
Abstract
Fungal deterioration is one of the major factors that significantly contribute to mummy cartonnage damage. Isolation and molecular identification of thirteen fungal species contributing to the deterioration of ancient Egyptian mummy cartonnage located in El-Lahun regions, Fayoum government, Egypt was performed. The most dominant deteriorated fungal species are Aspergillus flavus (25.70%), Aspergillus terreus (16.76%), followed by A. niger (13.97%). A newly synthesized series of tetrahydro-[1,2,4]triazolo[3,4-a]isoquinoline chalcone derivatives were synthesized and evaluated for their antifungal activities in vitro against the isolated deteriorated fungal species (Aspergillus flavus, A. niger, A. terreus, Athelia bombacina, Aureobasidium iranianum, Byssochlamys spectabilis, Cladosporium cladosporioides, C. ramotenellum, Penicillium crustosum, P. polonicum, Talaromyces atroroseus, T. minioluteus and T. purpureogenus). The most efficient chalcone derivatives are new chalcone derivative numbers 9 with minimum inhibitory concentration (MIC) ranging from 1 to 3 mg/mL followed by chalcone derivatives number 5 with MIC ranging from 1 to 4 mg/mL.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The scientific study of ancient human mummified skeletal remains has gained much attention over the past years with the improvement and progression of laboratory techniques (DeAraujo et al. 2016). The cartonnage in ancient Egypt appeared as an alternative from the deceased features through which the spirit can reach the body again if the mummy was damaged (Ali et al. 2016). Several factors lead to the deterioration of mummies including environmental conditions, physical damage, biological damage (Abdel-Maksoud and El-Amin 2013). Microorganisms (bacteria, actinomycetes, and fungi) cause the most serious damage to mummies (Naji et al. 2014). Ancient human remains and mummies were damaged by microbial species in processes that liquefying, degrading, or mineralizing different materials causes corrosion, fouling, rotting, declining, and disfiguring (DeAraujo et al. 2016).
Many fungal and bacterial species require available moisture for their development on the object surface (Valentin 2001). A temperature range for the growth of microorganisms is 30 °C (Valentin 2002), which controls the reaction, particularly collagen, which is the main component of mummy skin (Maekawa 1998), High temperatures can prevent the microbial growth but can cause the mummified skin to stiffen and become more susceptible to cracking and chemical breakdown. Increasing the relative humidity (RH) of museum air was believed to be the main factor responsible for microbial growth (Zaitseva 2010). A high relative humidity (RH) that arrives at 65℅ or higher aids in the decomposition of mummies (Valentin 2002). pH value aid in the growth of fungi and bacteria, whereas the fungi prefer acidic environments pH 6 is suitable for growth (Abdel-Maksoud and El-Amin 2013).
The deteriorating fungi can colonize books, clothing, manuscripts, mummies, and paintings causing cracking, crushing, discoloration, spots, and loss of material strength by enzymatic degradation, the production of metabolites, and mechanical attacks (Mansour et al. 2020). The fungal metabolites that accelerate the deterioration of the different archaeological substrata are carbon dioxide, water, organic acids (acetic, citric, oxalic acids, and others), and inorganic acids, specific pigments (melanin black pigments, and other colored pigments), volatile organic compounds (alcohols, aldehydes, and ketones), nitrogen-containing compounds (amino-acids, and beta-glucans), lipids and polyols (glycerol, and others), enzymes (cellulase, protease, amylase, and gelatinase), and secondary metabolites (mycotoxins and antibiotics) (Farooq et al. 2002; Borrego and Perdomo 2012; Lavin et al. 2016; Kakakhel et al. 2019).
Xerophilic and halophilic fungi share in mummy degradation due to xerophiles can grow on dry meat, and halophilic fungi can also tolerate high salinity. Xerophiles fungi (Cladosporium cladosporioides, Penicillium chrysogenum, and Aspergillus flavus) can produce mycotoxin that may also affect human health by breathing the high concentration of aflatoxins and fungal spore when opening the burial chamber after thousand years of being closed (Mansour 2018). Xerophilic fungi including Aspergillus penicillioides, Eurotium repens, E. rubrum, Penicillium roqueforti, and Wallemia sebi grow faster under acidic conditions (Gock et al. 2003). During the post-excavation storage and repair phases, waterlogged archaeological objects is at a significant risk of biological destruction (Geweely 2022).
Fungal species require necessary nutrients (nitrogen, vitamins, and minerals) for growth, which can obtain it from the meat of mummies (a major source of fats, protein, and vitamins) (Jay et al. 2008; Baltic and Boskovic 2015).
Exposure to fungal spores, metabolites, mycotoxins, and result in critical negative health impacts on the skin and respiratory system of library workers, conservators, and visitors (Sterflinger and Pinzari 2012). So, it is crucial to inhibit microbial proliferation to preserve archaeological objects and human health. Several physical and chemical strategies have been employed for heritage conservation including ultraviolet, laser, gamma rays, and traditional chemical compounds. These methods have various limitations including temporary actions, high cost, and toxic effects (Michaelsen et al. 2013; Sequeira et al. 2017). Various mechanical methods are incapable of preventing the growth of microorganisms completely. Many microorganisms have become resistant to the applied conventional antifungal agents (Kakakhel et al. 2019). To overcome these limits, the development of new and more efficient antimicrobial agents is required for application in materials preservation (Campana et al. 2020).
There is an urgent need to develop an alternative green eco-friendly strategy achieving high efficiency, cost-effectiveness, long-term effect, and safety for the preservation of historical artifacts against microbial colonization (Fouda et al. 2019). The isolation and screening of natural products are an important part of biocide discovery, where several successful drugs and molecules have been discovered via this route (Menezes and Diederich 2019). Novel inhibition of some pathogenic fungal and bacterial species by new synthetic phytochemical coumarin derivatives (Geweely 2009a). The development of new and more efficient antimicrobial agents for application in monument preservation is required to inhibit or prevent fungal growth on archaeological artifacts (Campana et al. 2020).
Chalcones (1,3-diaryl-2-propen-1-ones) are bio-precursors of flavonoids and isoflavonoids in plants (de Mello et al. 2018). Nowakowska (Nowakowska 2007) stated that chalcone forms the backbone of many natural products and is widely distributed as the main component in vegetables, fruits, teas, and other plants with a great interest for their biological activities. The biological profile of chalcone attracted the attention of organic chemists worldwide to design and develop new chalcone derivatives (Rani et al. 2019). Chalcones exhibited a variety of biological activities including antibacterial (Nanjundaswamy et al. 2022), antifungal (Lahtchev et al. 2008), anticancer (Mohamed et al. 2012, 2014, 2018, 2022; Tantawy et al. 2019; Fathi et al. 2021; Srilaxmi et al. 2021; Helmy et al. 2021; WalyEldeen et al. 2022, 2023; Kamel et al. 2022; Sroor et al. 2022) anti-inflammatory (Rojas et al. 2003), antimalarial (Larsen et al. 2005), and antiviral (Cheenpracha et al. 2006). The mechanism of action for chalcones can be act via the α,β-unsaturated carbonyl moiety as Michael acceptor, thioredoxin reductase, and inhibition of microtubule formation (Menezes and Diederich 2019).
These can contribute to a better understanding and developing new flavonoid-based antifungal agents as multi-target agents in the treatment of fungal deterioration. The different bioactivities and simple structures of natural chalcones have attracted the attention of many researchers, and many structural alterations have been conducted to improve activities (Wang et al. 2016; Jin 2019).
The aim of the present work is conservation of the tested ancient Egyptian mummy cartonnage from microbial deterioration using novel chalcone derivatives as eco-friendly, efficient, safe, and new alternative preservatives for cultural heritage. In our study, eight [1,2,4]triazolo[3,4-a] isoquinoline chalcone derivatives were targeted against thirteen fungal species isolated from the tested ancient Egyptian mummy cartonnage located in El-Lahun regions, Fayoum government, Egypt.
Materials and methods
Source of isolation
A deteriorated ancient Egyptian mummy cartonnage from the Supreme Council of Antiquities (SCA) excavation in El-Lahun regions, Fayoum government, Egypt was tested. The ancient Egyptian mummy cartonnage belonged to the late period of ancient Egypt and its number of the registry is 236. The cartonnage composed of multilayers of fabric structure (linen bandages and papyrus wrappings), which are covered by a gesso layer and colored with different pigments.
The tested ancient Egyptian mummy cartonnage was found in a bad state with many deterioration aspects Fig. 1. It suffered from loss in painting layer accompanied with the occurrence of damage a long two sides of cartonnage, and downfall of gesso layer. Fabric structure suffered from severe fragile and damaged parts with change in its color tending toward dark brown or black, while some linen parts were roasted. The presence of longitudinal cracks in the head and feet area was also observed. The mummy tissues were decayed which result in the appearance of bones.

The tested ancient Egyptian mummy cartonnage belonged to the Supreme Council of Antiquities (SCA) excavation in El-Lahun regions, Fayoum, Egypt. A The whole ancient Egyptian mummy cartonnage with damage a long two sides of cartonnage with downfall of gesso layer, B Appearance of small holes and degradation of linen bandage with longitudinal cracks, C Missing parts in lower area of feet accompanied with loss in structure cohesion of fabric layer, D Decay of mummy tissues with roasting of linen bandages, and E Dryness and gaps in linen bandages with change in its color to dark brown and black resulting in appearance of bone
Isolation and identification of deteriorated fungal species from the tested ancient Egyptian mummy cartonnage
The fungal species were isolated from the twelve different deteriorated parts (top of the vertex, face mask, neck, chest area, abdomen area, left and right sides of cartonnage, leg, the lower part of feet, bones, mummy tissues, and roasted linen) of the tested ancient Egyptian mummy cartonnage by swabbing with sterile cotton swabs. In the laboratory, Potato dextrose agar dishes were streaked with the inoculated swabs. The plates were incubated at 27 °C for 7 days (Naji et al. 2014). The isolated fungi were identified microscopically (Samson et al. 1981; Moubasher 1993).
Molecular identification of the isolated fungal species from the tested ancient Egyptian mummy cartonnage
Molecular identification was performed for the fungal isolates from tested ancient Egyptian mummy cartonnage. DNA extraction was performed by Quick-DNA™ Fungal Microprep Kit (Zymo Research) according to manufacturers’ protocol. The fungal sample (2 mg) was mixed with 95 μL water, 95 μL solid tissue buffer (blue) and 10 μL proteinase K and incubated at 55 °C for 2 h. Then, the mixture was centrifuged at 12,000×g for one minute. The aqueous supernatant has been transferred to a clean tube with the addition of 600 μL genomic binding buffer and completely mixed. It was transferred in the collecting tube of the Zymo-Spin™ IIC-XL column and centrifuged at 12,000×g for 1 min. A DNA pre-wash buffer (400 μL) was added in a new collection tube of the Zymo-Spin™ IIC-XL column and centrifuged at 12,000×g for 1 min. The genomic DNA wash buffer (700 μL) was added to the Zymo-Spin™ IIC-XL column followed by centrifugation at 12,000 × g for 1 min and the flow-through was discarded. The genomic DNA wash buffer (200 μL) was again added and centrifuged at 12,000×g for 1 min, then the collection tube was discarded. The Zymo-Spin™ IIC-XL column was transferred to a sterilized Eppendorf tube and the elution buffer was added directly to the column matrix, and then incubated for five minutes, followed by centrifugation at 12,000×g for 1 min to elute the DNA. The ultra-purified DNA was stored at − 20 °C (Phoku et al. 2014).
The polymerase chain reaction (PCR) was carried out in a 50 μl final volume using MyTaq™ Red Mix (Sigma). According to manufacturers’ protocol, the PCR reactions set up included 25 μL of MyTaq™ Red Mix, 8 μL DNA template, 15 μL Nuclease Free Water, 1 μL of 20 μM ITS1 forward primer, and 1 μL of 20 μM ITS4 primer. The two primers: ITS1 forward primer (5′-TCC GTA GGT GAA CCT GCG G-3′) and ITS4 reverse primer (5′-TCC TCC GCT TAT TGA TAT GC- 3′) (Wirya et al. 2020). Amplification of the PCR products was performed using the thermal cycling conditions: initial denaturing cycle for 6 min at 94 °C (1 cycle), 35 cycles of denaturation for 45 s at 94 °C, annealing for 45 s at 56 °C, and elongation for one minute at 72 °C with a final extension cycle for 5 min at 72 °C.
PCR amplified products were evaluated by electrophoresis with low melting 2% TBE agarose gel (2 g agarose in 98 ml Tris–Borate- EDTA buffer). Ethidium bromide was added to the solution and mixed thoroughly. Electrophoresis was done for 15 min at 70 Volt. The electrophoresed DNA was visualized by UV light (Phoku et al. 2014).
Sequencing was made to purified PCR product on GATC Company by the mean of ABI 3730 XL DNA sequencer using forward and reverse primers only by combining the traditional Sanger technology with the new 454 technology. BLAST program available at NCBI Gen Bank databases (National Center for Biotechnology Information) was used to align DNA strand forward and reverse sequence. Isolate identification was performed based on the analysis of hits from mega-blast (highly similar sequences) output (Bhore et al. 2010).
Chemistry
Melting points were determined on a Stuart melting point device and they are uncorrected. The 1H and 13C-NMR spectra were recorded in DMSO-d6 as solvent at 300 MHz and 75 MHz, respectively on Varian Gemini NMR spectrometer using Tetramethylsilane (TMS) as internal standard. Chemical shifts are reported in δ units (ppm). The IR spectra were recorded as KBr using a Bruker-vector 22 spectrophotometer FTIR. Mass spectra were measured on a Shimadzu GMSS -QP-1000 EX mass spectrometer at 70 eV. The elemental analyses were performed at the Microanalytical Center, Cairo University.
Synthesis of tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)-3-arylprop-2-en-1-one (3–10)
A mixture of 3-acetly[1,2,4]triazolo[3,4-a]isoquinolines 1a,b (0.351 g, 1 mmol) and the appropriate arylaldehydes 2a–e (1 mmol) was dissolved in 20 mL ethanol. Potassium hydroxide (20%, 5 mL) was added to this mixture at 0–5 °C. The reaction mixture was stirred at room temperature for 3 h, then poured over ice containing HCl (1 mL). The mixture was filtered to obtain the solid, washed with water, and dried. The crude product was crystallized from ethanol to afford the chalcone derivatives 3–10.

(E)-3-(4-Bromophenyl)-1-(1-(4-bromophenyl)-8,9-dimethoxy-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)prop-2-en-1-one (3)
Yield: (74%) as an orange solid (from ethanol); m.p 78–80 °C. IR (KBr, cm−1): 1656 (CO); 1H NMR (300 MHz, DMSO-d6): δ, ppm: 2.74 -2.82 (m, 2H, H6), 3.44 (s, 3H, OMe), 3.47–3.48 (m, 1H, H5), 3.72 (s, 3H, OMe), 3.80–4.11 (m, 1H, H5), 6.61 (s, 1H, H10b), 6.80 (s, 1H, H7), 6.92 (s, 1H, H10), 6.97–7.72 (m, 10H, Ar–H + vinyl-H); MS (EI): m/z = 597 (M+). Anal. Calcd. for C27H23Br2N3O3 (597.31): C, 54.29; H, 3.88; N, 7.04. Found: C, 54.43; H, 3.72; N, 7.19.

(E)-1-(1-(4-Bromophenyl)-8,9-dimethoxy-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)-3-(4-methoxyphenyl)prop-2-en-1-one (4)
Yield: (78%) as a yellow solid (from ethanol); m.p 144–146 °C. IR (KBr, cm−1): 1653 (CO); 1H NMR (300 MHz, DMSO-d6): δ, ppm: 2.58–2.72 (m, 2H, H6), 3.41 (s, 3H, OMe), 3.45–3.55 (m, 4H, H5 + OMe), 3.72 (s, 3H, OMe), 4.22–4.33 (m, 1H, H5), 6.61 (s, 1H, H10b), 6.80 (s, 1H, H7), 6.87 -7.95 (m, 11H, H10 + Ar–H + vinyl-H). MS (EI): m/z = 548 (M+). Anal. Calcd. for C28H26BrN3O4 (548.44): C, 61.32; H, 4.78; N, 7.66. Found: C, 61.51; H, 4.82; N, 7.50.

(E)-1-(1-(4-Bromophenyl)-8,9-dimethoxy-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)-3-(3,4,5-trimethoxyphenyl)prop-2-en-1-one (5)
Yield: (76%) as an orange solid (from ethanol); m.p 170–172 °C. IR (KBr, cm−1): 1655 (CO); 1H NMR (300 MHz, DMSO-d6): δ, ppm: 2.62–2.81 (m, 2H, H6), 3.48 (s, 3H, OMe), 3.72–3.74 (m, 7H, 2OMe + H5), 3.84 (s, 6H, 2 OMe), 4.23–4.34 (m, 1H, H5), 6.61 (s, 1H, H10b), 6.80 (s, 1H, H7), 6.87 (s, 1H, H10), 7.10 (s, 2H, Ar–H), 7.33–7.61 (m, 6H, Ar–H + vinyl-H). MS (EI): m/z = 608 (M+). Anal. Calcd. for C30H30BrN3O6 (608.49): C, 59.22; H, 4.97; N, 6.91. Found: C, 59.33; H, 5.12; N, 7.11.

(E)-3-(Benzo[d][1,3]dioxol-5-yl)-1-(1-(4-bromophenyl)-8,9-dimethoxy-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)prop-2-en-1-one (6)
Yield: (80%) as a red solid (from ethanol); m.p 168–170 °C. IR (KBr, cm−1): 1655 (CO); 1H NMR (300 MHz, DMSO-d6): δ, ppm: 2.64–2.83 (m, 2H, H6), 3.48 (s, 3H, OMe), 3.71–3.72 (m, 4H, OMe + H5), 4.23–4.32 (m, 1H, H5), 6.01 (s, 2H, -OCH2-), 6.61 (s, 1H, H10b), 6.79 (s, 1H, H7), 6.87 (s, 1H, H10), 6.96–7.56 (m, 9H, Ar–H + vinyl-H);13C NMR (75 MHz, DMSO-d6): δ, ppm: 26.9, 41.4, 55.4, 55.5, 77.2, 101.7, 106.9, 108.6, 111.8, 111.9, 116.2, 120.4, 125.7, 127.1, 128.3, 128.9, 131.9, 142.0, 142.8, 147.1, 148.1, 148.7, 149.7, 149.8, 179.1. MS (EI): m/z = 562 (M+). Anal. Calcd. for C28H24BrN3O5 (562.42): C, 59.80; H, 4.30; N, 7.47. Found: C, 59.92; H, 4.51; N, 7.62.

(E)-3-(4-Bromophenyl)-1-(8,9-dimethoxy-1-(4-methoxyphenyl)-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)prop-2-en-1-one (7)
Yield: (80%) as an orange solid (from ethanol); m.p 142–144 °C. IR (KBr, cm−1): 1650 (CO); 1H NMR (300 MHz, DMSO-d6): δ, ppm: 2.62–2.78 (m, 2H, H6), 3.39 (s, 3H, OMe), 3.42–3.52 (m, 1H, H5), 3.71 (s, 3H, OMe), 3.75 (s, 3H, OMe), 4.22–4.33 (m, 1H, H5), 6.56 (s, 1H, H10b), 6.73 (s, 1H, H7), 6.93 -7.72 (m, 11H, H10 + Ar–H + vinyl-H). MS (EI): m/z = 548 (M+). Anal. Calcd. for C28H26BrN3O4 (548.44): C, 61.32; H, 4.78; N, 7.66. Found: C, 61.44; H, 4.89; N, 7.78.

(E)-1-(8,9-Dimethoxy-1-(4-methoxyphenyl)-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)-3-(4-methoxyphenyl)prop-2-en-1-one (8)
Yield: (79%) as an orange solid (from ethanol); m.p 120–122 °C. IR (KBr, cm−1): 1658 (CO); 1H NMR (300 MHz, DMSO-d6): δ, ppm: 2.65–2.85 (m, 2H, H6), 3.40 (s, 3H, OMe), 3.43–3.48 (m, 1H, H5), 3.70 (s, 3H, OMe), 3.73 (s, 3H, OMe), 3.75 (s, 3H, OMe), 4.12–4.21 (m, 1H, H5), 6.56 (s, 1H, H10b), 6.73 (s, 1H, H7), 6.82 (s, 1H, H10), 6.95–7.00 (dd, 4H, Ar–H (J = 9.3) + 2 vinyl-H (J = 15.7 Hz), 7.37 (d, 2H, Ar–H, J = 8.7 Hz), 7.54 (d, 2H, Ar–H, J = 9.3 Hz), 7.72 (d, 2H, Ar–H, J = 8.7 Hz);MS (EI): m/z = 499 (M+). Anal. Calcd. for C29H29N3O5 (499.57): C, 69.72; H, 5.85; N, 8.41. Found: C, 69.86; H, 5.98; N, 8.65.

(E)-1-(8,9-Dimethoxy-1-(4-methoxyphenyl)-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)-3-(3,4,5-trimethoxyphenyl)prop-2-en-1-one (9)
Yield: (74%) as a red solid (from ethanol); m.p 170–172 °C. IR (KBr, cm−1): 1656 (CO); 1H NMR (300 MHz, DMSO-d6): δ, ppm: 2.64–2.83 (m, 2H, H6), 3.41 (s, 3H, OMe), 3.44–3.46 (m, 1H, H5), 3.70 (s, 3H, OMe), 3.75 (s, 6H, 2 OMe), 3.80 (s, 6H, 2 OMe), 4.24–4.36 (m, 1H, H5), 6.57 (s, 1H, H10b), 6.73 (s, 1H, H7), 6.87 (s, 1H, H10), 6.95–7.72 (m, 8H, Ar–H + vinyl-H). MS (EI): m/z = 559 (M+). Anal. Calcd. for C31H33N3O7 (559.62): C, 66.53; H, 5.94; N, 7.51. Found: C, 66.72; H, 6.11; N, 7.73.

(E)-3-(Benzo[d][1,3]dioxol-5-yl)-1-(8,9-dimethoxy-1-(4-methoxyphenyl)-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)prop-2-en-1-one (10)
Yield: (79%) as an orange solid (from ethanol); m.p 128–130 °C. IR (KBr, cm−1): 1658 (CO); 1H NMR (300 MHz, DMSO-d6): δ, ppm: 2.61–2.81 (m, 2H, H6), 3.40 (s, 3H, OMe), 3.42–3.52(m, 1H, H5), 3.71 (s, 3H, OMe), 3.74 (s, 3H, OMe), 4.24–4.32 (m, 1H, H5), 6.08 (s, 2H, -OCH2-), 6.57 (s, 1H, H10b), 6.73 (s, 1H, H7), 6.88 (s, 1H, H10), 6.95–7.53 (m, 9H, Ar–H + vinyl-H);13C NMR (75 MHz, DMSO-d6): δ, ppm: 26.8, 41.5, 55.2, 55.3, 55.5, 78.9, 101.7, 106.9, 108.6, 109.3, 112.0, 114.6, 117.7, 120.8, 125.3, 126.4, 128.6, 129.0, 136.8, 140.9, 146.8, 148.1, 148.5, 149.1, 149.5, 154.5, 178.6. MS (EI): m/z = 513 (M+). Anal. Calcd. for C29H27N3O6 (513.55): C, 67.83; H, 5.30; N, 8.18. Found: C, 67.94; H, 5.45; N, 8.23.
Effect of the new eight tested chalcone derivatives on the growth of the isolated thirteen deteriorated fungal species from the tested ancient Egyptian mummy cartonnage:-
The antifungal activities of the new eight chalcones (3–10) were evaluated using agar dilution method (Gupta and Jain 2015). Stock solutions of chalcone compounds were prepared in DMSO. Aliquots of the stock solution were diluted in melted potato dextrose agar at a concentration (1 mg/mL) with vigorously shaking, then poured in Petri plates (9 cm diameter) and let them solidify. Each plate was inoculated at the center with the fungal disc (6 mm) from 7-day-old culture, Itraconazole was used as a standard, and control experiments were performed under similar conditions without the chalcone compounds. All triplicate dishes were incubated at 27 °C for 7 days. Percent of inhibition = (dC − dT) × 100/dC. Where dC is the average diameter of the fungal colony in control and dT is the average diameter of the fungal colony in a treatment group.
Determination of minimum inhibitory concentration (MIC) for the isolated deteriorated fungal species
The minimum inhibitory concentration (MIC) of the new eight tested chalcone derivatives was determined by the agar dilution method. Stock solutions of synthesized compounds were prepared in DMSO. Aliquots of the stock solution were used to prepare series of subsequent concentrations (1, 2, 3, 4, 5, and 6 mg/mL). The plates were incubated at 27 °C for 7 days. MIC of the new eight tested chalcone derivatives expressed as the lowest concentration of chalcone derivatives (mg of the chalcone derivatives /ml of culture medium) at which no visible growth was occurred compared with control after the incubation time (Gupta and Jain 2015).
Results and discussion
Isolation and identification of deteriorated fungal species from the tested ancient Egyptian mummy cartonnage
The data revealed that thirteen fungal species (Aspergillus flavus, A. niger, A. terreus, Athelia bombacina, Aureobasidium iranianum, Byssochlamys spectabilis, Cladosporium cladosporioides, C. ramotenellum, Penicillium crustosum, P. polonicum, Talaromyces atroroseus, T. minioluteus, and T. purpureogenus) accounting 179 colonies were isolated from the twelve different parts (top of the vertex, face mask, neck, chest area, abdomen area, left and right sides of cartonnage, leg, the lower part of feet, bones, mummy tissues and roasted linen) of the tested ancient Egyptian mummy cartonnage, located in the Supreme Council of Antiquities (SCA) excavation, El-Lahun regions, Fayoum, Egypt (Table 1). The biodeterioration of the tested ancient Egyptian mummy cartonnage might be due to the enzymatic activities of the isolated fungal species as well as their ability to grow and feed on textiles and organic materials (skin and fabrics) as recorded by Kavkler et al. (2015) who indicated that organic materials (protein, acrylic, and calcium organic salts), inorganic materials (calcite, gypsum, and silicate fillers), and various additional materials (the animal glue in the paintings), might have contributed to the fungal growth and acted as additional nutrition sources (Mansour et al. 2020). stated that Cladosporium cladosporioides, Penicillium chrysogenum, and Aspergillus flavus producing the deterioration of a child's mummy covered by linen wrapping, in Dahshur, Giza, Egypt. Čavka et al. (2010) isolated seven saprophytic genera (Alternaria spp., Aspergillus fumigatus, Aspergillus nidulans, Chrysosporium spp., Monilia spp., Penicillium spp., and Rhizopus spp.) from different samples (oral, orbital and abdominal cavity and wrapping bandages) of the mummified body from the Archeological Museum in Zagreb, Croatia. Also, Skrlin et al. (2011) isolated Aspergillus, Bacillus, Penicillium, Sarcina, and Shewanella species from St. Marcian mummy in Rijeka, Croatia. The existence of the isolated thirteen fungal species on the tested ancient Egyptian mummy cartonnage may stay viable for prolonged periods due to that they were more resistant to low levels of water activity (aw) as recorded by Teixeira et al. (2018) who stated that the water stress and poor ventilation favor the proliferation of fungal species.
In the present investigation, the highest number of the isolated fungal colonies (33 colonies) was recorded on the roasted linen, followed by the left side of cartonnage (30 colonies), and the linen of the top of the vertex (26 colonies) of the tested ancient Egyptian mummy cartonnage which might be due to the ability of fungal species to feed and grow on cellulosic material by mycelial penetration, and secreting cellulosic enzymes leading to the loss of fiber strength and structure cohesion as recorded by Gutarowska et al. (2017) who recorded that historical cellulosic materials can easily decay by fungal species when exposed to high humidity. Also, Tiano (2002) stated that the most frequent deteriorating fungi on cellulosic textiles with high cellulolytic activities are Ascomycetes and Deuteromycetes Šimonovičová et al. (2015) isolated Aspergillus fumigatus, A. niger, Coprinellus xanthothrix, Penicillium polonicum, P. chrysogenum, and Rhizopus stolonifer from textile samples of mummified human remains in Sládkovičovo, Slovakia. In the Jordanian Museum, the historic textile objects were colonized by Alternaria alternate, and A. tenuissima, Aspergillus flavus, A. fumigatus, A. nidulans, A. niger, Penicillium asperum, and P. funiculosum, (Grabek-lejko et al. 2017). The support part of the mummy cartonnage consists mainly of cellulosic textile materials (linen and papyrus), and the binder material that was manufactured from animal or plant glues which make it more susceptible to microbial attack (Abdel-Kareem 2010). Ali et al. (2018) isolated Aspergillus fumigatus, A. niger, A. tamarii, Cladosporium sp. Fusarium solani, and Penicillium chrysogenum from the cartonnage surface, that consist of painted layer, gesso, and the organic materials (cotton, linen, or wool), which deteriorated by fungal species that result in changes in color, chemical, and physical characteristics of cartonnage. Alternaria, Aspergillus, Chaetomium, Cladosporium, and Penicillium sp. had been isolated from archaeological cellulosic materials (Fouda et al. 2019).
Five isolated fungal species (Aspergillus terreus, Cladosporium cladosporioides, C. ramotenellum, Penicillium crustosum, and P. polonicum) accounting 16 colonies were counted from the bone of the tested ancient Egyptian mummy. The obtained result agreed with Šimonovičová et al. (2015) who isolated different fungal species (Aspergillus candidus, A. terreus, A. venenatus, A. versicolor, A. westerdijkiae, Penicillium chrysogenum, and Talaromyces flavus) from a bone sample of mummified human remains in Sládkovičovo, Slovakia, where Aspergillus and Penicillium can produce metabolites that able to dissolve organic and inorganic constituents of bone tissue. Jans et al. (2004) stated that fungal colonization is regularly found in archaeological bone in favorable environment conditions (presence of oxygen and moisture) without destruction of the bone microstructure. The fungal metabolites (organic acids) can dissolve bone minerals, followed by collagen hydrolysis by fungal collagenase to harvest nutrients and to use the bone as a medium (Jans 2013). Piñar et al. (2013) found that Penicillium radicum was the dominant species in the mummies bone from the Capuchin Catacombs of Palermo, Italy. Penicillium species can be involved in the solubilization of the phosphorus that presents in bones minerals as tricalcium phosphate and calcium phosphates.
The tested mummy tissues were deteriorated by six fungal species (Aspergillus flavus, A. terreus, Aureobasidium iranianum, Cladosporium cladosporioides, C. ramotenellum, and Penicillium crustosum) accounting 9 colonies which may be due to the ability of fungal species to grow on dry meat, which is the major source of protein, fats, minerals, and vitamins in ancient mummies. The obtained results agree with Mansour (2018) who stated that xerophilic and halophilic fungi can help in the degradation of the mummy, due to its ability to grow on dry meat with a high level of salinity at minimum water content in a locked environment. The xerophilic fungus can also grow on leather or fabric bindings (Micheluz et al. 2015).
Arroyo (2009) found Aspergillus and Penicillium in the ancient proteinous materials. David (2008) mentioned that different fungal species are often seen in ancient tissues as a result of poor storage of the specimen. The proteinaceous objects with a high degree of impurities can increase their susceptibility to fungal attack (Kavkler et al. 2015). Piñar et al. (2013) found Botryotinia, Giberella, Didymella, Fusarium, Verticillium, Tritirachium, Coprinus, and Coniosporium on the surface and inside the mummy materials from the Capuchin catacombs of Palermo, Italy, while Phialosimplex species were the most dominated species on the mummies samples (skin, muscle, and hair), which are keratin- and collagen-rich materials.
The isolated low count of fungal colonies from the face mask of ancient mummy cartonnage, may be due to the presence of heavy metals in the paint layer as recorded by Tiano (2002) who indicated that the presence of heavy metals (chromium, copper, lead, and zinc) in some pigments can increase the resistance of the paint layer against fungal attack. The microbial examination of the tested mummy cartonnage showed that the presence of fungal growth on the paint layer is less frequent than on the fabric layer, which causes serious damage to the mummy. Brittleness and deep cracks were observed as result of fungal damage to archeological oil painting object (Geweely 2006).
In the present investigation, Aspergilli were the most dominant deteriorated genera in the twelve samples of the tested ancient Egyptian mummy cartonnage, where 101 colonies were recovered, represented 56.40% of the total fungal colony, and constituted three species (Aspergillus flavus, A. niger, A. terreus). Aspergilli and Penicillia genera are the most isolated fungal species from human remains, which can adapt well to the conditions offered by a mummified body characterized by low availability of water (Šimonovičová et al. 2015). The obtained data agree with Kraková et al. (2018) who found that Aspergillus species were the most widely distributed in the mummy from the castle of Krásna Hôrka, in Slovakia with an abundance of (25–51%), followed by Penicillium (3–41%), and Cladosporium (14–38%). Also, Naji et al. (2014) noticed that Aspergilli was the most common genera (48.94%) isolated from the mummy samples in the Yemen National Museum at Sana’a, where A. niger (25.53%) was the most collective species followed by A. flavus (10.63%), while A. fumigatus, A. candidus, and A. ustus were isolated in minimal frequency. Cladosporium and Penicillium were isolated in moderate frequency representing 14.89% and 12.76%, respectively. Aureobasidium pullulans, Chaetomium thermophilum, Mucor circinelloides, Scopulariopsis koningii, Stachybotrys chartarum, Trichoderma hamatum, and Ulocladium chartarum had been isolated in rare incidence from mummy samples. Lavin et al. (2016) found that Aspergillus niger, Aspergillus flavus, Penicillium sp., Cladosporium sp., Scopulariopsis sp., Fusarium sp. and Alternaria sp. produce different pigments which cause esthetic damage.
Aspergillus flavus was the most dominant isolated deteriorated fungal species, where the highest count (46 colonies) was recovered from the ten tested parts out of twelve of the tested ancient Egyptian mummy cartonnage accompanied with a high occurrence which constituted 25.70% of the total fungal count. Helmi et al. (2015) isolated Aspergillus flavus, and A. niger from archaeological funeral masks in Saqqara, Egypt, which caused various deterioration aspects (disintegration, discoloration, cracking, and stains). A. flavus can produce a broad range of hydrolytic enzymes which substantial for nutrition and penetration of different organic substrates (Mellon et al. 2007). It can develop also on both complex protein substrates and complex carbohydrate substrates (Teixeira et al. 2018). Aspergillus terreus was the second tested species in dominance (30 colonies), followed by A. niger (25 colonies) which constituted 16.76% and 13.97% of the total counts from the tested ancient Egyptian mummy, respectively. A. niger is a common contaminant on various materials, which produces toxic secondary metabolites (Lee 2004), as well as allergens (Ali et al. 2018). It can be adapted to different environmental conditions by different metabolic mechanisms (Zhang et al. 2017). The degradation action of A. niger is performed by the production of pectinases, hemicellulase, xylanase, and arabinase (Pařenicová et al. 2000).
Aspergillus spp. was the most frequent species followed by Penicillium spp. (24 colonies) and Cladosporium spp. (19 colonies). Kavkler et al. (2015) demonstrated that P. chrysogenum and Cladosporium spp. were the most frequently isolated from cellulosic and proteinaceous materials. Penicillium and Aspergillus species well-known for their cellulolytic and proteolytic activity on textiles and skin, which menacing the preservation of the mummies. Penicillium chrysogenum was isolated from a burial chamber in Upper Egypt. P. chrysogenum and C. cladosporioides can grow at low temperatures and moisture in an indoor environment. Xerophiles fungi (Aspergillus flavus Cladosporium cladosporioides, and Penicillium chrysogenum) can produce mycotoxin that may affect human health by breathing the high concentration of aflatoxins and fungal spore when opening the burial chamber after its one thousand years of being closed (Mansour 2018). Cladosporium cladosporioides and C. tenuissimum were isolated from five historical materials include cellulosic (i.e., cotton, flax, hemp), and proteinaceous (i.e., leather, wool) materials. They cause textile degradation mainly due to their abundant excretion of succinic acid (Kavkler et al. 2015).
Dothideomycetes was the most abundant phylum (29–50%) in (cellulosic material) Library and Archives in Poland, and Czechia. The species of Cladosporium (15.827–19.756%), Aureobasidium (3.833–16.789%), Toxicocladosporium (0.856–3.771%) and Conidiocarpus (0–3.255%) were the most common species in this phylum (Krakova et al. 2018). On the other hand, Krakova et al. (2018) detected new species contaminating cultural material includes Scorias spongiosa, Conidiocarpus caucasicus, Sydowia polyspora, Diaporthe longicolla, Byssochlamys spectabilis, and Graphiopsis chlorocephala, although their presence was in the range of 1–4%. Basidiomycota is not considered as the major fungal contaminant of archival documents, which isolated in lower concentration than Ascomycota species.
Talaromyces and Aureobasidium species were isolated with a relative density of 6.15% and 3.35% respectively. Talaromyces helices (Penicillium teleomorph) has been reported from archaeological cellulolytic materials, which uses cellulose as a carbon source to penetrate cracks and migrate between layer and causes detachment (Ali et al. 2014). The least isolated species in order of density was occupied by Athelia bombacina which represented 0.56% of the total count. Basidiomycota is not considered the major fungal contaminant of archival cellulosic materials. It found in lower concentration compared to Ascomycota species as recorded by Krakova et al. (2018) who stated that few Basidiomycota taxa were revealed on archival items. It is might due to the activation of the germination of basidiospores requires specific conditions, which make it difficult to obtain cultures of characteristic taxa as explained by Nitiu et al. (2019) who found that only Rhodotorula genera representative of the phylum Basidiomycota From Egyptian mummies in the museum of natural sciences of La Plata, Argentina.
Molecular identification of the isolated deteriorated fungal species
In the present study, the isolated deteriorated fungal species from the tested ancient Egyptian mummy cartonnage have been determined by PCR amplification and sequencing as follows: Aspergillus flavus (MT874383), A. niger (MT874510), A. terreus (MT874468), Athelia bombacina (MT874470), Aureobasidium iranianum (MT874508), Byssochlamys spectabilis (MT876602), Cladosporium cladosporioides (MT874517), C. ramotenellum (MT874471), Penicillium crustosum (MT874516), P. polonicum (MT874507), Talaromyces atroroseus (MT874506), T. minioluteus (MT874509), and T. purpureogenus (MT874504) (Table 2 and Fig. 2 and 3). The most dominant isolated fungal species from the tested ancient Egyptian mummy are belonging to Ascomycota and Basidiomycota phylum. Šimonovičová et al. (2015) recorded Ascomycota, Zygomycota, and Basidiomycota on different materials of mummified human remains (funeral clothes, skin, muscles, and bones) in the Kuffner family crypt in Sládkovičovo, Slovakia. The obtained results agree with Abdel-Maksoud and El-Amin (2013) who reported that the most dominant isolated fungal species from the mummy from the Late Period in ancient Egypt are belonging to Ascomycotina and Zygomycotina genera. The percentage (%) of fungi were: Pencillium egyptiacum (25%), Aspergillus fumigates (18.75%), Aspergillus niger (12.5%), Pencillium chrysogenum (12.5%), Rhizopus arrhizus (12.5%) and Rhizopus nigricans (12.5%), and Aspergillus flavus (6.25%).

Phylogenetic analysis of the fungal species isolated from the different deteriorated parts of the tested ancient Egyptian mummy cartonnage belonged to the Supreme Council of Antiquities (SCA) excavation in El-Lahun regions, Fayoum, Egypt. (1) Aspergillus flavus (MT874383), (2) A. niger (MT874510), (3) A. terreus (MT874468), (4) Athelia bombacina (MT874470), (5) Aureobasidium iranianum (MT874508), and (6) Byssochlamys spectabilis (MT876602)

Phylogenetic analysis of the fungal species isolated from the different deteriorated parts of the tested ancient Egyptian mummy cartonnage belonged to the Supreme Council of Antiquities (SCA) excavation in El-Lahun regions, Fayoum, Egypt. (7) Cladosporium cladosporioides (MT874517), (8) C. ramotenellum (MT874471), (9) Penicillium crustosum (MT874516), (10) P. polonicum (MT874507), (11) Talaromyces atroroseus (MT874506), (12) T. minioluteus (MT874509), and (13) T. purpureogenus (MT874504)
Synthesis of chalcone derivatives
The starting 3-acetyl-8,9-dimethoxy-1-phenyl-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinoline 1 was obtained following the reported procedures via the reactions of 3,4-dihydro-6,7-dimethoxyisoquinoline with nitrilimines (Elwan et al. 1996; Hassaneen et al. 2011). Claisen–Schmidt condensation of compound acetyl compound 1 with the mole equivalents of substituted aldehydes 2 in the presence of potassium hydroxide solution at room temperature affords of the corresponding chalcone derivatives 3–10 (Scheme 1). The constitution of the formed chalcones was proved by careful inspection of the different spectral tools. 1H NMR spectra of chalcone derivative number 6 as a representative example indicated two characteristic singlet signals at 3.48 and 3.72 ppm for two methoxy groups. The isoquinoline-H5 and H6 groups appeared as multiplets at 2.64, 3.71, and 4.23 ppm. The –OCH2O– group appeared as a singlet at 6.01 ppm. The isoquinoline H10b, H7, and H10 appeared as three singlets at 6.01, 6.61, and 6.87 ppm. Besides, it revealed the vinyl and aromatic protons as multiplets at δ 6.96–7.56 ppm. Moreover, the 13C NMR spectra of compound 6 indicated 26 different types of carbon signals appeared at their expected positions.

Synthesis of novel tetrahydro-[1,2,4]triazolo[3,4-a]isoquinoline chalcones 3–10
Effect of the new eight tested chalcone derivatives on the growth of the isolated deteriorated fungal species from the tested ancient Egyptian mummy cartonnage
The tested eight chalcone derivatives at a concentration of 1 mg/mL showed antifungal activities against thirteen fungal species that were collected from the tested ancient Egyptian mummy cartonnage as shown in Table 3. The predominant majority of the new tested chalcone derivatives provided effective growth inhibition on the tested isolated fungal species which may be due to the presence of nitrogen atoms as recorded by Srilaxmi et al. (2021) who stated that hetero-aromatic skeletons consist of nitrogen atoms are very significant intermediates because of their numerous biological applications. Wang et al. (2010) showed that the synergistic effect of biological activity depends on the combination of chalcone and 1,2,3-triazole are conjugated. Prasath et al. (2015) synthesized quinolinyl chalcones having a pyrazole ring, which was screened for antibacterial and anti-fungal activity against various gram-positive and gram-negative bacteria, and fungal strains. The compounds proved to be most potent with 20.2 mm against Candida. albicans and 21.4 mm activity against Escherichia coli, respectively, where the presence of electron-withdrawing groups improved the antimicrobial activities. The presence of reactive α, β-unsaturated enone group (_CO–CH=CH–) in chalcones derivatives enable them to exhibit a wide range of biological activities (ur Rashid et al. 2019). Chalcones are well-known chemical groups associated with several biological activities such as antibiotic, anti-inflammatory, and antitumor activities (Silva et al. 2022).
Structural activity relationship (SAR)

The structure can be seen as α,β-unsaturated enone group attached to two rings (A-ring and B-ring). A-ring represents the acetyl part of chalcone, which, is the aryl group attached to [1,2,4]triazolo[3,4-a]isoquinoline group. B-ring represents the aldehyde part. All compounds have two fixed methoxy groups in the base structure. The additional electron-donating groups such as methoxy groups increase the activity as shown in Table 1.
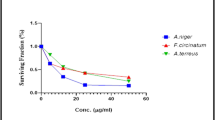
The new tested chalcone derivative 9 is the best inhibitor for the isolated deteriorated fungal species, which has the highest significant inhibition (100%) on the growth of Athelia bombacina, Cladosporium cladosporioides, C. ramotenellum and Talaromyces atroroseus. The activity of the chalcone compound may be due to the presence of six methoxy groups (three methoxy group in A-ring and three methoxy groups in B-ring). The results were found to be in accordance with that reported by Verma et al. (2020) who stated that the compounds bearing electron-donating group such as methyl on phenyl ring exhibited better activity against fungal strains. Also, the antimicrobial activity may refer to the less bulk compound as recorded by Geweely (2009b) who stated that the new complex of a tetrahedral geometry (less bulk) that enhances the rate of entrance through the candidal cell and accordingly increase its anticandidal activity and it can be used in future as novel, more active drug for designing new chemotherapeutic agents. Mohamed et al. (2012) suggested that chalcone derivatives containing electron-releasing groups as OCH3 increase the antimicrobial activity.
The 2′,4′-dihydroxy-3′-methoxychalcone and the 2′,4′-dihydroxychalcone applied on Alternaria alternate, Colletotrichum truncatum, Fusarium equiseti, F. graminearum, F. verticillioides, Phomopsis longicolla and Sclerotium bataticola, which completely suppressed the growth of C. truncatum and P. longicolla at the concentration of 6.25 µg/mL (Jimenez et al. 2014). Also, Jayasinghe et al. (2004) found that five chalcones isolated from Artocarpus nobilis have antifungal efficiency against Cladosporium cladosporioides, and Aspergillus niger.
The new chalcone derivative 5 and 7 came next in rank of antifungal activity and causes complete significant inhibition (100%) of Cladosporium cladosporioides followed by Athelia bombacina (97.6%) and P. polonicum (93.5%), while the least percent of inhibition was shown on Byssochlamys spectabilis (31.1%). It has one two methoxy and one bromo group in A-ring and three methoxy groups in B-ring. The presence of halogen substituent (Br) causes more bulk compound which may also act as electron-withdrawing group is responsible for the antimicrobial activity (Rani et al. 2019). The activity of compound 5 is less than activity of compound 9 which may be due to the presence of one bromine group (Br) (as an electron-withdrawing group) instead of methoxy group (OMe) in A-ring. Burmaoglu et al. (2020) reported that many organo-halogen contains bromine have been recorded to have potential antimicrobial and antioxidant efficiency and also inhibited protein tyrosine phosphatase, and aldose reductase activity.
The tested chalone derivatives 8 and 10 presented moderate inhibitory activity against tested isolated fungal species. ElSohly et al. (2001) isolated chalcones from the Maclura tinctoria, which showed inhibitory activity against Candida albicans and Cryptococcus neoformans. Chalcones are multifunctional molecules, which can show multiple biological activities. licochalcone A act as cell cycle inhibitor (Díaz-Tielas et al. 2016). Most chalcones inhibit the biosynthesis of the yeast cell wall and thus unfold their antifungal potential. The effects of several substitutions of synthetic chalcones against Candida albicans depended on their ability to interact with sulfhydryl groups. Chalcones have been also tested against Cryptococcus neoformans, Candida, Microsporum, Trichophyton and Penicillium species (Lahtchev et al. 2008).
The new synthesized chalcone compound 4 have no activity against Byssochlamys spectabilis and Talaromyces atroroseus, which may be due to the presence of bromine substituent (Br) as an electron-withdrawing group (Br) and methoxy group (OMe) as an electron-donating group, while chalcone derivative 6 have no activity against A. niger, and Talaromyces atroroseus. Also, new chalcone derivative 3 has no activity against Talaromyces atroroseus and T. purpureogenus. The antifungal activity of chalcones has been largely attributed to the reactive enone moiety. The enone unit binds to thiol groups of certain proteins in microbial species. The reactions of chalcones are facilitated by electron-withdrawing (EW) groups at p-position for the thiol attack, which showed antifungal properties (Lahtchev et al. 2008). The presence of an enone linkage would be a structural requirement necessary but not by itself sufficient for the antifungal activity (Nowakowska 2007). López et al. (2001) reported that electron-withdrawing groups in the para-position increased the potency of chalcone, while the presence of these groups in the ortho-position of the ring could introduce important effects that result from the size of substituents and the repulsion between them.
Determination of minimum inhibitory concentration (MIC) for the isolated deteriorated fungal species
The MIC of the eight tested chalcone derivatives on the thirteen isolated deteriorated fungal species from the tested ancient Egyptian mummy cartonnage, El-Lahun regions, Fayoum government, Egypt are presented in Table 4. The MIC of the most efficient chalcone derivatives 13 was ranging from 1 to 3 mg/mL for all isolated deteriorated fungal species. Athelia bombacina, Cladosporium cladosporioides, C. ramotenellum, and Talaromyces atroroseus were the most susceptible species to the lowest concentration of chalcone derivative number 9 (1 mg/mL). Chalcone derivatives with heterocycle moiety (azoles) are another effective approach to improve antifungal activity. Chalcone derivatives, in which quinolinyl N-oxide was used as a B-ring in chalcones, showed antifungal activities against C. gattii (MIC = 7.80 μg/mL) and Paracoccidioides brasiliensis (MIC = 1.90 μg/mL) (de Carvalho Tavares et al. 2011). A chalcone derived from carbazoles was assayed for antibacterial activity against two gram-positive (S. aureus and B. subtilis) and two gram-negative bacterial strains (E. coli and K. pneumonia) and antifungal activity against Fusarium oxysporum, Aspergillus niger, and Aspergillus flavus (Ashok et al. 2016). Konidala et al. (2021) found that the antimicrobial screening of the chalcone compounds showed potent antifungal activity with MIC value of 10 µM and 11 µM against Aspergillus niger.
Azole and non-azole chalcones derivatives showed effective antifungal activity against C. albicans due to inhibit ergosterol biosynthesis and down-regulated ERG11(Sterol 14-demethylase) gene expression (Ahmad et al. 2017). Pyrazine analogs of chalcones showed antifungal activities against Candida spp. (MIC = 7.81 µg/ mL), Aspergillus fumigatus (MIC = 31.25 µg/mL), and Trichophyton mentagrophytes (MIC = 31.25 µg/mL) (Kucerova-Chlupacova et al. 2016). Imidazole–chalcone derivative, exerted effective activities against Candida strains with MIC values ranging from 0.78 to 3.125 µg/mL and significantly decreased ergosterol level of C. krusei (Osmaniye et al. 2018). The series of 1,4-disubstituted-1,2,3- triazole derivatives of chalcones showed potent activities against Candida strains (MIC = 6.5–12.5 µg/mL), C. neoformans (MIC = 12.5 µg/mL), Aspergillus fumigatus and A. niger (MIC = 12.5 µg/mL) (Kant et al. 2016). The high oxidizing power and spontaneous decomposition also make safety, as a common sanitizing agent (Geweely 2011). Some Dehydroacetic acid-chalcone-triazole hybrids were synthesized and evaluated for their antifungal effects by Lal et al. (2018).
The structure–activity relationship showed that the presence of electron-donating groups (OCH3 and CH3) on the phenyl ring highly enhances the antimicrobial properties (Xu et al. 2019). Mohamed et al. (2012) found that chalcone, compounds containing electron-releasing groups such as OCH3 increase the antibacterial activity against Staphylococcus aureus, Streptococcus faecalis, Bacillus subtilis, Escherichia coli, Pseudomonas aeuroginosa, Neisseria gonorrhoeae with MIC 20 mg/mL. Andrade et al. (2018) showed that chalcones have antifungal activity against Trichophyton rubrum with a MIC of 7.5 μg/mL, while 2′4′-dihydroxychalcone inhibited the growth of Candida spp. at a concentration of 15.6 μg/mL, and the action of chalcone was involved in the ergosterol and fungal membrane.
The growth of Cladosporium cladosporioides was found more susceptible to the new tested chalcone derivative 5 when exposed to the lowest concentration (1 mg/mL). The tested chalcone derivative 5 at 4 mg/mL showed complete significant inhibition against Aspergillus flavus, and Byssochlamys spectabilis. The MIC of the chalcone derivative 5 was ranging from 1 to 4 mg/mL for all isolated deteriorated fungal species. The structure–activity relationship revealed that the position of the bromo atom on the benzene ring is highly favored for antimicrobial activity. This may be attributed to the greater hydrophobic effect of bromo group at 4-position (Abdullah et al. 2014). Chalcone derivatives with electron-withdrawing substituents (-Br) at 2 and 4-position on phenyl ring showed increases in the cytotoxicity level of tested compounds (Xu et al. 2019). The introduction of different electronic properties of two functional groups (–Br and –OMe) on phenyl rings exhibited superior antimicrobial activity (Yadav et al. 2017).
The MIC of the chalcone derivative 8 was ranging from 2 to 5 mg/mL for all isolated deteriorated fungal species. Aspergillus flavus, A. terreus, Cladosporium cladosporioides, and T. minioluteus were completely inhibited at 5 mg/mL of chalcone derivative 8. Dimeric chalcone isolated from Mallotus philippinensis exhibited inhibitory activity that inhibits 50% of the microbial growth of (MIC50) are 4–8 and 16 μg/mL against Cryptococcus neoformans and Aspergillus fumigatus, respectively (Kulkarni et al. 2014). Prenylated chalcone isolated from the Maclura tinctoria leaves showed antifungal activities against Candida albicans and C. neoformans (MIC50 = 15 μg/mL and 7 μg/mL, respectively) (ElSohly et al. 2001). Licochalcone-A showed antifungal activity against C. albicans and Trichophyton rubrum with minimum inhibitory concentration (MIC) 62.5–150 μM and 11.52 μM, respectively by induction of genes related to the ergosterol biosynthesis pathway and genes encoding enzymes involved in cell-wall synthesis and glyoxylate cycle (Seleem et al. 2016).
The growth of four fungal species (Aureobasidium iranianum, C. ramotenellum, Penicillium crustosum, and T. purpureogenus) were the most sensitive species to new chalcone derivative 7 with percent of inhibition at 100% at 3 mg/mL, while Aspergillus flavus, A. niger, Athelia bombacina, and P. polonicum were completely inhibited with 5 mg/mL. The MIC of the chalcone derivative 7 was ranging from 3 to 5 mg/mL for all isolated deteriorated fungal species. An unsubstituted chalcone showed antifungal activity against Trichophyton rubrum with a MIC of 7.5 μg/mL and it reduced ergosterol content, while the enzymatic activity of Fas Cell Surface Death Receptor (FAS gene) was inhibited with values of 68.23 and 17.1 μg/mL (Bitencourt et al. 2013). 2′4′-dihydroxychalcone at a concentration of 15.6 μg/mL inhibited the growth of Candida spp. by involving in the ergosterol and fungal membrane (Andrade et al. 2018). Jin (2019) showed that the chalcone skeleton was also found to be fundamental for antifungal activity. Lahtchev et al. (2008) demonstrated that chalcones may react with some proteins involved in cell separation and DNA was not the main target. Burmaoglu et al. (2017) reported that the fluoro atoms in the ring of 2′, 4′, 6′-trimethoxychalcone did not enhance antifungal activity. Diazenyl chalcones showed antifungal activity against Aspergillus fumigatus (MIC = 31.25 μg/mL) and C. albicans (MIC = 15 μg/mL) (Kaur and Narasimhan 2018).
The MIC of the chalcone derivative 10 was ranging from 2 to 6 mg/mL for all isolated deteriorated fungal species. C. ramotenellum, and Penicillium crustosum were completely inhibited with 2 mg/mL of chalcone derivative 10. Nowakowska (2007) showed that the most active chalcone compound does not have an electron-withdrawing group in the para-position of ring A, but also does not have a substituent in ortho-position. This compound showed strong antifungal activities against Microsporum canis (MIC ¼ 25 mg/mL), Microsporum gypseum (1.5 mg/mL), Trichophyton mentagrophytes (3 mg/mL), Trichophyton rubrum (3 mg/mL) and Epidermophyton floccosum (0.5 mg/mL).
The least and last antifungal activities were shown with the new chalcone derivatives 3, 4, and 6, where the MIC of three chalcone derivatives was ranging from 3 to 6 mg/mL. A. niger require a higher concentration (6 mg/mL) of chalcone derivatives 4, and 6 for complete inhibition. Kucerova-Chlupacova et al. (2016) reported that 2-bromo or 4-bromo substitution had an inhibiting effect on the growth of T. interdigitale (MIC 3.9–7.81 µmol/L), while halogenated chalcone derivatives also inhibited the growth of Candida spp. Wei et al. (2016) examined chalcone derivatives against Gram-positive (Staphylococcus aureus and Streptococcus mutans), four Gram-negative (Escherichia coli, Salmonella typhimurium, and Pseudomonas aeruginosa) and fungus (Candida albicans) strain with MIC ranging from 1 to 64 µg/mL. Muškinja et al. (2016) have reported a series of ferrocenyl chalcones showed antibacterial and antifungal activity against five species of bacteria (Staphylococcus aureus, Bacillus cereus, B. subtilis, Escherichia coli, and Proteus mirabilis) and five species of fungi (Aspergillus niger, Candida albicans, Penicillium italicum, Mucor mucedo, Trichoderma viride). The measured MIC values of synthesized compounds were in the range of 0.0352–0.8873 mg/mL. Xu et al. (2019) suggested that chalcone derivatives were a promising candidate for the development of new antimicrobial agents in the future.
Conclusion
The essential and crucial component of both the Egyptian and the world's cultural legacy are mummies. Its susceptibility to infection by fungal colonies influenced by the range of materials utilized and environmental variables. The impact of microbial activity on the deterioration of mummy cartonnage is a global problem along with its disinfection over time is challenging for preserving them for the next generation. The Ascomycota (Aspergillus flavus, Aspergillus terreus and by A. niger) made up majority of the isolated core phyla. Application of the most efficient new chalcone derivative 9 {(E)-1-(8,9-Dimethoxy-1-(4-methoxyphenyl)-1,5,6,10b-tetrahydro-[1,2,4]triazolo[3,4-a]isoquinolin-3-yl)-3-(3,4,5 trimethoxyphenyl)prop-2-en-1-one} with three methoxy groups as an electron-donating group and one methoxy group (electron-withdrawing group) can be successfully used with minimum inhibitory concentration (MIC) ranging from 1 to 3 mg/mL against fungal deterioration instead of using physical and chemical disinfection to avoid the unfavorable effects on the artifacts, public health and environment.
Data availability
Data available at request.
References
Abdel-Kareem O (2010) Fungal deterioration of historical textiles and approaches for their control in Egypt. E-Preserv Sci 7:40–47
Abdel-Maksoud G, El-Amin AR (2013) The investigation and conservation of a gazelle mummy from the Late Period in ancient Egypt. Mediterr Archaeol Archaeom 13:45–67
Abdullah MI, Mahmood A, Madni M et al (2014) Synthesis, characterization, theoretical, anti-bacterial and molecular docking studies of quinoline based chalcones as a DNA gyrase inhibitor. Bioorg Chem 54:31–37
Ahmad A, Wani MY, Patel M et al (2017) Synergistic antifungal effect of cyclized chalcone derivatives and fluconazole against Candida albicans. MedChemComm 8:2195–2207
Ali DMI, Abdel-Rahman TM, El-Badawey NF, Ali EAM (2014) Control of fungal paper deterioration by antifungal drugs, essential oils, gamma and laser irradiation. Egypt J Bot 54:219–246. https://doi.org/10.21608/ejbo.2014.489
Ali MF, El Sheikha AM, Ali AE (2016) Analytical study and conservation of gilded mummiform cartonnage from the greco-roman period in Cairo museum. Mediterr Archaeol Archaeom 16:127–137. https://doi.org/10.5281/zenodo.53070
Ali MF, Mansour MMA, Badr NM, Salem MZM (2018) A study of biodeterioration and chromatic alterations of painted and gilded mummy cartonnage at the Saqqara Museum Storeroom Egypt. Archaeometry 60:845–858. https://doi.org/10.1111/arcm.12340
Andrade JT, Santos FRS, Lima WG et al (2018) Design, synthesis, biological activity and structure-activity relationship studies of chalcone derivatives as potential anti-Candida agents. J Antibiot (tokyo) 71:702–712
Arroyo I (2009) The role of fungi in the deterioration of movable and immovable cultural heritage. E-Conservation 9:40–50
Ashok D, Ravi S, Ganesh A et al (2016) Microwave-assisted synthesis and biological evaluation of carbazole-based chalcones, aurones and flavones. Med Chem Res 25:909–922
Baltic MZ, Boskovic M (2015) When man met meat: meat in human nutrition from ancient times till today. Procedia Food Sci 5:6–9
Bhore SJ, Ravichantar N, Loh CY (2010) Screening of endophytic bacteria isolated from leaves of Sambung Nyawa [Gynura procumbens (Lour.) Merr.] for cytokinin-like compounds. Bioinformation 5:191
Bitencourt TA, Komoto TT, Massaroto BG et al (2013) Trans-chalcone and quercetin down-regulate fatty acid synthase gene expression and reduce ergosterol content in the human pathogenic dermatophyte Trichophyton rubrum. BMC Complement Altern Med 13:1–6
Borrego S, Perdomo I (2012) Aerobiological investigations inside repositories of the National Archive of the Republic of Cuba. Aerobiologia (bologna) 28:303–316
Burmaoglu S, Algul O, Gobek A et al (2017) Design of potent fluoro-substituted chalcones as antimicrobial agents. J Enzyme Inhib Med Chem 32:490–495
Burmaoglu S, Akin E, Kaya R, Kazancioglu M (2020) Synthesis of novel organohalogen chalcone derivatives and screening of their molecular docking study and some enzymes inhibition effects. J Mol Struct 1208:127868. https://doi.org/10.1016/j.molstruc.2020.127868
Campana R, Sabatini L, Frangipani E (2020) Moulds on cementitious building materials—problems, prevention and future perspectives. Appl Microbiol Biotechnol 104:509–514. https://doi.org/10.1007/s00253-019-10185-7
Čavka M, Glasnović A, Janković I et al (2010) Microbiological analysis of a mummy from the archeological museum in Zagreb. Coll Antropol 34:803–805
Cheenpracha S, Karalai C, Ponglimanont C et al (2006) Anti-HIV-1 protease activity of compounds from Boesenbergia pandurata. Bioorg Med Chem 14:1710–1714
David R (2008) Egyptian mummies and modern science. Cambridge University Press
de Carvalho TL, Johann S, de Almeida Alves TM et al (2011) Quinolinyl and quinolinyl N-oxide chalcones: synthesis, antifungal and cytotoxic activities. Eur J Med Chem 46:4448–4456
de Mello MVP, Abrahim-Vieira BDA, Domingos TFS et al (2018) A comprehensive review of chalcone derivatives as antileishmanial agents. Eur J Med Chem 150:920–929. https://doi.org/10.1016/j.ejmech.2018.03.047
DeAraujo A, Vasanthakumar A, Sepulveda M et al (2016) Investigation of the recent microbial degradation of the skin of the Chinchorro mummies of Ancient Chile. J Cult Herit 22:999–1005. https://doi.org/10.1016/j.culher.2015.11.004
Díaz-Tielas C, Graña E, Reigosa MJ, Sánchez-Moreiras AM (2016) Biological activities and novel applications of chalcones. Planta Daninha 34:607–616. https://doi.org/10.1590/S0100-83582016340300022
ElSohly HN, Joshi AS, Nimrod AC et al (2001) Antifungal chalcones from Maclura tinctoria. Planta Med 67:87–89. https://doi.org/10.1055/s-2001-10621
Elwan NM, Abdelhadi HA, Abdallah TA, Hassaneen HM (1996) Synthesis of [1,2,4]triazolo[3,4-a]isoquinolines and pyrrolo[2,1-a]isoquinolines using α-keto hydrazonoyl halides. Tetrahedron 52:3451–3456. https://doi.org/10.1016/0040-4020(96)00024-5
Farooq A, Choudhary I, Tahara S et al (2002) Detoxification of terpinolene by plant pathogenic fungus Botrytis cinerea. Zeitschrift Für Naturforsch C 57:863–866
Fathi EM, Sroor FM, Mahrous KF et al (2021) Design, synthesis, in silico and in vitro anticancer activity of novel bis-furanyl-chalcone derivatives linked through alkyl spacers. ChemistrySelect 6:6202–6211. https://doi.org/10.1002/slct.202100884
Fouda A, Abdel-Maksoud G, Ali Abdel-Rahman M et al (2019) Eco-friendly approach utilizing green synthesized nanoparticles for paper conservation against microbes involved in biodeterioration of archaeological manuscript. Int Biodeterior Biodegrad 142:160–169. https://doi.org/10.1016/j.ibiod.2019.05.012
Geweely NSI (2006) Non-toxic fumigation and alternative control techniques against fungal colonization for preserving archaeological oil painting. Int J Bot 2:353–362
Geweely NS (2009a) Novel inhibition of some pathogenic fungal and bacterial species by new synthetic phytochemical coumarin derivatives. Ann Microbiol 59:359–368
Geweely NS (2009b) Anticandidal cytotoxicity, antitumor activities, and purified cell wall modulation by novel Schiff base ligand and its metal (II) complexes against some pathogenic yeasts. Arch Microbiol 191:687–695
Geweely NSI (2011) Evaluation of ozone for preventing fungal influenced corrosion of reinforced concrete bridges over the River Nile Egypt. Biodegradation 22:243–252
Geweely NS (2022) A novel comparative review between chemical, natural essential oils and physical (ozone) conservation of archaeological objects against microbial deterioration. Geomicrobiol J 39:531–540. https://doi.org/10.1080/01490451.2022.2043959
Gock MA, Hocking AD, Pitt JI, Poulos PG (2003) Influence of temperature, water activity and pH on growth of some xerophilic fungi. Int J Food Microbiol 81:11–19
Grabek-lejko D, Tekiela A, Kasprzyk I (2017) Risk of biodeterioration of cultural heritage objects, stored in the historical and modern repositories in the Regional Museum in Rzeszow (Poland). A case study. Int Biodeterior Biodegrad 123:46–55. https://doi.org/10.1016/j.ibiod.2017.05.028
Gupta D, Jain DK (2015) Chalcone derivatives as potential antifungal agents: synthesis, and antifungal activity. J Adv Pharm Technol Res 6:114–117. https://doi.org/10.4103/2231-4040.161507
Gutarowska B, Pietrzak K, Machnowski W, Milczarek JM (2017) Historical textiles—a review of microbial deterioration analysis and disinfection methods. Text Res J 87:2388–2406. https://doi.org/10.1177/0040517516669076
Hassaneen HM, Hassaneen HME, Mohammed YS, Pagni RM (2011) Synthesis, reactions and antibacterial activity of 3-acetyl[1,2,4]triazolo[3,4-a]isoquinoline derivatives using chitosan as heterogeneous catalyst under microwave irradiation. Zeitschrift Für Naturforsch B 66:299–310. https://doi.org/10.1515/znb-2011-0313
Helmi FM, Ali NM, Ismael SM (2015) Nanomaterials for the inhibition of microbial growth on ancient Egyptian funeral masks. Mediterr Archaeol Archaeom 15:87–95
Helmy MT, Sroor FM, Mahrous KF et al (2021) Anticancer activity of novel 3-(furan-2-yl)pyrazolyl and 3-(thiophen-2-yl)pyrazolyl hybrid chalcones: synthesis and in vitro studies. Arch Pharm (weinheim). https://doi.org/10.1002/ardp.202100381
Jans MME (2013) Microbial bioerosion of bone—a review. J Petrol 369:1689–1699. https://doi.org/10.1017/CBO9781107415324.004
Jans MME, Nielsen-Marsh CM, Smith CI et al (2004) Characterisation of microbial attack on archaeological bone. J Archaeol Sci 31:87–95. https://doi.org/10.1016/j.jas.2003.07.007
Jay JM, Loessner MJ, Golden DA (2008) Modern food microbiology. Springer Science & Business Media
Jayasinghe L, Balasooriya BAIS, Padmini WC et al (2004) Geranyl chalcone derivatives with antifungal and radical scavenging properties from the leaves of Artocarpus nobilis. Phytochemistry 65:1287–1290. https://doi.org/10.1016/j.phytochem.2004.03.033
Jimenez CM, Sampietro DA, Sgariglia MA et al (2014) Isolation, identification and usefulness of antifungal compounds from Zuccagnia punctata for control of toxigenic ear rot pathogens. Nat Prod Commun 9:1934578X1400901014
Jin Y (2019) Recent advances in natural antifungal flavonoids and their derivatives. Bioorg Med Chem Lett 29:126589. https://doi.org/10.1016/j.bmcl.2019.07.048
Kakakhel MA, Wu F, Gu JD et al (2019) Controlling biodeterioration of cultural heritage objects with biocides: a review. Int Biodeterior Biodegrad 143:104721. https://doi.org/10.1016/j.ibiod.2019.104721
Kamel MG, Sroor FM, Othman AM et al (2022) Structure-based design of novel pyrazolyl–chalcones as anti-cancer and antimicrobial agents: synthesis and in vitro studies. Monatsh Chem 153:211–221. https://doi.org/10.1007/s00706-021-02886-5
Kant R, Kumar D, Agarwal D et al (2016) Synthesis of newer 1, 2, 3-triazole linked chalcone and flavone hybrid compounds and evaluation of their antimicrobial and cytotoxic activities. Eur J Med Chem 113:34–49
Kaur H, Narasimhan B (2018) Synthesis, characterization, antimicrobial and antioxidant potential of diazenyl chalcones. Curr Top Med Chem 18:844–856
Kavkler K, Gunde-Cimerman N, Zalar P, Demšar A (2015) Fungal contamination of textile objects preserved in Slovene museums and religious institutions. Int Biodeterior Biodegrad 97:51–59. https://doi.org/10.1016/j.ibiod.2014.09.020
Konidala SK, Kotra V, Danduga RCSR et al (2021) Design, multistep synthesis and in-vitro antimicrobial and antioxidant screening of coumarin clubbed chalcone hybrids through molecular hybridization approach. Arab J Chem 14:103154. https://doi.org/10.1016/j.arabjc.2021.103154
Kraková L, Šoltys K, Puškárová A et al (2018) The microbiomes of a XVIII century mummy from the castle of Krásna Hôrka (Slovakia) and its surrounding environment. Environ Microbiol 20:3294–3308. https://doi.org/10.1111/1462-2920.14312
Krakova L, Soltys K, Otlewska A et al (2018) Comparison of methods for identification of microbial communities in book collections: culture-dependent (sequencing and MALDI-TOF MS) and culture-independent (Illumina MiSeq). Int Biodeterior Biodegrad 131:51–59. https://doi.org/10.1016/j.ibiod.2017.02.015
Kucerova-Chlupacova M, Vyskovska-Tyllova V, Richterova-Finkova L et al (2016) Novel halogenated pyrazine-based chalcones as potential antimicrobial drugs. Molecules 21:1421
Kulkarni RR, Tupe SG, Gample SP et al (2014) Antifungal dimeric chalcone derivative kamalachalcone E from Mallotus philippinensis. Nat Prod Res 28:245–250
Lahtchev KL, Batovska DI, Parushev SP et al (2008) Antifungal activity of chalcones: a mechanistic study using various yeast strains. Eur J Med Chem 43:2220–2228. https://doi.org/10.1016/j.ejmech.2007.12.027
Lal K, Yadav P, Kumar A et al (2018) Design, synthesis, characterization, antimicrobial evaluation and molecular modeling studies of some dehydroacetic acid-chalcone-1, 2, 3-triazole hybrids. Bioorg Chem 77:236–244
Larsen M, Kromann H, Kharazmi A, Nielsen SF (2005) Conformationally restricted anti-plasmodial chalcones. Bioorg Med Chem Lett 15:4858–4861. https://doi.org/10.1016/j.bmcl.2005.07.012
Lavin P, Saravia SGD, Guiamet P (2016) Scopulariopsis sp. and Fusarium sp. in the documentary heritage: evaluation of their biodeterioration ability and antifungal effect of two essential oils. Microb Ecol 71:628–633. https://doi.org/10.1007/s00248-015-0688-2
Lee MW (2004) Fungal facts: solving fungal problems in heritage collections. J Am Inst Conserv 43:114–116
López SN, Castelli MV, Zacchino SA et al (2001) In vitro antifungal evaluation and structure-activity relationships of a new series of chalcone derivatives and synthetic analogues, with inhibitory properties against polymers of the fungal cell wall. Bioorg Med Chem 9:1999–2013. https://doi.org/10.1016/S0968-0896(01)00116-X
Maekawa S (1998) Conservation of the royal mummy collection at the Egyptian Museum. Oxyg Museum Cases, Maekawa, S ed, Getty Conserv Institute, Los Angeles 1–5
Mansour M (2018) Impact of storage conditions on biodeterioration of ancient egyptian child mummies by xerophilic fungi. Egypt J Archaeol Restor Stud 8:97–107. https://doi.org/10.21608/ejars.2018.23501
Mansour MMA, El-hefny M, Salem MZM, Ali HM (2020) The biofungicide activity of some plant essential oils for the cleaner production of model linen fibers similar to those used in Ancient Egyptian mummification. Processes 8:79. https://doi.org/10.3390/pr8010079
Mellon JE, Cotty PJ, Dowd MK (2007) Aspergillus flavus hydrolases: their roles in pathogenesis and substrate utilization. Appl Microbiol Biotechnol 77:497–504
Menezes JCJMDS, Diederich MF (2019) Natural dimers of coumarin, chalcones, and resveratrol and the link between structure and pharmacology. Eur J Med Chem 182:111637. https://doi.org/10.1016/j.ejmech.2019.111637
Michaelsen A, Pinzari F, Nicoletta B, Guadalupe P (2013) Monitoring the effects of different conservation treatments on paper-infecting fungi. Int Biodeterior Biodegrad 84:333–341
Micheluz A, Manente S, Tigini V et al (2015) The extreme environment of a library: Xerophilic fungi inhabiting indoor niches. Int Biodeterior Biodegrad 99:1–7. https://doi.org/10.1016/j.ibiod.2014.12.012
Mohamed MF, Mohamed MS, Shouman SA et al (2012) Synthesis and biological evaluation of a novel series of chalcones incorporated pyrazole moiety as anticancer and antimicrobial agents. Appl Biochem Biotechnol 168:1153–1162. https://doi.org/10.1007/s12010-012-9848-8
Mohamed MF, Mohamed MS, Fathi MM et al (2014) Chalcones incorporated pyrazole ring inhibit proliferation, cell cycle progression, angiogenesis and induce apoptosis of MCF7 cell line. Anticancer Agents Med Chem 14:1282–1292
Mohamed MF, Hassaneen HM, Abdelhamid IA (2018) Cytotoxicity, molecular modeling, cell cycle arrest, and apoptotic induction induced by novel tetrahydro-[1,2,4]triazolo[3,4-a]isoquinoline chalcones. Eur J Med Chem 143:532–541. https://doi.org/10.1016/j.ejmech.2017.11.045
Mohamed MF, Ibrahim NS, Saddiq AA et al (2022) Theoretical and molecular mechanistic investigations of novel (3-(furan-2-yl)pyrazol-4-yl) chalcones against lung carcinoma cell line (A549). Naunyn Schmiedebergs Arch Pharmacol. https://doi.org/10.1007/s00210-022-02344-x
Moubasher AH (1993) Soil fungi in Qatar and other Arab countries. The Centre for Scientific and Applied Research, University of Qatar
Muškinja J, Burmudžija A, Ratković Z et al (2016) Ferrocenyl chalcones with O-alkylated vanillins: synthesis, spectral characterization, microbiological evaluation, and single-crystal X-ray analysis. Med Chem Res 25:1744–1753
Naji KM, Abdullah QYM, Al-Zaqri AQM, Alghalibi SM (2014) Evaluating the biodeterioration enzymatic activities of fungal contamination isolated from some ancient Yemeni mummies preserved in the national museum. Biochem Res Int 2014:9. https://doi.org/10.1155/2014/481508
Nanjundaswamy S, Gurumallappa HMK et al (2022) Synthesis, crystal structure, in-silico ADMET, molecular docking and dynamics simulation studies of thiophene-chalcone analogues. J Mol Struct 1247:131365. https://doi.org/10.1016/J.MOLSTRUC.2021.131365
Nitiu DS, Mallo AC, Elíades LA et al (2019) Fungal monitoring in an exhibition room with Egyptian mummies in the museum of natural sciences of La Plata, Argentina. Int J Conserv Sci 10:291–306
Nowakowska Z (2007) A review of anti-infective and anti-inflammatory chalcones. Eur J Med Chem 42:125–137. https://doi.org/10.1016/j.ejmech.2006.09.019
Osmaniye D, Kaya Cavusoglu B, Saglik BN et al (2018) Synthesis and anticandidal activity of new imidazole-chalcones. Molecules 23:831
Pařenicová L, Kester HCM, Benen JAE, Visser J (2000) Characterization of a novel endopolygalacturonase from Aspergillus niger with unique kinetic properties. Febs Lett 467:333–336
Phoku JZ, Barnard TG, Potgieter N, Dutton MF (2014) Fungi in housefly (Musca domestica L.) as a disease risk indicator—a case study in South Africa. Acta Trop 140:158–165. https://doi.org/10.1016/j.actatropica.2014.08.019
Piñar G, Piombino-Mascali D, Maixner F et al (2013) Microbial survey of the mummies from the capuchin catacombs of palermo, italy: biodeterioration risk and contamination of the indoor air. FEMS Microbiol Ecol 86:341–356. https://doi.org/10.1111/1574-6941.12165
Prasath R, Bhavana P, Sarveswari S et al (2015) Efficient ultrasound-assisted synthesis, spectroscopic, crystallographic and biological investigations of pyrazole-appended quinolinyl chalcones. J Mol Struct 1081:201–210
Rani A, Anand A, Kumar K, Kumar V (2019) Recent developments in biological aspects of chalcones: the odyssey continues. Expert Opin Drug Discov 14:249–288. https://doi.org/10.1080/17460441.2019.1573812
Rojas J, Payá M, Dominguez JN, Ferrándiz ML (2003) ttCH, a selective inhibitor of inducible nitric oxide synthase expression with antiarthritic properties. Eur J Pharmacol 465:183–189
Samson RA, Hoekstra ES, Van Oorschot CAN (1981) Introduction to food-borne fungi. Centraalbureau voor Schimmelcultures
Seleem D, Benso B, Noguti J et al (2016) In vitro and in vivo antifungal activity of lichochalcone-A against Candida albicans biofilms. PLoS ONE 11:e0157188
Sequeira SO, Phillips AJL, Cabrita EJ, Macedo MF (2017) Antifungal treatment of paper with calcium propionate and parabens: short-term and long-term effects. Int Biodeterior Biodegrad 120:203–215
Silva LS, Véras JH, Fernandes AS et al (2022) Novel sulfonamide-chalcone hybrid stimulates inflammation, angiogenesis and upregulates vascular endothelial growth factor (VEGF) in vivo. Microvasc Res 139:104253. https://doi.org/10.1016/J.MVR.2021.104253
Šimonovičová A, Kraková L, Pangallo D et al (2015) Fungi on mummified human remains and in the indoor air in the Kuffner family crypt in Sládkovičovo (Slovakia). Int Biodeterior Biodegrad 99:157–164. https://doi.org/10.1016/j.ibiod.2014.12.011
Skrlin J, Kavur L, Mlinarić EM et al (2011) Microbiological analysis of the mummy of St. Marcian Coll Antropol 35:923–924
Srilaxmi D, Sreenivasulu R, Mak K, Rao M (2021) Design, synthesis, anticancer evaluation and molecular docking studies of chalcone linked pyrido [4, 3-b ] pyrazin-5 (6 H ) -one derivatives. J Mol Struct. https://doi.org/10.1016/j.molstruc.2020.129851
Sroor FM, Mohamed MF, Abdullah GK et al (2022) Anticancer activity of new bis-(3-(thiophen-2-yl)-1H-pyrazol-4-yl)chalcones: synthesis, in-silico, and in-vitro studies. Polycycl Aromat Compd. https://doi.org/10.1080/10406638.2022.2046616
Sterflinger K, Pinzari F (2012) The revenge of time: fungal deterioration of cultural heritage with particular reference to books, paper and parchment. Environ Microbiol 14:559–566
Tantawy MA, Sroor FM, Mohamed MF et al (2019) Molecular docking study, cytotoxicity, cell cycle arrest and apoptotic induction of novel chalcones incorporating thiadiazolyl isoquinoline in cervical cancer. Anticancer Agents Med Chem 20:70–83. https://doi.org/10.2174/1871520619666191024121116
Teixeira FS, Alves T, Sgubin L et al (2018) Disinfection of ancient paper contaminated with fungi using supercritical carbon dioxide. J Cult Herit 30:110–116. https://doi.org/10.1016/j.culher.2017.09.009
Tiano P (2002) Biodegradation of cultural heritage : decay mechanisms and control methods. Semin Artic New Univ Lisbon, Dep Conserv Restor 7–12
Ur Rashid H, Xu Y, Ahmad N et al (2019) Promising anti-inflammatory effects of chalcones via inhibition of cyclooxygenase, prostaglandin E2, inducible NO synthase and nuclear factor κb activities. Bioorg Chem 87:335–365. https://doi.org/10.1016/j.bioorg.2019.03.033
Valentin N (2001) Microbial contamination and insect infestation in Spanish museums, archives and libraries. Coalit 5–6
Valentin N (2002) Non-toxic methods and systems to control biodeterioration in historic objects of large size. In: 5th European Commission Conference, Cultural Heritage Research. pp 52–56
Verma SK, Verma R, Xue F et al (2020) Antibacterial activities of sulfonyl or sulfonamide containing heterocyclic derivatives and its structure-activity relationships (SAR) studies: a critical review. Bioorg Chem 105:104400. https://doi.org/10.1016/j.bioorg.2020.104400
WalyEldeen AA, El-Shorbagy HM, Hassaneen HM et al (2022) [1,2,4] Triazolo [3,4-a]isoquinoline chalcone derivative exhibits anticancer activity via induction of oxidative stress, DNA damage, and apoptosis in Ehrlich solid carcinoma-bearing mice. Naunyn Schmiedebergs Arch Pharmacol 395:1225–1238. https://doi.org/10.1007/s00210-022-02269-5
WalyEldeen AA, Sabet S, El-Shorbagy HM et al (2023) Chalcones: Promising therapeutic agents targeting key players and signaling pathways regulating the hallmarks of cancer. Chem Biol Interact 369:110297. https://doi.org/10.1016/J.CBI.2022.110297
Wang X-L, Wan K, Zhou C-H (2010) Synthesis of novel sulfanilamide-derived 1, 2, 3-triazoles and their evaluation for antibacterial and antifungal activities. Eur J Med Chem 45:4631–4639
Wang Y, Dong H, Zhao F et al (2016) The synthesis and synergistic antifungal effects of chalcones against drug resistant Candida albicans. Bioorg Med Chem Lett 26:3098–3102. https://doi.org/10.1016/j.bmcl.2016.05.013
Wei Z-Y, Chi K-Q, Yu Z-K et al (2016) Synthesis and biological evaluation of chalcone derivatives containing aminoguanidine or acylhydrazone moieties. Bioorganic Med Chem Lett 26:5920–5925
Wirya G, Gargita I, Sudiarta I (2020) Molecular identification of fungi the causal agent of strawberry wilt disease ball. Int J Biosci Biotechnol 7:64–73
Xu M, Wu P, Shen F et al (2019) Bioorganic chemistry chalcone derivatives and their antibacterial activities : current development. Bioorg Chem 91:103133. https://doi.org/10.1016/j.bioorg.2019.103133
Yadav P, Lal K, Rani P et al (2017) Efficient synthesis and antimicrobial evaluation of 2-((1-substituted-1 H-1, 2, 3-triazol-4-yl)-1-naphthaldehydes and their oxime derivatives. Med Chem Res 26:1469–1480
Zaitseva N (2010) A polysaccharide extracted from Sphagnum moss as antifungal agent in archaeological conservation. Masters Abstr Int 49
Zhang L, Xiao W, Wang Y et al (2017) Chassis and key enzymes engineering for monoterpenes production. Biotechnol Adv 35:1022–1031. https://doi.org/10.1016/j.biotechadv.2017.09.002
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). The authors have no relevant financial support.
Author information
Authors and Affiliations
Contributions
NSG: The idea, the experimental part of microbiology, data interpretation and writing the microbiology part. MMS: The experimental part and writing of the microbiology part. RAA: Saving our needs from antiquities in addition to revision. HMH: The idea and the experimental part of Chemistry. IAA: The idea, data interpretation, and writing the Chemistry part in addition to correspondence.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Communicated by Erko Stackebrandt.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Geweely, N.S., Soliman, M.M., Ali, R.A. et al. Novel eco-friendly [1,2,4]triazolo[3,4-a]isoquinoline chalcone derivatives efficiency against fungal deterioration of ancient Egyptian mummy cartonnage, Egypt. Arch Microbiol 205, 57 (2023). https://doi.org/10.1007/s00203-022-03395-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00203-022-03395-7