolic markers by mass spectrometry: current techniques, limitations of the “gold standard” method, and anticipated future directions. Mass Spectrom Rev 34:2–23. https://doi.org/10.1002/mas.21408
Zhang S, Jian W, Sullivan S et al (2014) Development and validation of an LC-MS/MS based method for quantification of 25 hydroxyvitamin D2 and 25 hydroxyvitamin D3 in human serum and plasma. J Chromatogr B Anal Technol Biomed Life Sci 961:62–70. https://doi.org/10.1016/j.jchromb.2014.05.006
Kobold U (2012) Approaches to measurement of Vitamin D concentrations mass spectrometry. Scand J Clin Lab Invest 72:54–59
Giuliani S, Barbieri V, Di Pierro AM et al (2019) LC–MS/MS based 25(OH)D status in a large Southern European outpatient cohort: gender- and age-specific differences. Eur J Nutr 58:2511. https://doi.org/10.1007/S00394-018-1803-1
Skuladottir SS, Ramel A, Eymundsdottir H et al (2021) Serum 25-hydroxy-vitamin D status and incident hip fractures in elderly adults: looking beyond bone mineral density. J Bone Miner Res. https://doi.org/10.1002/JBMR.4450
Wang N, Chen Y, Ji J et al (2020) The relationship between serum vitamin D and fracture risk in the elderly: a meta-analysis. J Orthop Surg Res. https://doi.org/10.1186/S13018-020-01603-Y
Feng Y, Cheng G, Wang H, Chen B (2017) The associations between serum 25-hydroxyvitamin D level and the risk of total fracture and hip fracture. Osteoporos Int 28:1641–1652. https://doi.org/10.1007/S00198-017-3955-X
Valcour A, Blocki F, Hawkins DM, Rao SD (2012) Effects of age and serum 25-OH-vitamin D on serum parathyroid hormone levels. J Clin Endocrinol Metab 97:3989–3995. https://doi.org/10.1210/JC.2012-2276
Pilz S, Dobnig H, Fischer JE et al (2008) Low vitamin d levels predict stroke in patients referred to coronary angiography. Stroke 39:2611–2613. https://doi.org/10.1161/STROKEAHA.107.513655
Herrmann M, Sullivan DR, Veillard AS et al (2015) Serum 25-hydroxyvitamin D: a predictor of macrovascular and microvascular complications in patients with type 2 diabetes. Diabetes Care 38:521–528. https://doi.org/10.2337/DC14-0180/-/DC1
O’Hartaigh B, Neil Thomas G, Silbernagel G et al (2013) Association of 25-hydroxyvitamin D with type 2 diabetes among patients undergoing coronary angiography: cross-sectional findings from the LUdwigshafen Risk and Cardiovascular Health (LURIC) study. Clin Endocrinol (Oxf) 79:192–198. https://doi.org/10.1111/CEN.12024
Vinh quôc Luong K, Thi Hoàng Nguyên L (2012) Vitamin D and Parkinson’s disease. J Neurosci Res 90:2227–2236
Harrison SR, Li D, Jeffery LE et al (2020) Vitamin D, autoimmune disease and rheumatoid arthritis. Calcif Tissue Int 106:58–75. https://doi.org/10.1007/S00223-019-00577-2
Zeng S, Chu C, Doebis C et al (2021) Reference values for free 25-hydroxy-vitamin D based on established total 25-hydroxy-vitamin D reference values. J Steroid Biochem Mol Biol 210:105877. https://doi.org/10.1016/j.jsbmb.2021.105877
Ross AC, Manson JAE, Abrams SA et al (2011) The 2011 report on dietary reference intakes for calcium and vitamin D from the institute of medicine: what clinicians need to know. J Clin Endocrinol Metab 96:53–58. https://doi.org/10.1210/JC.2010-2704
Rizzoli R, Boonen S, Brandi ML et al (2013) Vitamin D supplementation in elderly or postmenopausal women: A 2013 update of the 2008 recommendations from the European Society for Clinical and Economic Aspects of Osteoporosis and Osteoarthritis (ESCEO). Curr Med Res Opin 29:305–313
Maeda SS, Borba VZC, Camargo MBR et al (2014) Recomendações da Sociedade Brasileira de Endocrinologia e Metabologia (SBEM) para o diagnóstico e tratamento da hipovitaminose D. Arq Bras Endocrinol Metabol 58:411–433. https://doi.org/10.1590/0004-2730000003388
Pérez-López FR, Brincat M, Erel CT et al (2012) EMAS position statement: vitamin D and postmenopausal health. Maturitas 71:83–88. https://doi.org/10.1016/j.maturitas.2011.11.002
Aguiar M, Atapattu N, Bhatia V et al (2016) Global consensus recommendations on prevention and management of nutritional rickets. J Clin Endocrinol Metab 101:394–415. https://doi.org/10.1210/jc.2015-2175
(2014) Recommendations abstracted from the american geriatrics society consensus statement on vitamin D for prevention of falls and their consequences. J Am Geriatr Soc 62:147–152. https://doi.org/10.1111/jgs.12631
Płudowski P, Karczmarewicz E, Bayer M et al (2013) Practical guidelines for the supplementation of vitamin D and the treatment of defcits in Central Europe—recommended vitamin D intakes in the general population and groups at risk of vitamin D deficiency. Endokrynol Pol 64:319–327. https://doi.org/10.5603/EP.2013.0012
Ramasamy I (2020) Vitamin D metabolism and guidelines for vitamin D supplementation. Clin Biochem Rev 41:103–126
Sempos CT, Heijboer AC, Bikle DD et al (2018) Vitamin D assays and the definition of hypovitaminosis D: results from the first international conference on controversies in vitamin D. Br J Clin Pharmacol 84:2194–2207
Souberbielle JC, Deschenes G, Fouque D et al (2016) Recommendations for the measurement of blood 25-OH vitamin D. Ann Biol Clin (Paris) 74:7–19. https://doi.org/10.1684/abc.2015.1107
Bikle DD, Schwartz J (2019) Vitamin D binding protein, total and free vitamin D levels in different physiological and pathophysiological conditions. Front Endocrinol (Lausanne) 10:317. https://doi.org/10.3389/FENDO.2019.00317/BIBTEX
Roizen JD, Casella A, Lai M et al (2018) Decreased serum 25-hydroxyvitamin D in aging male mice is associated with reduced hepatic Cyp2r1 abundance. Endocrinology 159:3083–3089. https://doi.org/10.1210/en.2017-03028
Roizen JD, Long C, Casella A et al (2019) Obesity Decreases hepatic 25-hydroxylase activity causing low serum 25-hydroxyvitamin D. J Bone Miner Res 34:1068–1073. https://doi.org/10.1002/jbmr.3686
Aatsinki S-M, Elkhwanky M-S, Kummu O et al (2019) Fasting-induced transcription factors repress vitamin D bioactivation, a mechanism for vitamin d deficiency in diabetes. Diabetes 68:918–931. https://doi.org/10.2337/DB18-1050
Best CM, Pressman EK, Queenan RA et al (2019) Longitudinal changes in serum vitamin D binding protein and free 25-hydroxyvitamin D in a multiracial cohort of pregnant adolescents. J Steroid Biochem Mol Biol 186:79–88. https://doi.org/10.1016/j.jsbmb.2018.09.019
Moon RJ, Crozier SR, Dennison EM et al (2015) Tracking of 25-hydroxyvitamin D status during pregnancy: the importance of vitamin D supplementation. Am J Clin Nutr 102:1081–1087. https://doi.org/10.3945/AJCN.115.115295
Papapetrou PD (2010) The interrelationship of serum 1,25-dihydroxyvitamin D, 25-hydroxyvitamin D and 24,25-dihydroxyvitamin D in pregnancy at term: a meta-analysis. Hormones (Athens) 9:136–144. https://doi.org/10.14310/HORM.2002.1263
Figueiredo ACC, Cocate PG, Adegboye ARA et al (2018) Changes in plasma concentrations of 25-hydroxyvitamin D and 1,25-dihydroxyvitamin D during pregnancy: a Brazilian cohort. Eur J Nutr 57:1059–1072. https://doi.org/10.1007/s00394-017-1389-z
Park H, Wood MR, Malysheva OV et al (2017) Placental vitamin D metabolism and its associations with circulating vitamin D metabolites in pregnant women. Am J Clin Nutr 106:1439–1487. https://doi.org/10.3945/ajcn
Kovacs CS (2014) Bone development and mineral homeostasis in the fetus and neonate: roles of the calciotropic and phosphotropic hormones. Physiol Rev 94:1143–1218. https://doi.org/10.1152/PHYSREV.00014.2014
Melamed ML, Chonchol M, Gutiérrez OM et al (2018) The role of vitamin D in CKD stages 3 to 4: report of a scientific workshop sponsored by the national kidney foundation. Am J Kidney Dis 72:834–845
Ali FN, Arguelles LM, Langman CB, Price HE (2009) Vitamin D deficiency in children with chronic kidney disease: uncovering an epidemic. Pediatrics 123:791–796. https://doi.org/10.1542/peds.2008-0634
Christodoulou M, Aspray TJ, Schoenmakers I (2021) Vitamin D supplementation for patients with chronic kidney disease: a systematic review and meta-analyses of trials investigating the response to supplementation and an overview of guidelines. Calcif Tissue Int 109:157–178. https://doi.org/10.1007/S00223-021-00844-1/FIGURES/4
Johnson JM, Maher JW, DeMaria EJ et al (2006) The long-term effects of gastric bypass on vitamin D metabolism. Ann Surg 243:701. https://doi.org/10.1097/01.SLA.0000216773.47825.C1
Schafer AL (2017) Vitamin D and intestinal calcium transport after bariatric surgery. J Steroid Biochem Mol Biol 173:202–210. https://doi.org/10.1016/J.JSBMB.2016.12.012
Borges JLC, de Miranda IS, Sarquis MMS et al (2018) Obesity, bariatric surgery, and vitamin D. J Clin Densitom 21:157–162. https://doi.org/10.1016/J.JOCD.2017.03.001
Manson JE, Cook NR, Lee I-M et al (2019) Marine n−3 fatty acids and prevention of cardiovascular disease and cancer. N Engl J Med 380:23–32. https://doi.org/10.1056/nejmoa1811403
Scragg R, Stewart AW, Waayer D et al (2017) Effect of monthly high-dose vitamin D supplementation on cardiovascular disease in the vitamin D assessment study: a randomized clinical trial. JAMA Cardiol 2:608–616. https://doi.org/10.1001/jamacardio.2017.0175
Pilz S, Trummer C, Theiler-Schwetz V et al (2022) Critical appraisal of large vitamin D randomized controlled trials. Nutrients 14:303
Autier P, Boniol M, Pizot C, Mullie P (2014) Vitamin D status and ill health: a systematic review. Lancet Diabetes Endocrinol 2:76–89. https://doi.org/10.1016/S2213-8587(13)70165-7
Bae JH, Choe HJ, Holick MF, Lim S (2022) Association of vitamin D status with COVID-19 and its severity: vitamin D and COVID-19: a narrative review. Rev Endocr Metab Disord 1:1–21. https://doi.org/10.1007/s11154-021-09705-6
Zelzer S, Prüller F, Curcic P et al (2021) Vitamin D metabolites and clinical outcome in hospitalized COVID-19 patients. Nutrients 13:2129. https://doi.org/10.3390/nu13072129
Hewison M, Burke F, Evans KN et al (2007) Extra-renal 25-hydroxyvitamin D3–1α-hydroxylase in human health and disease. J Steroid Biochem Mol Biol 103:316–321. https://doi.org/10.1016/j.jsbmb.2006.12.078
Myrtle JF, Norman AW (1971) Vitamin D: A cholecalciferol metabolite highly active in promoting intestinal calcium transport. Science 171:79–82. https://doi.org/10.1126/science.171.3966.79
Mizwicki MT, Bula CM, Bishop JE, Norman AW (2005) A perspective on how the vitamin D sterol/vitamin D receptor (VDR) conformational ensemble model can potentially be used to understand the structure-function results of A-ring modified Vitamin D sterols. J Steroid Biochem Mol Biol 97:69–82
Ralston SH, Uitterlinden AG (2010) Genetics of osteoporosis. Endocr Rev 31:629–662
Altieri B, Grant WB, Della Casa S et al (2017) Vitamin D and pancreas: the role of sunshine vitamin in the pathogenesis of diabetes mellitus and pancreatic cancer. Crit Rev Food Sci Nutr 57:3472–3488. https://doi.org/10.1080/10408398.2015.1136922
Tirabassi G, Salvio G, Altieri B et al (2017) Adrenal disorders: is there Any role for vitamin D? Rev Endocr Metab Disord 18:355–362
Muscogiuri G, Altieri B, de Angelis C et al (2017) Shedding new light on female fertility: the role of vitamin D. Rev Endocr Metab Disord 18:273–283. https://doi.org/10.1007/S11154-017-9407-2
Seamans KM, Cashman KD (2009) Existing and potentially novel functional markers of vitamin D status: a systematic review. Am J Clin Nutr. https://doi.org/10.3945/ajcn.2009.27230D
Chapuy MC, Chapuy P, Meunier PJ (1987) Calcium and vitamin D supplements: effects on calcium metabolism in elderly people. Am J Clin Nutr 46:324–328. https://doi.org/10.1093/AJCN/46.2.324
Barger-Lux MJ, Heaney RP, Dowell S et al (1998) Vitamin D and its major metabolites: serum levels after graded oral dosing in healthy men. Osteoporos Int 8:222–230. https://doi.org/10.1007/S001980050058
Vieth R, Ladak Y, Walfish PG (2003) Age-related changes in the 25-hydroxyvitamin D versus parathyroid hormone relationship suggest a different reason why older adults require more vitamin D. J Clin Endocrinol Metab 88:185–191. https://doi.org/10.1210/JC.2002-021064
Vaes AMM, Tieland M, de Regt MF et al (2018) Dose–response effects of supplementation with calcifediol on serum 25-hydroxyvitamin D status and its metabolites: a randomized controlled trial in older adults. Clin Nutr 37:808–814. https://doi.org/10.1016/j.clnu.2017.03.029
Saponaro F, Saba A, Zucchi R (2020) An update on vitamin d metabolism. Int J Mol Sci 21:1–19
Lee SM, Riley EM, Meyer MB et al (2015) 1,25-Dihydroxyvitamin D3 controls a cohort of vitamin D receptor target genes in the proximal intestine that is enriched for calcium-regulating components. J Biol Chem 290:18199–18215. https://doi.org/10.1074/JBC.M115.665794
Goltzman D (2018) Functions of vitamin D in bone. Histochem Cell Biol 149:305–312
Sonoda J, Pei L, Evans RM (2008) Nuclear receptors: decoding metabolic disease. FEBS Lett 582:2–9
Goltzman D, Mannstadt M, Marcocci C (2018) Physiology of the calcium-parathyroid hormone-vitamin D axis. Front Horm Res 50:1–13. https://doi.org/10.1159/000486060
van de Peppel J, van Leeuwen JPTM (2014) Vitamin D and gene networks in human osteoblasts. Front Physiol. https://doi.org/10.3389/fphys.2014.00137
Suda T, Takahashi N, Abe E (1992) Role of vitamin D in bone resorption. J Cell Biochem 49:53–58. https://doi.org/10.1002/jcb.240490110
Holick MF (2010) Vitamin D: evolutionary, physiological and health perspectives. Curr Drug Targets 12:4–18. https://doi.org/10.2174/138945011793591635
Yang R, Chen J, Zhang J et al (2020) 1,25-Dihydroxyvitamin D protects against age-related osteoporosis by a novel VDR-Ezh2-p16 signal axis. Aging Cell. https://doi.org/10.1111/ACEL.13095
Chen L, Yang R, Qiao W et al (2019) 1,25-Dihydroxyvitamin D exerts an antiaging role by activation of Nrf2-antioxidant signaling and inactivation of p16/p53-senescence signaling. Aging Cell. https://doi.org/10.1111/acel.12951
Colotta F, Jansson B, Bonelli F (2017) Modulation of inflammatory and immune responses by vitamin D. J Autoimmun 85:78–97
Munger KL, Levin LI, Hollis BW et al (2006) Serum 25-hydroxyvitamin D levels and risk of multiple sclerosis. J Am Med Assoc 296:2832–2838. https://doi.org/10.1001/jama.296.23.2832
Kautzky-willer A, Pacini G, Barnas U, et al (1995) Intravenous calcitriol normalizes insulin sensitivity in uremic patients. Kidney Int 47:200–206. https://doi.org/10.1038/ki.1995.24
Bricio-Barrios JA, Palacios-Fonseca AJ, del Toro-Equihua M, Sanchez-Ramirez CA (2016) Effect of calcitriol supplementation on blood pressure in older adults. J Nutr Gerontol Geriatr 35:243–252. https://doi.org/10.1080/21551197.2016.1206499
Ashraf A, Alvarez JA (2010) Role of vitamin D in insulin secretion and insulin sensitivity for glucose homeostasis. Int J Endocrinol. https://doi.org/10.1155/2010/351385
Wang TJ (2016) Vitamin D and cardiovascular disease. Annu Rev Med 67:261–272. https://doi.org/10.1146/annurev-med-051214-025146
Thomas RL, Jiang L, Adams JS et al (2020) Vitamin D metabolites and the gut microbiome in older men. Nat Commun 11:1–10. https://doi.org/10.1038/s41467-020-19793-8
Ooi JH, Li Y, Rogers CJ, Cantorna MT (2013) Vitamin D regulates the gut microbiome and protects mice from dextran sodium sulfate-induced coliti. J Nutr 143:1679–1686. https://doi.org/10.3945/jn.113.180794
Davies M, Heys SE, Selby PL et al (1997) Increased catabolism of 25-hydroxyvitamin D in patients with partial gastrectomy and elevated 1,25-dihydroxyvitamin D levels. Implications for metabolic bone disease. J Clin Endocrinol Metab 82:209–212. https://doi.org/10.1210/JCEM.82.1.3644
Brumbaugh PF, Haussler DH, Bressler R, Haussler MR (1974) Radioreceptor assay for 1 alpha,25-dihydroxyvitamin D3. Science 183:1089–1091. https://doi.org/10.1126/SCIENCE.183.4129.1089
Dokoh S, Morita R, Fukunaga M et al (1978) Competitive protein binding assay for 1,25-dihydroxy-vitamin D in human plasma. Endocrinol Jpn 25:431–436. https://doi.org/10.1507/ENDOCRJ1954.25.431
Peacock M, Taylor GA, Brown W (1980) Plasma 1,25(OH)2 vitamin D measured by radioimmunoassay and cytosol radioreceptor assay in normal subjects and patients with primary hyperparathyroidism and renal failure. Clin Chim Acta 101:93–101. https://doi.org/10.1016/0009-8981(80)90060-1
Hussein H, Ibrahim F, Boudou P (2015) Evaluation of a new automated assay for the measurement of circulating 1,25-dihydroxyvitamin D levels in daily practice. Clin Biochem 48:1160–1162. https://doi.org/10.1016/j.clinbiochem.2015.05.018
Dabek JT, Harkonen M, Wahlroos AH (1981) Assay for plasma 25-hydroxyvitamin D2 and 25-hydroxyvitamin D3 by “high-performance” liquid chromatography. Clin Chem 27:1346–1351
Eisman JA, Shepard RM, DeLuca HF (1977) Determination of 25-hydroxyvitamin D2 and 25-hydroxyvitamin D3 in human plasma using high-pressure liquid chromatography. Anal Biochem 80:298–305. https://doi.org/10.1016/0003-2697(77)90648-0
Jones G (1978) Assay of vitamins D2 and D3, and 25-hydroxyvitamins D2 and D3 in human plasma by high-performance liquid chromatography. Clin Chem 24:287–298. https://doi.org/10.1093/CLINCHEM/24.2.287
Oftebro H, Falch JA, Holmberg I, Haug E (1988) Validation of a radioreceptor assay for 1,25-dihydroxyvitamin D using selected ion monitoring GC-MS. Clin Chim Acta 176:157–168. https://doi.org/10.1016/0009-8981(88)90203-3
Maunsell Z, Wright DJ, Rainbow SJ (2005) Routine isotope-dilution liquid chromatography-tandem mass spectrometry assay for simultaneous measurement of the 25-hydroxy metabolites of vitamins D2 and D3. Clin Chem 51:1683–1690. https://doi.org/10.1373/CLINCHEM.2005.052936
Tsugawa N, Okano T (2006) Bone and bone related biochemical examinations. Hormone and hormone related substances. Vitamin D (25D, 1,25D); measurements and clinical significances. Clin Calcium 16:36–42
Hollis BW, Horst RL (2007) The assessment of circulating 25(OH)D and 1,25(OH)2D: where we are and where we are going. J Steroid Biochem Mol Biol 103:473. https://doi.org/10.1016/J.JSBMB.2006.11.00
Valcour A, Zierold C, Podgorski AL et al (2016) A novel, fully-automated, chemiluminescent assay for the detection of 1,25-dihydroxyvitamin D in biological samples. J Steroid Biochem Mol Biol 164:120–126. https://doi.org/10.1016/J.JSBMB.2015.08.005
Zittermann A, Ernst JB, Becker T et al (2016) Measurement of circulating 1,25-Dihydroxyvitamin D: comparison of an automated method with a liquid chromatography tandem mass spectrometry method. Int J Anal Chem 2016:25. https://doi.org/10.1155/2016/8501435
Dirks NF, Ackermans MT, Lips P et al (2018) The when, what & how of measuring vitamin D metabolism in clinical medicine. Nutrients. https://doi.org/10.3390/NU10040482
Zittermann A, Ernst JB, Becker T et al (2016) Measurement of circulating 1,25-Dihydroxyvitamin D: comparison of an automated method with a liquid chromatography tandem mass spectrometry method. Int J Anal Chem. https://doi.org/10.1155/2016/8501435
Strathmann FG, Laha TJ, Hoofnagle AN (2011) Quantification of 1α,25-dihydroxy vitamin D by immunoextraction and liquid chromatography-tandem mass spectrometry. Clin Chem 57:1279–1285. https://doi.org/10.1373/CLINCHEM.2010.161174
Pauwels S, Jans I, Billen J et al (2017) 1β,25-Dihydroxyvitamin D3: a new vitamin D metabolite in human serum. J Steroid Biochem Mol Biol 173:341–348. https://doi.org/10.1016/j.jsbmb.2017.02.004
Dirks NF, Martens F, Vanderschueren D et al (2016) Determination of human reference values for serum total 1,25-dihydroxyvitamin D using an extensively validated 2D ID-UPLC–MS/MS method. J Steroid Biochem Mol Biol 164:127–133
Higgins V, Truong D, White-Al Habeeb NMA et al (2018) Pediatric reference intervals for 1,25-dihydroxyvitamin D using the DiaSorin LIAISON XL assay in the healthy CALIPER cohort. Clin Chem Lab Med 56:964–972. https://doi.org/10.1515/cclm-2017-0767
Taylor-Miller T, Allgrove J (2021) Endocrine diseases of newborn: epidemiology, pathogenesis, therapeutic options, and outcome “current insights into disorders of calcium and phosphate in the newborn.” Front Pediatr 9:42
Taylor-Miller T, Allgrove J (2021) Endocrine diseases of newborn: Epidemiology, pathogenesis, therapeutic options, and outcome “Current Insights Into Disorders of Calcium and Phosphate in the Newborn.” Front Pediatr 9:600490. https://doi.org/10.3389/fped.2021.600490
Jean G, Souberbielle JC, Chazot C (2017) Vitamin D in chronic kidney disease and dialysis patients. Nutrients 9:328
Glass AR, Cerletty JM, Elliott W et al (1990) Ketoconazole reduces elevated serum levels of 1,25-dihydroxyvitamin D in hypercalcemic sarcoidosis. J Endocrinol Invest 13:407–413. https://doi.org/10.1007/BF03350691
Saggese G, Bertelloni S, Baroncelli GI, Nero G (1993) Ketoconazole decreases the serum ionized calcium and 1,25-dihydroxyvitamin D levels in tuberculosis-associated hypercalcemia. Am J Dis Child 147:270–273. https://doi.org/10.1001/ARCHPEDI.1993.02160270032013
Ganguly A, Tamblyn JA, Finn-Sell S et al (2018) Vitamin D, the placenta and early pregnancy: effects on trophoblast function. J Endocrinol 236:R93–R103. https://doi.org/10.1530/JOE-17-0491
Kleerekoper M, Nelson DA, Peterson EL et al (1994) Reference data for bone mass, calciotropic hormones, and biochemical markers of bone remodeling in older (55–75) postmenopausal white and black women. J Bone Miner Res 9:1267–1276. https://doi.org/10.1002/JBMR.5650090817
Bikle DD, Ettinger B, Sidney S et al (1999) Differences in calcium metabolism between black and white men and women. Miner Electrolyte Metab 25:178–184. https://doi.org/10.1159/000057442
Aloia J, Mikhail M, Dhaliwal R et al (2015) Free 25(OH)D and the vitamin D paradox in African Americans. J Clin Endocrinol Metab 100:3356–3363. https://doi.org/10.1210/JC.2015-2066
Schmitt CP, Mehls O (2004) The enigma of hyperparathyroidism in hypophosphatemic rickets. Pediatr Nephrol 19:473–477. https://doi.org/10.1007/S00467-004-1443-Y
De Boer IH, Sachs MC, Chonchol M et al (2014) Estimated GFR and circulating 24,25-dihydroxyvitamin D3 concentration: a participant-level analysis of 5 cohort studies and clinical trials. Am J Kidney Dis 64:187–197. https://doi.org/10.1053/j.ajkd.2014.02.015
Berg AH, Powe CE, Evans MK et al (2015) 24,25-dihydroxyvitamin D3and vitamin D status of community-dwelling black and white Americans. Clin Chem 61:877–884. https://doi.org/10.1373/clinchem.2015.240051
Schlingmann KP, Kaufmann M, Weber S et al (2011) Mutations in CYP24A1 and idiopathic infantile hypercalcemia. N Engl J Med 365:410–421. https://doi.org/10.1056/nejmoa1103864
Herrmann M (2019) Towards a personalized assessment of vitamin D status. Clin Chem Lab Med. https://doi.org/10.1515/CCLM-2019-1213
Ginsberg C, Katz R, de Boer IH et al (2018) The 24,25 to 25-hydroxyvitamin D ratio and fracture risk in older adults: the cardiovascular health study. Bone 107:124–130. https://doi.org/10.1016/j.bone.2017.11.011
Bosworth CR, Levin G, Robinson-Cohen C et al (2012) The serum 24,25-dihydroxyvitamin D concentration, a marker of vitamin D catabolism, is reduced in chronic kidney disease. Kidney Int 82:693–700. https://doi.org/10.1038/ki.2012.193
Wagner D, Hanwell HE, Schnabl K et al (2011) The ratio of serum 24,25-dihydroxyvitamin D(3) to 25-hydroxyvitamin D(3) is predictive of 25-hydroxyvitamin D(3) response to vitamin D(3) supplementation. J Steroid Biochem Mol Biol 126:72–77. https://doi.org/10.1016/J.JSBMB.2011.05.003
Francic V, Ursem SR, Dirks NF et al (2019) The effect of vitamin D supplementation on its metabolism and the vitamin D metabolite ratio. Nutrients. https://doi.org/10.3390/nu11102539
Lehmann U, Riedel A, Hirche F et al (2016) Vitamin D3 supplementation: response and predictors of vitamin D3 metabolites—a randomized controlled trial. Clin Nutr 35:351–358. https://doi.org/10.1016/j.clnu.2015.04.021
Aloia J, Fazzari M, Shieh A et al (2017) The vitamin D metabolite ratio (VMR) as a predictor of functional biomarkers of bone health. Clin Endocrinol (Oxf) 86:674–679. https://doi.org/10.1111/cen.13319
Mena-Bravo A, Calderón-Santiago M, Luque de Castro MD, Priego-Capote F (2019) Evaluation of short-term storage prior to analysis of vitamin D3 and metabolites in human serum by liquid chromatography coupled to tandem mass spectrometry. Talanta 198:344–349. https://doi.org/10.1016/J.TALANTA.2019.02.030
Wise SA, Phinney KW, Tai SSC et al (2017) Baseline assessment of 25-hydroxy vitamin D assay performance: a vitamin D standardization program (VDSP) interlaboratory comparison study. J AOAC Int 100:1244–1252
Tai SSC, Nelson MA, Bedner M et al (2017) Development of standard reference material (SRM) 2973 vitamin D metabolites in frozen human serum (High Level). J AOAC Int 100:1294–1303. https://doi.org/10.5740/JAOACINT.17-0182
Phinney KW, Tai SSC, Bedner M et al (2017) Development of an improved standard reference material for vitamin D metabolites in human serum. Anal Chem 89:4907–4913. https://doi.org/10.1021/acs.analchem.6b05168
Tai SSC, Nelson MA (2015) Candidate reference measurement procedure for the determination of (24R),25-dihydroxyvitamin D3 in human serum using isotope-dilution liquid chromatography-tandem mass spectrometry. Anal Chem 87:7964–7970. https://doi.org/10.1021/ACS.ANALCHEM.5B01861
Carter GD, Ahmed F, Berry J et al (2019) External quality assessment of 24,25-dihydroxyvitamin D 3 (24,25(OH) 2 D 3) assays. J Steroid Biochem Mol Biol 187:130–133. https://doi.org/10.1016/j.jsbmb.2018.11.010
Dirks NF, Ackermans MT, De Jonge R, Heijboer AC (2019) Reference values for 24,25-dihydroxyvitamin D and the 25-hydroxyvitamin D/24,25-dihydroxyvitamin D ratio. Clin Chem Lab Med. https://doi.org/10.1515/cclm-2018-1096
Tang JCY, Nicholls H, Piec I et al (2017) Reference intervals for serum 24,25-dihydroxyvitamin D and the ratio with 25-hydroxyvitamin D established using a newly developed LC–MS/MS method. J Nutr Biochem 46:21–29. https://doi.org/10.1016/j.jnutbio.2017.04.005
Ketha H, Kumar R, Singh RJ (2016) LC-MS/MS for identifying patients with CYP24A1 mutations. Clin Chem 62:236–242. https://doi.org/10.1373/clinchem.2015.244459
Cooke NE, McLeod JF, Wang X, Ray K (1991) Vitamin D binding protein: genomic structure, functional domains, and mRNA expression in tissues. J Steroid Biochem Mol Biol 40:787–793. https://doi.org/10.1016/0960-0760(91)90304-N
Nykjaer A, Dragun D, Walther D et al (1999) An endocytic pathway essential for renal uptake and activation of the steroid 25-(OH) vitamin D3. Cell 96:507–515. https://doi.org/10.1016/S0092-8674(00)80655-8
Henderson CM, Fink SL, Bassyouni H et al (2019) Vitamin D-binding protein deficiency and homozygous deletion of the GC gene. N Engl J Med 380:1150–1157. https://doi.org/10.1056/nejmoa1807841
Safadi FF, Thornton P, Magiera H et al (1999) Osteopathy and resistance to vitamin D toxicity in mice null for vitamin D binding protein. J Clin Invest 103:239–251. https://doi.org/10.1172/JCI5244
Bikle DD (2021) The free hormone hypothesis: when, why, and how to measure the free hormone levels to assess vitamin D, thyroid, sex hormone, and cortisol status. JBMR Plus. https://doi.org/10.1002/jbm4.10418
Lefevre ML, Siu AL, Bibbins-Domingo K et al (2015) Screening for vitamin D deficiency in adults: U.S. Preventive services task force recommendation statement. Ann Intern Med 162:133–140. https://doi.org/10.7326/M14-2450
Bikle DD, Halloran BP, Gee E et al (1986) Free 25-hydroxyvitamin D levels are normal in subjects with liver disease and reduced total 25-hydroxyvitamin D levels. J Clin Invest 78:748–752. https://doi.org/10.1172/JCI112636
Schwartz JB, Christopher Gallagher J, Jorde R et al (2018) Determination of Free 25(OH)D concentrations and their relationships to total 25(OH)D in multiple clinical populations. J Clin Endocrinol Metab 103:3278–3288. https://doi.org/10.1210/jc.2018-00295
Martin CB, Oshiro BT, Sands LAD et al (2020) Vitamin-D dysregulation in early- and late-onset preeclampsia: a gestational-age matched study. J Steroid Biochem Mol Biol. https://doi.org/10.1016/J.JSBMB.2020.105729
Lundgren S, Carling T, Hjälm G et al (1997) Tissue distribution of human gp330/megalin, a putative Ca(2+)-sensing protein. J Histochem Cytochem 45:383–392. https://doi.org/10.1177/002215549704500306
Zheng G, Bachinsky DR, Stamenkovic I et al (1994) Organ distribution in rats of two members of the low-density lipoprotein receptor gene family, gp330 and LRP/alpha 2MR, and the receptor-associated protein (RAP). J Histochem Cytochem 42:531–542. https://doi.org/10.1177/42.4.7510321
Bhan I, Powe CE, Berg AH et al (2012) Bioavailable vitamin D is more tightly linked to mineral metabolism than total vitamin D in incident hemodialysis patients. Kidney Int 82:84–89. https://doi.org/10.1038/ki.2012.19
Rivera-Paredez B, Hidalgo-Bravo A, León-Reyes G et al (2021) Total, bioavailable, and free 25-hydroxyvitamin D equally associate with adiposity markers and metabolic traits in mexican adults. Nutrients. https://doi.org/10.3390/NU13103320/S1
Heo YJ, Lee YJ, Lee K et al (2021) Total, bioavailable and free 25-hydroxyvitamin D levels as functional indicators for bone parameters in healthy children. PLoS ONE 16:e0258585. https://doi.org/10.1371/JOURNAL.PONE.0258585
Rockwell MS, Kostelnik SB, McMillan RP, Lancaster M, Enette Larson-Meyer D, Hulver MW (2021) An association between bioavailable 25-hydroxyvitamin D and bone mineral density in a diverse cohort of collegiate athletes. Med Sci Sports Exerc. https://doi.org/10.1249/MSS.0000000000002807
Bikle DD, Gee E, Halloran B et al (1986) Assessment of the free fraction of 25-hydroxyvitamin d in serum and its regulation by albumin and the vitamin d-binding protein. J Clin Endocrinol Metab 63:954–959. https://doi.org/10.1210/jcem-63-4-954
Arnaud J, Constans J (1993) Affinity differences for vitamin D metabolites associated with the genetic isoforms of the human serum carrier protein (DBP). Hum Genet 92:183–188. https://doi.org/10.1007/BF00219689
Lauridsen AL, Vestergaard P, Nexo E (2001) Mean serum concentration of vitamin D-binding protein (Gc globulin) is related to the Gc phenotype in women. Clin Chem 47:753–756. https://doi.org/10.1093/clinchem/47.4.753
Fu L, Yun F, Oczak M et al (2009) Common genetic variants of the vitamin D binding protein (DBP) predict differences in response of serum 25-hydroxyvitamin D [25(OH)D] to vitamin D supplementation. Clin Biochem 42:1174–1177. https://doi.org/10.1016/J.CLINBIOCHEM.2009.03.008
Takiar R, Lutsey PL, Zhao D et al (2015) The associations of 25-hydroxyvitamin D levels, vitamin D binding protein gene polymorphisms, and race with risk of incident fracture-related hospitalization: twenty-year follow-up in a bi-ethnic cohort (the ARIC Study). Bone 78:94–101. https://doi.org/10.1016/J.BONE.2015.04.029
Malik S, Fu L, Juras DJ et al (2013) Common variants of the vitamin D binding protein gene and adverse health outcomes. Crit Rev Clin Lab Sci 50:1–22. https://doi.org/10.3109/10408363.2012.750262
Speeckaert M, Huang G, Delanghe JR, Taes YEC (2006) Biological and clinical aspects of the vitamin D binding protein (Gc-globulin) and its polymorphism. Clin Chim Acta 372:33–42. https://doi.org/10.1016/J.CCA.2006.03.011
Makris K, Bhattoa HP, Cavalier E et al (2021) Recommendations on the measurement and the clinical use of vitamin D metabolites and vitamin D binding protein—a position paper from the IFCC Committee on bone metabolism. Clin Chim Acta 517:171–197. https://doi.org/10.1016/j.cca.2021.03.002
Denburg MR, Hoofnagle AN, Sayed S et al (2016) Comparison of two ELISA methods and mass spectrometry for measurement of vitamin D-binding protein: implications for the assessment of bioavailable vitamin d concentrations across genotypes. J Bone Miner Res 31:1128–1136. https://doi.org/10.1002/jbmr.2829
Hollis BW, Bikle DD (2014) Vitamin D-binding protein and vitamin D in blacks and whites. N Engl J Med 370:878–881. https://doi.org/10.1056/nejmc1315850
Nielson CM, Jones KS, Chun RF et al (2016) Role of assay type in determining free 25-hydroxyvitamin D levels in diverse populations. N Engl J Med 374:1695–1696. https://doi.org/10.1056/nejmc1513502
Bikle DD, Siiteri PK, Ryzen E et al (1985) Serum protein binding of 1, 25-Dihydroxyvitamin D: a reevaluation by direct measurement of free metabolite levels. J Clin Endocrinol Metab 61:969–975. https://doi.org/10.1210/jcem-61-5-969
Heureux N, Lindhout E, Swinkels L (2017) A direct assay for measuring free 25-hydroxyvitamin D. J AOAC Int 100:1318–1322
Peris P, Filella X, Monegal A et al (2017) Concordance between direct and indirect measurements of free 25-OH vitamin D. Clin Chim Acta 475:169–171. https://doi.org/10.1016/j.cca.2017.10.016
Lopez-Molina M, Santillan C, Murillo M et al (2018) Measured free 25-hydroxyvitamin D in healthy children and relationship to total 25-hydroxyvitamin D, calculated free 25-hydroxyvitamin D and vitamin D binding protein. Clin Biochem 61:23–27. https://doi.org/10.1016/J.CLINBIOCHEM.2018.08.007
Jukic AMZ, Hoofnagle AN, Lutsey PL (2018) Measurement of vitamin D for Epidemiologic and clinical research: shining light on a complex decision. Am J Epidemiol 187:879–890
Kennel KA, Drake MT, Hurley DL (2010) Vitamin D deficiency in adults: when to test and how to treat. Mayo Clin Proc 85:752–758
Heijboer AC, Blankenstein MA, Kema IP, Buijs MM (2012) Accuracy of 6 routine 25-hydroxyvitamin D assays: influence of vitamin D binding protein concentration. Clin Chem 58:543–548. https://doi.org/10.1373/clinchem.2011.176545
Kim HJ, Ji M, Song J et al (2017) Clinical utility of measurement of vitamin D-binding protein and calculation of bioavailable vitamin D in assessment of vitamin D status. Ann Lab Med 37:34–38. https://doi.org/10.3343/alm.2017.37.1.34
Jeng L, Yamshchikov AV, Judd SE et al (2009) Alterations in vitamin D status and anti-microbial peptide levels in patients in the intensive care unit with sepsis. J Transl Med 7:1–9. https://doi.org/10.1186/1479-5876-7-28
Dahl B, Schiødt FV, Ott P et al (2003) Plasma concentration of Gc-globulin is associated with organ dysfunction and sepsis after injury. Crit Care Med 31:152–156. https://doi.org/10.1097/00003246-200301000-00024
van Hoof HJC, de Sévaux RGL, van Baelen H et al (2001) Relationship between free and total 1,25-dihydroxyvitamin D in conditions of modified binding. Eur J Endocrinol 144:391–396. https://doi.org/10.1530/eje.0.1440391
Funding
Open access funding provided by Medical University of Graz. The authors did not receive support from any organisation for the submitted work.
Author information
Authors and Affiliations
Contributions
MH conceived the idea; all authors performed the literature search, wrote the manuscript and critically review it.
Corresponding author
Ethics declarations
Conflict of interest
There is no conflict of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Alonso, N., Zelzer, S., Eibinger, G. et al. Vitamin D Metabolites: Analytical Challenges and Clinical Relevance. Calcif Tissue Int 112, 158–177 (2023). https://doi.org/10.1007/s00223-022-00961-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00223-022-00961-5