
Abstract
Key message
Eight soybean genomic regions, including six never before reported, were found to be associated with resistance to soybean rust (Phakopsora pachyrhizi) in the southeastern USA.
Abstract
Soybean rust caused by Phakopsora pachyrhizi is one of the most important foliar diseases of soybean [Glycine max (L.) Merr.]. Although seven Rpp resistance gene loci have been reported, extensive pathotype variation in and among fungal populations increases the importance of identifying additional genes and loci associated with rust resistance. One hundred and ninety-one soybean plant introductions from Japan, Indonesia and Vietnam, and 65 plant introductions from other countries were screened for resistance to P. pachyrhizi under field conditions in the southeastern USA between 2008 and 2015. The results indicated that 84, 69, and 49% of the accessions from southern Japan, Vietnam or central Indonesia, respectively, had negative BLUP values, indicating less disease than the panel mean. A genome-wide association analysis using SoySNP50K Infinium BeadChip data identified eight genomic regions on seven chromosomes associated with SBR resistance, including previously unreported regions of Chromosomes 1, 4, 6, 9, 13, and 15, in addition to the locations of the Rpp3 and Rpp6 loci. The six unreported genomic regions might contain novel Rpp loci. The identification of additional sources of rust resistance and associated genomic regions will further efforts to develop soybean cultivars with broad and durable resistance to soybean rust in the southern USA.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Soybean rust (SBR) caused by Phakopsora pachyrhizi Syd. is one of the most important foliar diseases of soybean [Glycine max (L.) Merr.]. This obligate biotrophic fungus has spread from East Asia to every continent where soybean is grown. After becoming established in South America, it was first observed in the continental USA in November 2004 (Schneider et al. 2005). Severe epidemics can result in yield losses of 70% or more by slowing pod filling and reducing seed weight (Ogle et al. 1979; Kumudini et al. 2008). Although SBR is less of a threat in the USA than it is in parts of South America, it caused > 60% yield loss in unprotected soybean fields in southern Alabama in 2012 (Sikora et al. 2014). In 2013, yield losses of up to 40% occurred in some fields in Alabama and Mississippi (Allen et al. 2014), and statewide yield losses to SBR in Alabama were 4% in 2020 (Allen et al. 2021; Sikora and Conner 2021). While losses can be reduced with timely fungicide applications, this adds to production costs. In addition, extensive use of fungicides in Brazil has diminished the efficacy of some classes of fungicides (Klosowski et al. 2016; Dalla Lana et al. 2018). Soybean cultivars with SBR resistance could therefore play a valuable role in more sustainable disease management strategies.
Resistance to SBR has been reported in G. max and other Glycine species, including the perennial species G. tomentella (Bromfield and Hartwig 1980; Hartman et al. 1992). The appearance of resistance reactions can vary considerably, but reactions to SBR have often been classified into three main infection types (Bromfield et al. 1980). Susceptible plants develop a “TAN” infection type, with two to four uredinia per lesion and profuse sporulation, particularly on the abaxial side of the leaf. A Type 0 infection type is characterized by the absence of macroscopically visible lesions or urediniospores and essentially indicates an immune reaction. Type 0 reactions are uncommon; most resistant soybean hosts have incomplete resistance and develop reddish-brown (RB) lesions, with zero to two uredinia and limited urediniospore production. The intensity of sporulation from both RB- and TAN-type lesions can be influenced by temperature and moisture, as well as by the genotype of the host and aggressiveness of the pathogen (Bromfield 1984).
Race-specific soybean resistance to SBR is conditioned by Rpp genes from at least seven reported loci (Bromfield and Hartwig 1980; Childs et al. 2018a; Li et al. 2012). Of these, the Rpp1, Rpp3, and Rpp5 loci are known to have more than one resistance allele (Garcia et al. 2008; Chakraborty et al. 2009; Harris et al. 2015). Most resistance alleles are dominant but recessive alleles have also been reported (Calvo et al. 2008; Garcia et al. 2008). In some cases, the degree of dominance depends on the allele in the susceptible parent (Garcia et al. 2011; Lemos et al. 2011). Due to the extent of pathogenic diversity among P. pachyrhizi populations, no Rpp gene conditions universal resistance (Bromfield et al. 1980; Bonde et al. 2006). Investigations of the virulence and aggressiveness of P. pachyrhizi isolates from the southern USA have revealed extensive pathotype diversity (Pham et al. 2009; Twizeyimana and Hartman 2012; Walker et al. 2011, 2014b). Confirmation that resistance is effective against fungal populations in more than one location and growing season is therefore needed to ensure broad resistance.
Although more than 100 plant introductions (PIs) have been reported to be resistant to P. pachyrhizi in the USA, the resistance genes from many of those PIs reside at either the Rpp1, Rpp3 or Rpp5 locus (Garcia et al. 2008; Pierozzi et al. 2008; Kendrick et al. 2011; Ray et al. 2011; Hossain et al. 2015). Harris et al. (2015) found that SBR resistance genes in 52 out of 75 PIs mapped to the Rpp3 region on chromosome (Chr) 6 and that 37 of the 52 PIs had a SNP haplotype identical to that of PI 462312, the accession in which the Rpp3 locus was first discovered. The high percentage of PIs with a resistance allele at one of these loci limits the number of potential resistance gene pyramids, or combinations of Rpp genes that could provide broader and more durable resistance (Lemos et al. 2011; Yamanaka et al. 2013; Yamanaka and Hossain 2019).
Prior to the first report of SBR in the continental USA in late 2004, Miles et al. (2006) screened 16,595 PIs from the USDA Soybean Germplasm Collection for resistance to a mixture of P. pachyrhizi isolates from Thailand, Brazil, Paraguay, and Zimbabwe. Seedling assays were conducted in a biosafety level 3 greenhouse, and based on the criteria used to define resistance, the authors concluded that 805 accessions (< 4.9%) had resistance to the four isolates. When the 805 PIs were tested in the field between 2006 and 2008, however, only 64 (8%) were confirmed to be resistant to seven southern US populations of the rust fungus (Walker et al. 2011). Furthermore, few of the PIs that Miles et al. (2008) subsequently found to be resistant in South America were also resistant to the US populations (Walker et al. 2011, 2014a, 2014b).
Genome-wide association studies (GWAS) to search for significant genotype–phenotype associations using single nucleotide polymorphism (SNP) markers have been successfully used to understand genetic architecture in panels of germplasm lines and to identify regions of the soybean genome associated with phenotypic variation in traits like resistance to soybean cyst nematode (Heterodera glycines), Phytophthora sojae, and other soybean pathogens (Guo et al. 2006; Ray et al. 2011; Hwang et al. 2014; Vaughn et al. 2014; Chang et al. 2016; Qin et al. 2017). The GWAS results can also be used to identify germplasm with traits of interest and SNP markers for marker-assisted selection in breeding programs.
Objectives of this study were to (1) evaluate soybean germplasm accessions for resistance to field populations of P. pachyrhizi under in the southeastern USA and (2) identify genomic regions associated with soybean rust resistance in those accessions, many of which had been reported to be susceptible to four foreign isolates. Germplasm accessions from a variety of countries and regions were screened initially, but screening was later focused primarily on PIs from Vietnam, southern Japan, or central Indonesia after preliminary evaluations showed that most of the accessions with resistance originated from those three geographic regions.
Materials and methods
Materials and experimental design
Two hundred and fifty-six germplasm accessions from Japan, Indonesia, Vietnam, China and several other countries were screened for their reactions to SBR in multiple year-location field tests, along with susceptible controls and PIs with Rpp genes at known loci (Tables 1 and S1). Most of the accessions tested were chosen specifically because they had had either susceptible or mixed reactions (i.e., both TAN and RB reactions) to four foreign isolates in earlier greenhouse assays (Miles et al. 2006). After the extent of pathotype diversity of P. pachyrhizi populations in different countries became apparent (Miles et al. 2008; Twizeyimana et al. 2008), however, we hypothesized that some of the accessions susceptible to the foreign isolates would have resistance to US pathotypes of the fungus. Each accession tested in a certain year was planted at two locations that year (Table 1). PIs with low-to-moderate disease in the first year they were tested were re-evaluated in the following year to confirm resistance, while PIs that were clearly susceptible were not retested in subsequent years unless they had been reported to have an Rpp gene that was effective in South America. The 2008 tests included accessions from more diverse geographical origins and maturity groups (MGs 0 to X) than the tests planted in subsequent years. After 2008, most of the accessions screened originated from southern Japan, Vietnam, or central Indonesia, and were in MGs IV through IX. This was because data from the earlier screenings had shown that few PIs from any other regions were resistant in the southern USA. Seeds were obtained from the USDA Soybean Germplasm Collection in Urbana, IL, USA.
Susceptible public cultivars representing MGs IV through VII were included in each field tests to provide an indication of SBR disease pressure and development in each year-location environment. Ten susceptible checks were planted in each test, and altogether 16 different susceptible checks were tested during the study. Ten germplasm accessions with known Rpp genes were generally planted in each test also to permit detection of possible pathotype differences among P. pachyrhizi populations in different years and locations. PIs carrying the Rpp1 through Rpp6 genes were planted, including Rpp1, Rpp2 and Rpp5 accessions with different resistance alleles. The Japanese cultivar Hyuuga (PI 506764), which has resistance alleles at the Rpp3 and Rpp5 loci (Kendrick et al. 2011), was also included in each field test.
Field tests were conducted in the southeastern USA between 2008 and 2016 (Table 1). The field tests were planted in at least two locations most years, and usable SBR reaction data were obtained from 14 different year-location environments. Screening nurseries were planted at the University of Florida’s North Florida Research and Education Center in Quincy, Florida, and at the University of Georgia Attapulgus Research and Education Center in southwestern Georgia every year of the study. Tests were also planted at Auburn University’s Gulf Coast Research and Extension Center in Fairhope, Alabama, in 2008 and 2009, and at Louisiana State University’s Central Research Station in Baton Rouge in 2008.
The field tests in Georgia were planted in mid-July, and the tests at most other locations were planted in early to mid-August to delay flowering until precipitation levels became more favorable for P. pachyrhizi and disease development later in the growing season. The field plots consisted of single rows arranged with a randomized complete block design, usually with two replications. Depending on the test site, rows ranged from 1.2 to 2.0 m in length, with 0.9 m between rows. Planting and field management methods used for the 2008 to 2013 tests were identical to those described in Walker et al. (2011, 2014b), and the 2015 and 2016 tests in Quincy, FL, were conducted the same way as earlier tests at that location. In Attapulgus, GA artificial lighting was used during the nighttime for one month after emergence of seedlings to delay plant maturation, and agricultural streptomycin was applied to the plots there to reduce the incidence of bacterial pustule (Xanthomonas axonopodis pv. glycines), as described in Walker et al. (2011).
Rating of reactions to SBR in field evaluations
Infection with P. pachyrhizi at the SBR screening nurseries occurred naturally through infection with urediniospores from nearby soybean fields and patches of kudzu (Pueraria lobata). SBR disease severity was rated either on plants in the field, or on leaflets collected from test plots between mid-October and early December, when SBR symptoms and signs (i.e., urediniospores) were visible on susceptible checks. At least 10 leaflets were collected randomly from the mid-canopy, placed in labeled bags, and kept in a refrigerator until they could be examined with a compound microscope. Reactions to SBR in some test locations were evaluated using different rating methods and scales in 2008, when tests were grown in four different states. The scales and methods used at each location between 2008 and 2011 were described in Walker et al. (2014b), and the methods used at Quincy in 2011 were also used for the 2013 to 2015 assays. The inclusion of rating data from susceptible checks helped to mitigate differences in rating methods and persons. Data from a test site were used only if SBR severity and sporulation were high on plants known to be susceptible.
For both severity and sporulation, higher rating values indicated more disease. SBR severity was rated on plants at each nursery location, usually on a scale of 1–5. A 1–9 scale was used at Baton Rouge in 2008, however, and the 2008 test in Fairhope, AL was rated once using a 0 to 8 scale, as described in Walker et al. (2011, 2014b). The ratings from the longer scales were subsequently converted to a 1 to 5 scale so that those ratings could be included in the calculation of the best linear unbiased predictor (BLUP) value for each of the soybean PIs with rating data from multiple locations. For SBR severity, a rating of 1 meant that no rust lesions were visible, while a rating of 5 indicated that the density of SBR lesions on plants or leaf samples was as high as that observed on susceptible checks from the same field test. In Quincy, FL and in some other nurseries, sporulation (i.e., accumulation of urediniospores on the abaxial side of infected leaves) was also rated on a 1 to 5 scale in which 1 meant that no urediniospores were visible, and 5 indicated profuse sporulation, equivalent to that seen on leaves of the susceptible checks. When rating data for both sporulation and SBR severity were collected, a rust index (RI) value was calculated from the two types of ratings as described in Walker et al. (2014a, b). The RI rating was the square root of the product of the severity and sporulation ratings, so it was also in the range of 1.0–5.0.
Genotype data and statistical analyses
SNP marker data from 285 accessions that had previously been genotyped with the Illumina SoySNP50K Beadchip (Song et al. 2015) were downloaded from the SoyBase (Grant et al. 2010). SNPs with a minor allele frequency (MAF) less than 0.05 were filtered from the dataset, leaving 31,114 SNPs that were used to analyze population structure and carry out the GWAS.
To assess the degree of population structure, principal coordinate analysis (PCoA) was conducted in the GAPIT Version 3 R package (Wang and Zhang 2020). Eigenvalues from the GAPIT output were used to calculate the variation explained by each principal coordinate (PC). A scatter plot of the first two PCs was created with the ggplot2 R package (Wickham 2016). A neighbor joining tree was calculated in TASEL 5 (Bradbury et al. 2007) and visualized with FigTree Version 1.1.4 software (http://tree.bio.ed.ac.uk/software/figtree/). The 20 accessions with the lowest disease ratings (all with BLUPs less than − 0.756) were labeled in the PC plot to facilitate the assessment of the importance of geographic sources of resistance in the panel. Accession names in the PCoA plot and neighbor joining tree were colored based on the country of origin of each PI to make visual assessment more informative.
BLUP values were calculated from the disease ratings of PIs using JMP Pro 14 (JMP, SAS Institute Inc., Cary, NC, USA), which permitted us to overcome the limitations of an incomplete dataset and the differences among test environments. BLUP values were calculated by treating genotype, environment, and genotype × environment interactions as random variables using the Standard Least Squares and restricted maximum-likelihood (REML) methods (Shaw 1987). The BLUP values were then used in the GWAS analysis to identify genomic regions associated with SBR resistance, similarly to the methods described by Steketee et al. (2020).
A GWAS analysis was conducted using the Fixed and random model Circulating Probability Unification (FarmCPU) method (Liu et al. 2016a, b), which eliminates some false positives by running iterations of a fixed-effect model to test markers and a random effects model to define kinship using multiple associated markers. The first three principal coordinates were included in the GWAS model to account for covariance between population structure and phenotype. To determine significant SNPs and correct for multiple testing, the false discovery rate (FDR) for each marker was included in the FarmCPU output. SNPs with an FDR less than 0.05 (p < 1.445 × 10–5) were considered significant.
Results
Disease evaluations
Informative data were obtained from field evaluations in 2008, 2009, 2011, 2012, 2013, 2015, and 2016. Altogether, 256 PIs with unknown or inconclusive resistance to US P. pachyrhizi populations were screened in at least two different year-location environments. Disease data were also collected on the 16 susceptible check cultivars and 16 accessions with mapped Rpp resistance genes. We added PI 605823 to the latter group after the discovery of the Rpp7 gene locus. In fact, we developed the populations that Childs et al. (2018b) used to map Rpp7 after early data from this study showed that PI 605823 was resistant in our field test locations.
SBR severity and urediniospore production on susceptible checks demonstrated that disease pressure was generally high by the time the PI reactions were rated in each test. Rust index (RI) ratings for susceptible checks were between 3.5 and 5.0 in most tests, resulting in positive BLUP values that were often higher than 1.00 (Table 2). In contrast, RI ratings on highly resistant Rpp gene differentials like PI 200492 (Rpp1) and PI 567102B (Rpp6) were typically between 1.0 and 2.0, and BLUP values less than − 1.00. Differences in the reactions of some accessions with known Rpp genes between test environments indicated pathotype variation among some of the P. pachyrhizi populations.
BLUP values calculated from the disease ratings ranged from − 1.46 (least disease) to 1.60 (most disease). One hundred and thirty-eight accessions had BLUP values of − 0.10 or lower, and the seven most resistant accessions had BLUPs less than − 1.00 (Tables 2 and S1). The nine lines with the least disease were PI 567104B (Rpp6), PI 200492 (Rpp1), PI 567090, PI 567102B (Rpp6), PI 200532, PI 567061, PI 635999, PI 605823 (Rpp7) and PI 200547 (Table 2). PI 635999 is the Vietnamese cultivar ‘DT 2000’, which has alleles at the Rpp3 and Rpp4 loci. Of the 99 accessions with BLUP values lower than -0.40, 44 were from Japan, 21 were from central Indonesia (mainly Java), and 24 were from northern Vietnam. Among the few resistant accessions not from those areas were PI 518295 from Taiwan and PI 476905A from an unknown location in China. Of the other 23 accessions from China, only PI 594796 and PI 594742 had negative BLUP values, and those were only − 0.07 and − 0.01, respectively (Table S1).
Other effective genes included the Rpp3 gene in PI 462312, the allele(s) of Rpp2 in PI 230,970 and PI 417125, the Rpp5 alleles of PI 200487 and PI 471904, and at least one of the two Rpp genes in ‘Hyuuga’ (PI 506764) (Table 2). The Rpp4 gene from PI 459025B was not effective against most of the P. pachyrhizi populations. Other highly resistant accessions with no known Rpp genes included PI 567090, PI 567061, PI 566984, PI 566975 and PI 567046 from central Indonesia; and PI 200532, PI 200547 and PI 416806 from southern Japan (Table 2).
Accessions known to have resistance at the Rpp1, Rpp3, Rpp4 and Rpp5 loci showed wide variation in the amount of disease that they developed, indicating different resistance alleles (Tables 2 and S1). Unlike the highly effective Rpp1 gene of PI 200492, the Rpp1 allele in PI 561356, the Rpp1-b gene from PI 594538A, and the Rpp1 allele(s) of PI 594760B and PI 594767A did not protect plants against the pathotypes encountered. The Rpp3 allele in PI 567099A was also ineffective compared to the Rpp3 gene in PI 462312. The Rpp5 allele(s) in PI 200487 from southern Japan and in PI 471904 from East Java provided resistance, whereas the Rpp5 allele from PI 200526 and the recessive rpp5 allele from PI 200456 did not suppress disease in those two Japanese accessions (Table S1). The Rpp4 allele that Harris et al. (2015) detected in PI 605791A also conditioned resistance, unlike the allele of the Rpp4 gene of PI 459025B (Silva et al. 2008). Rpp6 alleles in PI 567068A, PI 567076 and PI 567129 conditioned resistance but were less effective than the original Rpp6 gene(s) in PI 567102B and PI 567104B. At least five PIs with single Rpp genes had less disease than the cultivar Hyuuga (PI 506764), which has resistance alleles at the Rpp5 and Rpp3 loci (Table 2). It is possible that the Rpp5 allele in Hyuuga contributed little to resistance against the fungal pathotypes encountered.
Population structure in the germplasm panel
The principal coordinate analysis (PCoA) (Fig. 1) and the dendrogram generated by the neighbor-joining analysis (Fig. 2) revealed population structure in the panel of germplasm accessions. In the color-coded scatter plot of the first and second PCs, clustering of lines was strongly influenced by geographical origin, with separation between most of the PIs from Japan, Indonesia, and Vietnam (Fig. 1). The 20 most-resistant accessions, based on their low BLUP values, showed a strong degree of clustering along PC 1. Ten of these accessions were from Java, eight were from southern Japan, and two were from northern Vietnam. In the neighbor-joining tree, most of the accessions from southern Japan formed a cluster that was separate from the PIs from Indonesia, Vietnam and China (Fig. 2). The majority of the accessions from Indonesia, China and Vietnam also formed distinct clusters, though there were occasional exceptions to this origin-based clustering, especially for some accessions from Vietnam. The presence of population structure was therefore accounted for in the GWAS model.

Plot of principal coordinate analysis for a panel of soybean accessions evaluated for their reactions to soybean rust in the southeastern USA. Dots representing plant introductions (PIs) are color-coded based on country of origin. Clusters of PIs from Japan, Indonesia and Vietnam are mostly independent from one another, and the susceptible checks from the USA also formed an independent cluster

Dendrogram depicting the genetic relationship among soybean plant introductions in a panel of germplasm accessions evaluated for their reactions to soybean rust in the southeastern USA. The grouping patterns also indicate that the majority of the accessions from Vietnam, Indonesia and especially Japan formed distinct groups based on country of origin. The genetic similarities among US cultivars used as checks in the disease assays are also evident
Genomic regions associated with resistance to SBR
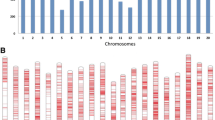
GWAS analysis using the BLUP disease rating values and 31,114 SNP markers resulted in the detection of eight significant SNPs in eight genomic regions on seven chromosomes (P < 1.445 × 10–5; − log10(P) = 4.8) (Fig. 3; Table 3). Single genomic regions were detected on Chrs 1, 4, 9, 15, and 18, and two distinct regions were found on Chr 6 (Fig. 3). The significant SNPs were also evident in the quantile–quantile (QQ) plot of the expected vs. observed p-values for the SNPs used in the GWAS (Fig. 4). The effect of the favorable allele at the significant SNPs (i.e., the allele in most of the resistant PIs) ranged from − 0.24 to − 0.10 (Table 3). The significant SNP marker with the highest significance level (ss715594707) was located at position 47,460,008 on Chr 6 in the Wm82.a2 reference assembly, which is approximately 2–3 Mb from the estimated location of the Rpp3 locus (Fig. 3; Hyten et al. 2009). The R2 value for this region was 0.23, considerably higher than those of the other genomic regions, which ranged from 0.02 to 0.08 (Table 3). SNP marker ss715594707 had a MAF of 0.25 in the panel. A second significant region on Chr 6 (ss715594035) was detected at position 2,748,236, a region not previously reported to be associated with SBR resistance, perhaps because its R2 contribution was only 0.02.

Manhattan plot generated from a genome-wide association analysis of a panel of soybean accessions evaluated for their reactions to soybean rust in the southeastern USA. The X-axis shows the location of SNPs along each chromosome in the genome, and the Y-axis shows the − log10 of the p-values. The significance threshold was − log10(P) = 4.84

Quantile–quantile (QQ) plot of expected vs. observed p-values for each SNP marker used in the GWAS analysis
Besides the two regions detected on Chr 6, the markers with the highest − log10(P) values, in descending order of probability, were on Chrs 4, 9, 15, 13, 1, and 18 (Table 3). The effects of the favorable alleles at the SNP markers in those genomic regions ranged from − 0.192 for the marker on Chr 4 to − 0.134 for the marker on Chr 18 (Table 3). The Chr 18 marker, ss715632525, is in a region of Chr 18 that contains the Rpp6 locus discovered in PI 567102B and PI 567104B (Li et al. 2012; Liu et al. 2016a, b) (Tables 2, 3 and S1). No Rpp loci have been reported previously in the other five genomic regions with significant markers.
Discussion
This study demonstrated that (1) numerous soybean PIs previously reported to be susceptible to P. pachyrhizi isolates from other continents were actually resistant to field populations of P. pachyrhizi in the southern USA; (2) the majority of the resistant accessions originated from either Japan, Vietnam or Indonesia; and (3) GWAS analysis of BLUP values calculated from disease reactions in multiple years and locations could be used to identify eight genomic regions putatively associated with resistance to SBR. Six of those regions have not previously been reported as far as we know.
Rpp genes and genomic regions of the soybean genome associated with resistance to SBR have been identified in previous studies using biparental molecular mapping, bulked segregant analysis or GWAS with seedling reaction data from greenhouse assays (Chang et al. 2016; Garcia et al. 2008; Harris et al. 2015; Hyten et al. 2007, 2009). In most of those assays, plants were inoculated with an isolate collected in a single location and growing season. To our knowledge, this is the first GWAS of SBR resistance that used field data from multiple years and locations, and it is only the second GWAS study for SBR resistance. Unlike greenhouse assays, in which seedling reactions are typically classified into three infection types two weeks after inoculation, our plant reaction data provided assessments of the resistance of adult plants to different field populations of the rust fungus based on sporulation intensity and disease severity. Calculation of BLUP values from the semi-quantitative disease ratings allowed us to obtain estimates of the relative resistance of the germplasm accessions from an unbalanced data set collected in multiple locations over several years. GWAS analysis of the BLUP values made it possible to identify regions of the soybean genome associated with resistance to P. pachyrhizi pathotypes prevalent in the southern USA, including regions containing the Rpp3 and Rpp6 resistance genes.
The disease ratings demonstrated SBR resistance in at least 132 germplasm accessions that Miles et al. (2006) had previously reported to be susceptible or only partly resistant to four foreign P. pachyrhizi isolates. This was less surprising after the extent of pathotype diversity among and within P. pachyrhizi populations became evident (Pham et al. 2009; Twizeyimana et al. 2009; Akamatsu et al. 2013). The BLUP values calculated from the disease rating data indicated the relative levels of resistance or susceptibility of the accessions, and several of the PIs with the lowest BLUP values also had high levels of resistance in previous field and greenhouse tests conducted in the southern USA (Walker et al. 2011, 2014a, 2014b). The reactions of some of the PIs also showed that pathotypes of P. pachyrhizi populations from fields in the southern USA differ from some South American pathotypes (Garcia et al. 2008; Miles et al. 2008). This illustrates the importance of verifying that soybean germplasm has resistance to local and regional populations of the rust fungus.
Phakopsora pachyrhizi populations have high levels of pathogenic diversity within and among populations (Yamaoka et al. 2002; Akamatsu et al. 2013; Twizeyimana and Hartman 2012). It is therefore important to identify germplasm with Rpp genes that condition resistance to local pathotypes across years and locations. Data from Stone et al. (2022) reaffirmed that the pathotypes of the Fort Detrick isolates used by Miles et al. (2006) are very different from pathotypes in US P. pachyrhizi populations. After finding that only about 50 of the 805 PIs that Miles et al. (2006) selected were resistant to P. pachyrhizi pathotypes in the southern USA (Walker et al. 2011), we hypothesized that some accessions with susceptible or mixed reactions to the four foreign isolates would have resistance to at least some US pathotypes. This study proved that hypothesis to be correct; 84 accessions that Miles et al. (2006) reported to be susceptible to SBR had BLUP values ≤ − 0.10 and can thus be considered resistant.
Soybean cultivars with pyramids of two or more Rpp genes are likely to have broader and more durable resistance than cultivars with single Rpp genes (Yamanaka et al. 2013; Yamanaka and Hossain 2019). Since many of the known resistant soybean accessions have Rpp genes at the same loci, however, it is important to continue searching for additional Rpp loci and novel alleles. We focused primarily on evaluating PIs originating from Japan, Vietnam and Indonesia after results from early tests revealed that most of the resistant accessions with resistance in the USA were from those regions (Walker et al. 2011, 2014b). In addition, most of the named Rpp genes were discovered in PIs from southern Japan (Rpp1, Rpp2 and at least three alleles at the Rpp5 locus), central Indonesia (Rpp6), or Vietnam (Rpp7). Although PI 462312 (‘Ankur’), the source of the Rpp3 gene, was selected in Uttar Pradesh in north-central India, it was selected from a cross between unknown parents made in the USA, so the original source of the resistance gene is not known (Germplasm Resources Information Network). In this study, 84% of the accessions from Japan, 69% of those from Vietnam, and 49% of those from Indonesia had negative BLUP values, compared to only 11% of the 27 Chinese PIs tested. Nevertheless, some PIs from Japan, Vietnam or central Indonesia were susceptible in our tests; the Japanese accessions PI 200456, PI 200526 (Shiranui) and PI 224270 (Hougyoku) had positive BLUP values, even though Yamanaka et al. (2010) had reported that they were highly resistant to a bulk fungal isolate from central Japan. In contrast, PI 200487 was resistant in central Japan and also in our field tests.
PI 635999 (‘DT 2000’) and PI 423972 were also resistant in field studies conducted in Hanoi, Vietnam, in 2006 and 2007 (Pham et al. 2010). Some other accessions were susceptible in our tests but showed resistance to field populations of P. pachyrhizi in northern Vietnam. Among those were the Rpp differentials PI 230970 (Rpp2), which had an RB reaction in Hanoi, and PI 459025B (Rpp4), which had an intermediate reaction there. In contrast, PI 200492 (Rpp1), PI 462312 (Rpp3) and PI 417089A (allele at Rpp3 locus) were resistant in our tests but developed TAN infection types in Hanoi (Pham et al. 2010). The reactions of some Rpp gene differentials in 2006 field tests in eastern Paraguay also differed from their reactions in our tests (Miles et al. 2008). PI 200492 and PI 615437, which has an Rpp3 resistance allele, were both susceptible in Paraguay, whereas PI 230970 and some PI 567099A plants (with the recessive rpp3 allele) were resistant there. These results attest to the pathogenic diversity among P. pachyrhizi populations.
Accessions from other countries that were resistant in our tests included PIs 368039, 379621 and 518295 from Taiwan, PI 423972 from Nepal, and PIs 203398, 417503 and 628932 from Brazil. PI 368039 was one of the few resistant PIs from this study that was also highly resistant to an unpurified rust isolate from central Japan (Yamanaka et al. 2010). The resistant accession PI 476897 is reported to be from China but was obtained from a germplasm collection in Hanoi, Vietnam. Harris et al. (2015) reported that PI 518295 has a resistance allele at the Rpp1 locus, PI 417503 has one at the Rpp3 locus, and PI 476905A has a resistance gene at the Rpp6 locus. It was somewhat surprising that very few of the accessions from southern China were resistant in our tests, especially since at least seven of them have Rpp genes that are effective against foreign isolates and populations of the fungus (Supplemental Table S1). Six of 24 Chinese PIs screened are known or thought to have a resistance gene at the Rpp1 locus, but like the Rpp1-b gene of PI 594538A and the Rpp1 allele from PI 561356, none of their alleles provided resistance in the southern USA (Supplemental Table S1; Walker et al. 2011, 2014b). A few other Chinese accessions in our study had negative BLUP values that were only slightly less than zero, so they are unlikely to be of value as sources of SBR resistance genes.
A high percentage of soybean accessions with resistance in the USA have a resistance allele at the Rpp3 locus (Harris et al. 2015), and many of them originated from southern Japan (Supplemental Table S1). At least 14 (28%) of the accessions with the 50 lowest BLUP values in the present study have a resistance allele at the Rpp3 locus, so it is not surprising that a marker near this locus had the highest significance level in the GWAS (Fig. 3). Alleles at the Rpp3 locus provided resistance to accessions from Indonesia (e.g., PI 567046A and PI 567034), Vietnam (PI 635999) and several from Japan, such as PI 416826A and PI 200488. Hyuuga and PI 462312 had similar BLUP means (− 0.64 and − 0.66), suggesting that the Rpp5 allele in Hyuuga may not have enhanced resistance against US rust populations. Resistance genes at the Rpp6 locus on Chr 18 appear to be less common, but the allele from PI 567102B and PI 567104B conditioned very high levels of resistance, resulting in the lowest and fourth lowest BLUP means. PI 567090, which had the third lowest BLUP value, has a resistance allele at the Rpp3 locus and another on Chr 18, likely at the Rpp6 locus (Harris et al. 2015).
The reactions of most of the Rpp gene differentials to field populations in this study differed considerably from the reactions of these accessions to a diverse collection of international and pre-2005 US isolates in a recent study by Stone et al. (2022). In that study, all 16 isolates defeated the Rpp1 gene in PI 200492 to some extent, even though PI 200492 had one of the lowest BLUP values in the present study. In contrast, the Rpp2 and Rpp4 genes, which were ineffective in this study, conditioned resistance to all or most of the 14 isolates in the Stone et al. (2022) assays, as did the Rpp1-b gene from PI 594538A and the Rpp1 alleles in three other accessions from China. PI 567102B (Rpp6) was the only highly resistant accession from the present study that also had resistance to most of the 16 isolates. These results and those of Pham et al. (2009) indicate a possible pathotype shift between the founder populations of P. pachyrhizi discovered in late 2004 and the predominant pathotypes of fungal populations along the Gulf Coast of the USA a few years later.
Detection of the genomic regions containing the Rpp3 and Rpp6 loci demonstrated that the GWAS methods used (i.e., the FarmCPU model and the FDR significance threshold) were effective. Of the eight genomic regions that were significant at P < 1.445 × 10–5; − log10(P) = 4.8, only ss715594707 on Chr 6 and ss715632525 on Chr 18 were locations with known Rpp loci. The former is near the Rpp3 locus, a common location for genes that condition resistance to SBR in the USA (Supplemental Table S1; Harris et al. 2015), and the latter is near the Rpp6 locus. Since at least four of the 20 PIs with the lowest BLUP values (ranging from − 1.457 to − 0.754) are known or thought to have an SBR resistance gene at the Rpp3 locus (Harris et al. 2015; Vuong et al. 2016), it was not surprising that a marker near the locus was significantly associated with disease. In contrast, resistance alleles at the Rpp6 locus have only been reported in a few PIs, but those alleles have conditioned much higher levels of resistance than any known Rpp3 alleles. This was demonstrated by the low BLUP values of the Indonesian accessions PI 567102B and PI 567104b and the significance of the ss715632525 marker near the Rpp6 locus on Chr 18. Since PI 567,090 also has the same base residue at that marker, it might also have a resistance gene at the Rpp6 locus. The Rpp6 gene is one of the few named genes that have been effective in both North and South America (Miles et al. 2008; Walker et al. 2014b). PIs 476905A, 567068A, 567076 and 567129 may also likely have a resistance gene at the Rpp6 locus (Harris et al. 2015), but their BLUP values ranged from − 0.35 to − 0.56, suggesting that they carry a different allele from the one in PI 567102B and PI 567104B.
When Chang et al. (2016) performed a GWAS for SBR resistance using infection type data from the Miles et al. (2006) greenhouse assays, they found one significant marker near the Rpp1 locus on Chr 18 and another on Chr 15. The Rpp1 and Rpp6 loci are both on Chr 18 but are located on different telomeres (Kim et al. 2012; Li et al. 2012; Yu et al. 2015). Although PI 200492 was one of the most resistant accessions in our study, we did not detect any significant markers close to the Rpp1 locus. At least 11 of the PIs in the panel have a resistance allele at the Rpp1 locus, but the allele(s) in seven Chinese accessions were ineffective against the US field pathotypes. As a result, the frequency of Rpp1 alleles with significant phenotypic effects in this study was low, which probably explains the reason that the locus was not detected using GWAS. Other than PI 200492, PIs 417120 and 423958 from Japan, and PI 518295 from Taiwan were among the few accessions screened that had an effective resistance allele at the Rpp1 locus (Supplemental Table S1).
The failure of the GWAS analysis to detect significant markers close to Rpp2 on Chr 16; (Yu et al. 2015), Rpp4 at the opposite end of Chr 18 from Rpp1 and Rpp6 (Silva et al. 2008), Rpp5 on Chr 3 (Garcia et al. 2008), or Rpp7 on Chr 19 (Childs et al. 2018b) was likely due to a low frequency of resistance genes and/or weak phenotypic effects. Although PI 605823 (Rpp7) and PI 200487 (allele at Rpp5) were among the 20 accessions with the lowest BLUP means for disease in this study, Rpp7 has not been reported in any other accessions, and PI 471904 appears to be the only other PI with an effective allele at the Rpp5 locus. A failure to detect genes that are present at a low frequency in a population is one limitation of GWAS analyses (Bandillo et al. 2015).
The detection of significant markers in regions of six chromosomes that have not been reported to have Rpp loci was unexpected, and the fact that the marker significance levels were higher than that of the marker close to the Rpp6 locus is encouraging. Some of the resistant accessions that we tested may have novel resistance genes in those regions that could be used in Rpp gene pyramids to improve resistance. Because a majority of the PIs in the GWAS panel had been reported by Miles et al. (2006) to be susceptible or to have had mixed reactions to SBR, few of them have been used to develop biparental mapping populations. It is possible that the SBR disease rating data that we used for the GWAS may have allowed contributions of Rpp loci in the six regions to be detected for the first time. Much of the phenotypic data used the BLUP values used for GWAS in this study reflected both disease severity and intensity of urediniospore production. Those semi-quantitative data provided a more accurate assessment of reactions that were intermediate between heavily sporulating TAN infection types and Type 0 or RB infection types with few uredinia and low sporulation. Yamanaka et al. (2010) also recognized the value of using semi-quantitative criteria (i.e., rating scales) for more accurate assessments of host reactions to SBR. Although Chang et al. (2016) also reported a novel putative resistance QTL between positions 10,659,000 and 10,859,000 bp on Chr 15, it did not correspond to the genomic region that we detected near position 15,694,109 on that chromosome.
Findings from this study should be useful for the development of cultivars with broader and more durable SBR resistance. Some accessions previously reported be susceptible to SBR should be re-evaluated for resistance in some other countries where rust is an economically important disease. Confirmation of an unknown Rpp locus in any of the six previously unreported genomic regions that were detected by GWAS would offer soybean breeders additional options for developing cultivars with novel combinations of resistance genes. Data collected early in this study led to the discovery of the Rpp7 locus in PI 605823 (Childs et al. 2018b). After the resistant accessions responsible for the significant GWAS markers are identified, their resistance can be characterized using a panel of pathogenically diverse P. pachyrhizi isolates, and biparental mapping populations can be created to fine-map the locations of the novel loci.
Data availability
Supplementary Table S1 contains the BLUP values for all accessions used for the GWAS analysis. The disease rating data for PIs grown in each year-location environment are available from the corresponding authors on reasonable request.
References
Akamatsu H, Yamanaka N, Yamaoka Y, Soares RM, Morel W, Ivanovich AJG, Bogado AN, Kato M, Yorinori JT, Suenaga K (2013) Pathogenic diversity of soybean rust in Argentina, Brazil, and Paraguay. J Gen Plant Pathol 79:28–40
Allen TW, Hollier CA, Sikora EJ (2014) A continuing saga: soybean rust in the continental United States, 2004 to 2013. Outlooks Pest Manag 25:167–174
Allen TW, Bissonnette K, Bradley CA, Damicone JP, Dufault NSS, Faske TR, Isakeit T, Kemerait RC, Koehler A, Langston D, Mueller JD, Padgett GB, Price PP, Sikora EJ, Small IM, Thiessen L, Young H (2021) Southern United States soybean disease loss estimates for 2020. In: Proceedings of the Southern Soybean Disease Workers 48th Annual Meeting, Virtual Content, Pensacola Beach, FL, 3 March 2021, pp 1–0
Bandillo N, Jarquin D, Song Q, Nelson R, Cregan P, Specht J, Lorenz A (2015) A population structure and genome-wide association analysis in the USDA Soybean Germplasm Collection. Plant Genome 8:1–13. https://doi.org/10.3835/plantgenome2015.04.0024
Bonde MR, Nester SE, Austin CN, Stone CL, Frederick RD, Hartman GL, Miles MR (2006) Evaluation of virulence of Phakopsora pachyrhizi and P. meibomiae isolates. Plant Disease 90(6):708–716. https://doi.org/10.1094/PD-90-0708
Bradbury PJ, Zhang Z, Kroon DE, Casstevens TM, Ramdoss Y, Buckler ES (2007) TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics 23:2633–2635. https://doi.org/10.1093/bioinformatics/btm308
Bromfield KR, Hartwig EE (1980) Resistance to soybean rust and mode of inheritance. Crop Sci 20:254–255
Bromfield KR, Melching JS, Kingsolver CH (1980) Virulence and aggressiveness of Phakopsora pachyrhizi isolates causing soybean rust. Phytopathology 70:17–21
Bromfield KR (1984) Soybean rust. Monogr. No. 11. Am. Phytopathol. Soc., St. Paul, MN
Calvo ÉS, Kiihl RAS, Garcia A, Harada A, Hiromoto DM (2008) Two major recessive soybean genes conferring soybean rust resistance. Crop Sci 48:1350–1354. https://doi.org/10.2135/cropsci2007.10.0589
Chakraborty N, Curley J, Frederick RD, Hyten DL, Nelson RL, Hartman GL, Diers BW (2009) Mapping and confirmation of a new allele at Rpp1 from soybean PI 594538A conferring RB lesion-type resistance to soybean rust. Crop Sci 49:783–790. https://doi.org/10.2135/cropsci2008.06.0335
Chang H-X, Lipka AE, Domier LL, Hartman GL (2016) Characterization of disease resistance loci in the USDA Soybean Germplasm Collection using genome-wide association studies. Phytopathology 106:1139–1151. https://doi.org/10.1094/PHYTO-01-16-0042-FI
Childs SP, Buck JW, Li Z (2018a) Breeding soybeans for resistance to soybean rust (Phakopsora pachyrhizi). Plant Breed 137:250–261. https://doi.org/10.1111/pbr.12595
Childs SP, King ZR, Walker DR, Harris DK, Pedley KF, Buck JW, Boerma HR, Li Z (2018b) Discovery of a seventh Rpp soybean rust resistance locus in soybean accession PI 605823. Theor Appl Genet 131:27–41. https://doi.org/10.1007/s00122-017-2983-4
Dalla Lana F, Paul PA, Godoy CV, Utiamada CM, da Silva LHCP, Siqueri FV, Forcelini CA, de Souza J-F, Miguel-Wruck DS, Borges EP, Juliatti FC, Campos HD, Nunes J Jr, Carneiro LC, Canteri MG, Ito MF, Meyer MC, Martins MC, Balardin RS, Furlan SH, Carlin VJ, Del Ponte EM (2018) Meta-analytic modeling of the decline in performance of fungicides for managing soybean rust after a decade of use in Brazil. Plant Dis 102:807–817. https://doi.org/10.1094/PDIS-03-17-0408-RE
Hwang E-Y, Song Q, Jia G, Specht JE, Hyten DL, Costa J, Cregan PB (2014) A genome-wide association study of seed protein and oil content in soybean. BMC Genomics 15(1):1. https://doi.org/10.1186/1471-2164-15-1
Garcia A, Calvo ÉS, Kiihl RS, Harada A, Hiromoto DM, Vieira LG (2008) Molecular mapping of soybean rust (Phakopsora pachyrhizi) resistance genes: Discovery of a novel locus and alleles. Theor Appl Genet 117:545–553. https://doi.org/10.1007/s00122-008-0798-z
Garcia A, Calvo ÉS, Kiihl RAS, Souto ER (2011) Evidence of a susceptible allele inverting the dominance of rust resistance in soybean. Crop Sci 51:32–40. https://doi.org/10.2135/cropsci2010.01.0037
Grant D, Nelson RT, Cannon SB, Shoemaker RC (2010) SoyBase, the USDA-ARS soybean genetics and genomics database. Nucleic Acids Res. https://doi.org/10.1093/nar/gkp798
Guo B, Sleper DA, Lu P, Shannon JG, Nguyen HT, Arelli PR (2006) QTLs associated with resistance to soybean cyst nematode in soybean: meta‐analysis of QTL locations. Crop Sci 46(2):595–602. https://doi.org/10.2135/cropsci2005.04-0036-2
Harris DK, Kendrick MD, King ZR, Pedley KF, Walker DR, Cregan PB, Buck JW, Phillips DV, Li Z (2015) Identification of unique genetic sources of soybean rust resistance from the USDA Soybean Germplasm Collection. Crop Sci 55:1–16. https://doi.org/10.2135/cropsci2014.09.0671
Hartman GL, Wang TC, Hymowitz T (1992) Sources of resistance to soybean rust in perennial Glycine species. Plant Dis 76:396–399
Hossain MM, Akamatsu H, Morishita M, Mori T, Yamaoka Y (2015) Molecular mapping of Asian soybean rust resistance in soybean landraces PI 594767A, PI 587905 and PI 416764. Plant Pathol 64:147–156. https://doi.org/10.1111/ppa.12226
Hyten DL, Hartman GL, Nelson RL, Frederick RD, Concibido VC, Narvel JM, Cregan PB (2007) Map location of the Rpp1 Locus that confers resistance to soybean rust in soybean. Crop Sci 47(2):837–838. https://doi.org/10.2135/cropsci2006.07.0484
Hyten DL, Hartman GL, Nelson RL, Frederick RD, Concibido VC, Narvel JM, Cregan PB (2009) Bulked segregant analysis using the goldengate assay to locate the Rpp3 locus that confers resistance to soybean rust in soybean. Crop Sci 49(1):265–271. https://doi.org/10.2135/cropsci2008.08.0511
Kendrick MD, Harris DK, Ha B-K, Hyten DL, Cregan PB, Frederick RD, Boerma HR, Pedley KD (2011) Identification of a second Asian soybean rust resistance gene in Hyuuga soybean. Phytopathology 101:535–543. https://doi.org/10.1094/PHYTO-09-10-0257
Kim K-S, Unfried JR, Hyten DL, Frederick RD, Hartman GL, Nelson RL, Song Q, Diers BW (2012) Molecular mapping of soybean rust resistance in soybean accession PI 561356 and SNP haplotype analysis of the Rpp1 region in diverse germplasm. Theor Appl Genet 125:1339–1352. https://doi.org/10.1007/s00122-012-1932-5
Klosowski A, May de Mio LL, Miessner S, Rodrigues R, Stammler G (2016) Detection of the F129L mutation in the cytochrome b gene in Phakopsora pachyrhizi. Pest Manag Sci 72:1211–1215. https://doi.org/10.1002/ps.4099
Kumudini S, Godoy CV, Board JE, Omielan J, Tollenaar M (2008) Mechanisms involved in soybean rust-induced yield reduction. Crop Sci 48:2334–2342. https://doi.org/10.2135/cropsci2008.01.0009
Lemos NG, de Lucca e Braccini A, Abdelnoor RV, de Oliveira MC, Suenaga K, Yamanaka N (2011)Characterization of genes Rpp2, Rpp5, and Rpp5 for resistance to soybean rust. Euphytica 182:53–64. https://doi.org/10.1007/s10681-011-0465-3
Li S, Smith JR, Ray JD, Frederick RD (2012) Identification of a new soybean rust resistance gene in PI 567102B. Theor Appl Genet 125:133–142. https://doi.org/10.1007/s00122-012-1821-y
Liu M, Li S, Swaminathan S, Sahu BB, Leandro LF, Cardinal AJ, Bhattacharyya MK, Song Q, Walker DR, Cianzio SR (2016a) Identification of a soybean rust resistance gene in PI 567104B. Theor Appl Genet 129:863–877. https://doi.org/10.1007/s00122-015-2651-5
Liu X, Huang M, Fan B, Buckler ES, Zhang Z (2016b) Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet 12:e1005767. https://doi.org/10.1371/journal.pgen.1005767
Miles MR, Frederick RD, Hartman GL (2006) Evaluation of soybean germplasm for resistance to Phakopsora pachyrhizi. Plant Health Prog. https://doi.org/10.1094/PHP-2006-0104-01-RS
Miles MR, Morel W, Ray JD, Smith JR, Frederick RD, Hartman GL (2008) Adult plant evaluation of soybean accessions for resistance to Phakopsora pachyrhizi in the field and greenhouse in Paraguay. Plant Dis 92:96–105. https://doi.org/10.1094/PDIS-92-1-0096
Ogle HJ, Byth DE, McLean R (1979) Effect of rust (Phakopsora pachyrhizi) on soybean yield and quality in south- eastern Queensland. Aust J Agric Res 30:883–893
Pham TA, Miles MR, Frederick RD, Hill CB, Hartman GL (2009) Differential responses of resistant soybean entries to isolates of Phakopsora pachyrhizi. Plant Dis 93:224–228
Pham TA, Hill CB, Miles MR, Nguyen BT, Vu TT, Vuong TD, VanToai TT, Hguyen HT, Hartman GL (2010) Evaluation of soybean for resistance to soybean rust in Vietnam. Field Crops Res 117:131–138. https://doi.org/10.1016/j.fcr.2010.02.011
Pierozzi PHB, Ribeiro AS, Moreira JUV, Laperuta LDC, Rachid LWF, Arias CAA, de Oliveira MF, de Toledo JFF (2008) New soybean (Glycine max Fabales, Fabaceae) sources of qualitative genetic resistance to Asian soybean rust caused by Phakopsora pachyrhizi (Uredinales, Phakopsoraceae). Genetics Mol Biol 31:505–511
Qin J, Song Q, Shi A, Li S, Zhang M, Zhang B (2017) Genome-wide association mapping of resistance to Phytophthora sojae in a soybean [Glycine max (L.) Merr.] germplasm panel from maturity groups IV and V. PLoS ONE 12(9):e0184613. https://doi.org/10.1371/journal.pone.0184613
Ray JD, Smith JR, Morel W, Bogado N, Walker DR (2011) Genetic resistance to soybean rust in PI567099A is at or near the Rpp3 locus. J Crop Improv 25:219–231. https://doi.org/10.1080/15427528.2011.555833
Schneider RW, Hollier CA, Whitam HK, Palm ME, McKenny JM, Hernandez JR, Levy L, DeVries-Paterson R (2005) First report of soybean rust caused by Phakopsora pachyrhizi in the continental United States. Plant Dis 89:774
Shaw RG (1987) Maximum-likelihood approaches applied to quantitative genetics of natural populations. Evolution 41:812–826
Sikora EJ, Conner K (2021) Observations from soybean rust monitoring and fungicide field demonstrations in Alabama in 2020. In: Proceedings of the southern soybean disease workers 48th annual meeting, virtual content, Pensacola Beach, FL, 3 March 2021
Sikora EJ, Conner K, Delaney D, Zhang L, Delaney M (2014) Observations on soybean rust and soybean vein necrosis virus in Alabama in 2013. In: Proceedings of the southern soybean disease workers 41st annual meeting, Pensacola, FL, USA, 5–6 March 2014
Silva DCG, Yamanaka N, Brogin RL, Arias CAA, Nepomuceno AL, Di Mauro AO, Pereira SS, Nogueira LM, Passianotto ALL, Abdelnoor RV (2008) Molecular mapping of two loci that confer resistance to Asian rust in soybean. Theor Appl Genetics 117(1):57–63. https://doi.org/10.1007/s00122-008-0752-0
Song Q, Hyten DL, Jia G, Quigley CV, Fickus EW, Nelson RL, Cregan PB (2015) Fingerprinting Soybean Germplasm and Its Utility in Genomic Research. G3 Genes Genomes Genet 5:1999–2006. https://doi.org/10.1534/g3.115.019000
Steketee CJ, Schapaugh WT, Carter TE, Li Z (2020) Genome-wide association analyses reveal genomic regions controlling canopy wilting in soybean. G3 Genes Genomes Genet 10:1413–1425. https://doi.org/10.1534/g3.119.401016
Stone CL, Smith JR, Ray JD, Gillen AM, Frederick RD (2022) Phenotypic reactions of 53 soybean genotypes to infection with each of 16 isolates of Phakopsora pachyrhizi. J Crop Improvement 36:25–39. https://doi.org/10.1080/15427528.2021.1904311
Twizeyimana M, Hartman GL (2012) Pathogenic variation of Phakopsora pachyrhizi isolates on soybean in the United States from 2006 to 2009. Plant Dis 96:75–81. https://doi.org/10.1094/PDIS-05-11-0379
Twizeyimana M, Ojiambo PS, Ikotun T, Ladipo JL, Hartman GL, Bandyopadhyay R (2008) Evaluation of soybean germplasm for resistance to soybean rust (Phakopsora pachyrhizi) in Nigeria Plant Disease 92(6):947–952. https://doi.org/10.1094/PDIS-92-6-0947
Twizeyimana M, Ojiambo PS, Ikotun T, Ladipo JL, Hartman GL, Bandyopadhyay R (2009) Pathogenic variation of Phakopsora pachyrhizi infecting soybean in Nigeria. Phytopathology® 99(4):353–361. https://doi.org/10.1094/PHYTO-99-4-0353
Vaughn JN, Nelson RL, Song Q, Cregan PB, Li Z (2014) The genetic architecture of seed composition in soybean is refined by genome-wide association scans across multiple populations. G3 Genes|Genomes|Genetics 4(11):2283–2294. https://doi.org/10.1534/g3.114.013433
Vuong TD, Walker DR, Nguyen BT, Nguyen TT, Dinh HX, Hyten DL, Cregan PB, Sleper DA, Lee JG, Shannon JG, Nguyen HT (2016) Molecular characterization of resistance to soybean rust (Phakopsora pachyrhizi Syd. & Syd.) in soybean cultivar DT 2000 (PI 635999). PLoS ONE 11(12):e0164493. https://doi.org/10.1371/journal.pone.0164493
Walker DR, Boerma HR, Phillips DV, Schneider RW, Buckley JB, Shipe ER, Mueller JD, Weaver DB, Sikora EJ, Moore SH, Hartman GL (2011) Evaluation of USDA soybean germplasm accessions for resistance to soybean rust in the southern United States. Crop Sci 51:678–693. https://doi.org/10.2135/cropsci2010.06.0340
Walker DR, Harris DK, King ZR, Li Z, Phillips DV, Buck JW, Nelson RL, Boerma HR (2014a) Soybean germplasm accession seedling reactions to soybean rust isolates from Georgia. Crop Sci 54:1433–1447. https://doi.org/10.2135/cropsci2013.09.0654
Walker DR, Harris DK, King ZR, Li Z, Boerma HR, Buckley JB, Weaver DB, Sikora EJ, Shipe ER, Mueller JD, Buck JW, Schneider RW, Marois JJ, Wright DL, Nelson RL (2014b) Evaluation of soybean germplasm accessions for resistance to Phakopsora pachyrhizi populations in the southeastern United States, 2009–2012. Crop Sci 54:1673–1689. https://doi.org/10.2135/cropsci2013.08.0513
Wang J, Zhang Z (2020) GAPIT Version 3: boosting power and accuracy for genomic association and prediction 1 2. bioRxiv 2020.11.29.403170
Wickham H (2016) ggplot2: elegant graphics for data analysis, 2nd edn. Springer, Switzerland
Yamanaka N, Hossain MM (2019) Pyramiding three rust-resistance genes confers a high level of resistance in soybean (Glycine max). Plant Breed 138:686–695. https://doi.org/10.1111/pbr.12720
Yamanaka N, Yamaoka Y, Kato M, Lemos NG, Passianotto AL, dos Santos JV, Benitez ER, Abdelnoor RV, Soares RM, Suenaga K (2010) Development of classification criteria for resistance to soybean rust and differences in virulence among Japanese and Brazilian rust populations. Trop Plant Pathol 35:153–162
Yamanaka N, Lemos NG, Uno M, Akamatsu H, Yamaoka Y, Abdelnoor RV, Braccini AL, Suenaga K (2013) Resistance to Asian soybean rust in soybean lines with the pyramided three Rpp genes Crop Breeding. Appl Biotechnol 13:75–82. https://doi.org/10.1590/S1984-70332013000100009
Yamaoka Y, Fujiwara Y, Kakishima M, Katsuya K, Yamada K, Hagiwara H (2002) Pathogenic races of Phakopsora pachyrhizi on soybean and wild plant host plants collected in Japan. J Gen Plant Pathol 68:52–56
Yu N, Kim M, King ZR, Harris DK, Buck JW, Li Z, Diers BW (2015) Fine mapping of the Asian soybean rust resistance gene Rpp2 from soybean PI 230970. Theor Appl Genet 128:387–396. https://doi.org/10.1007/s00122-014-2438-0
Acknowledgements
The authors particularly thank Dr. Randall Nelson (USDA-ARS, retired) for suggesting that some germplasm accessions should be re-tested for SBR resistance in the field, for recommending certain accessions, and for providing seed from the USDA Soybean Germplasm Collection in Urbana, Illinois. We also thank Drs. Raymond W. Schneider (now deceased) and Clark Robertson of the Louisiana State University AgCenter and Plant Pathology and Crop Physiology Department in Baton Rouge, LA, for planting and rating the 2008 test in Louisiana; Debbie Dalton, G. Kelly O’Brien and Maynard Douglas at the University of Florida’s North Florida Research and Education Center in Quincy, FL; Billy Mills at the University of Georgia’s Attapulgus Research and Education Center in Attapulgus, GA; and Malcomb Pegues at Auburn University’s Gulf Coast Research and Extension Center in Fairhope, AL.
Funding
This research was funded by the USDA-ARS, the Georgia Agricultural Experiment Stations, and United Soybean Board projects. The 2015 field test was also supported with funding from the USDA National Plant Disease Recovery System (NPDRS).
Author information
Authors and Affiliations
Contributions
DRW and ZL organized the study, compiled the data and wrote an early draft of the manuscript; DRW, DKH HRB, JWB, EJS, DBW, DLW and JJM grew the field tests and collected disease data; SCM and ZL calculated BLUP values, conducted the GWAS and generated the figures; all of the authors proofread and contributed to late drafts of the manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Consent for publication
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture. The US Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, or disability, and where applicable, sex, marital status, familial status, parental status, religion, sexual orientation, genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program. The USDA is an equal opportunity provider and employer.
Additional information
Communicated by Istvan Rajcan.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Walker, D.R., McDonald, S.C., Harris, D.K. et al. Genomic regions associated with resistance to soybean rust (Phakopsora pachyrhizi) under field conditions in soybean germplasm accessions from Japan, Indonesia and Vietnam. Theor Appl Genet 135, 3073–3086 (2022). https://doi.org/10.1007/s00122-022-04168-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-022-04168-y