
Abstract
Aims/hypothesis
FTO gene single nucleotide polymorphisms (SNPs) have been shown to be associated with obesity-related traits and type 2 diabetes. Several small studies have suggested a greater than expected effect of the FTO rs9939609 SNP on weight in polycystic ovary syndrome (PCOS). We therefore aimed to examine the impact of FTO genotype on BMI and weight in PCOS.
Methods
A systematic search of medical databases (PubMed, EMBASE and Cochrane CENTRAL) was conducted up to the end of April 2011. Seven studies describing eight distinct PCOS cohorts were retrieved; seven were genotyped for SNP rs9939609 and one for SNP rs1421085. The per allele effect on BMI and body weight increase was calculated and subjected to meta-analysis.
Results
A total of 2,548 women with PCOS were included in the study; 762 were TT homozygotes, 1,253 had an AT/CT genotype, and 533 were AA/CC homozygotes. Each additional copy of the effect allele (A/C) increased the BMI by a mean of 0.19 z score units (95% CI 0.13, 0.24; p = 2.26 × 10−11) and body weight by a mean of 0.20 z score units (95% CI 0.14, 0.26; p = 1.02 × 10−10). This translated into an approximately 3.3 kg/m2 increase in BMI and an approximately 9.6 kg gain in body weight between TT and AA/CC homozygotes. The association between FTO genotypes and BMI was stronger in the cohorts with PCOS than in the general female populations from large genome-wide association studies. Deviation from an additive genetic model was observed in heavier populations.
Conclusions/interpretation
The effect of FTO SNPs on obesity-related traits in PCOS seems to be more than two times greater than the effect found in large population-based studies. This suggests an interaction between FTO and the metabolic context or polygenic background of PCOS.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Obesity has reached epidemic proportions. In 2008, it was estimated that 1.46 billion adults worldwide were overweight, including 500 million obese individuals [1]. Both environmental and genetic factors contribute to this phenomenon. Several genes associated with obesity have recently been identified by large scale genome-wide association studies, one being the FTO gene located on chromosome 16q12.2 [2]. The FTO rs9939609 single nucleotide polymorphism (SNP) and some other variants have been associated with increased weight, raised BMI and several related traits [3–7]. This SNP has also been associated with higher odds of type 2 diabetes, although this effect can be entirely explained by its impact on BMI in affected and unaffected individuals [3, 8–11]. This initial finding in European white cohorts has been confirmed for other ethnicities [12], including Mexican [13] and Asian [14–16] as well as African populations [17]. The FTO rs9939609 SNP has two alleles, T and A, the latter being a variant associated with risk of obesity [18]. It has been demonstrated in multiple cohorts, mostly of white individuals, that AA homozygotic individuals are on average 3 kg heavier, with an increased BMI of about 0.8 kg/m2, than those without the risk allele [3].
Polycystic ovary syndrome (PCOS) is a common endocrine disorder affecting 6–12% of women of reproductive age, who present with hyperandrogenism, ovulatory dysfunction and polycystic ovaries [19–21]. Women with PCOS are frequently obese, which significantly affects the syndrome's clinical features by increasing the risk of insulin resistance, type 2 diabetes and cardiovascular disease [22]. A study of the Polish population recently demonstrated a substantially larger impact of the FTO rs9939609 polymorphism on weight in women with PCOS compared with the previously reported weight difference in the general population [23]. In this study, a 10 kg weight difference was found between AA and TT homozygote PCOS carriers. In addition, four other studies have reported a greater than expected effect of the FTO gene on weight in cohorts with PCOS [24–27]. However, all of these studies have been characterised by a limited sample size and statistical power. Furthermore, none of the CIs reported in the individual PCOS population studies excluded the 3 kg difference found in much larger studies of the general population.
Our study aims to perform a systematic review and meta-analysis to fully define the effect of the FTO gene variants on BMI and weight in PCOS women. Specifically, we intended to determine whether the FTO rs9939609 SNP and variants in linkage disequilibrium (LD) have a greater impact on these traits in individuals with PCOS than in the general population. A meta-analysis evaluating the relationship between FTO and PCOS has never been conducted and may provide new insight into this genetic association.
Methods
Search strategy
Relevant studies were identified by a systematic review of the literature via a search of the major medical databases (PubMed, EMBASE and Cochrane CENTRAL). The search strategy consisted of the following key words: FTO, rs9939609, fat mass and obesity associated protein, PCOS, polycystic, sclerocystic. Terms representing outcome measures were omitted in order to increase the sensitivity of the strategy and to elucidate all studies on a given population regardless of their design and methodology. This strategy was therefore considerably more sensitive than the common strategies used in systematic reviews. Databases were searched up to the end of April 2011 by two separate reviewers. References of retrieved articles and abstracts of presentations at international meetings (American Diabetes Association, European Association for the Study of Diabetes) were screened for the most up-to-date studies.
Inclusion and exclusion criteria
All types of study investigating women with PCOS were considered relevant, regardless of their design and hypothesis, provided that patients were genotyped for the FTO rs9939609 SNP or for other SNPs in strong LD. The authors of the identified publications were contacted in order to confirm the relevance of each study and were invited to collaborate. Detailed data of at least BMI or body weight had to be accessible for each patient. Unpublished raw data on which to perform the analyses were received from all of the authors except Tan et al [27], who performed their own calculations (using the same statistical methods) and submitted aggregated data (mean values with SDs for each group representing different genotypes). This procedure was associated with restriction of the local Bioethical Committee from external access to the raw data. Studies were excluded if patients were not genotyped for the rs9939609 polymorphism or any other SNP in LD, and if the authors declined or did not respond to the invitation to collaborate.
Outcome measures
Per allele change in effect on BMI and body weight were assessed.
Statistical analysis
Distributions of BMI were skewed in most cohorts; therefore, log10 transformation was performed in order to obtain a normal distribution. It was expected that the distribution of BMI in women with PCOS would differ from that of the general population; hence, data were standardised to the z score units of SD. z scores were obtained by dividing the difference between each logBMI value and the mean logBMI of the entire cohort by the SD of the logBMI values. The same transformation and standardisation was applied to body weight. Robust linear regression in the additive model was performed in order to test the association between genotypes and anthropometric measures. Individual study results were then pooled in a meta-analysis with a fixed inverse variance effect model. The overall effect was tested with the Z test assuming p < 0.05 as the level of significance. All effect sizes were presented as means with a 95% CI. Heterogeneity between studies was assessed by applying Cochran χ2 test and was considered significant at p < 0.1. Meta-analyses were carried out using Sophie ver. 1.5 Software developed by HTA Consulting (Krakow, Poland). Publication bias was assessed for the difference in BMI between the AA and TT genotypes with the use of both the Egger and Begg tests, with p < 0.05 considered statistically significant. Hardy–Weinberg equilibrium (HWE) was tested using the DeFinetti program (available from http://ihg.gsf.de/cgi-bin/hw/hwa1.pl, accessed 24 February 2012).
In addition, in order to allow a comparison with one of the external control cohorts, an association between the BMI and the FTO variants was also calculated after rank-based inverse normal transformation of the BMI values. Data regarding BMI were also adjusted for patients' ages by including the variable as a covariate in a linear regression model in order to exclude a potential confounding effect of this trait. To provide an approximate estimate of the per allele effect size in BMI units (kg/m2), the aggregated effect sizes in z score units were translated into BMI units using SD values from the entire PCOS population, yielding 8.619 kg/m2. The same procedure was applied to body weight, in which z score units were multiplied by the SD from the entire PCOS population, yielding 24.065 kg.
The effect of the FTO mutations on BMI in the subset of women with PCOS was statistically compared with the results obtained from genome-wide association studies. First, the data were compared with unpublished data provided by the GIANT Consortium, which investigated an association between rs1558902 FTO polymorphisms and BMI increase expressed in SD units. The available data from 80 studies included 109,955 adult women of European ancestry with characteristics that had previously been described [6]. These data were considered relevant as rs1558902 is localised to intron 1 and is in complete LD with rs9939609 (r 2 = 0.934 and D' = 0.966 according to HapMap release 22). Another reference source came from the published data from Frayling et al, who investigated 11 cohorts, including over 18,800 adult women [3]. Their findings reported an increase in BMI associated with the rs9939609 polymorphism, expressed as a per A-allele increase in z score units. Both reference results were unadjusted for age. Results from the entire female subpopulation were compared with those from the population with PCOS. Due to limited precision of the data published by Frayling et al, a conservative worst case scenario approach was adopted by using the highest possible mean value and SE that still fitted the common rule for rounding numerical values. Both the Z test and the χ2 test for heterogeneity were used to compare effect size in the population with PCOS for each reference group. However, owing to the consistent results of both statistical approaches, only one common p value was presented for each comparison [28].
To determine the genetic model exhibiting the best fit, a correlation between genotype and phenotype expressed in per allele increase in logBMI z score units was assessed using linear regression [29]. Ordinal variables were assigned to each genotype according to the number of A/C alleles (i.e. TT = 0, AT/AC = 1, AA/CC = 2). Additive, recessive and dominant models were tested, and the model with the lowest Akaike's information criterion (AIC) was considered best fitting. Meta-regression using the restricted maximum likelihood model was performed in order to explain deviation from the additive model.
Results
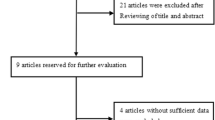
Twenty-nine records were identified in the systematic search of the medical databases, of which 14 were considered potentially relevant and underwent further assessment after exclusion of duplicates and abstract analysis. Six papers were subsequently excluded due to reasons presented in the PRISMA diagram (Fig. 1). The eight remaining studies were initially included, and their authors were contacted in order to confirm eligibility. The authors of seven studies, which included data regarding eight distinct cohorts, met the inclusion criteria and agreed to collaborate in this study. One study of the Chinese population presented data relevant to our hypothesis; however, since the authors did not respond to an invitation to participate and to provide data, this study was excluded [30].

PRISMA diagram of the study selection process
Study characteristics
The studies included were conducted in six European countries (Austria, Czech Republic, Germany, Poland, Romania, UK) and in the USA [23–27, 31, 32]. All of the study participants from the European centres were white, whereas in the American cohort participants were grouped as white 91%, Hispanic 3% and African–American 1%, with 5% categorised as unknown or other. The characteristics of the populations investigated are shown in Table 1. Overall, white participants represented 97% of the entire group. In six studies, patients were genotyped for the FTO rs9939609 SNP and in only one study for rs1421085 [32], which is in complete LD with rs9939609 (r 2 = 0.934 and D' = 0.966 according to HapMap release 22). The study size ranged from 136 to 469, with a total of 2,548 women with PCOS included in the analysis for whom at least BMI or body weight was available. Of these, 762 were TT homozygotic patients, 1,253 had an AT/CT genotype, and 533 were AA/CC homozygotic for rs9939609 and rs1421085. The distribution of the genotype was generally consistent across the cohorts, and the observed A/C allele frequency ranged from 42% to 52%. Genotype distribution of all cohorts was consistent with HWE.
The mean age of the population with PCOS ranged from 25 to 32 years old. Mean BMI ranged from 26.44 to 35.37 kg/m2, with patients weighing least in Austria and the heaviest study cohort originating from the USA. Mean weight ranged from 73.01 to 95.76 kg. Data regarding body weight were not available for one study [32].
Anthropometric traits
Data regarding BMI were available for a total of 2,510 women with PCOS. Of these, 746 were TT homozygotes, 1,239 women were AT/CT heterozygotes, and 525 patients had an AA/CC genotype. A strong association between the FTO variants and BMI was revealed. Each additional copy of the effect allele increased BMI by a mean of 0.19 z score units (95% CI 0.13, 0.24; p = 2.26 × 10−11), equivalent to approximately 1.62 kg/m2 (Fig. 2). No heterogeneity between the studies was noted (p = 0.9217, I 2 = 0%). This association remained almost unaffected even after data were adjusted for age (0.18 z score units, 95% CI 0.12, 0.23; p = 1.30 × 10−10, equivalent to 1.55 kg/m2).

Inverse variance (IV) meta-analysis of per allele (A/C) effect increase in log-transformed BMI, expressed in z score units. Forest plot and corresponding values represent point estimates for per allele effect together with 95% CI. Pooled weighted mean difference effect size with 95% CI is also shown. Each cohort is represented by the name of the first author. Test for heterogeneity: Q = 2.57, df = 7, p = 0.9217, I 2 = 0.00%. Test overall effect: Z = 6.69, p < 0.0001
Data regarding body weight were available for a total of 2,161 individuals with PCOS, including 648 women in the TT group, 1,068 women with an AT genotype, and 445 patients with an AA genotype. Meta-analysis of the seven cohorts revealed that, in the women with PCOS, each additional copy of the effect allele was associated with a body weight increase of a mean of 0.20 z score units (95% CI 0.14, 0.26; p = 1.02 × 10−10), which is equivalent to approximately 4.79 kg (Fig. 3). This effect was highly consistent across the studies and no evidence of heterogeneity was observed (p = 0.9388, I 2 = 0%).

Inverse variance (IV) meta-analysis of per allele (A) effect increase in log-transformed body weight, expressed in z score units. Forest plot and corresponding values represent point estimates for per allele effect together with 95% CI. Pooled weighted effect size with 95% CI is also shown. Each cohort is represented by the name of the first author. Test for heterogeneity: Q = 1.78, df = 6, p = 0.9388, I 2 = 0.00%. Test overall effect: z = 6.46, p < 0.0001
Comparison between PCOS cohorts and the general female population
The observed FTO-associated increase in BMI in women with PCOS was larger than the corresponding effect found in the 109,955 women from the GIANT Consortium cohort (p = 0.0002) (Fig. 4a) [6]. The association between examined polymorphisms and BMI in women with PCOS was also stronger than previously reported by Frayling et al [3] for the general female population (p = 0.0146) (Fig. 4b). When PCOS data were corrected for age, the impact of FTO on BMI remained significantly greater when comparing the effect sizes of both the GIANT Consortium cohort (p = 0.0005) and the female cohort described by Frayling et al [3] (p = 0.0298).

Comparison of the effect of the FTO polymorphism on BMI between women with PCOS and the general female population from the GIANT Consortium (a) and the study of Frayling et al [3] (b). White columns represent the per allele effect increase in the PCOS population, whereas grey columns represent the corresponding effect calculated for the general population. Data are expressed as either rank-based inverse normally transformed BMI (a) or log-transformed BMI z scores (b). Error bars represent SE. Comparison between cohorts was performed with a two-sided t test. † p = 0.0002 for comparison between women with PCOS and the general female population from the GIANT Consortium. *p = 0.0146 for comparison between women with PCOS and the general female population described by Frayling et al [3]
Genetic model analysis
Analysis of inheritance revealed that the study populations fitted different genetic models. The data from four trials [23, 25, 26, 31] fit very well with an additive model, whereas the recessive model was the best fit for the remaining populations [24, 27, 32] (Table 2). The degree of the deviation from an additive model was clearly associated with the mean BMI value in the TT subgroup, which was shown by meta-regression (p = 0.0033) (Fig. 5a). A single copy of the effect allele was associated with a significant increase in logBMI z score in only one trial, with the lowest mean BMI in the homozygous TT group [25]. A phenotypic manifestation of the heterozygotic FTO variant was also observed in three subsequent studies, with the lowest mean BMI in the TT group; however, the difference did not reach statistical significance, probably because of low power [23, 26, 31]. In the four remaining trials with a higher mean BMI in the TT group (≥28 kg/m2), a single copy of the effect allele did not cause any apparent gain in BMI (Fig. 5a). The association between the phenotypic manifestation of the heterozygous FTO polymorphism and the extent of obesity in each cohort was also replicated with respect to change in body weight z score units (p = 0.0041) (Fig. 5b). Conversely, phenotypic manifestation of homozygous AA/CC variants was independent of mean BMI or mean body weight in the TT group (data not shown).

Mixed-effect meta-regression analysis correlating the mean difference in (a) log-transformed BMI z score units between AT/CT and TT (dependent variable) and mean BMI in the TT group as independent variable or (b) log-transformed body weight z score units between AT and TT (dependent variable) and mean body weight in the TT group (independent variable). The solid lines indicate the predicted effects with 95% CI (dashed lines). Circles represent point estimates for each cohort. Study weight is represented by the diameter of the corresponding circle. Error bars represent the SE of each point estimate. Dotted lines represent a lack of phenotypic difference between the TT and AT/CT polymorphism. There was a statistically significant inverse correlation between mean BMI in the TT group and the AT-associated increase in logBMI (β = −0.0381, p = 0.0033). Similarly, a significant inverse correlation was observed between mean body weight in the TT group and the AT-associated increase in log-body weight (β = −0.0161, p = 0.0041)
Publication bias
No apparent publication bias was found by either visual inspection of the funnel plot or formal statistical tests: Egger's regression test (p = 0.217) or Begg's rank correlation test (Kendall's tau = 0.357, p = 0.275).
Discussion
Recent investigations have shed light on the genetic mechanism of adiposity and related traits. Several large studies have demonstrated an association between FTO gene polymorphisms and weight, BMI and related traits [3, 6, 7]. Moreover, the FTO rs9939609 SNP was also associated with PCOS, and its effect was mediated, as in the general population, by adiposity [26]. Interestingly, it was postulated in the within-patient analysis that the impact of this particular genetic variant could be larger in individuals with PCOS than in the general population [23]. A greater effect of rs9939609 SNP variants on weight-related variables in women with PCOS was also observed in several other studies [24–27]. However, none of these studies was adequately powered to confirm that the greater effect in women with PCOS was significantly different from that in the general population.
In this study, a meta-analysis demonstrated that each additional copy of the effect allele (A/C) was associated with an average increase in BMI of 0.19 z score units and a mean weight gain of 0.20 z score units. This corresponded to around a 3.3 kg/m2 and a 9.6 kg difference in BMI and body weight units, respectively, between TT and AA/CC homozygotes. Importantly, this effect was highly consistent between the PCOS studies included in this meta-analysis, with no evidence of heterogeneity between the analysed populations. The impact of FTO variants on BMI and weight is significantly stronger than those observed for the general female population as reported in the previous studies of FTO conducted on adult participants not selected for PCOS [3, 6, 15, 33–35].
Unexpectedly, the analysis revealed a variation between the studies with regard to the inheritance model of the FTO polymorphisms, which seemed to be related to the BMI in the investigated populations. Phenotypic manifestation of the heterozygous FTO variant existed only in studies with the lowest mean BMI in the TT group. In trials with a higher mean BMI in the TT group, a single variant of an effect allele was not associated with an apparent increase in obesity indices. For those studies, the recessive model fits best. To our knowledge, this phenomenon has not been observed in the general population.
The current study has several possible limitations. First, our PCOS cohorts with a mean age ranging from 25 to 32 years old were younger than those reported in the general female population studies. Effect sizes adjusted for age were not available for either reference group, either as unpublished data or from earlier publications [3, 6]. Nevertheless, the age adjustment in women with PCOS had a very limited impact on both the observed association between FTO and BMI and the results of comparisons between the patients with PCOS and the women in the general population, which remained almost unchanged. Moreover, the FTO-associated increase in BMI reported by Frayling et al [3] was comparable in the young and older cohorts, with no evidence of between-group heterogeneity. The per allele increase in logBMI z scores in the young female ALSPAC Mothers cohort (N = 6,376; mean age 28.4 years) and the NFBC1996 cohort (N = 2,306; each patient sampled at 31 years old) was not significantly different from the effect size observed in elderly women from the BWHS cohort (N = 3,244; mean age 68.8 years) as shown by the heterogeneity test (p = 0.79; I 2 = 0%, for all three cohorts) [3]. It thus seems unlikely that the effect of FTO variants on BMI in PCOS is due to the young age of the women examined.
Another possible limitation includes a lack of direct comparison with control cohorts consisting of women without PCOS matched with the analysed study populations. Such comparisons were either not made in the original studies [23, 25, 27] or were based on a very limited sample size, precluding quantitative analysis [24]. Nevertheless, we compared our study data with results, published and unpublished, from two large cohorts representing the general female population (GIANT Consortium, unpublished data) [3]. This analysis showed a statistically larger effect of the FTO variants in PCOS than in the reference groups [3, 6]. Second, although the possibility of a publication bias in reports examining the impact of the FTO gene in PCOS cannot be entirely excluded, it seems unlikely in light of the results of the statistical tests (Egger's and Begg's tests). Moreover, all studies regarding PCOS and the FTO gene were included even if they were originally designed not to test for the difference in impact of the FTO genotypes on weight or BMI.
The sample size of 2,548 individuals is smaller than those of many genetic studies comparing effect sizes. In part, this number reflects the limited number of studies in which PCOS women were genotyped for the FTO SNP as we included all available studies referring to FTO in PCOS. The problem of the limited number of PCOS patients who were genotyped may be, at least in part, associated with the need to follow a diagnostic algorithm for this disease that is costly and time-consuming. We acknowledge that, while our study tested the hypothesis that FTO genetic variation has a greater impact on BMI and body weight in the context of PCOS, a biological explanation for this phenomenon remains to be elucidated. The unique metabolic setting of PCOS, which includes significant alterations in sex steroid hormone levels (hyperandrogenaemia and significant extraglandular aromatisation of androgens), reduced sex hormone-binding globulin levels, and insulin resistance, may modify the influence of the FTO genotypes [36]. Alternatively, gene–gene interactions may contribute to the greater influence of FTO. Further studies are needed to elucidate the roles of biochemical and genetic factors affecting the action of FTO in PCOS.
The major studies that have demonstrated an impact of FTO on body weight have been conducted primarily on white populations [3, 37–39]. This initial finding in white cohorts was confirmed in a Mexican population as well as in multiple studies including a recent large-scale meta-analysis of the East and South Asian population [12, 14–16]. In spite of initial data suggesting that FTO had no clear impact on phenotype in African cohorts [40], the effect of the gene variants on obesity was eventually established in a new genome-wide association study in a population with a predominantly African ancestry in the USA [17]. The lack of effect in the initial African study may have been due to the impact of environmental circumstances, such as seasonal food shortages and high physical activity in the African population [40, 41].
Results from one study of the Chinese population, which could not be included in our meta-analysis due to a lack of numerical data, suggested no clear association between FTO allelic variation and obesity-related traits in both healthy individuals and women with PCOS [30]. Subjects from studies included in our meta-analysis were mainly white. Consequently, our findings cannot be generalised to other ethnic groups. Our observations might not be specific to PCOS, as there might exist other phenotypes and diseases in which the effect of the FTO genotypes is larger than in the general population. Finally, we should point out that a slightly greater effect of FTO genotypes was observed in heavier diabetic cohorts than in the general population [3]. Nevertheless, the magnitude of the current finding and its presence in obese and non-obese populations with PCOS suggest that it has a different mechanism in PCOS.
In conclusion, our meta-analysis, based on eight distinct cohorts, shows that the per allele effect of FTO polymorphisms on BMI and weight seems to be more than two times greater than the effect found in large population-based studies. Our results suggest that the metabolic context or specific polygenic background of PCOS modifies the influence of FTO on weight and BMI.
Abbreviations
- AIC:
-
Akaike's information criterion
- HWE:
-
Hardy–Weinberg equilibrium
- LD:
-
Linkage disequilibrium
- PCOS:
-
Polycystic ovary syndrome
- SNP:
-
Single nucleotide polymorphism
References
Finucane MM, Stevens GA, Cowan MJ et al (2011) National, regional, and global trends in body-mass index since 1980: systematic analysis of health examination surveys and epidemiological studies with 960 country-years and 9·1 million participants. Lancet 377:557–567
Peng S, Zhu Y, Xu F, Ren X, Li X, Lai M (2011) FTO gene polymorphisms and obesity risk: a meta-analysis. BMC Med 9:71
Frayling TM, Timpson NJ, Weedon MN et al (2007) A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 316:889–894
Scuteri A, Sanna S, Chen W-M et al (2007) Genome-wide association scan shows genetic variants in the FTO gene are associated with obesity-related traits. PLoS Genet 3:e115
Dina C, Meyre D, Gallina S et al (2007) Variation in FTO contributes to childhood obesity and severe adult obesity. Nat Genet 39:724–726
Speliotes EK, Willer CJ, Berndt SI et al (2010) Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat Genet 42:937–948
Loos RJF, Lindgren CM, Li S et al (2008) Common variants near MC4R are associated with fat mass, weight and risk of obesity. Nat Genet 40:768–775
Wellcome Trust Case Control Consortium (2007) Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature 447:661–678
Zeggini E, Weedon MN, Lindgren CM et al (2007) Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science 316:1336–1341
Scott LJ, Mohlke KL, Bonnycastle LL et al (2007) A genome-wide association study of type 2 diabetes in Finns detects multiple susceptibility variants. Science 316:1341–1345
Saxena R, Voight BF, Lyssenko V et al (2007) Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science 316:1331–1336
Al-Attar SA, Pollex RL, Ban MR et al (2008) Association between the FTO rs9939609 polymorphism and the metabolic syndrome in a non-Caucasian multi-ethnic sample. Cardiovasc Diabetol 7:5
Villalobos-Comparan M, Teresa Flores-Dorantes M, Teresa Villarreal-Molina M et al (2008) The FTO gene is associated with adulthood obesity in the Mexican population. Obesity 16:2296–2301
Tan JT, Dorajoo R, Seielstad M et al (2008) FTO variants are associated with obesity in the Chinese and Malay populations in Singapore. Diabetes 57:2851–2857
Chang Y-C, Liu P-H, Lee W-J et al (2008) Common variation in the fat mass and obesity-associated (FTO) gene confers risk of obesity and modulates BMI in the Chinese population. Diabetes 57:2245–2252
Li H, Kilpeläinen TO, Liu C et al (2012) Association of genetic variation in FTO with risk of obesity and type 2 diabetes with data from 96,551 East and South Asians. Diabetologia 55:981–995
Hassanein MT, Lyon HN, Nguyen TT et al (2010) Fine mapping of the association with obesity at the FTO locus in African-derived populations. Hum Mol Genet 19:2907–2916
Jess T, Zimmermann E, Kring SII et al (2008) Impact on weight dynamics and general growth of the common FTO rs9939609: a longitudinal Danish cohort study. Int J Obes (Lond) 32:1388–1394
Asunción M, Calvo RM, San Millán JL, Sancho J, Avila S, Escobar-Morreale HF (2000) A prospective study of the prevalence of the polycystic ovary syndrome in unselected Caucasian women from Spain. J Clin Endocrinol Metab 85:2434–2438
Diamanti-Kandarakis E, Kouli CR, Bergiele AT et al (1999) A survey of the polycystic ovary syndrome in the Greek island of Lesbos: hormonal and metabolic profile. J Clin Endocrinol Metab 84:4006–4011
March WA, Moore VM, Willson KJ, Phillips DIW, Norman RJ, Davies MJ (2010) The prevalence of polycystic ovary syndrome in a community sample assessed under contrasting diagnostic criteria. Hum Reprod 25:544–551
Teede H, Deeks A, Moran L (2010) Polycystic ovary syndrome: a complex condition with psychological, reproductive and metabolic manifestations that impacts on health across the lifespan. BMC Med 8:41
Kowalska I, Malecki MT, Straczkowski M et al (2009) The FTO gene modifies weight, fat mass and insulin sensitivity in women with polycystic ovary syndrome, where its role may be larger than in other phenotypes. Diabetes Metab 35:328–331
Ewens KG, Jones MR, Ankener W et al (2011) FTO and MC4R gene variants are associated with obesity in polycystic ovary syndrome. PLoS One 6:e16390
Wehr E, Schweighofer N, Möller R, Giuliani A, Pieber TR, Obermayer-Pietsch B (2010) Association of FTO gene with hyperandrogenemia and metabolic parameters in women with polycystic ovary syndrome. Metab Clin Exp 59:575–580
Barber TM, Bennett AJ, Groves CJ et al (2008) Association of variants in the fat mass and obesity associated (FTO) gene with polycystic ovary syndrome. Diabetologia 51:1153–1158
Tan S, Scherag A, Janssen OE et al (2010) Large effects on body mass index and insulin resistance of fat mass and obesity associated gene (FTO) variants in patients with polycystic ovary syndrome (PCOS). BMC Med Genet 11:12
Borenstein M, Hedges LV, Higgins JPT, Rothstein HR (2009) Introduction to meta-analysis. Wiley, Chichester
Balding DJ (2006) A tutorial on statistical methods for population association studies. Nat Rev Genet 7:781–791
Yan Q, Hong J, Gu W et al (2009) Association of the common rs9939609 variant of FTO gene with polycystic ovary syndrome in Chinese women. Endocrine 36:377–382
Vcelak J, Lukasova P, Vankova M et al (2008) FTO gene is associated not only with obesity-related quantitative traits but also with higher OGTT stimulated glycaemia and leptin levels. Diabetologia 51:S326
Attaoua R, Ait El Mkadem S, Radian S et al (2008) FTO gene associates to metabolic syndrome in women with polycystic ovary syndrome. Biochem Biophys Res Commun 373:230–234
Lango H, Weedon MN, Timpson NJ et al (2007) A common variant in the FTO gene region is associated with BMI in the general population and predisposes to adult and childhood obesity. Diabetologia 50:S22
Bennett AJ, Weedon MN, Zeggini E et al (2007) Common variation in the FTO (fused toes) gene is strongly associated with adiposity measures in the Northern Finnish Birth Cohort of 1966. Diabetologia 50:S76
Andreasen CH, Stender-Petersen KL, Mogensen MS et al (2008) Low physical activity accentuates the effect of the FTO rs9939609 polymorphism on body fat accumulation. Diabetes 57:95–101
Le TN, Nestler JE, Strauss JF 3rd, Wickham EP 3rd (2012) Sex hormone-binding globulin and type 2 diabetes mellitus. Trends Endocrinol Metab 23:32–40
Field SF, Howson JMM, Walker NM, Dunger DB, Todd JA (2007) Analysis of the obesity gene FTO in 14,803 type 1 diabetes cases and controls. Diabetologia 50:2218–2220
Price RA, Li W-D, Zhao H (2008) FTO gene SNPs associated with extreme obesity in cases, controls and extremely discordant sister pairs. BMC Med Genet 9:4
Hunt SC, Stone S, Xin Y et al (2008) Association of the FTO gene with BMI. Obesity 16:902–904
Hennig BJ, Fulford AJ, Sirugo G et al (2009) FTO gene variation and measures of body mass in an African population. BMC Med Genet 10:21
Kilpeläinen TO, Qi L, Brage S et al (2011) Physical activity attenuates the influence of FTO variants on obesity risk: a meta-analysis of 218,166 adults and 19,268 children. PLoS Med 8:e1001116
Acknowledgements
The authors are grateful to T. Frayling (Peninsula College of Medicine and Dentistry, University of Exeter, UK) for valuable comments regarding this manuscript. The authors wish to acknowledge A. Stozek, I. Zerda and K. Kapcia from HTA Consulting (Krakow, Poland) for their valuable contribution in systematic search, statistical support and preparation of figures, respectively.
Funding
MTM was supported by the Polish Ministry of Science Grant No. NN407 414436 and by the Jagiellonian University Medical College. MOG and JFSIII were supported in part by the National Institutes of Health (NIH) grant U54 HD034449. ST and AH were financed by the German Federal Ministry of Education and Research (BMBF 01KU0903; NGFNplus: 01GS0820). JV, PL and BB were supported by Internal Grant Agency of the Ministry of Health, CR NS 9839-4 /2008. IK and MS were supported by the grant 3-50709 from the Medical University of Bialystok, Poland. PW and PR were supported by HTA Consulting.
Duality of interest
The authors declare that there is no duality of interest associated with this manuscript.
Contribution statement
PW and PR developed the protocol; PW, AL, ST, PL searched the medical databases and carried out data analysis. PW, PR, KGE, SF, ST, EL, JV, RAt, MS, RAz, TMB, AH, BO-P, PL, BB, FG, IK, MOG, JFSIII, MMcC, MTM researched and interpreted data; PW, AL, PR, KGE, IK, JFSIII, MTM wrote the manuscript. KGE, SF, ST, EL, JV, RAt, MS, RAz, TMB, AH, BO-P, BB, FG, MOG, JFSIII, MMcC, PL, MTM critically reviewed the manuscript, designed the study and coordinated the project. All authors gave final approval of the version of the manuscript to be published.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Consortia
Corresponding author
Additional information
P. Wojciechowski, A. Lipowska and P. Rys contributed equally to this study.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Wojciechowski, P., Lipowska, A., Rys, P. et al. Impact of FTO genotypes on BMI and weight in polycystic ovary syndrome: a systematic review and meta-analysis. Diabetologia 55, 2636–2645 (2012). https://doi.org/10.1007/s00125-012-2638-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-012-2638-6