
Abstract
Strains LEOWEIH-7CT and LEPPI-3A were isolated from the Leopoldskroner Weiher, a lake located in the city of Salzburg, Austria. 16S rRNA gene similarities and phylogenetic reconstructions with 16S rRNA gene sequences as well as based on genome sequences revealed that the new strains belong to the A. antheringensis branch of the genus Aquirufa. Calculated whole-genome average nucleotide identity (gANI) and digital DNA–DNA hybridization (dDDH) values with the closely related type strains showed that the two strains represent a single new species. The strains grew aerobically and chemoorganotrophically, and the cells were rod shaped, on average 0.8 µm long and 0.3 µm wide, red pigmented and motile by gliding. The genome size of both strains was 2.6 Mbp and the G+C value was 41.9%. The genomes comprised genes predicted for the complete light-harvesting rhodopsin system and various carotenoids. We proposed to establish the name Aquirufa regiilacus sp. nov. for strain LEOWEIH-7CT (=DSM 116390T = JCM 36347T) as the type strain. Strain LEPPI-3A (=DSM 116391 = JCM 36348) also belongs to this new species. The calculated genome-based phylogenetic tree revealed that Aquirufa and some other genera currently allocated in the family Cytophagaceae need a reclassification. Aquirufa, Arundinibacter, Sandaracinomonas, and Tellurirhabdus should be designated to the family Spirosomataceae, the genus Chryseotalea to the family Fulvivirgaceae, and the genus Litoribacter to the family Cyclobacteriaceae. Furthermore, based on calculated gANI and dDDH values, Litoribacter alkaliphilus should be reclassified as a later heterotypic synonym of Litoribacter ruber.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The genus Aquirufa was first described in 2019 (Pitt et al. 2019) as belonging to the family Cytophagaceae of the phylum Bacteroidota. Due to genome-based phylogenetic analyses, it was known that the family Cytophagaceae with its members showed polyphyly at that time (Hahnke et al. 2016). In 2019 it was proposed to split the family Cytophagaceae into three distinct families (García-López et al. 2019). Besides Cytophagaceae and Flexibacteraceae, the latter validated in 2020 (Oren and Garrity 2020), the authors recommended using the validly published name Spirosomaceae proposed in 1978 (Larkin and Borrall 1978) and orthographically corrected in 2022 to Spirosomataceae (Arahal et al. 2022). Some genera were not considered in the reorganization of the family Cytophagaceae, because they were described at the same time or near after. Our phylogenetic reconstructions based on genome sequences showed that the genus Aquirufa and some other genera, all currently allocated in the family Cytophagaceae, need a reclassification. Furthermore, the genomic tree and subsequently calculated whole-genome average nucleotide identity (gANI) values revealed that Litoribacter alkaliphilus is a later heterotypic synonym of Litoribacter ruber.
The family Spirosomataceae comprises Gram-negative, rod-forming, aerobic or facultatively anaerobic, non-spore-forming and pigmented bacteria with variable motility; the major menaquinone is menaquinone-7 and the major polar lipid is phosphatidylethanolamine (García-López et al. 2019).
The genus Aquirufa is characterized by aerobic, chemoorganotrophic, rod-shaped, red-pigmented freshwater bacteria with gliding motility (Pitt et al. 2019) including at the time of writing seven species with validly published names (Pitt et al. 2019, 2020, 2022; Sheu et al. 2020). These bacteria seem to occur all over the world in various standing and running freshwater habitats (Pitt et al. 2022). Closely related to the genus Aquirufa is the genus Sandaracinomonas with the only species Sandaracinomonas limnophila, which was also isolated from a freshwater habitat (Chen et al. 2020). Genome-based phylogenetic trees showed that the members of the genus Aquirufa form two lineages (Pitt et al. 2022). Species of the A. antheringensis branch (A. antheringensis, A. lenticrescens) are characterized by genome sizes of around 2.5 Mbp and G+C contents of around 42%. The A. nivalisilvae branch (A. nivalisilvae, A. aurantiipilula, A. ecclesiirivi, A. beregesia, A. rosea) consists of species with genome sizes around 3 Mbp and G+C contents around 38%.
Within a citizen science project, we searched for new Aquirufa strains in freshwater habitats in Salzburg and Upper Austria. The habitats were sampled and bacterial cultures belonging to the genus were obtained. Two strains, LEOWEIH-7CT and LEPPI-3A, both isolated from water samples of a lake located in the city of Salzburg, represented a new species of the A. antheringensis branch.
So, we describe here the new species Aquirufa regiilacus sp. nov. with its type strain LEOWEIH-7CT. In addition, we propose the classification of Aquirufa and five other genera, which are currently placed in the family Cytophagaceae, to appropriate families and the reclassification of Litoribacter alkaliphilus.
Materials and methods
Home habitat and isolation
Strains LEOWEIH-7CT and LEPPI-3A originated from an artificial lake called Leopoldskroner Weiher in the city of Salzburg. This lake with an area of 13 ha was created more than 500 years ago, probably as a retention reservoir. Later the castle Leopoldskron was built near the shore, which is today together with the pond and the surrounding park under a preservation order. Strains LEOWEIH-7CT and LEPPI-3A were obtained from different water samples, both were taken from the surface water in November 2022 at the geographic coordinates 47.78326 N 13.04111 E and 47.78445 N 13.03846 E, respectively. Measurements revealed a neutral pH value for both samples and a conductivity of 399 µS cm−1 and 274 µS cm−1, respectively.
The isolation procedure started with filtering the water samples through 0.65 µm pore-size membrane filters and spreading the filtrate afterward on nutrient broth soytone yeast extract (NSY) agar plates (Hahn et al. 2004). We searched for colonies with red pigmentation, which is a characteristic of Aquirufa strains. They were picked and transferred into liquid NSY medium. Sanger sequencing of the marker gene gyrB (B subunit of the DNA gyrase) (Pitt et al. 2022) was used to search for strains representing new species. The candidate strains were obtained as pure cultures by alternating cultivation on NSY agar plates and in liquid NSY medium and subsequently stored in NSY medium with 15% glycerol at −80 °C. The strains were genome-sequenced and gANI values were calculated to verify the preliminary estimations.
Phenotypic and chemotaxonomic characterization
For the phenotypic and chemotaxonomic investigations, the same methods were used as described previously (Pitt et al. 2022). To reveal the temperature range of growth occulated NSY agar plates were exposed to increasing temperatures. We started at 5 °C and increased the temperature until no growth was observed. Anaerobic growth was tested in an anaerobic chamber by the use of standard NSY agar plates, and NSY plates supplemented with 2 g l−1 NaNO3, for testing NaCl tolerance served agar plates with various NaCl concentrations (0.1% w/v steps). Cell dimensions were measured with an epifluorescence microscope (UV filter). For that purpose, liquid cultures were fixed with 2% paraformaldehyde and stained with 4′,6-diamidino-2-phenylindole (DAPI). Motility of the strains was tested on soft agar plates (1 g l−1 yeast extract, 0.1 g l−1 K2HPO4, and 2.0 g l−1 agar). 20 µl of a well-growing culture was placed in the middle of standard NSY plates and on soft agar plates, incubated at 21 °C, and observed for one week. For chemotaxonomic characterization, the composition of cellular fatty acids, polar lipids, and respiratory quinones were analyzed. For this purpose, cells were cultured in liquid NSY medium at room temperature and harvested after three days by centrifugation. To determine the cellular fatty acid composition, biomass was saponified, methylated, and subsequently analyzed on an Agilent Technologies 6890 N instrument coupled to a flame ionization detector following the protocol described by Sasser (Sasser 1990). Equivalent chain length values were calculated analogously to the Microbial Identification System Sherlock to provide the peak naming according to the TSBA6 database. The extract was analyzed in addition by gas chromatography/mass spectrometry (GC/MS) to identify the fatty acids (Vieira et al. 2021). Double bond positions were determined by further derivatization to dimethyl disulfide adducts and following GC/MS analysis (Moss and Lambert-Fair 1989). The polar lipids were extracted and analyzed following the description of Tindall (Tindall 1990a, b) and the methods of Bligh and Dyer (Bligh and Dyer 1959). Two-dimensional silica gel thin-layer chromatography served to separate the polar lipids. The total lipids were detected by the use of dodecamolybdophosphoric acid (Dmp) and specific functional groups by using α-naphthol, ninhydrin, and molybdenum blue. Respiratory quinones were extracted by solid-phase extraction for analyses served reversed-phase HPLC coupled to a diode array detector as well as a high-resolution mass spectrometer. This method was described previously (Vieira et al. 2021).
Genomic characterization
For DNA extraction and genome sequencing, the method described by Hoetzinger et al. (Hoetzinger et al. 2017) was used. For this purpose, a shotgun library was paired-end sequenced with 2 × 150 bp (Illumina NovaSeq). The software SPAdes version 3.13.1 (Bankevich et al. 2012) was used for de novo genome assembly. The nucleotide coverage was calculated based on the k-mer coverage received from SPAdes by using the formula: nucleotide coverage = k-mer coverage * read length/(read length−k-mer length + 1). The software CheckM (Parks et al. 2015) provided by the online platform Galaxy Protologger (Hitch et al. 2021) was utilized for a quality check of the obtained genome sequences. Both genomes were annotated by the NCBI Prokaryotic Genome Annotation Pipeline (Tatusova et al. 2016) and the RAST annotation server (Aziz et al. 2008) and deposited at DDBJ/ENA/GenBank databases. They were also incorporated into the Integrated Microbial Genomes and Microbiomes Expert Review (IMG/MER) database (Chen et al. 2019) for further annotation. The IMG/MER tool Phylogenetic Profiler for Single Genes served for detection of the presence or absence of homologous protein-encoding genes in the regarded genomes. The settings were minimum 30% identity and max. E-value 1e−5. These data formed the basis of a Venn diagram, which was constructed with the online tool https://eulerr.co (Larsson 2021). The tool SEED viewer (Overbeek et al. 2005) was used for determining N50 and L50 values and an amino acid sequence-based comparison of the genomes of strains LEOWEIH-7CT and LEPPI-3A with the type strains of A. lenticrescens and A. antheringensis. The gANI and corresponding alignment fraction (AF) values were calculated with the IMG/MER system (Chen et al. 2019). Both parameters were obtained for all possible pairs involving all Aquirufa type strains. Additionally, the Type (Strain) Genome Server (Meier-Kolthoff et al. 2021) served for the determination of digital DNA–DNA hybridization (dDDH) values. The online tool Average Amino Acid Identity calculator from the Environmental Microbial Genomics Laboratory (Rodríguez-R and Konstantinidis 2016) served for the calculation of whole-genome (proteome) average amino acid identity (gAAI) values.
Phylogenetic reconstructions
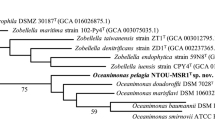
Phylogenetic reconstructions were performed by using almost full-length sequences of the 16S rRNA gene as well as by utilizing amino acid sequences of 119 single-copy marker genes (Parks et al. 2018) based on genome sequences. For the phylogenetic tree based on 16S rRNA gene sequences (Figure S1), at least all type species of the genera listed by the LPSN database (Meier-Kolthoff et al. 2021) for the families Cytophagaceae, Spirosomataceae, Flexibacteraceae and Fulvivirgaceae, as well as a selection of the type species of the family Cyclobacteriaceae, were included. The software MEGA X (Kumar et al. 2018) was used to align the sequences, and a neighbor-joining tree was constructed with the parameters Kimura 2 model (Kimura 1980), invariant sites, gamma-distributed (5 categories), and 1000 bootstrap replicates. All strains from Figure S1 with available genome sequences were considered for the phylogenetic tree based on genome sequences (Fig. 1). A RAxMl tree was calculated with amino acid sequences of 119 protein-encoding genes (Parks et al. 2018). One of the 120 genes recommended by Parks et al., i.e., the protein family TIGR0009, was not found in all considered genomes and therefore omitted. MAFFT version 7 (Katoh et al. 2005) served to align the concatenate amino acid sequences. GBLOCKS version 0.91b (Castresana 2000) was used to filter out highly variable positions of the alignment. The result was a reduction of the alignment fraction from 50665 to 36179 amino acid positions (in 485 selected blocks). This represented 71% of the original alignment positions. The CIPRES Science Gateway version 3.3 (Miller et al. 2010) served to construct a RAxML tree (Stamatakis 2014) with 100 bootstrap replicates.

Midpoint rooted phylogenomic RAxML tree calculated with amino acid sequences obtained from 119 single-copy genes from all available genomes of the taxa from Figure S1. The classification of the genera into families (colored letters) is based on their position in the tree. Bar, 0.1 substitutions per nucleotide position (colour figure online)
Results and discussion
Phenotypic and chemotaxonomic analyses
The phenotypic features of the type strain are shown in Table 1. The results for strain LEPPI-3A were very similar and are therefore not listed. The fatty acid composition of strain LEOWEIH-7CT is presented in Table S1, the patterns of the polar lipids in Figure S2. When growing exclusively in a liquid medium for several weeks, cultures of strain LEOWEIH-7CT became less colored. After spreading these paler cultures on agar plates, white colonies occurred among the normal red ones and could be purified. It seemed that this phenomenon was caused by a mutation.
Table 1 indicates the features which distinguished the new type strain from the closely related type strains of the A. antheringensis branch. The cell size of the new strain was smaller than the cell dimensions of the related strains, as well as the patterns of the fatty acids and the polar lipids differed in some details.
Phylogenetic analyses
BLAST analyses (Johnson et al. 2008) of strains LEOWEIH-7CT and LEPPI-3A revealed 16S rRNA gene sequence similarities with the type strains of A. antheringensis and A. lenticrescens of 99.42% and 99.47%, respectively. It is known that Aquirufa species within the A. antheringensis branch and the A. nivalisilvae branch, respectively, have identical or nearly identical 16S rRNA gene sequences (Pitt et al. 2022). The phylogenetic reconstruction based on 16S rRNA gene sequences (Figure S1) could neither resolve the relationships between the Aquirufa species nor was suitable for assignment of the incorporated genera to a family. Nevertheless, the tree revealed the affiliation of the two new strains with A. antheringensis and A. lenticrescens. The whole-genome-based phylogenetic reconstructions (Fig. 1) confirmed an affiliation and pointed out a relatively far phylogenetic distance between the new strains and the type strains of the two above-mentioned species.
In addition, the genome-based tree (Fig. 1) showed that the species of the genera Aquirufa, Arundinibacter, Sandaracinomonas, and Tellurirhabdus were placed on the branch of the family Spirosomataceae, the species of the genus Litoribacter on the branch of the family Cyclobacteriaceae, and the species of the genus Chryseotalea on the branch of the family Fulvivirgaceae. The branch of the genus Rhodocytophaga was strongly separated from the mentioned families. Recently, the new family ‘Rhodocytophagaceae’ including Rhodocytophaga and a newly described genus ‘Xanthocytophaga’ was proposed (Zhang et al. 2023) but so far not validated. The two species of the genus Litoribacter, L. ruber and L. alkaliphilus, possessed very short branches.
Genomic analyses
The results of genome sequencing were in the case of strain LEOWEIH-7CT 12 contigs with a coverage value of 598x, N50 289 kbp, L50 2 and in the case of strain LEPPI-3A 40 contigs with a coverage of 470x, N50 291 kbp, L50 4, and the genome size of both genomes was 2.6 Mbp. The authenticity of the genomes was tested with Sanger sequences obtained from sequencing the 16S rRNA gene and the gyrB gene. Quality checks with CheckM revealed for both strains genome completeness of 96.3% and detected no genome contamination, and the genome sequence similarity of the two genomes was 99.1%. A comparison of the genome sequences of the novel strains and the type strains of nearly related species is given in Fig. 2. Regarding the homologous protein-coding genes (Fig. 2, left), approximately 80% of the genes were found in all three species. A little more than 400 genes were found exclusively in the new species (blue area). Even if considering only one of the two new strains to ensure equality, the new species had twice as many exclusive genes as the type strains of the nearest related species. This corresponded with the genome-based phylogenetic tree (Fig. 1), which revealed a wide distance between the branch of the new species and the nearest related Aquirufa species. Comparisons of the protein sequences (Fig. 2, right) consolidate this impression. The amino acid sequence similarities of the proteins in comparison of strain LEOWEIH-7CT and the type strain of the closely related species were relatively low and corresponded with the average value (gAAI) of around 69%.

Genome sequence-based comparison of the two new strains (LEOWEIH-7CT and LEPPI-3A) and the type strains of the closely related species (A. antheringensis, 30S-ANTBACT and A. lenticrescens, 9H-EGSET). Left: Venn diagram of the protein-coding genes. Right: Protein sequence identities of strain LEOWEIH-7CT compared with strain LEPPI-3A (outer circle), with strain 9H-EGSET (middle circle) and strain 30S-ANTBACT (inner circle), upper scale: bidirectional best hits, lower scale: unidirectional best hits
The pairwise calculated gANI values of the strains LEOWEIH-7CT and LEPPI-3A and the type strains of the nearest related Aquirufa species (Table 2) were 77.3% and 78.0%, respectively, with relatively low AF values of around 50%. They ranged clearly under the accepted threshold of 95–96% utilized to delineate two prokaryotic species (Konstantinidis et al. 2006; Chun et al. 2018). The calculated dDDH values varied between 20.1 and 21.1% and were also far under the established threshold of 70% (Chun et al. 2018). On the other hand, the calculated values of 99.1% and 92.3%, respectively (Table 2), showed that strain LEOWEIH-7CT and LEPPI-3A belong to the same species. The calculated gANI as well as dDDH values of the type strains of L. alkaliphilus and L. ruber of 98.4% and 85.8%, respectively, indicated clearly that they should belong to the same species. The differentiation of L. alkaliphilus from L. ruber as a new species was based on a lab DNA–DNA relatedness value of 23 ± 1% (Subhash et al. 2013), which is far below the accepted threshold. So, it is unclear what caused the wrong estimation.
Like all so far described Aquirufa strains, the new strains contained genes predicted for the whole glycolysis, citrate cycle, and pentose phosphate pathway as well as the biosynthesis of the carotenoids zeta-carotene, lycopene, beta-carotene, and astaxanthin. Some genes which were detected in all or some of the strains of the A. antheringensis branch are listed in Table 2. As all strains belonging to this branch, the new strains owned genes putatively encoding for the entire light-harvesting bacteriorhodopsin/retinal system. Ten genes were predicted in the IMG/MER system for gliding motility and six genes for the Type IX secretion system membrane protein PorP/SprF, which is confined to the phylum Bacteroidota and could also be involved in gliding motility (McBride 2019).
Ecology of the new species
Two additional cultures, which were screened by sequencing the gyrB gene, were likely belonging to the proposed new species. They originated from the lake Hallstättersee and the river Vöckla, both located in Upper Austria (data not shown). Therefore, the new species seems to dwell in both standing and running freshwater systems. Since the 16S rRNA gene sequence similarities of the new species and the two closely related species (see above) are very high, BLAST searches with the 16S rRNA gene sequence are unsuitable for detecting the new species.
It is unclear if the species appears in the open water, free or attached to particles, or in the sediment. While the water samples from lake Leopoldskroner Weiher were taken from the lakeside, the sample from lake Hallstättersee originated from the water column, so a pelagic lifestyle is most probably. The capacity for gliding motility suggests at least a temporary association with particles. The genome size of 2.6 Mbp is quite small but in the upper range of genome sizes of typical pelagic freshwater bacteria (Chiriac et al. 2023), which is regarded as an adaption to a pelagic lifestyle. The same applies to the very high coding densities and the comparatively low G+C contents (Table 2) of the genomes of the new strains and the very small cell size. The occurrence of the light-harvesting rhodopsin system is generally discussed as an adaption of freshwater bacteria to low nutrient conditions (Chiriac et al. 2023), in any case, it is a hint on the occurrence of the new species in upper water layers, where enough light is available.
Conclusion
The calculated 16S rRNA gene identity values showed that strains LEOWEIH-7CT and LEPPI-3A belong to the genus Aquirufa. The calculated gANI, gAAI, and dDDH values indicated clearly that the strains represent a new species of the genus Aquirufa. The phylogenetic reconstructions based on genome sequences (Fig. 1) and the phenotypic and chemotaxonomic features of the type strain support this finding. For the new species with the type strain LEOWEIH-7CT, we propose the name Aquirufa regiilacus sp. nov.
In addition, based on the genomic phylogenetic tree of Fig. 1, we propose the reclassification of the genera Aquirufa, Arundinibacter, Sandaracinomonas, and Tellurirhabdus to the family Spirosomataceae, and the genus Chryseotalea to the family Fulvivirgaceae, as well as the genus Litoribacter to the family Cyclobacteriaceae. According to the calculated gANI and dDDH values, we propose to classify Litoribacter alkaliphilus as a later heterotypic synonym of Litoribacter ruber.
Description of Aquirufa regiilacus sp. nov. (re.gi.i.la’cus. L. masc. n. regius, royal; L. masc. n. lacus, lake; N.L. gen. n. regiilacus, of a royal lake)
Cells form rods, about 0.3 µm wide and 0.8 µm long. Colonies grown on NSY or R2A agar are circular, and convex with a smooth surface; they are bright red, in older stages dark red pigmented. Liquid cultures (NSY or R2A medium) have a red–orange coloring. Cells show motility on soft agar. Growth occurs in 0–0.3% (w/v) NaCl and at 5–32 °C. Major fatty acids are iso-C15:0, C16:1ω7c and anteiso-C15:0. Polar lipids are three unidentified polar lipids, one unidentified aminophospholipid, one unidentified aminolipid, and phosphatidylethanolamine. Menaquinone-7 is the major respiratory quinone. The genome of the type strain has a size of 2.6 Mbp and a G+C content of 41.9 mol%. Genes putatively encoding for the complete light-harvesting bacteriorhodopsin system and biosynthesis of several further carotenoids occur in the genome.
The type strain is LEOWEIH-7CT (=DSM 116390T = JCM 36347T), isolated from lake Leopoldskroner Weiher (City of Salzburg, Austria).
The accession numbers of the 16S rRNA gene sequence and the genome sequence are OR064354 and JAVNWW000000000, respectively.
Data availability
The Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession JAVNWW000000000 for strain LEOWEIH-7CT and JARDXH000000000 for strain LEPPI-3A. These are the versions described in this paper. The accession of the 16S rRNA gene sequence deposited at DDBJ/ENA/GenBank of strain LEOWEIH-7CT is OR064354. All genomes in Table 2 are available in the IMG/MER system (ID numbers see Table 2).
Abbreviations
- A.:
-
Aquirufa
- gANI:
-
Whole-genome average nucleotide identity
- dDDH:
-
Digital DNA–DNA hybridization
- NSY medium:
-
Nutrient broth soytone yeast extract medium
- gyrB:
-
B subunit of the DNA gyrase
- GC/MS:
-
Gas chromatography/mass spectrometry
- IMG/MER:
-
Integrated Microbial Genomes and Microbiomes Expert Review
- AF:
-
Alignment fraction
- gAAI:
-
Whole-genome average amino acid identity
- L. :
-
Litoribacter
References
Arahal DR, Busse H-J, Bull CT et al (2022) Judicial opinions 112–122. Int J Syst Evol Microbiol. https://doi.org/10.1099/ijsem.0.005481
Aziz RK, Bartels D, Best AA et al (2008) The RAST server: rapid annotations using subsystems technology. BMC Genomics 9:75. https://doi.org/10.1186/1471-2164-9-75
Bankevich A, Nurk S, Antipov D et al (2012) SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol 19:455–477. https://doi.org/10.1089/cmb.2012.0021
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem 37:911–917
Castresana J (2000) Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol 17:540–552. https://doi.org/10.1093/oxfordjournals.molbev.a026334
Chen IMA, Chu K, Palaniappan K et al (2019) IMG/M vol 5.0: an integrated data management and comparative analysis system for microbial genomes and microbiomes. Nucleic Acids Res 47:D666–D677. https://doi.org/10.1093/nar/gky901
Chen W-M, Cai C-Y, Sheu S-Y (2020) Sandaracinomonas limnophila gen. nov., sp. nov., a new member of the family Cytophagaceae isolated from a freshwater mesocosm. Int J Syst Evol Microbiol 70:2178–2185. https://doi.org/10.1099/ijsem.0.003532
Chiriac M-C, Haber M, Salcher MM (2023) Adaptive genetic traits in pelagic freshwater microbes. Environ Microbiol 25:606–641. https://doi.org/10.1111/1462-2920.16313
Chun J, Oren A, Ventosa A et al (2018) Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int J Syst Evol Microbiol 68:461–466. https://doi.org/10.1099/ijsem.0.002516
García-López M, Meier-Kolthoff JP, Tindall BJ et al (2019) Analysis of 1000 type-strain genomes improves taxonomic classification of Bacteroidetes. Front Microbiol 10:2083. https://doi.org/10.3389/fmicb.2019.02083
Hahn MW, Stadler P, Wu QL, Pöckl M (2004) The filtration-acclimatization method for isolation of an important fraction of the not readily cultivable bacteria. J Microbiol Methods 57:379–390. https://doi.org/10.1016/j.mimet.2004.02.004
Hahnke RL, Meier-Kolthoff JP, García-López M et al (2016) Genome-based taxonomic classification of Bacteroidetes. Front Microbiol 7:2003. https://doi.org/10.3389/fmicb.2016.02003
Hitch TCA, Riedel T, Oren A, Overmann J, Lawley TD, Clavel T (2021) Automated analysis of genomic sequences facilitates high-throughput and comprehensive description of bacteria. ISME Commun 1:16. https://doi.org/10.1038/s43705-021-00017-z
Hoetzinger M, Schmidt J, Jezberova J, Koll U, Hahn MW (2017) Microdiversification of a pelagic Polynucleobacter species is mainly driven by acquisition of genomic islands from a partially interspecific gene pool. Appl Environ Microbiol 83:19. https://doi.org/10.1128/aem.02266-16
Johnson M, Zaretskaya I, Raytselis Y, Merezhuk Y, McGinnis S, Madden TL (2008) NCBI BLAST: a better web interface. Nucleic Acids Res 36:W5–W9. https://doi.org/10.1093/nar/gkn201
Katoh K, Kuma K, Toh H, Miyata T (2005) MAFFT version 5: improvement in accuracy of multiple sequence alignment. Nucleic Acids Res 33:511–518. https://doi.org/10.1093/nar/gki198
Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide-sequences. J Mol Evol 16:111–120. https://doi.org/10.1007/BF01731581
Konstantinidis KT, Ramette A, Tiedje JM (2006) The bacterial species definition in the genomic era. Philos Trans R Soc B Biol Sci 361:1929–1940. https://doi.org/10.1098/rstb.2006.1920
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35:1547–1549. https://doi.org/10.1093/molbev/msy096
Larkin JM, Borrall R (1978) Spirosomaceae, a new family to contain the genera Spirosoma Migula 1894, Flectobacillus Larkin et al1977, and Runella Larkin and Williams1978. Int J Syst Evol Microbiol 28:595–596. https://doi.org/10.1099/00207713-28-4-595
Larsson J (2021) eulerr: area-proportional euler and venn diagrams with ellipses. R package version 6.1.1. https://CRAN.R-project.org/package=eulerr
McBride MJ (2019) Bacteroidetes gliding motility and the type IX secretion system. Microbiol Spectr. https://doi.org/10.1128/microbiolspec.psib-0002-2018
Meier-Kolthoff JP, Carbasse JS, Peinado-Olarte RL, Göker M (2021) TYGS and LPSN: a database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res 50:D801–D807. https://doi.org/10.1093/nar/gkab902
Miller MA, Pfeiffer W, Schwartz T (2010) Creating the CIPRES science gateway for inference of large phylogenetic trees. In: Proceedings of the gateway computing environments workshop (GCE). IEEE, New Orleans, LA, pp 1–8
Moss CW, Lambert-Fair MA (1989) Location of double bonds in monounsaturated fatty acids of Campylobacter cryaerophila with dimethyl disulfide derivatives and combined gas chromatography-mass spectrometry. J Clin Microbiol 27:1467–1470. https://doi.org/10.1128/jcm.27.7.1467-1470.1989
Oren A, Garrity GM (2020) List of new names and new combinations previously effectively, but not validly, published. Int J Syst Evol Microbiol 70:2960–2966. https://doi.org/10.1099/ijsem.0.004156
Overbeek R, Begley T, Butler RM et al (2005) The subsystems approach to genome annotation and its use in the project to annotate 1000 genomes. Nucleic Acids Res 33:5691–5702. https://doi.org/10.1093/nar/gki866
Parks DH, Imelfort M, Skennerton CT, Hugenholtz P, Tyson GW (2015) CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res 25:1043–1055. https://doi.org/10.1101/gr.186072.114
Parks DH, Chuvochina M, Waite DW et al (2018) A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat Biotechnol 36:996–1004. https://doi.org/10.1038/nbt.4229
Pitt A, Schmidt J, Koll U, Hahn MW (2019) Aquirufa antheringensis gen. nov., sp. nov. and Aquirufa nivalisilvae sp. nov., representing a new genus of widespread freshwater bacteria. Int J Syst Evol Microbiol 69:2739–2749. https://doi.org/10.1099/ijsem.0.003554
Pitt A, Koll U, Schmidt J, Hahn MW (2020) Aquirufa ecclesiirivi sp. nov. and Aquirufa beregesia sp. nov., isolated from a small creek and classification of Allopseudarcicella aquatilis as a later heterotypic synonym of Aquirufa nivalisilvae. Int J Syst Evol Microbiol 70:4602–4609. https://doi.org/10.1099/ijsem.0.004319
Pitt A, Koll U, Schmidt J et al (2022) Aquirufa lenticrescens sp. nov. and Aquirufa aurantiipilula sp. nov.: two new species of a lineage of widespread freshwater bacteria. Arch Microbiol 204:356. https://doi.org/10.1007/s00203-022-02950-6
Rodríguez-R L, Konstantinidis K (2016) The enveomics collection: a toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Preprints 4:e1900v1
Sasser M (1990) Identification of bacteria by gas chromatography of cellular fatty acids. USFCC Newsl 20:16
Sheu S-Y, Chen T-Y, Chen W-M (2020) Aquirufa rosea sp. nov., isolated from a freshwater lake. Int J Syst Evol Microbiol 70:3145–3153. https://doi.org/10.1099/ijsem.0.004147
Stamatakis A (2014) RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30:1312–1313. https://doi.org/10.1093/bioinformatics/btu033
Subhash Y, Tushar L, Sasikala C, Ramana CV (2013) Mongoliicoccus alkaliphilus sp. nov. and Litoribacter alkaliphilus sp. nov., isolated from salt pans. Int J Syst Evol Microbiol 63:3457–3462. https://doi.org/10.1099/ijs.0.049924-0
Tatusova T, DiCuccio M, Badretdin A et al (2016) NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res 44:6614–6624. https://doi.org/10.1093/nar/gkw569
Tindall BJ (1990a) A comparative study of the lipid composition of Halobacterium saccharovorum from various sources. Syst Appl Microbiol 13:128–130. https://doi.org/10.1016/S0723-2020(11)80158-X
Tindall BJ (1990b) Lipid composition of Halobacterium lacusprofundi. FEMS Microbiol Lett 66:199–202
Vieira S, Huber KJ, Neumann-Schaal M et al (2021) Usitatibacter rugosus gen. nov., sp. nov. and Usitatibacter palustris sp. nov., novel members of Usitatibacteraceae fam. nov. within the order Nitrosomonadales isolated from soil. Int J Syst Evol Microbiol. https://doi.org/10.1099/ijsem.0.004631
Zhang X, Feng G-D, Liu Y et al (2023) Description of Xanthocytophaga agilis sp. nov. and Xanthocytophaga flavus sp. nov. of the new genus Xanthocytophaga gen. nov. and the proposal of Rhodocytophagaceae fam. nov. within the order Cytophagales. Arch Microbiol 205:345. https://doi.org/10.1007/s00203-023-03685-8
Acknowledgements
The authors thank all Citizen Scientists who contributed to this work. Many thanks to the students of the school BORG Nonntal with their teacher Markus Herbst, and especially Edna Jahja and Ioanna Polo for taking and working on the water samples from the Leopoldskroner Weiher. Anna Stephens, Lucie Metznerová, Zoe Huber, Martin Windberger, as well as Joachim Göbl (BRG Schloss Wagrain) for their assistance in laboratory work. The authors thank Isabella Kammerer (BG Vöcklabruck) and Florian Wolff (GUE Austria) for further water samples. They paid special thanks to Edna Jahja for creating the species name and to Aharon Oren for his kind nomenclatorial advice. They are also grateful to Anika Wasner, Birgit Grün, and Gesa Martens for technical assistance in chemotaxonomic analyses.
Funding
Open access funding provided by University of Innsbruck and Medical University of Innsbruck. This study was supported by the Austrian Federal Ministry of Education, Science and Research with the program Sparkling Science, project SPSC_01_051-Aquirufa and by the Austrian Research Promotion Agency with the program Talents.
Author information
Authors and Affiliations
Contributions
All authors contributed to the investigations and data analyses. AP created the figures and tables and wrote the manuscript. All authors provided comments on the manuscript.
Corresponding author
Ethics declarations
Conflict of interests
The authors have no relevant financial or non-financial interests to disclose. The authors declare no competing interests.
Additional information
Communicated by Yusuf Akhter.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pitt, A., Lienbacher, S., Schmidt, J. et al. Description of a new freshwater bacterium Aquirufa regiilacus sp. nov., classification of the genera Aquirufa, Arundinibacter, Sandaracinomonas, and Tellurirhabdus to the family Spirosomataceae, classification of the genus Chryseotalea to the family Fulvivirgaceae and Litoribacter to the family Cyclobacteriaceae, as well as classification of Litoribacter alkaliphilus as a later heterotypic synonym of Litoribacter ruber. Arch Microbiol 206, 79 (2024). https://doi.org/10.1007/s00203-023-03801-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00203-023-03801-8