
Abstract
Seeds, i.e. embryos, may be genetically different from either of their parents and moreover may express their own heterosis. The objective was to genetically analyse embryo heterosis for their own weight (i.e. seed weight) in comparison with their seedlings’ heterosis, taking the large-seeded crop (Vicia faba L.) as model. A specific diallel mating scheme was used, based on four parental lines, creating 76 seed genotypes in generations P, F1, F2 and BC. Mature seed weight was assessed for these embryo genotypes in 3 years at one German location, and young plant biomass yield of seedlings emerging from these seeds in two greenhouse experiments. The quantitative genetic analysis showed an average of 10.6% mid-parent heterosis for mature seed weight and 14.5% mid-parent heterosis for juvenile biomass. In both traits, the embryos contributed markedly and significantly via their own genes to the genetic variation. For mature embryo weight heterosis, apparently the parental difference in seed weight was decisive, whereas for juvenile biomass heterosis, genetic unrelatedness of parents had priority.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The seeds of annual crop plants are of crucial importance, as humankind depends on them both as prerequisite to start the next farming season and as staple foods. On the one hand, seed traits such as size and vigour play a major role in the establishment of promising crop stands; on the other hand, the harvest is composed of seeds, and seed traits such as quality affect their market value.
Two genetic phenomena are tightly connected: (1) seeds, in fact their embryos, may have genes and genotypes that are different from those of their seed parent plant, and may have their own genetic impact on their traits; and (2) embryos may have their own degree of heterozygosity and heterosis. The degree of heterozygosity of embryos is independent of both parents, except in the case of self-fertilization and of autopolyploidy.
A well-known case is the erucic acid content of Brassica napus seed which is determined by the embryo genotype (Stefansson and Hougen 1964). A more striking cause of genetic differences among seeds and between seeds and seed parents than segregation are genetically different pollen sources. The term ‘xenia’ (Focke 1881) was first coined to describe the effects of different pollen sources on seed and fruit. Tschermak (1931) defined ‘genuine xenia’ if the pollen source influences colour, shape, and chemical composition of the seed, and ‘metaxenia’ if pollen influences the fruit. Tschermak (1932) discussed so-called ‘false xenia’, i.e. greater seed size of crossed seeds than selfed seeds, as reported by Nicolaisen (1931) in rye and pea. Kiesselbach (1960) reported kernel yield increase in maize caused by cross-pollen instead of self-pollen; this principle is used today when growing TopCross® hybrids in maize (Thomison and Geyer 1999). False xenia presumably reflects very early heterosis (embryos’ own heterosis, not heterosis of its mother plant).
Heterosis and its complement, inbreeding depression, denote the general experience that the F1 hybrid of two genetically different, inbred parents surpasses them in performance. Heterosis is defined as the difference between trait expression of this F1 hybrid and the average of its homozygous parents. Classical genetic explanations for heterosis are the dominance and overdominance hypothesis, modified by inclusion of epistasis (Crow 1999; Becker and Link 2000; Syed and Chen 2005; Frascaroli et al. 2007; Lee et al. 2007).
Heterosis is usually analysed for traits that are expressed by the green plant. Nonetheless, the earliest stage of the plant, the embryo, may already express heterosis. The assessment of embryo heterosis is hampered by (1) the fact that homozygous seed of the male parent cannot develop side by side with the hybrid seed and with homozygous seed of the female parent on the same seed parent plant (same “environment”), and (2) the existence of different parental gene doses in embryo and endosperm. The first item is usually addressed by producing hybrid seed on both parents and taking their average as F1 value, to be compared to the average of the homozygous seed from the two parents. The second item is of relevance because seed traits are a complex result of endosperm and embryo features and because the triploid endosperm carries two doses of the maternal genes. This point is experimentally addressed either by excising the embryos rather early, and growing them on standardized in vitro conditions (e.g. Meyer et al. 2007, in case of maize), by biometrically separating endosperm effects from embryo effects (e.g. Lou et al. 2005), or by the use of seed that is nearly devoid of endosperm (Dure 1975) such as cotton (Dani and Kohel 1989) or faba bean (Duc et al. 2001).
Maize embryos excised 6 days after pollination showed heterosis, the hybrid embryos of the two reciprocal cross directions being on average 31% longer than the corresponding homozygous parental embryos (Meyer et al. 2007). Few genes showed a markedly higher expression in the hybrid embryos, mainly related to signal transduction and regulatory processes. Duc et al. (2001) reported from Vicia faba heterotic increments in mature seed size (2–8%) due to cross-fertilization compared to selfing, based on one seed parent and two pollen parents.
Embryo heterosis may be the primer of the heterosis of seedling plants. In thale cress (Arabidopsis thaliana), heterosis could be detected already 10 days after sowing and for biomass it was from 42 to 118%, depending on the light intensity (60–240 μm m2 s−1; Meyer et al. 2004). Hoecker et al. (2006) assessed the lateral root density in maize seedlings 5 days after emergence. They used four parental inbred lines and established all six hybrids, in both reciprocal directions. The average heterosis was 51%.
A decade ago, novel quantitative genetic models were developed to analyse differences between seed genotypes and generations (Zhu and Weir 1994). Based on these tools, the objectives here were to genetically analyse mature embryo heterosis in comparison with young plants’ heterosis, taking the grain legume V. faba as model. The faba beans offer large seeds and a large variation of this trait, and it is a partially allogamous crop (Link 1990), thus intermediate between honest selfers and outcrossers. The hypothesis here was that embryo heterosis is reflected in mature seed weight and that this heterosis responds to parental relatedness. When denoting a seed kernel, here the focus is on its embryo, neglecting the testa.
Materials and methods
The first experiment focussed on heterosis of mature seeds. It was conducted across 3 years at one location, the experimental station Reinshof, Göttingen, Germany (51.5°N, 9.9°E). Reinshof is located near the river Leine at 150 m above sea level and has high soil fertility. Average annual rainfall is 645 mm. The three seasons for this experiment, 2004–2006, were favourable for faba beans. The experiments were carried out in so-called isolation cages (Link 1990), 3 m high, 3 m wide and 20 m long, covered with a white mesh (2 mm × 2 mm) that prevents access of pollinator insects (honeybees, bumblebees). Thus, uncontrolled cross-pollination is prevented. Plants were grown inside such cages with 0.4 m between rows, 0.1 m spacing within rows and 1 m of row length. There was one replicate in 2004 and two replicates in 2005 and in 2006. Replicates were sown in individual isolation cages. Sowing was early in April and harvesting was in September. Herbicides and insecticides were applied as necessary.
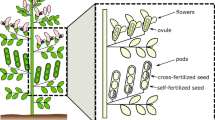
The experiment was based on four inbred lines (Table 1), three of which were related to each other. The lines ‘HP58’ and ‘HP69’ were developed by single seed descent from the relatively wide cross between the small-seeded, German tick bean line ‘Hedin’ (Table 1) and the very large-seeded, Sicilian broad bean line ‘Pietranera’ (major, TSW about 1,800 g). HP58 and HP69 were chosen due to their large differences in seed size (Table 1) in spite of having a common genetic background. The line ‘Gobo’ is not related to the three other entries (Dieckmann 2007). These spring faba beans rarely produce tillers. Their earliest inflorescence appears typically at the 6th leaf, their ultimate number of fertile inflorescences is more than 8 with more than four flowers per inflorescence, a plant can yield more than 2 pods at an inflorescence and altogether more than 8 pods with two to four seeds per pod.
These 4 inbred lines and their 12 diallel F1 hybrids (six combinations in two reciprocal crossing directions) were manually self-fertilized (tripped) and crossed following the diallel mating scheme of Dani and Kohel (1989; cf. Table 2). Tripping stands for manually opening the fully developed flower by holding the wing petals and pulling away the flag petal; this is repeated twice or thrice as a mechanical stimulation of selfing (Link 1990). Crossing stands for manual emasculating and cross-pollinating. The resulting seeds and their embryos were either highly inbred (selfing of inbred line), non-inbred (crossing among lines) or half inbred (selfing of an F1 hybrid or crossing between a parental line and its F1 hybrid; the above-mentioned basic relationship between three of the four lines was disregarded for this statement).
In the isolation cages, one row of seed parent plants was used to produce one embryo genotype through manual crossing (F1, BC, or RBC; cf. Table 2); selfing was carried out simultaneously (see below). From the ten plants per row, four to six were used; first and last plants were considered as border and not used as seed parents. Eventually occurring tillers of seed parents were removed, and main stems were trimmed to eight consecutive inflorescences with one pod per inflorescence. This was realized by cutting surplus young pods at latest 14 days after flowering. Only young pods in the second or third position in the inflorescence were left to grow on and become mature. For seed parent plants, the aim was to have either one selfed or one crossed pod per inflorescence, growing on alternate inflorescences, starting at the earliest, lowermost inflorescence, and with plants in a row alternating for selfed pod first and crossed pod first. Mature seeds were harvested and dried until equilibrium. Subsequently, they were individually weighed, recording their individual seed parent and their embryo genotype.
Nine rows were grown per parental inbred line to be used as seed parent, three rows for each of the three combinations per parent (i.e. 1 × 2; 1 × 3; and 1 × 4), and within a combination one row for each of the three cross-fertilized seed types produced on a parent (underlined in Table 2). Two rows were grown for each of the 12 hybrids (e.g. F1(1 × 2)) to serve as seed parent when producing its two backcross seed genotypes (italics in Table 2). Thus, (4 × 9) + (2 × 12) = 60 rows established one replicate. Further rows of these genotypes were grown to serve as the pollen source for manual crossing.
The individual mature seed weights of the crossed seeds (F1, BC, and RBC) were first averaged for the seeds of the individual seed parent plants and thereafter across the plants of a given row. The same applied for the selfed seeds (P and F2). The resultant data were analysed with QGA Station Version 1.0 software developed by Chen and Zhu (2003) for quantitative genetic studies (Zhu and Weir 1994). The following model was applied:
where μ is the population mean, E y the effect of the environment (i.e. year), G k the genotypic effect, GEyk the genotype × environment interaction effect, B yc the effect of replicate (i.e. isolation cage) and e ykcr = residual error effect.
The genotypic value of an embryo genotype was further decomposed, applying a generation mean analysis for the diallel crosses. In contrast to single cross generation mean analyses (Dani and Kohel 1989), here the four parents are allowed to unfold a conceptual reference population. Therefore, rather than metric effects (‘a’ and ‘d’; Falconer and Mackey 1996, p. 109), statistical genetic effects (‘average effect’ of an allele; ‘dominance deviation’ of a locus; Falconer and Mackey 1996, pp. 112–119) had to be employed. Consequently, the genetic effect A i (Table 3) termed ‘direct additive effect’ reflects the average impact of the alleles of parent i on mature seed weight; this impact being realized through the embryo that inherited these alleles. The effect D ii is called ‘direct dominance effects’ of the alleles of the ancestor i in homozygous status (D ii might be understood as ‘direct inbreeding effect’), and D ij is the ‘direct dominance effect’ of the alleles i and j in heterozygous status. The parameters C i and C j represent potential cytoplasmic differences between the parents. The further parameter Am i allows for additive maternal effects of the seed parent on its seed; and Dm ii allows for a dominant maternal effect of the homozygous parent i, and Dm ij for a dominant maternal effect when the seed parent is the hybrid i × j (Zhu 1997). As implemented in QGA Station, MINQUE (0/1) was used to estimate variance components; an adjusted unbiased prediction method was applied to predict the components of genetic effects (http://ibi.zju.edu.cn/software/qga; Zhu and Weir 1994; Zhu 1997). For each of the four paternal inbred lines, each of the A i , D ii , C i , Am i , Dm ii effects was estimated, thus amounting to 5 × 4 = 20 effects. Moreover, for the six combinations i × j, the D ij and the Dm ij effects were estimated, resulting in a total of 32 genetic effects, to be estimated from a total of 76 genotypes. The corresponding matrix of coefficients is shown in Table 3.
In addition, generation means of seed weight and plant weight were calculated and F1 versus P, F1 versus BC as well as RBC and F2 versus BC (cf. Table 2) were tested for significance of difference by Student’s t test, using the genotype means across the 3 years. Admittedly, heterosis of each cross could be directly taken as difference between the mean of the two reciprocal F1 hybrids and the mean of their two parents (F1 mean − P mean; cf. Table 3). The generation BC is expected to realize half-heterosis. This heterosis is realized on fully inbred seed parent plants. Two further half-heterotic seed generations, F2 and RBC, grew on non-inbred seed parents plants. QGA Station uses the complete data set to jointly estimate among others the effects D ii , D jj , and D ij . Thus, heterosis is deduced based on Eq. 1:
The selfed pods growing on the four seed parents were scored according to the number of seeds they contained, in order that the impact of this number on mature seed weight could be quantified.
The second experiment focussed on heterosis in seedling weight. All 76 seed genotypes (cf. Table 2) were assessed as young plants for their above-ground biomass in winter 2006/2007 in the greenhouse. A first set was sown on 1 November 2006, the second on 12 December 2006; both were harvested 27 days after sowing. The design was a randomized complete block with eight replicates per set, a replicate being one table of 2 m × 1.3 m with pots. Each table comprised all genotypes, with five 7 cm × 7 cm pots per genotype, one seedling each. A single buffer row of pots of cv Merkur was arranged at all edges of the table. Soil, watering and supplemental lighting (15 h day−1) were chosen to allow optimum plant growth. The experimental units were the five pots per genotype per table. Their average above-ground fresh weight was used for the statistical analyses. Water content determined after drying for 24 h at 60°C and further 12 h at 105°C was 93% with a very low variation among genotypes (CV% = 0.0043).
For the quantitative genetic analyses of the juvenile biomass experiment, the two sets were coded as environments, the eight tables as replicates. The decomposition of the plant genotype values was as for embryo genotypes with the mature seed weight data using QGA Station.
Spearman’s rank correlation coefficient was calculated between generations and between the two traits.
Results
Across the three seasons and across the plant genotypes and generations, an average of 7.3 pods and 20.6 seeds per seed parent plant were harvested and analysed. The average number of analysed crossed pods (F1, BC, RBC) per plant was 3.4, the average number of selfed pods (P, F2) per plant was 3.9. The average number of seeds per pod was 2.9 in crossed pods and 3.2 in selfed pods; the difference was significant (P > 0.05).
On average across the four parents, selfed seeds in two-seeded pods were 0.28% lighter and in four-seeded pods they were 1.49% lighter than in three-seeded pods. The largest difference from a single parent’s average was, for two-seeded pods, +1.1% in Gobo, for three-seeded pods +1.3% (Hedin), and for four-seeded pods −1.9% (HP69). Linear regression of mature seed weight on seed number per pod explained 0.047% of the variation; it was not significant.
The differences among embryo genotypes for seed weight and young plant weight were highly significant (P < 0.001). The mature mean seed weight of hybrid seeds was 921.4 mg (Fig. 1), being 91.8 mg or 11.1% higher than the mature mean seed weight of selfed seed on parents; this difference was highly significant (P < 0.02). The smallest hybrid seeds [516 mg; F1(Hedin × HP69)] grew on the smallest-seeded parent Hedin (cf. Table 1). The largest hybrid seeds (1,680 mg) grew on HP58 and were the seed genotype F1(HP58 × HP69). When taking this F1 hybrid as pollinator for HP58, almost the same average seed weight of 1,670 mg was attained, being the largest value of the BC generation. Nevertheless, the average of this BC generation was significantly (P < 0.05) lower than F1; it was 877.8 mg, thus almost exactly halfway between P and F1 (Fig. 1). These backcrossed seeds grew on the parental lines, hence reflecting their large differences in seed weight, whereas the backcrossed RBC and the F2 seeds grew on the 12 F1 hybrid seed parents and had a much narrower size range (with max. values of 1,296 and 1,201 mg, respectively). The seeds of the half inbred generations RBC and F2 were slightly larger than the half inbred BC seeds growing on the parental inbred lines, but this difference was not significant (cf. Fig. 1; for RBC vs. BC: P < 0.62).

Generation means for faba bean mature seed weight, realized for seed growing on inbred and non-inbred seed parent plants
Mature seed weight and juvenile plant weight were, based on the 76 genotypes, correlated (r = 0.91**). Thus, about 83% of the variation in young plant weight of these genotypes was a reflection of the variation in sown seed weight. Hedin yielded the lightest seedlings (6.4 g) and HP58 the heaviest (13.7 g) among the four parents. The overall heaviest seedlings resulted from the genotype RBC[HP58 × (Gobo × HP58)] with 14.6 g. Analysed per generation, the variance of plant weight explained by seed weight was between 64% (parents) and 93% (F2). For plant weight, the mean superiority of F1 over parents was 14.3%; the difference between the two generation means was highly significant (P < 0.02). Again, the BC generation mean was nearly exactly the average of P and F1 (Fig. 2). Half inbred plantlets from seed that grew on F1 seed parent (RCB, F2) were significantly (P < 0.03) heavier than half inbred plantlets from seed that grew on homozygous plants (BC).

Generation means of above-ground faba bean plantlet weight; plantlets developed from seed that grew on inbred and non-inbred seed parent plants
The diallel analysis isolated the genetic components of mature seed weight and young plant weight variation (Tables 4, 5). Embryos’ direct additive effects and direct dominance effects were highly significant sources of variation. The estimates of direct additive effects varied from +179 mg (HP58) to −84 mg (Gobo), the estimates of direct dominance effects from +172 mg (D ij for the combination of HP58 and HP69) to −77 mg (D ii for HP58). For seed weight, one standard deviation (SD) of the direct dominance effects amounted to 44 mg, thus to 26% of one SD of the direct additive variance (171 mg). One maternal additive SD was 64% of one direct additive SD; maternal dominance effects were not significantly different from zero. Cytoplasmic effects proved to be significant, albeit rather small.
For plant weight, the analyses also showed high significance for direct additive and dominance variance, and again small cytoplasmic differences among the parents existed and were transmitted to their offspring. One SD of seeds’ direct dominance effects amounted to 594 mg and thus to 56% of one SD (1,054 mg) of embryos’ direct additive effects. In contrast to mature seed weight, here the maternal dominance effects rather than the maternal additive effects were significant.
Discussion
Seed size of faba bean is of high relevance, not least for variety identification and classification (Crofton 1997). Furthermore, it is one of the traits that underwent major changes in its expression and genetic makeup under domestication (Suso and Cubero 1986). Here, mature seed weight was studied focussing on its embryos’ heterotic impact.
One of the yield components in the experiments, number of seeds per pod, could not be controlled, and it varied between two to four seeds per pod. The number of seeds in crossed pods was smaller than in selfed pods. This difference (2.9 < 3.2) corroborated our earlier findings. Since the selfing (tripping) and crossing (emasculation and pollination) are partly different procedures, a possible impact of this difference on the estimate of embryo heterosis should be addressed. This issue was observed by analysing the impact of seed number per pod on single seed weight. The slope of the regression predicted an average decrease of seed weight of 1.02% per increase of seed number by one seed. The regression coefficient was not significantly different from zero. Thus, the differences between generations in seeds per pod can account for only very small differences in seed weight.
The genotypic value of parents for the trait ‘mature seed weight’, assessed from selfed seed, had a major effect on the seed weight of crossed seed growing on them. Weight of selfed seed of HP58 averaged 1,459 mg, and the three F1 seed genotypes and six BC seed genotypes growing on HP58 weighed between 1,419 and 1,680 mg. Selfed seed on Hedin weighed 519 mg, whereas crossed seed on Hedin weighed 465–660 mg. Only two such cases occurred with crossed seed being lighter than parental selfed seed, both were BC involving Hedin and Gobo as parents; apparently these two lines expressed a negative, specific interaction. Across all genotypes, the rank correlations were r = 0.867** between P and F1, r = 0.856** between P and BC, and r = 0.934** between F2 (the F2 denoting the genotypic value of F1 seed parent) and RCB (P < 0.01 throughout); the last comparison does not include a heterotic difference between selfed and crossed seed. Thus, the very high amount of 75–87% of the variation of crossed seed weight was determined by the seed weight of the seed parent. The determination is composed of maternal effects, cytoplasmic effects and the seed parent’s contribution to the genotype of the embryo (Table 3). In a subsequent project, the chemical composition of homozygous and heterozygous mature seed will be studied, to allow for a more sophisticated view into embryo and seed heterosis.
Lemontey et al. (1999) described how the seed parent influenced pea mature seed size by controlling mitotic activity during early embryogenesis and final endoreduplication level and probably the cotyledon cell size in mature seeds. In faba bean, early embryo growth was mainly maternally controlled, whereas the transition into maturation implied a switch to filial control (Weber et al. 2005; Sreenivasulu et al. 2006), suggesting that a regulatory network initiated maturation and the accumulation of storage products. This included transcriptional and physiological reprogramming mediated by sugar and hormone-responsive pathways. Regulatory transcription factors were essential elements governing seed-specific gene expression and coordinating environmental and hormonal signals.
The significant difference between F1 and BC amounted to 5.0%. These two seed generations were both produced by manual crossing, the mean number of seeds per pod of these two generations was actually the same (2.89). Hence, the higher seed weight of the F1 seeds cannot be attributed to a smaller number of seeds per pod; the alternative explanation is embryo heterosis. The comparison of the generation means F1 and P demonstrated a significant difference of 11.1%, understood as embryo heterosis.
Based on Eq. 1, using the direct dominance estimates from the diallel analysis, the average heterosis amounted to 10.6%, being very similar to the 11.1%. Duc et al. (2001) reported 2–8% heterotic increment of faba bean hybrid seed weight over selfed seed weight. Davies (1975) gave examples for seed weight heterosis in pea: in one cross, hybrid seed with the small-seeded parent as seed parent yielded a heterotic increment of 52%, for the reciprocal hybrid it was 9%, hence hybrid seed was on average 20% heavier than selfed seed. On average across 15 crosses, mean seed heterosis was 18%.
The plant weight heterosis taken directly from F1 versus P, being 14.3%, was as well very similar to the estimate based on Eq. 1, which was 14.5%.
The apparent linear relationship between inbreeding coefficient and both seed weight (Fig. 1) and plant weight (Fig. 2), based on comparing the generations F1, BC and P, corroborates general experience and simple quantitative genetic models for heterosis (Falconer and Mackey 1996). The results of RBC when compared with BC are a hint at superior maternal impact delivered by a hybrid seed parent versus an inbred seed parent. This phenomenon has long been known for maize. Schnell and Singh (1978) reported that three-way hybrids grown from seed that grew on hybrid seed parents resulted in significantly higher yield than the same genotypes grown from seed that developed on fully inbred seed parents; this reflects the initial advantage of seedlings whose seed parents were a hybrid instead of a pure line.
For mature seed weight, the difference between RBC and F2 compared with BC, i.e. the seed parent heterotic impact, was positive, albeit not significant. This is corroborated by the non-significant impact of maternal dominance effects on seed weight (Table 4). In contrast, for plant weight, the differences between RBC and BC and between F2 and BC were significant and the maternal dominance effects were indeed a significant source of variation (Table 5). The conclusion is that hybrid seed parents did not produce significantly heavier mature seeds than inbred seed parents. Seemingly the maximum of eight pods that was allowed to grow on a seed parent plant allowed a very good assimilate supply even to seed growing on the inbred seed parents. On the other hand, the seed harvested from hybrid seed parents developed into larger young plants than isogenic, approximately similar weighing seed harvested from inbred seed parents. Since the differences in weight were small, a difference in the composition of these two types of seed, e.g. in energy content, could still explain these findings. Composition of these seed was not studied here.
The 12 F1 hybrid results for seed weight were highly significantly correlated (Fig. 3; r = 0.748**) with their heterosis as estimated by Eq. 1, showing the impact of embryo heterosis on the phenotypic mature weight, in spite of the large impact of the seed parent seed size. The two reciprocal F1 hybrids of the three crosses with HP58 were very different in seed size (Fig. 3). Nevertheless, it is important to notice that in all three crosses the hybrid seed became larger than the HP58 selfed seed, although these hybrid seeds were pollinated by a much smaller seeded pollinator (F1(l × s) vs. P(l)). For the cross of HP69 with HP58, the difference between the F1 seed HP58 × HP69 and the selfed seed HP58 was significant (F1[P i × P i ] − P i ; P < 0.05). This is a further indication of embryo heterosis. Since the pollinator P i is small-seeded, its pollen grains are considered unlikely to transfer genes for large seeds, thus the effect is very probably due to heterosis. Following quantitative genetic notation, Eq. 2 shows (cf. Table 3) that a significant, positive heterotic increment HI can result only if the heterotic value [D ij − D ii ] surpasses the additive value [A i − A j ] (NB: A j , the smaller-seeded parent’s additive impact, and D ii , the larger-seeded parent’s inbreeding effect, are negative contributions):
Heterosis according to Eq. 1 for the two traits was correlated (r = 0.657; P > 0.10; Fig. 4); with only six data pairs, the relationship was not significantly different from zero. For plant weight, the crosses between the basically unrelated parents, i.e. the three crosses with Gobo, showed more heterosis than the crosses between the basically related parents. The contrary was true for seed weight. The three unrelated crosses with Gobo ranked first, third and fourth for plant weight heterosis, and only second, fourth and sixth for seed weight. The highest heterosis for seed weight occurred in the cross between the related lines HP69 and HP58, whereas the highest plant weight heterosis occurred in the unrelated cross between Gobo and HP58. A strong influence of the parental genetic distance on the amount of heterosis is under dispute even for heterosis of plants. Lee et al. (2007) showed that, in maize, substantial genome-wide heterozygosity was not a requirement for the expression of high heterosis. Chakraborty (1981) had already stressed that, with a limited number of loci coding a trait (like N < 10), a tight correlation between trait heterozygosity (representing heterosis) and genome-wide heterozygosity (representing parental relationship) is not expected. Grain legume seed weight is probably based on few loci (Maughan et al. 1996; Liu et al. 2007). The present data allow the conclusion that a large parental genetic difference for seed size, such as between HP69 and HP58, is more important for seed heterosis than a low genetic relationship like that between Gobo and HP58. The conclusion for young plant biomass is the contrary: many loci might be implied and crossing related parents does not allow maximum offspring heterozygosity of this trait. Further research has to uncover the genes that actually code for the large differences in seed size of faba bean, possibly throwing light on this main feature of domestication of plants.

Relationship between mature seed weight of faba bean hybrid seed (with either large- or small-seeded parent as seed parent) and embryo heterosis. Parental values are given in addition

Relationship between embryo heterosis for mature seed weight and young plant heterosis for above-ground biomass yield
V. faba is partially allogamous with on average about 50% of cross-fertilization (Link 1990). Thus, a repeatedly mentioned idea (Berthélem, pers. comm., 1989) has been to increase the frequency of hybrid seed in a seed lot by sorting, i.e. by mechanical selection in favour of the larger and heavier, presumably hybrid, seeds. Unfortunately, the variation in seed size and weight from pod to pod and plant to plant and the variation due to segregation seem to be too large compared to the size of seeds’ direct heterosis to make this idea become implementable and useful in the production of hybrid-enriched seed lots. Dieckmann (2007) studied the influence of genetically unrelated versus self-pollen on grain yield, using pollen-sterile faba beans in related experiments (self-pollen was provided by isogenic maintainer lines). No yield advantage provoked by unrelated pollen was verified. In maize, corresponding findings were more promising. As in faba bean, embryos of maize hybrid seeds growing on homozygous seed parents were 12% heavier than selfed embryos (Djisbar and Gardner 1989). Heterosis for embryo size was even larger (31%) in immature maize embryos (Meyer et al. 2007). Such findings found an agricultural application. The past decade has seen US corn fields planted to high-oil corn following the TopCross® patent, where a male-sterile version of an otherwise conventional hybrid is mixed with some 10% of an unrelated high oil pollinator maize. This results in a markedly increased embryo size, i.e. oil content of the seed growing on the conventional hybrid plants, and normally in no corresponding dry matter yield reduction per field area, i.e. the crop produces a higher energy yield (Thomison and Geyer 1999; Weingartner et al. 2002). Different from this approach, conventional single cross hybrids realize full hybrid vigour in the whole plant except in the seed, which are half way inbred. One way to overcome this disadvantage would be apomictic production of seeds on hybrid cultivars.
References
Becker HC, Link W (2000) Heterosis and hybrid breeding. Vortr Pflanzenzüchtg 48:319–327
Chakraborty R (1981) The distribution of the number of heterozygous loci in an individual in natural populations. Genetics 98:461–466
Chen G, Zhu J (2003) QGA Station 1.0. Software for classical quantitative genetics. Developed by Guobo Chen and Jun Zhu. Institute of Bioinformatics, Zhejiang University, China. http://ibi.zju.edu.cn/software/qga/index.htm (cited 21 Nov 2007)
Crofton GRA (1997) The principal seed characters of field beans (Vicia faba L. (Partim)) in relation to variety classification. Plant Var Seeds 10:81–94
Crow JF (1999) Dominance and overdominance. In: Coors JG, Pandey S (eds) Genetics and exploitation of heterosis in crops. American Society of Agronomy, Madison, WI, pp 49–58
Dani RG, Kohel RJ (1989) Maternal effects and generation mean analysis of seed-oil content in cotton (Gossypium hirsutum L.). Theor Appl Genet 77:569–575
Davies DR (1975) Studies of seed development in Pisum sativum. Planta 124:297–302
Dieckmann S (2007) Genetische Analyse von Heterosis im jüngsten Stadium einer Pflanze, als Embryo, am Beispiel der Fababohne (Vicia faba L.). PhD thesis. University of Goettingen, Germany
Djisbar A, Gardner FP (1989) Heterosis for embryo size and source and sink components of maize. Crop Sci 29:985–992
Duc G, Moessner A, Moussy F, Mousset-Déclas C (2001) A xenia effect on number and volume of cotyledon cells and on seed weight in faba bean (Vicia faba L.). Euphytica 117:169–174
Dure LSIII (1975) Seed formation. Ann Rev Plant Physiol 26:259–278
Falconer DS, MacKey TFC (1996) Introduction to quantitative genetics, 4th edn. Longman Scientific, New York
Focke OW (1881) Die Pflanzenmischlinge. Bornträger, Berlin
Frascaroli E, Canè MA, Landi P, Pea G, Gianfranceschi L, Villa M, Morgante M, Pè ME (2007) Classical genetic and quantitative trait loci analyses of heterosis in a maize hybrid between two elite inbred lines. Genetics 176:625–644
Hoecker N, Keller B, Piepho H-P, Hochholdinger F (2006) Manifestation of heterosis during early maize (Zea mays L.) root development. Theor Appl Genet 112:421–429
Kiesselbach TA (1960) The significance of xenia effects on the kernel weight of corn. Res Bull Neb Agric Exp St 191:1–30
Lee EA, Ash MJ, Good B (2007) Re-examining the relationship between degree of relatedness, genetic effect, and heterosis in maize. Crop Sci 47:629–635
Lemontey C, Mousset-Déclas C, Munier-Jolain N, Boutin JP (1999) Maternal genotype influences pea seed size by controlling both mitotic activity during early embryogenesis and final endoreduplication level and cotyledon-cell size in mature seed. J Exp Botany 51:167–175
Link W (1990) Autofertility and rate of cross-fertilization: crucial characters for breeding synthetic varieties in faba beans (Vicia faba L.). Theor Appl Genet 79:713–717
Liu B, Fujita T, Yan Z-H, Sakamoto S, Xu D, Abe J (2007) QTL mapping of domestication-related traits in soybean (Glycine max). Ann Bot. doi:10.1093/aob/mcm149
Lou X, Zhu J, Zhang Qu, Zang R, Chen Y, Yu Zh, Zhao Y (2005) Genetic control of the opaque-2 gene and background polygenes over some kernel traits in maize (Zea mays L.). Genetica 124:291–300
Maughan PJ, Saghai Maroof MA, Buss GR (1996) Molecular-marker analysis of seed-weight: genomic locations, gene action, and evidence for orthologous evolution among three legume species. Theor Appl Genet 93:574–579
Meyer C, Törjék O, Mecher M, Altmann T (2004) Heterosis of biomass production in Arabidopsis. Establishment during early development. Plant Physiol 134:1813–1823
Meyer S, Pospisil H, Scholten S (2007) Heterosis associated gene expression in maize embryos 6 days after fertilization exhibits additive, dominant and overdominant pattern. Plant Mol Biol 63:381–391
Nicolaisen W (1931) Über quantitative Xenien bei Roggen und Erbsen. Zeitschr Pflanzenzüchtg 17:265–276
Schnell FW, Singh IS (1978) Epistasis in three-way crosses involving early flint and dent inbred lines of maize. Maydica 23:233–238
Sreenivasulu N, Radchuk V, Strickert M, Miersch O, Weschke W, Wobus U (2006) Gene expression patterns reveal tissue-specific signaling networks controlling programmed cell death and ABA-regulated maturation in developing barley seeds. Plant J 47:310–327
Stefansson BR, Hougen FW (1964) Selection of rape plants (Brassica napus) with seed oil practically free from erucic acid. Can J Plant Sci 44:359–364
Suso MJ, Cubero JI (1986) Genetic changes under domestication in Vicia faba. Theor Appl Genet 72:364–372
Syed NH, Chen ZJ (2005) Molecular marker genotypes, heterozygosity and genetic interactions explain heterosis in Arabidopsis thaliana. Heredity 94:295–304
Thomison PR, Geyer AB (1999) Evaluation of TC blends® used in high oil maize production. Plant Var Seeds 12:99–112
Tschermak E (1931) Über Xenien bei Leguminosen. Zeitschr Züchtg (Reihe A, Pflanzenzüchtg) 16:73–81
Tschermak E (1932) Bemerkungen über echte und falsche Größen-Xenien. Zeitschr Züchtg (Reihe A, Pflanzenzüchtg) 17:447–450
Weber H, Borisjuk L, Wobus U (2005) Molecular physiology of legume seed development. Ann Rev Plant Biol 56:253–279
Weingartner U, Kaeser O, Long M, Stamp P (2002) Combining cytoplasmic male sterility and xenia increases grain yield of maize hybrids. Crop Sci 42:1848–1856
Zhu J (1997) Analysis methods for genetic models. Agricultural Publication House of China, Beijing
Zhu J, Weir BS (1994) Analysis of cytoplasmic and maternal effects. I. A genetic model for diploid plant seeds and animals. Theor Appl Genet 89:153–159
Acknowledgments
We acknowledge the financial support by DFG, Priority Program SPP1149 ‘Heterosis in Plants’ (coordinated by Prof. Dr. A.E. Melchinger). We acknowledge the helpful comments by Prof. Jun Zhu, Dr. Hans Weber and Dr. Lamiae Ghaouti, and the many thoughtful hints by Prof. Dr. Fred Stoddard.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by D. Hoisington.
Contribution to the special issue “Heterosis in Plants”.
With great respect we have pleasure in dedicating this paper to Professor Dr. Dr. h.c. H.H. Geiger, Hohenheim, in honour of his 70th birthday.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Dieckmann, S., Link, W. Quantitative genetic analysis of embryo heterosis in faba bean (Vicia faba L.). Theor Appl Genet 120, 261–270 (2010). https://doi.org/10.1007/s00122-009-1057-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-009-1057-7