
Abstract
Aims/hypothesis
A recent large clinical study has shown that empagliflozin has a lower rate of cardiovascular and all-cause mortality when compared with placebo in patients with type 2 diabetes. We investigated the effect of empagliflozin (compared with glimepiride) on the progression of atherosclerosis, and its possible mechanisms of action.
Methods
Forty-eight 5-week-old male ApoE −/− mice were fed a western diet for 20 weeks and divided into four groups: control (saline, 154 mmol/l NaCl), glimepiride 0.1 mg/kg, empagliflozin 1 mg/kg and empagliflozin 3 mg/kg (n = 12/group). Plaque size and composition in the aortic arch/valve areas and cardiovascular risk variables in the blood and tissues were evaluated. Insulin resistance was estimated by HOMA and adiponectin levels. Body composition was determined using dual-energy x-ray absorptiometry.
Results
After 8 weeks of treatment, the empagliflozin and glimepiride groups exhibited decreased blood glucose levels. Atherosclerotic plaque areas in the aortic arch/valve were significantly smaller in the empagliflozin groups than in the control or glimepiride groups. Insulin resistance and circulating concentrations of TNF-α, IL-6, monocyte chemoattractant protein-1 (MCP-1), serum amyloid A and urinary microalbumin decreased after empagliflozin treatment, and this significantly correlated with plaque size. Empagliflozin treatment reduced weight and fat mass, lipid droplets in the liver, fat cell size, mRNA expression of Tnf, Il6 and Mcp-1 (also known as Ccl2) and the infiltration of inflammatory cells in plaque and adipose tissue compared with the control or glimepiride group. Empagliflozin treatment increased adiponectin levels.
Conclusions/interpretation
Improvements in inflammation and insulin resistance seem to be mechanisms involved in the mitigation of atherosclerosis by empagliflozin.
Similar content being viewed by others

Introduction
Sodium/glucose cotransporter 2 (SGLT2), a 672 amino acid transporter, is mainly expressed within the proximal tubule of the kidney and is reported to be responsible for approximately 90% of renal glucose reabsorption [1]. Given the role of SGLT2 in glucose transport within the kidney, SGLT2 inhibitors have become a focus of developmental and clinical research in the field of diabetes. The first natural SGLT inhibitor to be reported, phlorizin, was isolated from apple tree root bark [2]. Treatment with phlorizin lowered both fasting and postprandial blood glucose levels, independent of insulin secretion, in a diabetic rat model induced by 90% pancreatectomy [3]. In humans, oral administration of phlorizin induces renal glycosuria, weight loss and blood pressure reduction [4], although rapid degradation by lactase–phlorizin hydrolase and poor absorption in the gastrointestinal tract mean that phlorizin has not been developed as a drug for treating diabetes mellitus.
Recently, several highly selective SGLT2 inhibitors derived from the basic phlorizin structure have been synthesised [5]. Unlike many other glucose-lowering therapies, SGLT2 inhibitors act independently of insulin secretion or action [6]. A systematic review and network meta-analysis of randomised controlled trials involving glucose-lowering treatments added to metformin revealed that SGLT2 inhibitors yielded similar HbA1c reduction and less pronounced hypoglycaemia, as well as the additional benefit of weight loss, compared with dipeptidyl peptidase-4 inhibitors, thiazolidinediones and sulfonylureas [7–9]. Empagliflozin effectively lowered blood glucose and provided additional clinical benefits, including body weight and blood pressure reduction [10]. Dapagliflozin reduced fat mass as well as body weight in patients with type 2 diabetes inadequately controlled with metformin [11]. Thus, SGLT2 inhibitors have been found to induce weight loss and decrease blood pressure in patients with diabetes through glucosuria and other indirect mechanisms such as an energy loss of approximately 800–1300 kJ/day [12, 13].
Recently, a large, randomised controlled study showed that patients with type 2 diabetes who were at high risk for cardiovascular events and received empagliflozin had a lower rate of the primary composite cardiovascular outcome and of death from any cause than those receiving placebo [14]. This positive result related mainly to the low rate of hospitalisation for heart failure. In this context, it would be interesting to investigate whether empagliflozin has anti-atherosclerotic properties and a specific role in atheromatous plaque formation. A recent study found that dapagliflozin or ipragliflozin suppressed macrophage foam cell formation [15]. In another recent study, administration of ipragliflozin inhibited the progression of atherosclerosis in a mouse model of repetitive glucose spikes [16]. However, no mechanistic studies have compared the anti-atherosclerotic effects of SGLT2 inhibitors with those of active competitors such as sulfonylurea. Therefore, this study aimed to investigate the effectiveness of empagliflozin in mitigating atherosclerosis, by focusing on its possible mechanisms of action.
Methods
Animal study
Study animals and care
Forty-eight male ApoE −/− mice (Japan SLC, Shizuoka, Japan) were used at 5 weeks old for the generation of the spontaneous atherosclerosis model [17]. At 7 weeks of age, all mice were fed a western diet containing 1.5 g cholesterol (43% energy from carbohydrate and 41% energy from fat; D12079B; Research Diets, New Brunswick, NJ, USA). The animals were kept under 12 h/12 h light/dark cycles with free access to food and water.
This study was approved by the Institutional Animal Care Committee, Seoul National University Bundang Hospital (06-2014-168). Animal experiments were performed in compliance with the Guide for Experimental Animal Research of the Laboratory for Experimental Animal Research, Clinical Research Institute, Seoul National University Bundang Hospital, South Korea.
Treatment with study medication
After being raised for 20 weeks, the mice were divided into four groups according to drug administration: (1) control (normal saline, 154 mmol/l NaCl); (2) glimepiride 0.1 mg/kg; (3) empagliflozin 1 mg/kg and (4) empagliflozin 3 mg/kg (n = 12 per group). The mice were maintained on the high-fat and high-carbohydrate diet. After 8 weeks of oral treatment with study medication, body composition was determined by dual-energy x-ray absorptiometry (DXA) using a PIXImus II apparatus (GE Lunar, Madison, WI, USA). The treatment was randomly assigned by a computer random generator. The experimenters were not blind to the treatment, but they were blind for measurement of experimental outcomes. See electronic supplementary material (ESM) Methods, ‘Animal experiment with ApoE −/− mice’ section. After 8 weeks, the aorta, liver and abdominal subcutaneous and visceral fat tissues were rapidly excised.
Aortic atherosclerosis in ApoE −/− mice
To measure the atherosclerotic lesion area, the aortic arch prepared using the en face method was stained with Oil Red O solution. [18]. Atheroma composition around the aortic valve area was measured by the same method using Oil Red O staining to identify lipid droplets, Masson’s trichrome staining to identify fibrous tissue and CD68 (1:200) immunofluorescence to identify inflammatory cell infiltration. The presence of histiocytes was also examined in Haematoxylin and Eosin (H&E)-stained sections. See ESM Methods.
Biochemical markers associated with cardiovascular risk
At the end of study, after mice had been fasted for 8 h, blood samples were procured and biochemical variables were measured using standard methods. Briefly, plasma glucose (glucose oxidase method [YSI 2300-STAT; Yellow Springs Instruments, Yellow Springs, OH, USA]) and insulin were measured, and HOMA-IR and HOMA-β were calculated [19]. Additionally, lipids (triacylglycerol, HDL-cholesterol and LDL-cholesterol), liver enzymes (aspartate aminotransferase [AST] and alanine aminotransferase [ALT]) and creatine concentration were determined enzymatically (Hitachi 747 Chemistry Analyzer; Hitachi, Tokyo, Japan). TNF-α, IL-6 and monocyte chemoattractant protein-1 (MCP-1) were also measured (Multiplex Assay Kit [RADPK-81K], Millipore, Billerica, MA, USA), as were adiponectin and C-reactive protein concentrations (ELISA; Millipore and BD Biosciences Pharmingen, Heidelberg, Germany, respectively), and serum amyloid A (ELISA; Tridelta Development, Maynooth, Ireland). Additionally, urinary albumin excretion (competitive ELISA; Exocell, Philadelphia, PA, USA) and urinary creatine (enzymatic kit, Stanbio Labs, Boerne, TX, USA) were measured and urinary albumin excretion was expressed as a total urinary albumin (mg)/creatinine (g) ratio. For further details, see ESM Methods.
Histology of liver and adipose tissues
After mice had been treated for 8 weeks with study drugs, the areas and size of lipid droplets that had accumulated in the liver and adipose tissues were measured using a microscope for picture acquisition and the corresponding software for quantification (Image J software 1.50i; National Institutes of Health, Bethesda, MA, USA) [20]. See ESM Methods.
Quantitative real-time PCR
Total RNA was extracted from tissue samples and converted to cDNA. The expression of tnf-α (also known as tnf), Il6 and monocyte chemoattractant protein-1 (Mcp-1, also known as ccl2) was analysed using the Power SYBR Green PCR Master Mix (Applied Biosystems, Warrington, UK) on an ABI 7500 (Applied Biosystems). 18S rRNA was used as an internal control. All experiments were repeated five times. See ESM Methods for details.
Western blot analysis
For western blot analysis, tissues were lysed and resolved on 10% SDS-PAGE and transferred onto a polyvinylidene difluoride (PVDF) membrane (Millipore). For analysis of Akt and GSK, the following primary antibodies were used: anti-Akt, anti-p-Akt, anti-GSK-3β and anti-p-GSK-3β antibodies (1:1000) (Cell Signaling Laboratories, Beverly, MA, USA). An HRP-conjugated anti-rabbit IgG secondary antibody (1:5000) was used (see ESM Methods for full details).
Double immunofluorescence staining of CD68 and CD11c
Immunofluorescence staining of CD68 and CD11c was performed using anti-CD68 (1:200) (Abcam, Cambridge, MA, USA) and anti-CD11 antibodies (1:200) (Abcam). Texas Red X-conjugated goat anti-mouse IgG and Alexa 488-conjugated goat anti-rabbit IgG (1:500) (Invitrogen, Grand Island, NY, USA) were used as secondary antibodies. Sections were mounted and images acquired using fluorescence microscopy (IX81, Olympus, Tokyo, Japan). See ESM for further details.
Cell studies
Rat aortic smooth muscle cells (RAoSMCs) (Bio-bud, Seoul, South Korea) and human umbilical vein endothelial cells (HUVECs) (Lonza, San Diego, CA, USA) were used for in vitro experiments. These cells were negative for mycoplasma before the experiment.
Proliferation of RAoSMCs and HUVECs
The proliferation of RAoSMCs and HUVECs was determined via a bromodeoxyuridine (BrdU) incorporation assay using a commercial ELISA kit (Roche, Mannheim, Germany) [21]. Cells were seeded at 5 × 103 cells/well in a 96-well plate and starved for 24 h in DMEM (plus 0.1% (wt/vol.) FBS). At 1 h prior to induction of cellular proliferation (with platelet-derived growth factor [PDGF; 10 ng/ml] for RAoSMCs or vascular endothelial growth factor [VEGF; 10 ng/ml] for HUVECs), cells were pretreated with empagliflozin (0.1–100 μmol/l) and BrdU incorporation was analysed by spectrophotometry at 450 nm (SoftMax Pro 5.4.1 software, Molecular Devices, Sunnyvale, CA, USA). See ESM Methods for full details.
Migration of RAoSMCs
The migration of RAoSMCs was determined via a wound-healing assay. Briefly, cells were grown to confluence and then starved. Following this, a linear wound was made in the cell monolayer and the medium was supplemented with PDGF (10 ng/ml). Cells were allowed to migrate for 24 h and images of the intersections of the linear wound and each grid line were taken (three fields per well). See ESM Methods for details.
Statistical analyses
Results are reported as the means ± SD or SEM, as indicated. ANOVA and ANCOVA were used to test mean differences between groups. Spearman’s correlation analysis was used for correlation. For all tests, p < 0.05 was considered to be statistically significant.
Results
Animal studies
Atheroma burden in the aorta of ApoE −/− mice
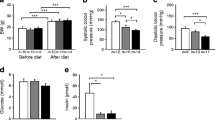
Plaque accumulation in the aortic arch of ApoE −/− mice is shown in Fig. 1a; quantification analysis showed that the plaque area decreased significantly in the empagliflozin groups compared with the control or glimepiride groups after adjusting for weight (Fig. 1b). The atheroma burden in the aortic valve area was also significantly lower in the empagliflozin-treated mice than in the control or glimepiride-treated mice (Fig. 1c, d).

Atheroma burden in the aorta of ApoE −/− mice (n = 12 per group) after 8 weeks of treatment with glimepiride or empagliflozin. (a, c) Representative images of atheromas stained using Oil Red O of the aortic arch (scale bar, 0.5 cm) (a) and aortic valve areas (scale bar, 100 μm) (c) from ApoE −/− mice. The red colour indicates plaque accumulation. (b, d) The atheromatous plaque area (% of total area) was smaller in the aortic arch (b) and aortic valve (d) of empagliflozin-treated mice compared with control or glimepiride-treated mice. Data are means ± SEM. Weight-adjusted ANCOVA was used. *p < 0.05 vs control; † p < 0.05 vs glimepiride
Composition of atheroma in the aortic valve area of ApoE −/− mice
The area occupied by lipid droplets in the atheromas was significantly smaller in empagliflozin-treated mice than in control or glimepiride-treated mice (Fig. 2a, b). There were fewer histiocytes in the atheroma in the empagliflozin groups than in the control and glimepiride groups (Fig. 2c, d).

(a) Oil Red O staining for lipid droplets in atheromas (n = 12 per group) in the aortic valve area (scale bars, 50 μm). (b) The area taken up by lipid droplets was significantly smaller in empagliflozin-treated mice than in the control or glimepiride-treated mice (% of total area). (c) The per cent of histiocytes (indicated by arrows) in H&E sections (scale bar, 50 μm). (d) Significantly fewer histiocytes infiltrated into atheromas in empagliflozin-treated mice than in control or glimepiride-treated mice (% of total area). Data are means ± SEM. *p < 0.05 vs control; † p < 0.05 vs glimepiride
Lower expression levels of CD68 in the aortic valve area were found in the empagliflozin-treated mice than in the other two groups (Fig. 3a, b). The area of connective tissue stained blue with Masson trichrome was slightly greater in the empagliflozin-treated mice than in the control or glimepiride-treated mice, but this difference was not statistically significant (ESM Fig. 1a, b).

(a) Immunohistochemical staining of CD68 in the aortic valve area (n = 12 per group) (scale bars, 100 μm). (b) CD68-positive cells were significantly less abundant in empagliflozin-treated mice than in control or glimepiride-treated mice (% of total cells). Data are means ± SEM.*p < 0.05 vs control; † p < 0.05 vs glimepiride
Change in weight and fat during glimepiride or empagliflozin treatment
For 8 weeks (from 27 weeks of age), weight changes were compared among the control, glimepiride and empagliflozin groups. After 6 weeks of treatment, the weight of mice in the empagliflozin groups (1 and 3 mg/kg) began to fall in relation to that recorded in the control or glimepiride groups, and the difference was maintained thereafter (Fig. 4a). The mass and percentage of fat, assessed using DXA analysis, were lower in the empagliflozin groups (1 and 3 mg/kg) than in the control or glimepiride group (Fig. 4b). Representative DXA images are shown in Fig. 4c.

(a) Weight change during the 8 weeks of treatment (n = 12 per group) (black circles, control; black triangles, glimepiride 0.1 mg/kg; white squares, empagliflozin 1 mg/kg; reverse white triangles, empagliflozin 3 mg/kg). (b) Changes in the mass and percentage of fat measured by DEXA after 8 weeks of treatment in each arm (black bars, control; white bars, glimepiride 0.1 mg/kg; light grey bars, empagliflozin 1 mg/kg; dark grey bars, empagliflozin 3 mg/kg). (c) Representative DEXA images from the four groups. Data are means ± SEM. *p < 0.05 vs control; † p < 0.05 vs glimepiride
Biochemical variables after glimepiride or empagliflozin treatment
In the ApoE −/− mice that were fed a western diet, fasting glucose levels were lower in the empagliflozin and glimepiride treatment groups than they were in the control group (Table 1). Fasting insulin levels were lower in the empagliflozin groups than in the glimepiride group. HOMA-IR, a surrogate marker of insulin resistance, was lower in the empagliflozin-treated groups than it was in the control group, which was consistent after adjusting for final body weight. In this study, ALT and AST levels were significantly lower in the empagliflozin groups compared with the control or glimepiride groups. There was no significant deterioration in kidney function (evaluated by creatinine level) in the glimepiride or empagliflozin groups, and no differences were found after comparing with the control group (Table 1). Triacylglycerol levels were significantly lower in the empagliflozin groups than those detected in the glimepiride or control groups. HDL-cholesterol levels were higher in the high-dose empagliflozin group (3 mg/kg) than in the control group.
Levels of high-sensitivity C-reactive protein (hsCRP) were significantly decreased in the empagliflozin groups (1 or 3 mg/kg) compared with the control group. The level of serum amyloid A (SAA) was also lower in empagliflozin-treated mice than in the control or glimepiride-treated mice. However, SAA level did not differ between the two empagliflozin groups. The circulating concentrations of TNF-α and IL-6 were decreased by empagliflozin treatment but not by control or glimepiride treatment. MCP-1 concentrations were also decreased significantly by empagliflozin treatment. There was a significant increase in the circulating adiponectin concentrations after empagliflozin treatment. The urinary microalbumin/creatinine ratio was lower in the empagliflozin group than in the control or glimepiride groups.
Correlation between atheroma area and markers of inflammation or insulin resistance
Atheroma volume was correlated positively with HOMA-IR and hsCRP and TNF-α levels and negatively with adiponectin levels (Fig. 5). There was a non-significant correlation between atheroma area and MCP-1 but not IL-6. Atheroma volume was also correlated significantly with urinary microalbuminuria (data not shown).

Correlation between atheroma volume and markers related to insulin resistance (a, d) or inflammation (b, c) (n = 12 per group) (grey circles, control; white squares, glimepiride 0.1 mg/kg; black triangles, empagliflozin 1 mg/kg; black diamonds, empagliflozin 3 mg/kg)
Area and size of lipid droplets in the liver
After 8 weeks of treatment, the liver of mice in the empagliflozin groups appeared to display a lesser degree of fatty changes compared with the control or glimepiride groups (Fig. 6a). The lipid droplet area (Fig. 6b) and size of lipid droplets (Fig. 6c) in the liver of empagliflozin-treated mice were smaller than those in the control or glimepiride groups.

(a) Liver histology revealed by H&E staining. All images were acquired at magnification ×200 (n = 12 per group). (b) The percentage area of lipid droplets in the liver (% of total area). (c) Lipid droplet size in the liver. Data are means ± SEM. *p < 0.05 vs control; † p < 0.05 vs glimepiride
Fat cell size and crown-like structures in adipose tissue
After 8 weeks of treatment, crown-like structures were found in the control and glimepiride groups but not in the empagliflozin groups (Fig. 7a). The fat cells in abdominal visceral adipose tissue of mice in the empagliflozin groups were smaller in size than those in the control or glimepiride groups (Fig. 7b).

(a) Histology of abdominal visceral adipose tissues stained by H&E (n = 12 per group) (scale bars, 20 μm). The arrows indicate crown-like structures. (b) Comparison of fat cell size (% of control), glimepiride and empagliflozin groups. Data are means ± SEM. *p < 0.05 vs control; † p < 0.05 vs glimepiride
Comparison of the mRNA expression of inflammatory markers and immunofluorescence staining of CD68 and CD11c in adipose tissue
Levels of Tnf, Il6 and Mcp-1 mRNA expression in abdominal visceral adipose tissue were lower in the empagliflozin groups than in the control or glimepiride groups (Fig. 8a). Immunofluorescence staining for inflammatory cells showed that empagliflozin treatment reduced the immunopositivity of CD68 compared with control or glimepiride treatment (Fig. 8b). In the M1-specific assays, M1 macrophage infiltration (CD11c) was lower in fat tissues of the empagliflozin groups compared with other groups (Fig. 8b).

(a) Expression of Tnf (white bars), Il6 (grey bars) and Mcp-1 (black bars) mRNA in visceral adipose tissue. Data are means ± SEM and are representative of five individual experiments. (*p < 0.05 vs control; † p < 0.05 vs glimepiride). (b) Immunohistochemical staining of CD68 and CD11c in visceral adipose tissue (scale bar, 20 μm). The arrows indicate merged cells
Expression of p-Akt and p-GSK in the liver and abdominal visceral fat
The relative expression of p-Akt vs Akt in the liver of mice in the empagliflozin groups was greater than that detected in the control group (Fig. 9a). The relative expression of p-GSK vs GSK in the liver of mice in the empagliflozin groups was greater than that observed for the glimepiride or control groups (Fig. 9b). The relative expression of p-Akt vs Akt and p-GSK vs GSK in the abdominal visceral fat was greater in the empagliflozin groups than it was in the glimepiride or control group and the relative expression of these enzymes in the glimepiride group was greater than that detected in the control group (Fig. 9c, d).

Western blot analysis of p-Akt and p-GSK in liver (a, b) and visceral fat tissues (c, d). Data are means ± SEM and are representative of five individual experiments. (*p < 0.05 vs control; † p < 0.05 vs glimepiride, ‡ p < 0.05 vs 1 mg/kg empagliflozin)
Safety of empagliflozin
Urinary-tract and genital infections have been reported after treatment with SGLT2 inhibitors [22]. The mice in our study showed no sign of significant infection, particularly of the genital areas.
Cell studies
Effect of empagliflozin treatment on the proliferation of RAoSMCs
To determine whether empagliflozin affects vascular smooth muscle cell proliferation in vitro, we measured incorporation of BrdU. As expected, treatment of RAoSMCs with platelet-derived growth factor (PDGF) significantly stimulated BrdU incorporation. However, empagliflozin treatment (0.1–100 μmol/l) had no effect on RAoSMC proliferation, suggesting the absence of a direct effect of empagliflozin on the proliferation of RAoSMCs (ESM Fig. 2).
Effect of empagliflozin on the PDGF-directed migration of RAoSMCs
To determine the effect of empagliflozin on RAoSMC migration, we performed a wound-healing assay on cell monolayers. Empagliflozin treatment did not inhibit the PDGF-directed migration of RAoSMCs (ESM Fig. 3).
Effect of empagliflozin treatment on the proliferation of HUVECs
Similar to the observations made in RAoSMCs, empagliflozin treatment (0.1–100 μmol/l) had no effect on vascular endothelial growth factor-stimulated HUVEC proliferation (ESM Fig. 4). This suggests a neutral effect of empagliflozin on endothelial cell proliferation.
Discussion
This study demonstrated that treatment with empagliflozin reduced atheromatous plaque formation in the aortic arch of an atherosclerosis-prone mouse model. Body weight and the amount and percentage of fat were also lower in the empagliflozin groups than in the control or glimepiride groups. Empagliflozin decreased fasting insulin levels and HOMA-IR, suggesting that insulin resistance was improved. Lipid profiles, liver enzyme activity and hsCRP and adiponectin levels were all improved by empagliflozin, suggesting it had a positive effect on insulin sensitivity and that it had an anti-inflammatory action.
It was reported that empagliflozin improved glucose tolerance and insulin sensitivity in db/db mice [23]. A study showed that dapagliflozin induced a significant reduction in glucose excursion in an OGTT [24]. Treatment with TS-071 (luseogliflozin), another recently developed SGLT2 inhibitor, improved glucose tolerance without stimulating insulin secretion in Zucker diabetic fatty rats [25]. In the present study, fasting insulin levels and HOMA-IR were decreased by empagliflozin treatment compared with control or glimepiride treatment. This finding corresponds to the results of a clinical study in which empagliflozin treatment decreased insulin secretion and tissue glucose disposal while improving insulin sensitivity in patients with type 2 diabetes [26]. Improvement of insulin sensitivity might be a main contributing factor to the cardiovascular protection afforded by empagliflozin.
Examination of the downstream molecules in the insulin signalling cascade showed that empagliflozin treatment increased the expression of p-Akt and p-GSK-3β in both the liver and visceral fat. Glimepiride treatment also increased the expression of p-Akt and p-GSK-3β in visceral fat but to a significantly lower extent than that induced by empagliflozin treatment. This finding demonstrates that the improvement of insulin signalling by empagliflozin was not induced by the amelioration of glucotoxicity, because the final plasma glucose levels detected in the treatment groups were comparable. Comparison between mice that lost weight and mice that maintained weight in the empagliflozin groups showed that Akt expression levels increased in both types of mouse. This suggests that improvement of Akt expression is independent of weight loss. In consequence, empagliflozin may improve insulin sensitivity via the modulation of the Akt–GSK-3β pathway.
In this study, empagliflozin treatment resulted in reduced fat mass and fat percentage compared with control or glimepiride treatment. Furthermore, fat cells in visceral adipose tissue were smaller in size in the empagliflozin-treated mice than in control or glimepiride-treated mice. Clinical studies have reported that SGLT2 inhibitors reduce body fat mass [11, 27]. In our study, the mRNA expression of Tnf, Il6 and Mcp-1 was significantly lower in the fat tissues of empagliflozin-treated mice when compared with control or glimepiride-treated mice. In the same fat tissue, immunopositivity for CD68, a marker of the various cells of the macrophage lineage, was lower in the empagliflozin group vs the control or glimepiride group. Furthermore, M1 macrophage infiltration (CD11c) was less pronounced in the fat tissue of mice treated with empagliflozin than in the other groups. M1 macrophages are typically induced by lipopolysaccharide or IFN-γ and are potent effector cells that produce pro-inflammatory cytokines [28]. These findings highlight the anti-inflammatory effect of empagliflozin on target tissues and suggest that inflammation of fat, a main driving factor of increased cardiovascular risk [29, 30], can be alleviated by empagliflozin.
In our study, empagliflozin decreased AST and ALT levels. The area and size of lipid droplets that accumulated in the liver was also smaller in the empagliflozin group than in the other groups. Alteration in lipid droplet size was associated with the expression of the perilipin gene, which is a key player in hepatic steatosis [31], although the total number of lipid droplets was not reported to differ [32]. Nonalcoholic fatty liver disease is one of the phenotypes of insulin resistance or metabolic syndrome. There is substantial evidence to show that fat accumulation in the liver is associated with type 2 diabetes and cardiovascular disease [33, 34].
In this study, empagliflozin treatment reduced the circulating levels of hsCRP, TNF-α, IL-6 and MCP-1. The inflammation process is involved in the initiation and progression of atherosclerosis [35]. C-reactive protein has been reported to promote atherosclerosis by directly increasing the transcytosis of LDL across vascular endothelial cells and increasing LDL retention in vitro and in vivo [36]. MCP-1 has been also reported to play an important role in the pathogenesis of insulin resistance, thrombogenicity and atherosclerosis [35, 37]. Several lines of evidence suggest that monocytes containing MCPs and macrophages influence the growth of other cell types within atherosclerotic lesions [38]. A study in db/db mice demonstrated that long-term treatment with empagliflozin suppressed pancreatic islet cell inflammation, as well as inflammation in the liver and white adipose tissues [23]. This anti-inflammatory effect was concomitant with a reduction in the levels of major inflammatory and immune factors, a finding that was similar to that in our own study.
Decreased levels of circulating adiponectin may act as a mediator of the pathophysiology of obesity, type 2 diabetes and atherosclerosis [39, 40]. In a different context, several studies have reported the cardioprotective effect of glucagon-like peptide-1 (GLP-1) treatment or increased GLP-1 levels in the setting of ischaemic injury or heart failure [41, 42]. Thus, increased levels of adiponectin and GLP-1 after empagliflozin treatment seem to be a mechanism by which empagliflozin mitigates the progression of atherosclerosis [43].
Chronic treatment with SGLT2 inhibitors shifts substrate utilisation from carbohydrates to lipids [26]. The serum levels of triacylglycerol decreased to a greater extent in the empagliflozin-treated mice compared with control or glimepiride-treated mice. High-dose empagliflozin treatment also increased HDL-cholesterol levels significantly compared with control or glimepiride treatment. These favourable changes may contribute to a decrease in plaque formation, as shown in clinical studies [44, 45]. Interestingly, clinical studies with SGLT2 inhibitors have reported inconsistent results for LDL-cholesterol level [14, 46]. In our study, empagliflozin treatment caused a non-significant decrease in LDL-cholesterol level. An animal study reported reduced intestinal cholesterol absorption after empagliflozin treatment [46]. At present, we cannot conclude whether SGLT2 inhibitors increase LDL-cholesterol or whether effects of SGLT2 inhibitors on LDL-cholesterol level affect the development of atherosclerosis.
In this study, glimepiride was selected as the active control. Glimepiride was used as a reasonable control drug in the PERISCOPE study, which investigated the anti-atherosclerotic effect of pioglitazone [47]. Another clinical study reported that glimepiride treatment did not affect vascular endothelial function, despite significant reduction in blood glucose level [48]. In contrast, metformin attenuated angiotensin-II-induced atheromatous plaque formation in ApoE −/− mice [49]. In terms of clinical relevance, the current diabetes guidelines state that a sulfonylurea is more suitable for comparison with empagliflozin (or another SGLT-2 inhibitor) as a second-line drug after metformin treatment.
Human and rodent studies have shown that SGLT2 inhibitors lower blood pressure by inducing osmotic diuresis and natriuresis [14, 50]. Thus, although we did not measure blood pressure in our study, a reduction in blood pressure after empagliflozin treatment may have contributed to the reduction in albuminuria and plaque formation. To evaluate insulin resistance and beta cell function, we used HOMA-IR and HOMA-β instead of more sophisticated methods. Although they have not been validated in animal studies as in human studies, these variables are frequently used in rodent studies [21, 51].
The present study, focussing on the effect of empagliflozin regarding cardiovascular protection, suggests possible underlying mechanisms to explain the superior results of empagliflozin vs placebo obtained in the EMPA-REG study [14]. Although more detailed mechanistic studies are needed, our study suggests that the improvement in insulin resistance and alleviation of the inflammatory process in the circulation, liver and fat tissues are attributable to the anti-atherosclerotic property of empagliflozin, in addition to its induction of weight loss, decrease in triacylglycerol/HDL-cholesterol ratio and reduction of microalbuminuria. Compositional changes in atheromatous plaques, such as reduced lipid droplet area and reduced infiltration of inflammatory cells, were also found in the empagliflozin groups.
In conclusion, empagliflozin seems to be effective against the macrovascular complications of diabetes in addition to its known glucose-lowering property.
Abbreviations
- ALT:
-
Alanine aminotransferase
- AST:
-
Aspartate aminotransferase
- BrdU:
-
Bromodeoxyuridine
- DEXA:
-
Dual-energy x-ray absorptiometry
- GLP-1:
-
Glucagon-like peptide-1
- H&E:
-
Haematoxylin and Eosin
- hsCRP:
-
High-sensitivity C-reactive protein
- HOMA-β:
-
HOMA of beta cell function
- HUVEC:
-
Human umbilical vein endothelial cell
- MCP-1:
-
Monocyte chemoattractant protein-1
- PDGF:
-
Platelet-derived growth factor
- RAoSMC:
-
Rat aortic smooth muscle cell
- RT:
-
Room temperature
- SGLT2:
-
Sodium/glucose cotransporter 2
References
Kanai Y, Lee WS, You G, Brown D, Hediger MA (1994) The human kidney low affinity Na+/glucose cotransporter SGLT2. Delineation of the major renal reabsorptive mechanism for D-glucose. J Clin Invest 93:397–404
Ehrenkranz JR, Lewis NG, Kahn CR, Roth J (2005) Phlorizin: a review. Diabetes Metab Res Rev 21:31–38
Kahn BB, Shulman GI, DeFronzo RA, Cushman SW, Rossetti L (1991) Normalization of blood glucose in diabetic rats with phlorizin treatment reverses insulin-resistant glucose transport in adipose cells without restoring glucose transporter gene expression. J Clin Invest 87:561–570
Chasis H, Jolliffe N, Smith HW (1933) The action of phlorizin on the excretion of glucose, xylose, sucrose, creatinine and urea by man. J Clin Invest 12:1083–1090
Vallon V (2015) The mechanisms and therapeutic potential of SGLT2 inhibitors in diabetes mellitus. Annu Rev Med 66:255–270
Idris I, Donnelly R (2009) Sodium-glucose co-transporter-2 inhibitors: an emerging new class of oral antidiabetic drug. Diabetes Obes Metab 11:79–88
Ferrannini E, Seman L, Seewaldt-Becker E, Hantel S, Pinnetti S, Woerle HJ (2013) A Phase IIb, randomized, placebo-controlled study of the SGLT2 inhibitor empagliflozin in patients with type 2 diabetes. Diabetes Obes Metab 15:721–728
Goring S, Hawkins N, Wygant G et al (2014) Dapagliflozin compared with other oral anti-diabetes treatments when added to metformin monotherapy: a systematic review and network meta-analysis. Diabetes Obes Metab 16:433–442
Stenlof K, Cefalu WT, Kim KA et al (2013) Efficacy and safety of canagliflozin monotherapy in subjects with type 2 diabetes mellitus inadequately controlled with diet and exercise. Diabetes Obes Metab 15:372–382
Liakos A, Karagiannis T, Athanasiadou E et al (2014) Efficacy and safety of empagliflozin for type 2 diabetes: a systematic review and meta-analysis. Diabetes Obes Metab 16:984–993
Bolinder J, Ljunggren O, Kullberg J et al (2012) Effects of dapagliflozin on body weight, total fat mass, and regional adipose tissue distribution in patients with type 2 diabetes mellitus with inadequate glycemic control on metformin. J Clin Endocrinol Metab 97:1020–1031
List JF, Woo V, Morales E, Tang W, Fiedorek FT (2009) Sodium-glucose cotransport inhibition with dapagliflozin in type 2 diabetes. Diabetes Care 32:650–657
Perkins BA, Cherney DZ, Partridge H et al (2014) Sodium-glucose cotransporter 2 inhibition and glycemic control in type 1 diabetes: results of an 8-week open-label proof-of-concept trial. Diabetes Care 37:1480–1483
Zinman B, Wanner C, Lachin JM et al (2015) Empagliflozin, cardiovascular outcomes, and mortality in type 2 diabetes. N Engl J Med 373:2117–2128
Terasaki M, Hiromura M, Mori Y et al (2015) Amelioration of hyperglycemia with a sodium-glucose cotransporter 2 inhibitor prevents macrophage-driven atherosclerosis through macrophage foam cell formation suppression in type 1 and type 2 diabetic mice. PLoS One 10, e0143396
Nakajima K, Mita T, Osonoi Y et al (2015) Effect of repetitive glucose spike and hypoglycaemia on atherosclerosis and death rate in Apo E-deficient mice. Int J Endocrinol 2015:406394
Wu KK, Huan Y (2007) Diabetic atherosclerosis mouse models. Atherosclerosis 191:241–249
Lim S, Lee KS, Lee JE et al (2015) Effect of a new PPAR-gamma agonist, lobeglitazone, on neointimal formation after balloon injury in rats and the development of atherosclerosis. Atherosclerosis 243:107–119
Matthews DR, Hosker JP, Rudenski AS et al (1985) Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 28:412–419
Stringer DM, Zahradka P, Declercq VC et al (2010) Modulation of lipid droplet size and lipid droplet proteins by trans-10, cis-12 conjugated linoleic acid parallels improvements in hepatic steatosis in obese, insulin-resistant rats. Biochim Biophys Acta 1801:1375–1385
Lim S, Moon MK, Shin H et al (2011) Effect of S-adenosylmethionine on neointimal formation after balloon injury in obese diabetic rats. Cardiovasc Res 90:383–393
Vasilakou D, Karagiannis T, Athanasiadou E et al (2013) Sodium-glucose cotransporter 2 inhibitors for type 2 diabetes: a systematic review and meta-analysis. Ann Intern Med 159:262–274
Chen L, Klein T, Leung PS (2012) Effects of combining linagliptin treatment with BI-38335, a novel SGLT2 inhibitor, on pancreatic islet function and inflammation in db/db mice. Curr Mol Med 12:995–1004
Komoroski B, Vachharajani N, Feng Y, Li L, Kornhauser D, Pfister M (2009) Dapagliflozin, a novel, selective SGLT2 inhibitor, improved glycemic control over 2 weeks in patients with type 2 diabetes mellitus. Clin Pharmacol Ther 85:513–519
Yamamoto K, Uchida S, Kitano K et al (2011) TS-071 is a novel, potent and selective renal sodium-glucose cotransporter 2 (SGLT2) inhibitor with anti-hyperglycaemic activity. Br J Pharmacol 164:181–191
Ferrannini E, Muscelli E, Frascerra S et al (2014) Metabolic response to sodium-glucose cotransporter 2 inhibition in type 2 diabetic patients. J Clin Invest 124:499–508
Ridderstrale M, Andersen KR, Zeller C, Kim G, Woerle HJ, Broedl UC (2014) Comparison of empagliflozin and glimepiride as add-on to metformin in patients with type 2 diabetes: a 104-week randomised, active-controlled, double-blind, phase 3 trial. Lancet Diabetes Endocrinol 2:691–700
Gordon S, Taylor PR (2005) Monocyte and macrophage heterogeneity. Nat Rev Immunol 5:953–964
Apovian CM, Bigornia S, Mott M et al (2008) Adipose macrophage infiltration is associated with insulin resistance and vascular endothelial dysfunction in obese subjects. Arterioscler Thromb Vasc Biol 28:1654–1659
Farb MG, Bigornia S, Mott M et al (2011) Reduced adipose tissue inflammation represents an intermediate cardiometabolic phenotype in obesity. J Am Coll Cardiol 58:232–237
Carr RM, Ahima RS (2016) Pathophysiology of lipid droplet proteins in liver diseases. Exp Cell Res 340:187–192
Hesse D, Radloff K, Jaschke A et al (2014) Hepatic trans-Golgi action coordinated by the GTPase ARFRP1 is crucial for lipoprotein lipidation and assembly. J Lipid Res 55:41–52
Cho NH, Jang HC, Choi SH et al (2007) Abnormal liver function test predicts type 2 diabetes: a community-based prospective study. Diabetes Care 30:2566–2568
Lim S, Oh TJ, Koh KK (2015) Mechanistic link between nonalcoholic fatty liver disease and cardiometabolic disorders. Int J Cardiol 201:408–414
Freitas Lima LC, Braga VA, do Socorro de Franca Silva M et al (2015) Adipokines, diabetes and atherosclerosis: an inflammatory association. Front Physiol 6:304
Bian F, Yang X, Zhou F et al (2014) C-reactive protein promotes atherosclerosis by increasing LDL transcytosis across endothelial cells. Br J Pharmacol 171:2671–2684
Uchida Y, Takeshita K, Yamamoto K et al (2012) Stress augments insulin resistance and prothrombotic state: role of visceral adipose-derived monocyte chemoattractant protein-1. Diabetes 61:1552–1561
Lin J, Kakkar V, Lu X (2014) Impact of MCP-1 in atherosclerosis. Curr Pharm Des 20:4580–4588
Funahashi T, Nakamura T, Shimomura I et al (1999) Role of adipocytokines on the pathogenesis of atherosclerosis in visceral obesity. Intern Med 38:202–206
Kern PA, Di Gregorio GB, Lu T, Rassouli N, Ranganathan G (2003) Adiponectin expression from human adipose tissue: relation to obesity, insulin resistance, and tumor necrosis factor-α expression. Diabetes 52:1779–1785
Liu Q, Anderson C, Broyde A et al (2010) Glucagon-like peptide-1 and the exenatide analogue AC3174 improve cardiac function, cardiac remodeling, and survival in rats with chronic heart failure. Cardiovasc Diabetol 9:76
Goodwill AG, Mather KJ, Conteh AM, Sassoon DJ, Noblet JN, Tune JD (2014) Cardiovascular and hemodynamic effects of glucagon-like peptide-1. Rev Endocr Metab Disord 15:209–217
Lim S, Quon MJ, Koh KK (2014) Modulation of adiponectin as a potential therapeutic strategy. Atherosclerosis 233:721–728
Salazar MR, Carbajal HA, Espeche WG, Aizpurua M, Maciel PM, Reaven GM (2014) Identification of cardiometabolic risk: visceral adiposity index versus triglyceride/HDL cholesterol ratio. Am J Med 127:152–157
Wang D, Liu B, Tao W, Hao Z, Liu M (2015) Fibrates for secondary prevention of cardiovascular disease and stroke. Cochrane Database Syst Rev 10, CD009580
Briand F, Mayoux E, Brousseau E et al (2016) Empagliflozin, via switching metabolism toward lipid utilization, moderately increases LDL cholesterol levels through reduced LDL catabolism. Diabetes 65:2032–2038
Mazzone T, Meyer PM, Feinstein SB et al (2006) Effect of pioglitazone compared with glimepiride on carotid intima-media thickness in type 2 diabetes: a randomized trial. JAMA 296:2572–2581
Papathanassiou K, Naka KK, Kazakos N et al (2009) Pioglitazone vs glimepiride: differential effects on vascular endothelial function in patients with type 2 diabetes. Atherosclerosis 205:221–226
Vasamsetti SB, Karnewar S, Kanugula AK, Thatipalli AR, Kumar JM, Kotamraju S (2015) Metformin inhibits monocyte-to-macrophage differentiation via AMPK-mediated inhibition of STAT3 activation: potential role in atherosclerosis. Diabetes 64:2028–2041
Bailey CJ, Gross JL, Pieters A, Bastien A, List JF (2010) Effect of dapagliflozin in patients with type 2 diabetes who have inadequate glycaemic control with metformin: a randomised, double-blind, placebo-controlled trial. Lancet 375:2223–2233
Lim S, Choi SH, Shin H et al (2012) Effect of a dipeptidyl peptidase-IV inhibitor, des-fluoro-sitagliptin, on neointimal formation after balloon injury in rats. PLoS One 7, e35007
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This work was supported by research grants from the Korea Diabetes Association (06-2014-260) and Boehringer Ingelheim (06-2014-168). The funding agencies had no role in the study design, data collection or analysis, decision to publish or preparation of the manuscript. The sole responsibility for the content of this paper lies with the authors.
Duality of interest
The authors declare that there is no duality of interest associated with this manuscript.
Contribution statement
JHH, TJO and SL researched data and contributed to the experimental design and discussion. GL, HJM, DHL, KMK, SHC, HCJ, HSL, KSP and Y-BK researched data and contributed to the discussion. TJO and SL drafted the manuscript. All authors edited and revised the manuscript and approved the final version. SL is responsible for the integrity of the work as a whole.
Data availability
All relevant data were included in the article and/or the ESM files.
Additional information
Ji Hye Han and Tae Jung Oh contributed equally to this study.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM
(PDF 517 kb)
Rights and permissions
About this article

Cite this article
Han, J.H., Oh, T.J., Lee, G. et al. The beneficial effects of empagliflozin, an SGLT2 inhibitor, on atherosclerosis in ApoE −/− mice fed a western diet. Diabetologia 60, 364–376 (2017). https://doi.org/10.1007/s00125-016-4158-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-016-4158-2