
Abstract
Key message
Selection over 70 years has led to almost complete fixation of a haplotype spanning ~ 250 Mbp of chomosome 5H in European two-rowed spring barleys, possibly originating from North Africa.
Abstract
Plant breeding and selection have shaped the genetic composition of modern crops over the past decades and centuries and have led to great improvements in agronomic and quality traits. Knowledge of the genetic composition of breeding germplasm is essential to make informed decisions in breeding programs. In this study, we characterized the structure and composition of 209 barley cultivars representative of the European two-rowed spring barley germplasm of the past 190 years. Utilizing high-density SNP marker data, we identified a distinct centromeric haplotype spanning a ~ 250 Mbp large region on chromosome 5H which likely was first introduced into the European breeding germplasm in the early to mid-twentieth century and has been non-recombining and under strong positive selection over the past 70 years. Almost all cultivars in our panel that were released after 2000 carry this new haplotype, suggesting that this region carries one or several genes conferring highly beneficial traits. Using the global barley collection of the German Federal ex situ gene bank at IPK Gatersleben, we found the new haplotype at high frequencies in six-rowed spring-type landraces from Northern Africa, from which it may have been introduced into modern European barley germplasm via southern European landraces. The presence of a 250 Mbp genomic region characterized by lack of recombination and high levels of fixation in modern barley germplasm has substantial implications for the genetic diversity of the modern barley germplasm and for barley breeding.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
An understanding of the genetic structure and genetic diversity of crop germplasm is a prerequisite for crop improvement. The availability of diverse germplasm is essential for the maintenance of food security and for improved resilience of crops to abiotic and biotic stresses, especially under climate change (McCouch et al. 2013). Plant breeding, which is based on combining beneficial alleles controlling desirable traits, relies on the availability of wide germplasm pools from which alleles are selected. Over time, breeding has resulted in the constant improvement of crops in terms of yield, disease resistance and improved agronomic and quality traits. A prominent example of this is the Green Revolution and the introduction of semi-dwarfing genotypes in cereals which led to substantial improvements in yield and yield stability (Hedden 2003). However, a constant risk in breeding is the loss of genetic diversity in modern breeding germplasm and hence the loss of beneficial alleles. The consequences are stagnation of the breeding progress and the inability of breeding programs to adapt to different environments or changing market demands (McCouch et al. 2013; Dawson et al. 2015). While repeated intercrossing of elite cultivars or a narrow germplasm pool often leads to a reduction of genetic diversity, it may also generate new beneficial allelic combinations that lead to an improved phenotype (Rasmusson and Phillips 1997). In addition, new genetic diversity may be generated by introducing more exotic alleles into the existing germplasm. In barley (Hordeum vulgare L. ssp. vulgare), there are contradictory assessments of the overall genetic diversity over time, with some authors reporting a loss of genetic diversity, while most did not observe any significant changes (Donini et al. 2000; Russell et al. 2000; Koebner et al. 2003; Kolodinska Brantestam et al. 2007; Malysheva-Otto et al. 2007; Rajala et al. 2017; Brbaklić et al. 2021). Discrepancies between these studies can likely be attributed to the composition of the population under study, the geographic origin, the time period examined, the relative importance of loss of alleles through selection and the introgression of diverse material. Recent advances in next generation sequencing and high-density marker development have made it possible to not only study genome-wide patterns of genetic diversity but to also identify local changes of genetic diversity over time caused by selection and breeding. For example, the reduced diversity at the HvCEN locus on chromosome 2H in European two-rowed spring barleys was the result of selection for late flowering and increased yield associated with this locus (Tondelli et al. 2013). Similarly, the powdery mildew resistance gene mlo is now present in most European spring barleys as a result of selection and introgression of resistance alleles from Ethiopian barley (Jørgensen 1992; Friedt et al. 2011).
Barley is the fourth most important cereal crop globally, the second most important crop in Europe and is adapted to a wide range of environmental conditions (Friedt et al. 2011). Domesticated in the Fertile Crescent around 10,000 years ago, barley reached Europe approximately 6–8,000 years ago (von Bothmer et al. 2003; Jones et al. 2011). In Europe, latitudinal adaptation through selection for different flowering times and growth habit allowed the cultivation of barley in diverse day length and climatic environments from Northern Scandinavia to the Mediterranean region. Both spring and winter types are grown in Europe today. The main end uses are animal feed and malt (von Bothmer et al. 2003).
Historically, barley was grown as landraces, i.e. relatively heterogenous populations adapted to local conditions. The first selections from popular spring barley landraces were made in the 1800s in the UK and soon thereafter in other regions of Europe as well (Fischbeck 2003). Among these selections, lines such as ‘Chevalier’ and ‘Goldthorpe’ (selections from British landraces ‘Archer’ and ‘Spratt’, respectively), ‘Hana/Hanna’ (selection from Moravian landraces) or ‘Gull’ (selection from landraces from Gotland, Sweden) became widely successful due to their superior agronomic and malting qualities and were distributed across the continent (Fischbeck 2003; Hagenblad and Leino 2022). At the same time, selections from local landraces were made which were well-adapted to specific environmental conditions and sometimes outcompeted the imported selections but led to a decrease in genetic diversity (Fischbeck 2003). Lines from wider geographic crosses, however, ultimately made up the majority of the successful spring cultivars of that time (e.g. ‘Kenia’ or ‘Isaria’) laying the foundation of modern barley breeding in the mid-twentieth century (Fischbeck 2003; Milotova et al. 2008; Friedt et al. 2011) and leading to the development of widely used varieties such as ‘Golden Promise’ and ‘Triumph’. In the 1950s, crosses became genetically wider by using parents from different breeding programs and more exotic material was introduced such as the landrace H. laevigatum which provided resistance to diseases, e.g. powdery mildew (Blumeria graminis) and leaf rust (Puccinia hordei) (Giese et al. 1993; Hickey et al. 2012). The use of mutation breeding contributed important varieties such as ‘Diamant’ in the 1960s and 1970s which had superior agronomic and malting properties (Friedt et al. 2011). Constant genetic improvement of different agronomic and quality traits has been made over time, although high and stable yield has always been the main focus of barley breeding (Friedt et al. 2011; Laidig et al. 2017; FAOSTAT 2022). In addition, breeding for high protein content in feed barley and low protein content and large grain size, among many other traits, in malting barley has resulted in a range of different varieties suitable for different end uses. The introduction of semi-dwarfing genotypes during the green revolution reduced both plant height and harvest losses due to lodging. Further, yield and quality have been greatly increased by the exploitation of genes conferring different levels of resistance to a range of globally important diseases (Friedt et al. 2011).
Here, we present a high-resolution characterization of barley diversity in a panel of 209 landraces and cultivars representative of 190 years of breeding history in the European two-rowed spring barley germplasm (Schreiber et al. 2023). Based on a high-density SNP dataset comprising 1.5 million SNPs from transcript and whole genome shotgun (WGS) sequencing data derived from the entire diversity panel and a gene expression survey of six tissues sampled during plant development, the genetic and multi-omic characterization allowed us to track major haplotype and transcript changes in the European barley breeding history (Schreiber et al. 2023). To fully exploit this rich resource of high-density multi-omics datasets, we thoroughly studied the genetic and structural aspects of this panel. We characterized population substructure and identified regions exhibiting allelic changes over time, indicative of selection. In particular, we highlight a ~ 250 Mbp pericentromeric and centromeric region on chromosome 5H which has been under exceptionally strong selection during the past 70 years, almost reaching fixation in European two-rowed spring varieties released in the past 20 years. The two main haplotypes in this region differ in the expression of 48 genes and we find evidence of reduced recombination between these haplotypes in the past decades. Utilizing genotypic data from the German Federal ex situ gene bank at IPK Gatersleben we identified North African landraces as a potential geographic origin of this recently preferentially selected haplotype.
Materials and methods
Genotypic and expression datasets
The datasets used in this study are described in detail in Schreiber et al. (2023). Briefly, we utilized genotypic and transcript data of 209 barley varieties from the past 190 years which were selected for their representativeness of the European two-rowed spring barley germplasm (Schreiber et al. 2023, Online Resource 1). The genotypic dataset comprised 1,509,446 biallelic SNP markers derived from 150 bp paired-end RNAseq data from six tissues (seedling crown, seedling root, peduncle, developing inflorescence, spikelet at green anther stage and developing grain five days post anthesis), WGS skim sequencing at threefold haploid genome coverage and the 50 k Illumina Infinium iSelect array (Bayer et al. 2017). The SNPs were filtered for < 2% heterozygosity and minor allele frequency (MAF) > 0.025. Missing data were imputed using a haplotype-based approach implemented in Tassel 5.0 (Bradbury et al. 2007) and SNPs were pruned using an LD cutoff of 0.99 in Plink v1.9 (Purcell et al. 2007). The gene expression datasets were constructed from the same RNAseq data after pseudo-alignment against the barley reference transcriptome BaRTv2.0 using Salmon v.1.3.0 (Patro et al. 2017; Coulter et al. 2022). Expression was calculated as log-transformed counts per million (cpm). Unless otherwise stated, all physical positions of genetic features refer to the reference genome of ‘Barke’, a European two-rowed spring barley variety (Jayakodi et al. 2020).
Population genetics analyses
To visualize genetic patterns over time, a Discriminant Analysis of Principal Components (DAPC) was performed on the panel clustered by release period (1830–1959, 1960–1969, 1970–1979, 1980–1989, 1990–1999, 2000–2009, 2010–2014) using the function ‘dapc’ from the R package ‘adegenet’ v.2.1.4 and 600,000 randomly selected SNPs (Jombart 2008). The 'adegenet' function ‘xvalDapc’ was used to perform a cross-validation with 100 repetitions to identify the optimal number of PCs to retain for the analysis.
To visualize genetic variation and population structure, a Principal Coordinate Analysis (PCoA) was performed using all 1,509,446 SNPs in the dataset. A neighbor-joining tree was created based on a distance matrix using the ‘nj’ function in the R package ‘ape’ v.5.6–1 (Paradis and Schliep 2019). To select the number of principal components required to explain ~ 90% of the variance in the population and to identify the most likely number of subpopulations and to assign accessions to these subpopulations, k-means clustering using the function ‘find.cluster’ from the R package ‘adegenet’ v.2.1.4 was used with 10,000 iterations. Due to limited computational resources, the clustering of the European two-rowed spring barley panel was done on a subset of 1,000,000 randomly selected SNPs.
The polymorphism information content (PIC) was calculated in rolling windows of 10,000 SNPs using the following equation (Botstein et al. 1980):
with pi and pj being the frequency of the ith and jth allele, respectively and n the number of markers. Frequencies of reference alleles (of cultivar ‘Barke’) were calculated in rolling windows of 10,000 SNPs.
FST (Weir and Cockerham 1984) between subpopulations was calculated in 100 kbp windows with a step size of 10 kbp using the software vcftools 0.1.16 (Danecek et al. 2011). Pairwise FST between cultivar groups from different release periods was calculated using the ‘pairwise.WCfst’ function from the ‘hierfstat’ package v.0.5–10 (Goudet 2005). Variance within and between subgroups defined by release period was calculated using analysis of molecular variance (AMOVA) as implemented in the ‘poppr.amova’ function in the R package ‘poppr’ (Kamvar et al. 2014) and a subset of 400,000 randomly selected SNPs.
Haplotype characterization
Two major haplotypes were apparent in the centromeric region of chromosome 5H through FST analysis. Utilizing the gene expression data of six tissues, we studied patterns of gene expression in the genes located in this region and tested whether gene expression is haplotype-dependent. A PCoA of the log-transformed cpm values was performed on the cultivars to identify clusters of individuals with similar patterns of gene expression and to look for overlaps between gene expression clusters and haplotype clusters. Next, a PCoA of the log-transformed cpm values was performed on the genes and combined with a k-means cluster analysis to cluster genes according to their expression profiles across cultivars. Expression of genes with haplotype-dependent expression was visualized across all cultivars and tissues in a heatmap using the ‘pheatmap’ package in R. Dotplots of genome alignments were made using a custom R script utilizing lastz 1.04.03 (Harris 2007). GO term enrichment analysis was performed with the R package ‘topGO’.
Haplotype analysis in the collection of the German Federal ex situ gene bank at IPK Gatersleben
The origin and geographic distribution of the haplotype present in the cluster of more recent cultivars was analysed in a collection of approximately 20,000 domesticated barley accessions stored in the German Federal ex situ gene bank at IPK Gatersleben for which high-resolution SNP datasets generated by genotyping-by-sequencing (GBS) and morphological and agronomic passport data are available (Mascher, pers. comm.; Milner et al. 2019). Since the SNP dataset was mapped to the third version of the reference genome of cultivar ‘Morex’ (Mascher et al. 2021), we blasted 5000 bp sequences flanking the region of interest on chromosome 5H of cultivar ‘Barke’ (as identified by high FST values) against the ‘Morex’ V3 genome (67–320 Mbp on chromosome 5H) and extracted the SNPs in this region. The SNP dataset was filtered for < 20% missing data and a minor allele count of 1, resulting in 6536 SNP markers in the region. Using the k-means clustering approach described above we identified the number of clusters that sufficiently described the structure in the dataset and performed a PCoA on the domesticated barley accessions. Based on the cluster membership of accessions with known haplotypes (from the panel of 209 European cultivars), we identified gene bank accessions belonging to the cluster corresponding to the newer haplotype. Maps showing the distribution of this haplotype by country and different passport data categories (e.g. cultivation status, row type etc.) were made using the R package ‘maps’. Countries for which fewer than 15 accessions per respective category were present were excluded.
Results
Genetic changes in the population over time
To characterize genetic changes in the population over time, we divided the population into different groups according to release year, either in periods of 10 years or 20 years from 1960 onwards with landraces and cultivars released before 1959 forming a separate group.
Changes based on release year along the first axis were apparent in the DAPC analysis (Fig. 1). Cultivars with a greater difference in release year tended to be located further apart than cultivars released at a similar time. The clusters including cultivars released before 1980 showed considerable overlap, while the newer clusters showed more variation within and between clusters suggesting an increase in genetic diversity. These observations are reflected in pairwise FST values between the groups based on release period, which tended to increase with temporal distance between time periods (Online Resource 2). Most of the genetic variation in the panel was found between samples within a release period (86%), whereas smaller fractions were attributed to variation between release periods (14%) (Online Resource 3).

Discriminant Analysis of Principal Components (DAPC) of the European two-rowed spring barley panel with pre-defined groups according to release period using 600,000 randomly selected SNPs. Principal component (PC) 1 represents a development of the population over time. Increased variation between and within more recent clusters suggests an increase in genetic diversity after the 1980s
PIC values differed greatly between release periods in the pericentromeric and centromeric region of chromosome 5H, which showed very low PIC values in groups of cultivars released before 1960 and after 2000 and higher PIC values in the cultivar groups released between 1960 and 2000 (Fig. 2a). Reference allele frequencies increased continuously over time in this region: the groups of cultivars released before 1979 were characterized by reference allele frequencies between 0 and 0.36, whereas the group of cultivars released between 1980 and 1999 had reference allele frequencies around 0.75 and the cultivar group released after 2000 had reference allele frequencies between 0.89 and 0.96 (Fig. 2b). Thus, older cultivars have a different haplotype that has decreased in frequency over time whereas modern cultivars released after 2000 almost exclusively carry the same haplotype as the reference cultivar ‘Barke’.

Polymorphic information content (PIC) (a) and frequencies of the reference alleles (from cultivar ‘Barke’) (b) in rolling windows of 10,000 SNPs in cultivar groups based on release period. Regions of interest are highlighted by vertical red dotted lines and described in the text. The pericentromeric and centromeric region on chromosome 5H (69–320 Mbp) is characterized by low PIC values in early and recent cultivar groups and a strong increase in reference allele frequency in cultivar groups released after the 1980s, providing evidence for a change in haplotype over time. Centromeric positions were estimated by mapping centromeric Morex V3 genes to Barke and are highlighted by vertical grey dotted lines (color figure online)
Another region in which PIC values varied greatly was identified at 330.42 Mbp on chromosome 1H. PIC values decreased over time from 0.33 in the group of cultivars released before 1959 to almost reaching zero in the group of cultivars released after 2000 (Fig. 2a). This development was due to an increase of the ‘Barke’ allele over time, which reached almost complete fixation among the newest cultivars in our panel (Fig. 2b). The entire pericentromeric and centromeric region of chromosome 1H, generally a region of low diversity in spring barleys (Mascher et al. 2017; Schreiber et al. 2023), was characterized by an increase of the ‘Barke’ allele over the last decades.
Similarly, the newest cultivar group showed low PIC values at 81.4 Mbp on chromosome 2H and 45.6 Mbp on chromosome 6H (Fig. 2a). These regions have potentially been subjected to a decrease in genetic diversity in breeding programs in recent years as frequencies of the ‘Barke’ alleles of > 0.95 were observed in the newest cultivars whereas these alleles were present at much lower frequencies in the older cultivar groups (Fig. 2b). Conversely, regions with high PIC values on chromosome 3H, 6H and 7H in the more recent groups may have experienced recent introgressions from other germplasm sources, which is further supported by a drop in reference allele frequency, suggesting that new alleles not present in ‘Barke’ have been recently introduced to the germplasm (Fig. 2).
Clustering analysis and population structure
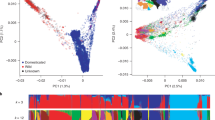
In addition to the a priori clustering of cultivars by release year described above, overall population structure was analysed using several different approaches. The PCoA (Fig. 3) and the neighbor-joining tree (Online Resource 4) suggest a moderate degree of clustering into two groups which to some extent is attributable to the release year of the cultivars. The first principal coordinate (PCo1) placed the cultivars on a gradient corresponding to the release year, explaining 16.5% of the total variation and the neighbour-joining tree clustered most of the older cultivars released before 1990 into a separate group which included only few cultivars released after 1990. A clustering according to geographic origin was not apparent (Online Resource 5). In addition, we used k-means clustering to identify the most likely number of subpopulations. Approximately 110 principal components (PCs) were required to capture ~ 90% of the variation in the dataset, underlining the lack of major structure in the population (Online Resource 6). With 110 PCs retained the lowest Bayesian information criterion (BIC) value, an indicator of the best fit was reached at k = 9 clusters. Given the lack of strong population structure observed in the PCoA, such a high number of subpopulations seemed unlikely. Hence, and since we were interested in the main drivers of population differentiation, we followed the ‘elbow method’ instead and concluded that two or three clusters would provide a reasonable description of the data (Online Resource 4, Online Resource 7). Assuming two subpopulations, the panel was divided into a slightly older (cluster 1, most cultivars released before 1990) and a slightly younger population (cluster 2, most cultivars released after 1980) (Table 1, Online Resource 1, Online Resource 7), coinciding with the clusters apparent in the PCoA (Fig. 3). At k = 3, the cluster of more recent cultivars was further divided into two clusters consisting of cultivars of similar age and geographic origin (clusters 2 and 3) (Table 1, Online Resource 1, Online Resource 7). Two cultivars that belonged to the older cluster at k = 2 were assigned to one of the younger clusters (cluster 3) at k = 3.

Principal coordinate analysis (PCoA) of the European two-rowed spring barley panel using the genotypes of 1,509,447 SNP markers. Each dot represents a cultivar, color-coded according to the period in which they were registered as commercial cultivars
FST analysis detected genomic regions characterized by different allele frequencies in these clusters (Fig. 4). FST values above 0.7 between 68.78 and 320.04 Mbp on chromosome 5H indicate very different allele frequencies and therefore support the presence of different haplotypes in this region in the two clusters at k = 2 (Fig. 4a). This is confirmed by low PIC values in this region and low reference allele frequencies in the old cluster (cluster 1) and high reference allele frequencies in the new cluster (cluster 2) (Fig. 5a and b). In addition, FST values above 0.8 were found at 267 Mbp on chromosome 3H and at 34 Mbp on chromosome 4H (Fig. 4a). At k = 3, the regions on chromosomes 5H, 3H and 4H were associated with FST values above 0.7 between clusters 1 and 2 and 1 and 3, respectively (Fig. 4b and c). In addition, loci with FST values above 0.6 at 248 Mbp on chromosome 6H and at 316 Mbp on chromosome 7H were apparent between clusters 1 and 2 (Fig. 4b). The differentiation between the two newer clusters 2 and 3 was due to an almost complete fixation for opposite alleles between 254.2 and 381.1 on chromosome 3H (FST > 0.8 in most 10 kbp windows in the region) in addition to small regions on chromosomes 5H, 6H and 7H (Fig. 4d). While cluster 3 is almost fixed for the ‘Barke’ haplotype in the region on chromosome 3H, cluster 2 is almost fixed for a different haplotype (Fig. 5c and d). Interestingly, the region of fixation in cluster 3 is larger than in cluster 2. The genomic regions at 267 Mbp on chromosome 3H, at 248 Mbp on chromosome 6H and at 316 Mbp on chromosome 7H coincided with the approximate centromere positions and may be caused by mapping errors to these highly repetitive regions.

FST between clusters in 100 kbp windows with a step size of 10 kbp along the genome between a clusters 1 and 2 at k = 2, b clusters 1 and 2 at k = 3, c clusters 1 and 3 at k = 3, and d clusters 2 and 3 at k = 3. Clusters were determined by k-means clustering under the assumption of two or three clusters, respectively. Regions with high FST values between different clusters are highlighted by vertical red dotted lines and described in the text. The pericentromeric and centromeric region on chromosome 5H (68–320 Mbp) is characterized by high FST values between the two clusters at k = 2 as well as between clusters 1 and 2 and 1 and 3 at k = 3 respectively. FST values of 0.7–1 in this region suggest almost complete fixation of different haplotypes in the different clusters. Centromeric positions were estimated by mapping centromeric Morex V3 genes to Barke and are highlighted by vertical grey dotted lines (color figure online)

Polymorphic information content (PIC) (a, c) and frequencies of the reference alleles (from cultivar ‘Barke’) (b, c) in rolling windows of 10,000 SNPs in the different k-means clusters at k = 2 (a, b) and k = 3 (c, d). Regions of interest are highlighted by vertical red dotted lines and described in the text. The pericentromeric and centromeric region on chromosome 5H (69–320 Mbp) is characterized by low PIC values in all clusters. Low reference allele frequencies in older cultivars (cluster 1 at k = 2 and k = 3) and high reference allele frequencies in newer cultivars (cluster 2 at k = 2 and clusters 2 and 3 atk = 3) illustrate the haplotype change over time in this region. Centromeric positions were estimated by mapping centromeric Morex V3 genes to Barke and are highlighted by vertical grey dotted lines (colour figure online)
Analysis of haplotypes and gene expression on chromosome 5H
In the chromosome 5H region characterized by high FST values between the older cluster 1 and the more recent cluster 2 (at k = 2), including the flanking 10 kbp, 1080 BaRTv2.0 gene models are present, of which 827 had an expression of > 1 cpm in > 30% of the cultivars in at least one of the six tissues. A PCoA of the log-transformed cpm expression values of these genes separated the cultivars into the two k-means clusters along the first PC in all six tissues (Fig. 6, orange and green groups); however, six cultivars (‘Georgie’, ‘Vada’, ‘Reggae’, ‘Abacus’, ‘Apex’ and ‘Livet’) formed two separate clusters between the two main haplotype groups. Across all six tissues we identified 46 genes which formed a separate cluster (red cluster, Fig. 7, Online Resource 8) whose expression was strongly haplotype-dependent (Online Resources 9–14). In the haplotype predominant in older cultivars (henceforth referred to as haplotype 1) these genes were characterized by no or low expression, whereas in cultivars carrying the new haplotype (haplotype 2) the genes were moderately to highly expressed (Online Resource 9–14). In addition, we identified two genes highly expressed in cultivars carrying haplotype 1 and lowly expressed in cultivars carrying haplotype 2. These patterns were consistent across all six tissues. In the six cultivars forming two distinct clusters, expression of these genes was more variable and did not follow the described pattern. Here, two patterns in gene expression were apparent with cultivars ‘Georgie’, ‘Vada’ and ‘Abacus’ forming one group and ‘Reggae’, ‘Apex’ and ‘Livet’ forming another group. ‘Georgie’ and ‘Abacus’ are derived from crosses with ‘Vada’ which is a cross between ‘Gull’ and H. laevigatum. A PCoA of the 119,914 SNPs in the region placed the six cultivars into the same two distinct clusters between haplotype 1 and 2, confirming that these cultivars indeed carry distinct haplotypes (Online Resource 15). We define the H. laevigatum-derived haplotype as haplotype 3 and the haplotype found in ‘Reggae’, ‘Apex’ and ‘Livet’ as haplotype 4.

Principal coordinate analysis (PCoA) of the expression of the 827 genes in the haplotype region on chromosome 5H in six tissues. Cultivars are color-coded according to the haplotype they carry

Principal coordinate analysis (PCoA) of the expression of the 827 genes in the haplotype region on chromosome 5H in six tissues. Each dot represents a gene, color-coded by the k-means cluster they were assigned to under the assumption of the presence of four distinct gene clusters (k = 4). A group of in total 46 genes (red) is apparent along PC2 which does not cluster with the rest of the genes in this region. These are the genes which are up-regulated in haplotype 2. In addition, one or two genes cluster separately on the opposite end of PC2 from the red cluster. These are the genes which are down-regulated in haplotype 2 (color figure online)
The pronounced changes of FST values at the beginning and at the end of the 5H region and graphical genotypes of 10,000 randomly selected markers (Fig. 4, Online Resource 16, Online Resource 17) illustrate the distinctness and the lack of recombination between haplotype 1 and 2. Except for cultivars ‘Novello’, ‘Heron’, ‘Fairytale’, ‘Chieftain’, ‘Berenice’, ‘Nomad’, ‘Kym’ and ‘Drost’ the start of the haplotype block is very conserved (Online Resource 17a and c), and the end of the haplotype block is conserved in all cultivars (Online Resource 17b and 17d). No signs of recombination are apparent. Of the recently published 20 reference genomes representing the first version of a barley pan-genome (Jayakodi et al. 2020) we identified haplotype 1 in the accessions and cultivars ‘Golden Promise’, ‘Morex’, ‘HOR 13821’, ‘HOR 3081’ and ‘HOR 3365’ and haplotype 2 in ‘Barke’, ‘Igri’, ‘HOR 13942’ and ‘RGT Planet’. Genome alignments show an insertion at the beginning of the haplotype block in ‘Barke’ and ‘RGT Planet’ (ca. 61–68 Mbp in ‘Barke’) which is not present in accessions and cultivars carrying haplotype 1, as well as two inversions at 79–84 Mbp (Online Resources 18a and b, Online Resource 19). In addition, the end of the haplotype block is characterized by an insertion in haplotype 2 (ca. 312–316 Mbp in ‘Barke’) followed by a potential translocation of a haplotype 2 segment further upstream into haplotype 1 (Online Resources 18a and c).
A Blast search showed that the genomic region of BaRT2v18chr5HG229090 at 81.11 Mbp was not found in any of the accessions carrying haplotype 1 in the pan-genome but is present in haplotype 2 accessions. In addition, BaRT2v18chr5HG238070, BaRT2v18chr5HG238380, BaRT2v18chr5HG238390, BaRT2v18chr5HG238400, BaRT2v18chr5HG238410, BaRT2v18chr5HG238630, BaRT2v18chr5HG238640 at 307.73–317.47 Mbp at the end of the haplotype region are absent in haplotype 1 genomes. BaRT2v18chr5HG238320 (311.83 Mbp), BaRT2v18chr5HG238440 and BaRT2v18chr5HG238450 (315.86–315.88 Mbp) showed Blast hits further downstream in haplotype 1 accessions, suggesting that they are present on the translocated segment.
A GO enrichment analysis of the 1080 genes located between 68.78 and 320.04 Mbp in chromosome 5H identified six significantly enriched terms suggesting an involvement of these genes in general processes such as calmodulin and abscisic acid binding, phosphatase activity and salicylic acid biosynthesis processes (Online Resource 20).
Occurrence of the new haplotype 2 in a global barley germplasm collection
The oldest cultivar in the panel carrying haplotype 2 is ‘Union’, a variety released in 1955 and with southern German pedigree which can be traced back to Bavarian landraces and landrace selections. Over the following 40 years, haplotype 2 quickly increased in frequency in the panel. All but one cultivar released after 2000 carry this haplotype (Fig. 8, Online Resource 1). We utilized the barley collection of the German Federal ex situ gene bank at IPK Gatersleben to identify the haplotypes of accessions in the pedigree of the cultivars in our panel and to make inferences about the origin and geographic distribution of haplotype 2 in a global context (Milner et al. 2019). Eighty-three of the 209 cultivars in our panel are part of the gene bank collection. Including duplicate entries, we retrieved 243 gene bank accessions that carried the same name as cultivars in our panel and that had matching passport data (country of origin, row type and growth habit) (Online Resources 21 and 22). As expected, these accessions formed distinct clusters according to their haplotypes (Fig. 9a). A few accessions known to carry haplotype 1 were apparent in the haplotype 2 cluster and vice versa, which was likely due to genetically distinct accessions carrying the same name and/or incorrect passport information. The k-means cluster analysis did not clearly identify an optimal number of subpopulations. Due to the relatively low number of SNP markers (n = 6535) the analysis was not powerful enough to consistently distinguish between haplotypes 1 and 3 and putative additional haplotypes at different values for k. However, at k > = 7 the cluster corresponding to haplotype 2 remained stable, hence we assumed k = 7 subpopulations (Online Resource 23).

Changes of chromosome 5H haplotype frequencies in the population over time. Colored lines indicate the percentage of cultivars released in a certain time period carrying the respective haplotype (color figure online)

PCoA of all domesticated barley accessions of the German Federal ex situ gene bank at IPK Gatersleben utilizing 6535 SNPs in the haplotype region on chromosome 5H. Colored dots represent accessions carrying the same names as cultivars and landraces from the European two-rowed spring barley panel, color-coded by haplotype. Grey dots represent gene bank accessions not present in the panel (color figure online)
Northern Africa became apparent as the geographic region with the highest percentage of landraces carrying haplotype 2 (Morocco 84.2%, Chad 63.1% Tunisia, 63.5%, Libya 58.9%, Algeria 56.3%), suggesting this region as the potential origin of this haplotype (Fig. 10a, Online Resource 24). In Europe, haplotype 2 was predominantly found in landraces originating from the Mediterranean region but also from Western Europe as well as the UK. A high frequency of haplotype 2 was also observed among Bolivian and Uruguayan landraces (69.2 and 58.3%, respectively). A north–south gradient of haplotype 2 frequency was apparent in European cultivars (Fig. 10b, Online Resource 24). Fifty-six percent of Spanish cultivars carried haplotype 2. In addition, haplotype 2 was widespread in Mexican cultivars (53.8%). In general, haplotype 2 occurred in higher frequencies among six-rowed and winter cultivars than among two-rowed and spring cultivars, underlining that this haplotype is not row type- or growth habit-specific.

Geographical distribution of haplotype 2 in a landraces and b cultivars in the IPK gene bank collection. Colors indicate the frequency of gene bank accessions from a given country carrying haplotype 2. Countries from which less than 15 accessions per respective category are present in the gene bank are shown in grey
Discussion
The European two-rowed barley germplasm analysed in this study did not separate into distinct clusters. A general differentiation according to the age of the varieties was apparent which likely reflects the breeding history of the population over time. This differentiation accounted for 16.7% of the genetic variation which is consistent with Tondelli et al. (2013) and Ovesná et al. (2013) who attributed 14% and 23% of the genetic variation to changes over time in European two-rowed spring barleys and Czech barleys, respectively. The DAPC analysis suggested a slight increase in genetic diversity in modern accessions after 1980. The reason for this may be introgressions of exotic germplasm, mainly as a source of disease resistance (Fischbeck 2003). The lack of strong clustering in the population is not surprising, given that most of the modern cultivars can be traced back to founder genotypes that were widely used in European breeding, such as ‘Kenia’, ‘Triumph’, ‘Diamant’, ‘Gull’ or ‘Binder’.
A major haplotype switch has occurred on chromosome 5H since the 1960s and 1970s
The most striking genetic change over time was observed on chromosome 5H, where a large haplotype block of ~ 250 Mbp was replaced by another haplotype over the course of the last 60 years. This region has been described previously as divergent between different European barley subpopulations. Tondelli et al. (2013) identified high FST values in the pericentromeric region on chromosome 5H between two clusters representing old and new European two-rowed spring barleys. Similarly, the pericentromeric region on chromosome 5H was characterized by high FST values between Polish cultivars released before 1945 and after 2000, respectively (Dziurdziak et al. 2022). Both authors reported low PIC values in the older group and higher PIC values in the newer group of lines studied. This is affected by the composition of the different clusters, i.e. the frequency of the alleles representing the two haplotypes in the clusters and is dependent on the relative impact that the diversity at this locus has on the clustering of the entire population. In our population, both clusters showed low PIC values in this region, suggesting that the division into subpopulations was mostly driven by the genetic differentiation in this region, underlining the importance of this genomic region for genetic differentiation of the whole population.
Using high-density SNP data and gene expression data we are now able to delineate this region more accurately and characterize it more comprehensively. The region between 68.78 and 320.04 Mbp is characterized by high FST values (> 0.7) which differ clearly from the adjacent regions with lower FST values. FST values remain fairly high along the entire region but are especially high at the start and the end of the region. The start and end points of this region are very conserved except in a few cultivars. We did not see any signs of recombination between the two haplotypes in modern European cultivars, which is not uncommon for centromeric and pericentromeric regions of barley chromosomes (Mayer et al. 2012; The International Barley Genome Sequencing Consortium 2012; Mascher et al. 2017). This region showed segregation bias in three out of 45 systematic crosses between 23 spring barleys, making it one of the large regions in which segregation distortion occurred most frequently in a study by Casale et al. (2022). Structural variation, mainly 5–7 Mbp insertions in cultivars carrying haplotype 2, was identified on either side of the haplotype block which may have further reduced collinearity of the two haplotypes and therefore reduced the ability for meiotic recombination. In Arabidopsis, structural rearrangements frequently occur in centromeric and pericentromeric regions (Jiao and Schneeberger 2020). Whereas the haplotype block itself is collinear except for a small inversion at 210 Mbp and a larger inversion around 250 Mbp, the rearrangements in the flanking regions may have had a detrimental effect on recombination and may have thus facilitated the divergence of the two haplotypes.
Within the short time span of approximately 60 years, haplotype 2 has completely replaced the older haplotype 1 in our population which strongly suggests that this region harbors one or more loci associated with beneficial traits which have been under strong selection. We identified 46 genes across the region which were consistently upregulated in all six tissues in haplotype 2 compared to haplotype 1 and two genes which were downregulated in haplotype 2. The annotation of these genes did not suggest an involvement in specific pathways or traits (Online Resources 8 and 20). One of the 46 differentially expressed genes is annotated as a disease resistance protein (CC-NBS-LRR class, BaRT2v18chr5HG236990) family gene (Online Resource 8). Tondelli et al. (2013) and Dziurdzak et al. (2022) both suggested that the leaf rust resistance locus Rph2 might be the reason for selection of the new haplotype (Borovkova et al. 1997). Rph2 may thus have conferred resistance to isolates in the North African countries, which led to an increase in haplotype 2 frequency in the North African landraces. Indeed, Rph2 is one of the most common leaf rust resistance genes in North African barleys (Elmansour et al. 2017). Reinhold and Sharp (1982) showed that cultivars carrying Rph2 were resistant against some Moroccan isolates but susceptible to others. Rph2 was originally identified in mostly South American and Australian cultivars such as ‘Weider’, ‘Ricardo’, ‘Peruvian’, ‘Bolivia’, ‘Juliaca’, ‘Quinn’, ‘Purple Nepal’, ‘Modia’, ‘Marco’, ‘Reka I’ and ‘Morocco’ (Henderson 1945; Franckowiak et al. 1997; Sandhu et al. 2012). Today, resistance conferred by Rph2 has been overcome in many parts of the world including Europe, the US, North Africa, Australia and the Middle East (Reinhold and Sharp 1982; Niks et al. 2000; Mehnaz et al. 2021).
Few virulence tests have been conducted on the cultivars that were part of this study. Among these, ‘Armelle’, ‘Claret’, ‘Delta’, ‘Egmont’, ‘Favorit’, ‘Hart’, ‘Koral’, ‘Krystal’, ‘Rainbow’, ‘Rapid’, ‘Spartan’, ‘Union’ and ‘Zephyr’ were postulated to have Rph2 conferring seedling resistance, whereas ‘Abacus’, ‘Atlas’, ‘Blenheim’, ‘Bonus’, ‘Corniche’, ‘Derkado’, ‘Georgie’, ‘Landlord’, ‘Natasha’, ‘Optic’, ‘Orbit’, ‘Tyne’, ‘Vada’ and ‘Wisa’ were postulated to have other resistance genes but not Rph2 (Parlevliet 1983; Dreiseitl and Steffenson 2000; Golegaonkar et al. 2009). While this classification is generally in good agreement with the haplotypes present in these cultivars, in a few cultivars the postulated presence or absence of Rph2 did not coincide with the respective haplotype. ‘Delta’ and ‘Zephyr’ were postulated to carry Rph2 resistance but were found to have haplotype 1, whereas ‘Blenheim’, ‘Landlord’ and ‘Optic’ were postulated to not carry Rph2 but do carry haplotype 2. In addition, ‘Abacus’, ‘Georgie’ and ‘Vada’ do not carry Rph2 but carry the third haplotype likely derived from H. laevigatum through ‘Vada’. H. laevigatum carries the Rph20 resistance locus on chromosome 5H which, in contrast to Rph2, confers adult plant resistance to P. hordei. Rph20, however, is located at ~ 7 Mbp on the short arm of chromosome 5H and is therefore not a potential target of selection of haplotype 2. The expression of the NBS-LRR gene BaRT2v18chr5HG236990 and the genotype of SNP S5H_281577407 located within the gene correlate perfectly with the haplotypes we identified; hence, this gene is not a candidate for Rph2. We did not identify any SNPs whose genotypes correspond perfectly with the postulated presence or absence of Rhp2. This could be because it was suggested that there are more than two alleles of Rph2, while our dataset contains only biallelic SNPs and will therefore miss more diverse loci. The general good agreement between the presence of Rph2 and haplotype 2 is not a conclusive proof for Rph2 being the reason for selection of haplotype 2 but it provides enough evidence to warrant further studies. Ideally, an extensive screening of European and North African barley with isolates from North Africa and Europe representative of different time periods would shed light on the development of virulence over time.
Another potential reason for genetic differentiation is suggested by Contreras-Moreira et al. (2019) who found that the region of 71–348 Mbp (first version of the reference genome of ‘Morex’, Mascher et al. 2017) on chromosome 5H differentiated a group of Spanish six-rowed landraces from coastal regions of Southern Spain from other landraces and cultivars of the Spanish Barley Core Collection (SBCC). The coastal landraces carried a unique haplotype not found in the rest of the SBCC. In addition, this region was associated with agroclimatic variables related to the onset and duration of frost periods as well as vernalization potential. Lower frost tolerance is, however, an unlikely selection target for European barleys and does not explain why this haplotype has increased in frequency so quickly. It is more likely that the lower frost tolerance is linked with another unknown beneficial trait and has therefore been indirectly selected for. In an earlier characterization of the SBCC in terms of agronomic traits, it was shown that this group of landraces had significantly higher leaf rust resistance and higher grain yield under low-yielding conditions than the rest of the collection and was heading earlier than other landraces (Yahiaoui et al. 2014). It is however not clear whether grain yield and earliness are associated with the region on chromosome 5H in the SBCC. Fang et al. (2014) found high levels of linkage disequilibrium, a low number of haplotypes and FST values of > 0.8 in the pericentromeric region of chromosome 5H between wild barleys from the Western and the Eastern part of the Fertile Crescent. This region of high FST was associated with environmental variables related to precipitation. The Western group, which was collected between the Eastern Mediterranean and the Zagros Mountains, originates from a more humid climate than the Eastern group. The authors suggested that positive selection and/or structural rearrangements or suppression of recombination might have caused the differentiation in this region. While the two studies did not conclusively show that adaptation to climatic conditions is the reason for the genetic differentiation between groups in this genomic region, it is worthwhile to test this in further studies.
The haplotype frequency screening of the IPK gene bank identified Northern Africa as the geographic region with the highest proportion of accessions carrying haplotype 2. It is therefore likely that the haplotype first occurred in this region and that the haplotype confers a selective advantage under Northern African conditions. However, care must be taken when making inferences based on the gene bank data alone. While being one of the largest barley germplasm collections worldwide, it is not free from biases. For example, modern cultivars and breeding material are not well-represented in the collection due to proprietary reasons, so the frequencies of haplotype 2 shown in Fig. 10 are likely an underestimation as they do not reflect the strong selection for this haplotype in Europe in the last decades that we have seen in our panel. Furthermore, as many countries are underrepresented in the collection, passport data of many accessions are incomplete, and different collections may be subject to different sampling biases, the global haplotype distribution is equally incomplete and needs to be interpreted with caution.
Similarly, potential mismatches between gene bank genotypes and passport data became apparent when we attempted to identify the haplotypes of accessions that were progenies of the oldest carriers of haplotype 2 in our panel. The oldest accession in the panel, Union, is a cross between a cross of ‘Weihenstephaner Mehltauresistente II’ x ‘Donaria’ and ‘Firlbeck 621’. All accessions containing ‘Mehltauresistente’ (= resistant to powdery mildew) in their name were assigned to the haplotype 1 cluster, excluding this parent as the likely haplotype donor for ‘Union’. Two out of three accessions named ‘Donaria’ were part of the haplotype 1 cluster while the third one belonged to another group. The collection contains one accession called ‘Firlbecks III/621’ and two accessions called ‘Firlbecks III Neu’. The former possesses haplotype 2 while the latter two carry haplotype 1. It is therefore not possible to conclude that ‘Firlbeck 621’ is the haplotype 2 donor of ‘Union’. While ‘Firlbecks Union’, ‘UNION (BENDER) AUSGANGSFORM 4X’ and two accessions called ‘Union’ all have haplotype 2, there is also one accession named ‘UNION AUSGANGSFORM 2X’ which carries haplotype 1. ‘Firlbeck III’ is derived from a cross between ‘Firlbeck II’ and ‘Haisa’. ‘Firlbeck II’ is not part of the genebank collection and none of the accessions including the name ‘Haisa’ had haplotype 2 except for one accession simply named ‘Haisa’. ‘Haisa’ is likely ‘Haisa I’, a cross between ‘Heines Hanna’ and ‘Ackermann Isaria’. None of the accessions containing the names ‘Hanna’ and ‘Isaria’ carried haplotype 2. Despite these limitations of utilizing passport data and genetic information from gene bank collections we do believe, however, that the general patterns of haplotype distribution are reliable, simply due to the large number of accessions analysed.
Implications for barley breeding
The presence of a 250 Mbp genomic region characterized by lack of recombination and high levels of fixation in modern barley germplasm has substantial implications for barley breeding. Contemporary barley breeding relies primarily on the selection of favourable combinations of alleles generated by crosses between breeder strains (Fischbeck 2003). While it could be argued that this haplotype has remained largely unchanged since its introduction in the 1950s because it already represents the best combination of favourable alleles, it appears more likely that the lack of recombination has hampered the selection of better combinations. The region contains 1080 genes (2.7% of the 39,419 genes in BaRTv2.0) and it is very unlikely that they all carry the most beneficial allele, especially since this haplotype has developed in a completely different environment (North Africa, in landraces) compared to where it is “used” now (Europe, intensive agriculture).
Given the high prevalence of haplotype 2 in current European two-rowed spring barleys it is likely that haplotype 2 is also present at very high frequencies in the breeding material of most Central, Western and Southern European breeding programs. Hence, current barley breeding methods are unlikely to be able to increase genetic diversity in this region. Due to the low recombination rate in centromeric and pericentromeric regions of barley chromosomes (Mayer et al. 2012; Baker et al. 2014) crosses between genotypes carrying different haplotypes are not an efficient way of breaking the linkage and increasing diversity in this region. Instead, a more promising approach for increasing genetic variation and exploiting new allelic combinations is to screen for different 5H haplotypes present in landraces and unadapted material (e.g. crop wild relatives) for association with beneficial traits. This approach already proved successful in the past when H. laevigatum was widely used as a crossing partner in European barley breeding for its resistance against powdery mildew and leaf rust. A similar approach may be necessary to increase genetic variation in the centromeric regions of chromosomes 1H, 2H and 7H characterized by low diversity in spring barley (Mascher et al. 2017; Schreiber et al. 2023).
Conclusions
In conclusion, we identified a ~ 250 Mbp centromeric region on chromosome 5H which has been under strong selection in European barley cultivars in the past 70 years. We speculate that the haplotype 2 first occurred in North Africa and conferred a selective advantage over other haplotypes, possibly due to leaf rust resistance. It may then have been introduced to Southern European landraces and thus found its way into European cultivars via this route. Similarly, it may have been introduced to South America via Spanish landraces as well (Friedt et al. 2011). This proposed route is highly speculative and further studies will be required, especially to test the hypothesis of improved stress resistance (diseases, temperatures). To increase genetic variation in this region and break potential linkage between beneficial and detrimental traits we propose to screen and characterize germplasm carrying different haplotypes in this genomic region.
Data availability
This study is based on datasets which are described in detail in Schreiber et al. (2023). Raw data files for both RNA-sequence data and whole genome shotgun data have been deposited at the European Nucleotide Archive (ENA) under the project numbers PRJEB49069 and PRJEB48903, respectively. SNP data is available through the Germinate platform https://ics.hutton.ac.uk/germinate-barn/. Gene expression data is available on the e!DAL platform under the preview link https://doi.ipk-gatersleben.de/DOI/815787d8-4036-408b-999e-725f7645eacf/6e532074-b4d7-4393-9221-8fe643e100f2/2/1847940088. Code used in this study is available at https://github.com/rwonneberger/Wonneberger_Chr5H_haplotype_barley.
References
Baker K, Bayer M, Cook N et al (2014) The low-recombining pericentromeric region of barley restricts gene diversity and evolution but not gene expression. Plant J 79:981–992. https://doi.org/10.1111/tpj.12600
Bayer MM, Rapazote-Flores P, Ganal M et al (2017) Development and evaluation of a barley 50k iSelect SNP Array. Front Plant Sci. https://doi.org/10.3389/fpls.2017.01792
Borovkova IG, Steffenson BJ, Jin Y et al (1997) Identification and mapping of a leaf rust resistance gene in barley line Q21861. Genome 40:236–241. https://doi.org/10.1139/g97-033
Botstein D, White RL, Skolnick M, Davis RW (1980) Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Genet 32:314–331
Bradbury PJ, Zhang Z, Kroon DE et al (2007) TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics 23:2633–2635. https://doi.org/10.1093/bioinformatics/btm308
Brbaklić L, Trkulja D, Mikić S et al (2021) Genetic diversity and population structure of Serbian barley (Hordeum vulgare L.) collection during a 40-year long breeding period. Agronomy 11:118. https://doi.org/10.3390/agronomy11010118
Casale F, Van Inghelandt D, Weisweiler M et al (2022) Genomic prediction of the recombination rate variation in barley—a route to highly recombinogenic genotypes. Plant Biotechnol J 20:676–690. https://doi.org/10.1111/pbi.13746
Contreras-Moreira B, Serrano-Notivoli R, Mohammed NE et al (2019) Genetic association with high-resolution climate data reveals selection footprints in the genomes of barley landraces across the Iberian Peninsula. Mol Ecol 28:1994–2012. https://doi.org/10.1111/mec.15009
Coulter M, Entizne JC, Guo W et al (2022) BaRTv2: a highly resolved barley reference transcriptome for accurate transcript-specific RNA-seq quantification. Plant J 111:1183–1202. https://doi.org/10.1111/tpj.15871
Danecek P, Auton A, Abecasis G et al (2011) The variant call format and VCFtools. Bioinformatics 27:2156–2158. https://doi.org/10.1093/bioinformatics/btr330
Dawson IK, Russell J, Powell W et al (2015) Barley: a translational model for adaptation to climate change. New Phytol 206:913–931. https://doi.org/10.1111/nph.13266
Donini P, Law JR, Koebner RMD et al (2000) Temporal trends in the diversity of UK wheat. Theor Appl Genet 100:912–917. https://doi.org/10.1007/s001220051370
Dreiseitl A, Steffenson BJ (2000) Postulation of leaf-rust resistance genes in Czech and Slovak barley cultivars and breeding lines. Plant Breed 119:211–214. https://doi.org/10.1046/j.1439-0523.2000.00495.x
Dziurdziak J, Podyma W, Bujak H, Boczkowska M (2022) Tracking changes in the spring barley gene pool in Poland during 120 years of breeding. Int J Mol Sci 23:4553. https://doi.org/10.3390/ijms23094553
Elmansour H, Singh D, Dracatos PM, Park RF (2017) Identification and characterization of seedling and adult plant resistance to Puccinia hordei in selected African barley germplasm. Euphytica 213:119. https://doi.org/10.1007/s10681-017-1902-8
Fang Z, Gonzales AM, Clegg MT, et al (2014) Two Genomic Regions Contribute Disproportionately to Geographic Differentiation in Wild Barley. G3 (Bethesda) 4:1193–1203. https://doi.org/10.1534/g3.114.010561
FAOSTAT (2022). https://www.fao.org/faostat/en/#home
Fischbeck G (2003) Chapter 3 - Diversification through breeding. In: Diversity in barley (Hordeum vulgare). pp 29–52
Franckowiak JD, Jin Y, Steffenson BJ (1997) Recommended allele symbols for leaf rust resistance genes in barley. Barley Genetics Newsletter 27:36–44
Friedt W, Horsley R, Harvey B, et al (2011) Barley breeding history, progress, objectives, and technology. pp 160–220
Giese H, Holm-Jensen AG, Jensen HP, Jensen J (1993) Localization of the Laevigatum powdery mildew resistance gene to barley chromosome 2 by the use of RFLP markers. Theoret Appl Genetics 85:897–900. https://doi.org/10.1007/BF00225035
Golegaonkar PG, Singh D, Park RF (2009) Evaluation of seedling and adult plant resistance to Puccinia hordei in barley. Euphytica 166:183–197. https://doi.org/10.1007/s10681-008-9814-2
Goudet J (2005) hierfstat, a package for r to compute and test hierarchical F-statistics. Mol Ecol Notes 5:184–186. https://doi.org/10.1111/j.1471-8286.2004.00828.x
Hagenblad J, Leino MW (2022) Chevalier barley: the influence of a world-leading malting variety. Crop Sci 62:235–246. https://doi.org/10.1002/csc2.20668
Harris RS (2007) Improved pairwise alignment of genomic DNA. PhD thesis
Hedden P (2003) The genes of the green revolution. Trends Genet 19:5–9. https://doi.org/10.1016/S0168-9525(02)00009-4
Henderson M (1945) Studies of sources of resistance and inheritance of reaction to leaf rust, Puccinia anomala Rostr., in barley. University of Minnesota
Hickey LT, Lawson W, Platz GJ et al (2012) Origin of leaf rust adult plant resistance gene Rph20 in barley. Genome 55:396–399. https://doi.org/10.1139/g2012-022
Jayakodi M, Padmarasu S, Haberer G et al (2020) The barley pan-genome reveals the hidden legacy of mutation breeding. Nature 588:284–289. https://doi.org/10.1038/s41586-020-2947-8
Jiao W-B, Schneeberger K (2020) Chromosome-level assemblies of multiple Arabidopsis genomes reveal hotspots of rearrangements with altered evolutionary dynamics. Nat Commun 11:989. https://doi.org/10.1038/s41467-020-14779-y
Jombart T (2008) adegenet: a R package for the multivariate analysis of genetic markers. Bioinformatics 24:1403–1405. https://doi.org/10.1093/bioinformatics/btn129
Jones H, Civáň P, Cockram J et al (2011) Evolutionary history of barley cultivation in Europe revealed by genetic analysis of extant landraces. BMC Evol Biol 11:320. https://doi.org/10.1186/1471-2148-11-320
Jørgensen IH (1992) Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica 63:141–152. https://doi.org/10.1007/BF00023919
Kamvar ZN, Tabima JF, Grünwald NJ (2014) Poppr: an R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ. https://doi.org/10.7717/peerj.281
Koebner R, Donini P, Reeves J et al (2003) Temporal flux in the morphological and molecular diversity of UK barley. Theor Appl Genet 106:550–558. https://doi.org/10.1007/s00122-002-1065-3
Kolodinska Brantestam A, von Bothmer R, Dayteg C et al (2007) Genetic diversity changes and relationships in spring barley (Hordeum vulgare L.) germplasm of Nordic and Baltic areas as shown by SSR markers. Genet Resour Crop Evol 54:749–758. https://doi.org/10.1007/s10722-006-9159-4
Laidig F, Piepho H-P, Rentel D et al (2017) Breeding progress, genotypic and environmental variation and correlation of quality traits in malting barley in German official variety trials between 1983 and 2015. Theor Appl Genet 130:2411–2429. https://doi.org/10.1007/s00122-017-2967-4
Malysheva-Otto L, Ganal MW, Law JR et al (2007) Temporal trends of genetic diversity in European barley cultivars (Hordeum vulgare L.). Mol Breeding 20:309–322. https://doi.org/10.1007/s11032-007-9093-y
Mascher M, Gundlach H, Himmelbach A et al (2017) A chromosome conformation capture ordered sequence of the barley genome. Nature 544:427–433. https://doi.org/10.1038/nature22043
Mascher M, Wicker T, Jenkins J et al (2021) Long-read sequence assembly: a technical evaluation in barley. Plant Cell 33:1888–1906. https://doi.org/10.1093/plcell/koab077
Mayer KFX, Waugh R, Langridge P et al (2012) A physical, genetic and functional sequence assembly of the barley genome. Nature 491:711–716. https://doi.org/10.1038/nature11543
McCouch S, Baute GJ, Bradeen J et al (2013) Feeding the future. Nature 499:23–24. https://doi.org/10.1038/499023a
Mehnaz M, Dracatos PM, Park RF, Singh D (2021) Mining middle eastern and central Asian barley germplasm to understand diversity for resistance to Puccinia hordei. Causal Agent of Leaf Rust Agronomy 11:2146. https://doi.org/10.3390/agronomy11112146
Milner SG, Jost M, Taketa S et al (2019) Genebank genomics highlights the diversity of a global barley collection. Nat Genet 51:319–326. https://doi.org/10.1038/s41588-018-0266-x
Milotova J, Martynov SP, Dobrotvorskaya TV, Vaculova K (2008) Genealogical analysis of the diversity of spring barley cultivars released in former Czechoslovakia and modern Czech Republic. Russ J Genet 44:51–59. https://doi.org/10.1134/S1022795408010079
Niks R, Walther U, Jaiser H et al (2000) Resistance against barley leaf rust (Puccinia hordei) in West-European spring barley germplasm. Agronomie 20:769–782. https://doi.org/10.1051/agro:2000174
Ovesná J, Kučera L, Vaculová K et al (2013) Analysis of the genetic structure of a barley collection using DNA diversity array technology (DArT). Plant Mol Biol Rep 31:280–288. https://doi.org/10.1007/s11105-012-0491-x
Paradis E, Schliep K (2019) ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35:526–528. https://doi.org/10.1093/bioinformatics/bty633
Parlevliet JE (1983) Race-specific resistance and cultivar-specific virulence in the barley-leaf rust pathosystem and their consequences for the breeding of leaf rust resistant barley. Euphytica 32:367–375. https://doi.org/10.1007/BF00021445
Patro R, Duggal G, Love MI et al (2017) Salmon provides fast and bias-aware quantification of transcript expression. Nat Methods 14:417–419. https://doi.org/10.1038/nmeth.4197
Purcell S, Neale B, Todd-Brown K et al (2007) PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet 81:559–575. https://doi.org/10.1086/519795
Rajala A, Peltonen-Sainio P, Jalli M et al (2017) One century of Nordic barley breeding: nitrogen use efficiency, agronomic traits and genetic diversity. J Agric Sci 155:582–598. https://doi.org/10.1017/S002185961600068X
Rasmusson DC, Phillips RL (1997) Plant breeding progress and genetic diversity from De Novo variation and elevated epistasis. Crop Sci. https://doi.org/10.2135/cropsci1997.0011183X003700020001x
Reinhold M, Sharp EL (1982) Virulence types of Puccinia hordei from North America, North Africa, and the middle East. Plant Dis 66:1009–1011
Russell JR, Ellis RP, Thomas WTB et al (2000) A retrospective analysis of spring barley germplasm development from `foundation genotypes’ to currently successful cultivars. Mol Breed 6:553–568. https://doi.org/10.1023/A:1011372312962
Sandhu KS, Forrest KL, Kong S et al (2012) Inheritance and molecular mapping of a gene conferring seedling resistance against Puccinia hordei in the barley cultivar Ricardo. Theor Appl Genet 125:1403–1411. https://doi.org/10.1007/s00122-012-1921-8
Schreiber M, Wonneberger R, Haaning AM et al (2023) Genomic resources for a historical collection of cultivated two-row European spring barley genotypes. bioRxiv. https://doi.org/10.1101/2023.03.06.531259
The International Barley Genome Sequencing Consortium (2012) A physical, genetic and functional sequence assembly of the barley genome. Nature 491:711–716. https://doi.org/10.1038/nature11543
Tondelli A, Xu X, Moragues M et al (2013) Structural and temporal variation in genetic diversity of European spring two-row barley cultivars and association mapping of quantitative traits. Plant Genome. https://doi.org/10.3835/plantgenome2013.03.0007
von Bothmer R, Sato K, Komatsuda T, et al (2003) Chapter 2 - The domestication of cultivated barley. In: von Bothmer R, van Hintum T, Knüpffler H, Sato K (eds) Diversity in Barley (Hordeum vulgare). Elsevier, pp 9–27
Weir BS, Cockerham CC (1984) Estimating F-statistics for the analysis of population structure. Evolution 38:1358–1370. https://doi.org/10.2307/2408641
Yahiaoui S, Cuesta-Marcos A, Gracia MP et al (2014) Spanish barley landraces outperform modern cultivars at low-productivity sites. Plant Breed 133:218–226. https://doi.org/10.1111/pbr.12148
Acknowledgements
The authors would like to thank Axel Himmelbach, Ines Walde, Susanne König at the sequencing team at IPK Gatersleben for generating genomic and transcriptomic data utilized in this project and Anne Fiebig for submitting sequences to the European Nucleotide Archive. The authors would like to acknowledge and thank current and former members of the research group Genomics of Genetic Resources at IPK Gatersleben for help with tissue sampling: Mary Ziems, Mark Timothy Rabanus-Wallace, Hélène Pidon, Sudharsan Padmarasu, Mingjiu Li, Jayavardhan Reddy Kunam, Mohammed Rafaqat, Mohammad Awais, Jaqueline Pohl, Susanne König, Ines Walde, Manuela Knauft, Manuela Kretschmann and Beate Kamm. The authors would also like to thank Shane Heinen at the University of Minnesota for help with tissue sampling and the University of Minnesota Genomics Center for generating transcriptomic data for the project. The authors acknowledge the Research/Scientific Computing teams at The James Hutton Institute and NIAB for providing computational resources and technical support for the "UK's Crop Diversity Bioinformatics HPC" (BBSRC grant BB/S019669/1), use of which has contributed to the results reported. Work was supported by Rural and Environment Science and Analytical Services Division of the Scottish Government.
Funding
Open Access funding enabled and organized by Projekt DEAL. The project received funding in frame of the ERA-CAPS Research Programme with funding of (i) the German partners to NS through German Research Foundation (DFG) under the project references MU 3589/1-1 | STE 1102/15-1 | WA 3336/4-1, (ii) the Scottish partner to RW through the Biotechnology and Biological Sciences Research Council (BBSRC; award number BB/S004610/1), (iii) the US partner to GJM through the National Science Foundation (NSF; award number 1844331).
Author information
Authors and Affiliations
Contributions
RWo generated and analysed datasets and wrote the manuscript, MS and AH generated datasets and commented on previous versions of the manuscript, GJM, RW and NS designed the experiment, supervised the work and commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Communicated by lihuihui.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Online Resource 1
List of cultivars in the European two-rowed spring barley panel including country of origin, release year, chromosome 5H haplotype and cluster membership at k=2 and k=3
Online Resource 2
Pairwise FST between different release periods
Online Resource 3
Analysis of Molecular Variance (AMOVA) statistics quantifying the extent of genetic variation found between and within groups of cultivars defined by release periods (1830-1959, 1960-1979, 1980-1999, 2000-2014)
Online Resource 4
Neighbor-joining tree using the genotypes of 1,509,447 SNP markers. The cultivars are color-coded according to the period in which they were released. The inner ring panel shows the chromosome 5H haplotype of each cultivar, the outer ring panels show the k-means cluster membership of each cultivar at k=2 (outermost ring panel) and k=3 (central ring panel)
Online Resource 5
Principal coordinate analysis of the European two-rowed spring barley panel using the genotypes of 1,509,447 SNP markers. Each dot represents a cultivar, color-coded according to geographic origin
Online Resource 6
Left: Cumulative percentage of variance explained by different numbers of PCs. Selecting the number of PCs at which ~95% of the cumulative variance were explained, 110 PCs were retained for k-means clustering. Right: Bayesian Information Criterion (BIC) values at different numbers of assumed subpopulations (k). Low BIC values indicate a good fit of the model to the data. Using the elbow method to determine the value for k at which the curve flattens, two or three subpopulations were assumed to be likely
Online Resource 7
k-means cluster membership of cultivars at k=2 (A) and k=3 (B) superimposed on PCoA (see Fig. 3)
Online Resource 8
Annotation, gene IDs and genetic positions in reference genomes Barke and Morex V3 as well as the reference transcriptome BaRTv2 of the 48 genes differentially expressed in haplotype 1 and 2. These 48 genes form a distinct group in a PCoA of all genes located between 69 and 320 Mbp on chromosome 5H (see Fig. 7, red clusters). All genes were highly expressed in cultivars carrying haplotype 2 except for the two genes marked with asterisks which were highly expressed in cultivars carrying haplotype 1 and low/not expressed in cultivars carrying haplotype 2
Online Resource 9
Heatmap of the expression of the genes between 69 and 320 Mbp on chromosome 5H forming a separate k-means cluster and showing low or no expression in cultivars carrying the old haplotype and higher expression in cultivars carrying the new haplotype in the crown tissue of one week old seedlings. An exception is BaRT2v18chr5HG233580 which shows the opposite pattern. Gene expression is not correlated with haplotypes 3 and 4.
Online Resource 10
Heatmap of the expression of the genes between 69 and 320 Mbp on chromosome 5H forming a separate k-means cluster and showing low or no expression in cultivars carrying the old haplotype and higher expression in cultivars carrying the new haplotype in the root tissue of one week old seedlings. An exception is BaRT2v18chr5HG233580 which shows the opposite pattern. Gene expression is not correlated with haplotypes 3 and 4
Online Resource 11
Heatmap of the expression of the genes between 69 and 320 Mbp on chromosome 5H forming a separate k-means cluster and showing low or no expression in cultivars carrying the old haplotype and higher expression in cultivars carrying the new haplotype in the developing inflorescence tissue. Exceptions are BaRT2v18chr5HG233570 and BaRT2v18chr5HG233580 which show the opposite pattern. Gene expression is not correlated with haplotypes 3 and 4
Online Resource 12
Heatmap of the expression of the genes between 69 and 320 Mbp on chromosome 5H forming a separate k-means cluster and showing low or no expression in cultivars carrying the old haplotype and higher expression in cultivars carrying the new haplotype in the peduncle tissue. Exceptions are BaRT2v18chr5HG233570 and BaRT2v18chr5HG233580 which show the opposite pattern. Gene expression is not correlated with haplotypes 3 and 4
Online Resource 13
Heatmap of the expression of the genes between 69 and 320 Mbp on chromosome 5H forming a separate k-means cluster and showing low or no expression in cultivars carrying the old haplotype and higher expression in cultivars carrying the new haplotype in the spikelet tissue. Exceptions are BaRT2v18chr5HG233570 and BaRT2v18chr5HG233580 which show the opposite pattern. Gene expression is not correlated with haplotypes 3 and 4
Online Resource 14
Heatmap of the expression of the genes between 69 and 320 Mbp on chromosome 5H forming a separate k-means cluster and showing low or no expression in cultivars carrying the old haplotype and higher expression in cultivars carrying the new haplotype in the developing grain tissue. Gene expression is not correlated with haplotypes 3 and 4
Online Resource 15
PCA of all cultivars in the panel, using 110,914 SNPs at 68.78 - 320.04 Mbp on chromosome 5H color-coded by the haplotype the cultivars carry in this region
Online Resource 16
Alleles of 10,000 randomly selected SNP markers in the haplotype region between 68.78 and 320.04 Mbp on chromosome 5H and the flanking 10 Mbp on each side. Red indicates the homozygous reference genotype (cultivar ‘Barke’), yellow indicates the homozygous alternative genotype, green indicates the heterozygous genotype, grey indicates missing data. White columns indicate regions without SNP data. Accessions are sorted according to haplotype (right bar: orange = haplotype 1, green = haplotype 2, blue = haplotype 3, red = haplotype 4)
Online Resource 17
a) Alleles of all SNP markers 30 Mbp upstream and 4 Mbp downstream of the start of the haploblock region on chromosome 5H (38.78-72.78 Mbp). b) Alleles of all SNP markers 4 Mbp upstream and 30 Mbp downstream of the end of the haploblock region on chromosome 5H (316.04-350.04 Mbp). c) Alleles of all SNP markers 0.5 Mbp upstream and 0.5 Mbp downstream of the start of the haploblock region on chromosome 5H (68.28-69.28 Mbp). b) Alleles of all SNP markers 0.5 Mbp upstream and 0.5 Mbp downstream of the end of the haploblock region on chromosome 5H (319.54-320.54 Mbp). Black lines indicate the start (a, c) and end (b, d) of the haploblock region, respectively. Red indicates the homozygous reference genotype (cultivar ‘Barke’), yellow indicates the homozygous alternative genotype, green indicates the heterozygous genotype, grey indicates missing data. White columns indicate regions without SNP data. Accessions are sorted according to haplotype (right bar: orange = haplotype 1, green = haplotype 2, blue = haplotype 3, red = haplotype 4)
Online Resource 18
Dotplots showing genome alignments of cv. Morex (carrying haplotype 1) and cv. Barke (carrying haplotype 2) of a) the entire haplotype region on chromosome 5H illustrating the structural variation at the start and end of the region, b) the start of the haplotype region illustrating an insertion in Barke followed by two inversions further downstream and c) the end of the haplotype region illustrating an insertion in Barke followed by a deletion or potential translocation
Online Resource 19
Dotplots showing pairwise alignments of the chromosome 5H haplotypes in genomes of nine cultivars/accessions carrying haplotype 1 or 2. Names of the cultivars/accessions are shown on the bottom and left. Accessions carrying haplotype 1 are ‘Golden Promise’, ‘Morex’, ‘HOR13821’, ‘HOR3081’ and ‘HOR3365’. Accessions carrying haplotype 2 are ‘Barke, ‘Igri’, ‘HOR13942’ and ‘RGT Planet’.
Online Resource 20
GO enrichment of the 1080 genes located between 68.78 and 320.04 Mbp on chromosome 5H. Shown are the GO IDs, GO terms, the number of annotated genes per term in BartV2.0, the number of genes observed per term, the number of genes expected per term, the p-value of gene enrichment and the GO category
Online Resource 21
PCAs of all domesticated barley accessions of the German Federal ex-situ gene bank at IPK Gatersleben. Red dots represent the 243 accessions carrying the same names as cultivars and landraces from the European two-rowed spring barley panel. The figure was created using the IPK Bridge Portal (IPK Gatersleben - BRIDGE Web Portal (ipk-gatersleben.de)
Online Resource 22
PCoAs of all domesticated barley accessions of the German Federal ex-situ gene bank at IPK Gatersleben. Accessions are color-coded by geographic origin. NZ=New Zealand
Online Resource 23
Clustering of domesticated barley accessions of the IPK gene bank at different k. While the approach is not suitable to clearly distinguish haplotype 1, haplotype 3 and other potential haplotypes clustering together, haplotype 2 is clearly distinguished at k >= 7
Online Resource 24
Geographical distribution of haplotype 2 in groups of landraces and cultivars characterized by different row-types and growth habits in the IPK gene bank collection. Colors indicate the frequency of gene bank accessions from a given country carrying haplotype 2. Countries from which less than 15 accessions per respective category are present in the gene bank are shown in grey
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Wonneberger, R., Schreiber, M., Haaning, A. et al. Major chromosome 5H haplotype switch structures the European two-rowed spring barley germplasm of the past 190 years. Theor Appl Genet 136, 174 (2023). https://doi.org/10.1007/s00122-023-04418-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00122-023-04418-7