
Abstract
Fish are known to pose strong effects on invertebrate abundance, species richness and assemblage structure. Littoral vegetation may play a crucial role as a refuge for invertebrates vulnerable to fish predation. We studied relative densities and taxonomic composition of water mites, aquatic beetles and bugs in large lake-like ponds with different fish status (fish-free and containing fish) and mesohabitats (emergent littoral vegetation and open water zone). The macroinvertebrate taxa differed in their responses to the fish presence and in mesohabitat preferences. The density and species richness of water mites were greater in fish-containing ponds, while no differences were found between littoral and open-water habitats. In contrast, beetles were far more numerous and species-rich in fish-free ponds and in littoral vegetation. Total densities of aquatic bugs were non-significantly higher in fish-containing ponds, and they preferred littoral areas, but species richness was independent of fish presence and mesohabitat. No statistical interactions between fish presence and the densities of individual macroinvertebrate groups in the littoral habitat were detected, indicating that their use of emergent littoral vegetation was not an antipredator response to fish. The assemblages of the three macroinvertebrate taxa exhibited nested structures of a different order, consistent with their species richness patterns. Our research stresses the importance of littoral vegetation for the distribution and abundance of aquatic insects; however, high fish presence may not affect or may even benefit ecologically important macroinvertebrate groups, such as water mites or bugs.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Predator–prey relationships between fish and aquatic macroinvertebrates are crucial in structuring freshwater communities (Healey 1984; Evans 1989; Bendell and McNicol 1995; Hanson and Riggs 1995; Batzer et al. 2000). Fish are considered to be highly effective top predators of aquatic food chains; macroinvertebrates are a basic food for many species (Michel and Oberdorff 1995; Specziár et al. 1997; Kottelat and Freyhof 2007). Research involving manipulations of the fish presence or abundance has shown dramatic changes in the density, species composition, and size structure of macroinvertebrate communities (Eriksson et al. 1980; Morin 1984; Diehl 1992; Carlisle and Hawkins 1998; Zimmer et al. 2001; Leppä et al. 2003; Nieoczym et al. 2020; but see Pierce and Hinrichs 1997). Many macroinvertebrate taxa are virtually absent from fish-dominated waters (de Mendoza et al. 2012; Nieoczym and Kloskowski 2015; Toro et al. 2020). Fish also exert a strong impact on macroinvertebrate distribution within water bodies (Oscarson 1987; Tolonen et al. 2003; Schilling et al. 2009). However, less vulnerable species with defensive adaptations may persist in the presence of fish (Peckarsky 1982; Batzer and Wissinger 1996; Pope and Hannelly 2013).
The outcomes of interactions between fish and invertebrates in large waterbodies, consisting of distinct mesohabitats, are influenced by the presence and character of complex-structured vegetation (Gilinsky 1984; Diehl 1992; Rennie and Jackson 2005; Kajgrova et al. 2021). Aquatic vegetation enhances habitat and resource diversity and influences abiotic parameters such as light penetration depth, temperature, dissolved organic matter and oxygen concentration in the water (Diehl and Kornijów 1998; Lürig et al. 2021). The efficiency of fish predation is believed to be greater in open water with no or sparse vegetation than in habitats with a complex macrophyte structure (Peckarsky 1984; Lima and Dill 1990; Diehl and Kornijów 1998; de Mendoza et al. 2012; but see Carlisle and Hawkins 1998). Prey is less conspicuous to visual fish predators in vegetated habitat. Moreover, dense plant structure impairs the fish predator’s ability to move and to pursue the prey (Gilinsky 1984; Winfield 1986; Wirsing et al. 2010). Many aquatic macroinvertebrates, including numerous representatives of Odonata (larvae), Heteroptera and Coleoptera, have been reported to prefer or even restrict their distribution to vegetation-rich littoral habitats, which are believed to protect them against predators (Zimmer et al. 2000; Tolonen et al. 2003).
The primary goals of this study were to investigate the fish impact on the abundance and composition of aquatic macroinvertebrates – water mites (Hydrachnidia), aquatic beetles (Coleoptera) and bugs (Heteroptera) – in large lake-like ponds, focusing on the role of emergent littoral vegetation and the open water zone as the principal pond mesohabitats. These taxa, which are often dominant components of both temporary and more permanent pond communities (Nicolet et al. 2004; Buczyńska et al. 2007; Stryjecki 2007; Broyer and Curtet 2011; Nieoczym and Kloskowski 2015), are mostly mobile and can move between mesohabitats (Klausnitzer 1996; Papáček 2001; Bouchard et al. 2014; Proctor et al. 2015). We assessed how their assemblages varied between two mesohabitats of ponds with and without fish. We expected that the densities of these macroinvertebrate groups would be lower in ponds containing fish (cf. Papáček 2001; Leppä et al. 2003; Gioria 2014), and that many species would use the near-shore emergent vegetation to mitigate predation pressure by fish, although in the absence of fish they might be found in the open water zone as well (Bennett and Streams 1986; Oscarson 1987; Kloskowski et al. 2020). However, species with antipredator adaptations against fish, such as secretion of toxic substances or defensive morphological traits, would not need refuges in a structured vegetated habitat (Peckarsky 1984; Scrimshaw and Kerfoot 1987).
Methods
Characteristics of ponds
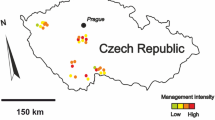
The study was carried out in human-made, lake-like fishponds scattered in eastern Poland (51°17–33’ N, 22°15–26’ E) in 2012–2013. The ponds were eutrophic water bodies used for semi-extensive culture of carp species, with a mean depth of about 0.7–1.4 m, filled with water in early spring and drained in autumn (see Korinek et al. 1987 for a description of carp pond ecosystems). Two types of ponds were included: 17 ponds without fish (hereafter ‘fish-free ponds’) and 24 ponds stocked with fish (‘fish-containing ponds’). The fish-free ponds were those that would be stocked with fish fry by local fisheries in late May and early June, so the fieldwork was terminated in both years on 6–10 June, before or shortly after the introduction of fish larvae to these ponds. Fish-containing ponds were stocked with 1- and 2-year-old common carp Cyprinus carpio in early spring at an individual weight range of about 30–600 g (biomass density 80–300 kg ha–1). Small amounts (stocking biomass of less than 10 kg/ha, similar or smaller individual size) of other carp species, i.e., grass carp Ctenopharyngodon idella, bighead carp Hypophthalmichthys nobilis, and crucian carp Carassius carassius, were added. The area of the fish-containing ponds was 1–14 ha, except one pond of 18 ha. The area of the fish-free ponds varied between 1 and 8 ha. An advantage of this study system for investigating the role of fish in determining the composition of aquatic macroinvertebrate assemblages was that all ponds were comparable in depth, profile and hydroperiod length, whereas natural fish-bearing and fishless lakes typically differ in some geomorphic and water chemistry characteristics and in their hydrological history (Schilling et al. 2008; Kurek et al. 2010).
We sampled macroinvertebrates and measured abiotic variables in two within-pond mesohabitats: near-shore emergent vegetation (hereafter ‘littoral habitat’) and the open water zone. The emergent vegetation habitat along the pond shores tended to be shallower than the rest of the pond, as it included the land-water interface zone; however, the central parts of the ponds were shallow as well. Hence, the use of the term ‘littoral’ with regard to near-shore vegetation is a simplification, because the shallow ponds are almost entirely littoral, and submerged macrophytes, unless adversely affected by fish activity, could occur throughout nearly the entire pond, especially later in the growing season. However, submerged macrophytes were scarce in the study ponds during the study period (May to early June); they did not reach the water surface and did not form dense aggregations. Stands of littoral vegetation surrounded the open water area mainly as narrow emergent macrophyte belts, usually 1–5 m wide, consisting primarily of Phragmites australis and Typha angustifolia, but also as large emergent vegetation patches up to 100 m wide. Thus, the structurally complex littoral mesohabitat clearly differed from the open-water mesohabitat, where submerged vegetation apparently lacked the complexity necessary for the effective protection of macroinvertebrates from fish. The areal extent of the vegetated habitat was determined from digitized aerial photos. The open-water abundance of submerged vegetation was measured as dry biomass of samples collected with a Bernatowicz grabber (for details on grabber construction see Nieoczym and Kloskowski 2015) at 10 haphazardly chosen sites roughly evenly spaced along the longer axis of the pond. The submerged vegetation was sampled within a week after macroinvertebrate sampling (see below). Among the dominant plants, Stuckenia pectinata was present in both types of ponds, while Potamogeton lucens and Chara sp. were noted mainly in fish-free ponds.
Abiotic parameters were measured at mid-depth in both within-pond habitats shortly before the activity traps were set in the ponds. Temperature, dissolved oxygen concentration and electrical conductivity were measured in situ using a multifunction meter (CX-401, Elmetron, PL). The load of total suspended solids was analysed immediately upon arrival at the laboratory using a PASTEL-UV SECOMAM spectrophotometer; the estimates were means from three water samples per mesohabitat per each pond.
Macroinvertebrate sampling
Pond macroinvertebrates were sampled using activity traps in 7 fish-free ponds and 10 fish-containing ponds in 2012 and in 10 vs. 14 in 2013. The samples were collected between 15 and 19 May in 2012 and between 17 May and 4 June in 2013 (the phenological season started slightly later in 2013). Traps were 1-L plastic cylinders with funnels 100 mm at the large end and 23 mm at the narrow end (Nieoczym and Kloskowski 2015). Ten traps were placed on the bottom of each pond in both mesohabitats (in total 20 traps per pond). In the littoral habitat, the traps were set along the shoreline in areas where the belts of emergent vegetation were at least 10 m wide, with approximately even distances between each trap. The traps were placed within extensive patches of scattered plants or next to patches of dense vegetation. The traps set in open water were located 5–15 m from the emergent vegetation border. After 24 h of trap deployment, the captured macroinvertebrates were preserved in 70% ethanol and then transported to the laboratory and identified, usually to species level. Macroinvertebrate abundances are presented as relative densities (number of individuals per 10 traps from each mesohabitat or per 20 traps for the whole pond). Funnel traps were used because they are effective at capturing mobile invertebrates, like most of the species studied here; they are less efficient at trapping sedentary species (Hyvönen and Nummi 2000; Zimmer et al. 2000; Bouchard et al. 2014; Liao et al. 2020). However, we were primarily interested in species able to move between mesohabitats. Water mites can be highly mobile and show strong exploratory behaviour (Pieczyński 1976; Proctor et al. 2015); mite species collected in this study have numerous swimming setae (Di Sabatino et al. 2010; Gerecke et al. 2016) enabling horizontal and vertical movements. Most of the locally recorded aquatic beetles and bugs are skilled swimmers; some species which do not swim actively can crawl on the bottom and submerged macrophytes (Jäch and Balke 2008; Schilling et al. 2008; Lytle 2015). Although the traps may sample only a single stratum of water, they are convenient to handle in dense vegetation and are not biased by the underestimation of mobile invertebrates scared away by field workers using hand-held devices (e.g., a dipnet or corer). Finally, the activity trap method avoids biases resulting from daytime/night-time sampling (Hilsenhoff 1987; Murkin et al. 1983). In total, water mites (deutonymphs and adults) constituted 34.3%, beetles (larvae and imagines) 22.1%, and bugs (nymphs and imagines) 18.5% of the collected macroinvertebrates. Other taxa found in the traps were Odonata, Oligochaeta, Diptera, Mollusca, Ephemeroptera, Hirudinea and Trichoptera, which contributed 2–5% each to overall macroinvertebrate abundance.
Statistical analyses
Canonical correspondence analysis (CCA) was conducted to determine how the environmental variables accounted for variation in the macroinvertebrate assemblages. We made separate analyses for water mites and aquatic insects (beetles and bugs were considered together due to the small number of bug species; when the two groups were considered separately similar CCA graphs were obtained, with fish status as the only significant environmental variable). We combined data from both mesohabitats, i.e., we used numbers of captured individuals from 20 traps per pond and averaged values of abiotic parameters recorded in the littoral habitat and open water zone. Fish status (fish-free vs. fish-containing ponds), the amount of emergent vegetation cover in the pond, and the abiotic parameters listed in Table S1 were included in the preliminary CCA analyses. We used the amount of emergent vegetation cover (expressed in ha) rather than the proportion of pond area covered by emergent vegetation because the latter was correlated with pond fish status. The data were screened using Pearson and biserial correlations to identify inter-correlations among environmental variables; redundant variables were omitted from further analysis (see Results). The forward-selection procedure was used; the statistical significance of added variables was tested using a Monte Carlo test (999 permutations). In addition, global Monte Carlo permutation tests were performed to test for the existence of a relationship between environmental characteristics and species composition (ter Braak and Šmilauer 2018). Since the fieldwork was carried out during two seasons, and the ponds occurred in four scattered aggregations (“pond clusters”) which received water from different sources, the year of research and pond cluster identity were included in the CCA analyses as covariables. The option of down-weighting rare species was applied. The ordination analyses were performed in Canoco 5.1 software (Biometris, Wageningen University & Research and Petr Šmilauer).
Generalized linear mixed models (GLMMs) with a log link and negative binomial distribution were used to compare the relative densities (numbers of individuals per 10 traps per mesohabitat) and species richness of the three groups of macroinvertebrates in fish-free and fish-containing ponds and in two mesohabitats (littoral habitat and open water zone). Pond identity nested within pond cluster was used as a random effect because two data points (catches in open water and in littoral habitat) were obtained from each pond. Only pond identity was used for analyses of species richness because models with nested random factors did not converge. The abiotic parameters of the ponds were compared in a similar manner, using GLMMs with identity link and normal distribution. The models initially contained all fixed factors and their interactions; however, non-significant interactions were removed to improve model fit. The GLMM analyses were conducted in Genstat v. 15 software (VSN International).
Assemblage nestedness of the three macroinvertebrate groups in the ponds was examined separately for each taxonomic group using NODF (a nestedness metric based on overlap and decreasing fill; Almeida-Neto et al. 2008). We obtained significances for presence-absence matrices of species occurrence by comparison with 500 null matrices constructed considering the proportional column and row totals algorithm. The analyses were performed using NeD software (Strona et al. 2014). To assess the role of fish presence in structuring species assemblages among the ponds, we calculated t tests on the ranks of assemblages from individual ponds in the maximally packed matrix with respect to the fish status of the ponds.
Results
Differences in environmental conditions between and within ponds
On average, the within-pond area covered by emergent macrophytes was 1.1 ± SE 0.3 ha with a range 0.2–5 ha in fish-free ponds (mean percentage of pond area covered by emergent macrophytes, 35.6 ± 3.8%) and 1.9 ± 0.3 ha, range 0.2–7 ha, in fish-containing ponds (percentage of pond covered 24.5 ± 3.0%). The average area of emergent vegetation did not significantly differ between fish-containing and fish-free ponds (t test, t39 = − 1.67, p = 0.103). The open-water abundance of submerged vegetation, was greater in fish-free ponds than in fish-containing ponds (48.0 ± 11.0 vs. 12.5 ± 5.1 g m− 2; t test, t39 = 3.22, p = 0.003).
Total suspended solids, dissolved oxygen concentration and electrical conductivity were higher in fish-containing ponds than in fish-free ponds, while water temperatures were similar. Dissolved oxygen concentrations were greater in open water, while conductivity was higher in littoral habitat. Water temperature and total suspended solids did not differ between mesohabitats (Table S1, S2).
Macroinvertebrate assemblage composition and distribution among ponds
In total 493 Hydrachnidia individuals belonging to 30 species were collected. The most frequently occurring species were Limnesia maculata (present in 36.6% of ponds), Piona coccinea (34.1%), P. neumani (29.3%), P. imminuta (26.8%), P. pusilla (22.0%) and Tiphys ornatus (22.0%). Heteroptera were represented by only 5 species (266 individuals), most frequently Ilyocoris cimicoides (58.5% of ponds), Sigara falleni (48.8%) and Notonecta glauca (12.2%). A total of 21 Coleoptera taxa (Hydroporus larvae were determined to genus level) were identified from the ponds (based on the sample of 313 individuals). The most frequently found species were Agabus undulatus (29.3% of ponds), Graphoderus austriacus, Rhantus latitans (22.0% each), Cybister lateralimarginalis, Noterus clavicornis, N. crassicornis (19.5% each) and Macroplea appendiculata (17.1%). A complete list of species with their average percentage abundances in ponds differing in fish status and in the two mesohabitats is presented in the Supplementary Materials (Table S3, S4).
Prior to CCA, the most inter-correlated variables were removed from the analysis. Water temperature was correlated with dissolved oxygen, total suspended solids, and electrical conductivity (all p < 0.021) and hence omitted. Pond area was significantly correlated with emergent vegetation cover and fish status (both p ≤ 0.02). Abundance of submerged macrophytes was also removed due to correlation with fish status (p = 0.003). According to CCA, water mite assemblages were most strongly correlated with dissolved oxygen concentration (λa = 0.36, F = 3.17, p = 0.001), explaining 11.0% of the overall variation, followed by electrical conductivity (λa = 0.2, F = 1.95, p = 0.029), explaining 6.1% of variation, and fish status in the pond (λa = 0.19, F = 1.75, p = 0.036), explaining 5.8% of variation (Fig. 1). Among the most common species, only P. pusilla was significantly correlated with dissolved oxygen concentration (r = 0.281), and P. coccinea was significantly associated with fish-containing ponds (r = 0.251). The inter-set correlations of the variables with the CCA ordination axes indicated that the first axis captured a gradient of dissolved oxygen concentration (r = − 0.736), a gradient of electrical conductivity (r = − 0.555), and differences in the fish status in the ponds (fish-free ponds, r = 0.608). The second axis was largely defined by fish status (fish-free ponds, r = 0.482) and total suspended solids (r = 0.471). Monte Carlo permutation tests on the trace value (F = 1.78, p = 0.003, trace value = 0.945) and on the value of axis 1 (F = 3.59, p = 0.001, eigenvalue = 0.460) indicated significant associations between species distribution and environmental variables.

Biplots from canonical correspondence analyses for pond water mites. The variables used in the model explained 29% of the variation in the water mite assemblage. The first axis explained 14% of the variation in water mite relative abundance, the second 8.1%. Significant variables (p < 0.05) are shown in bold. Species names are abbreviated using three letters each of genus and species, defined in Table S3
Fish status in the pond was the most important factor in the CCA model of the influence of environmental variables on the relative abundance of aquatic beetles and bugs (λa = 0.31, F = 3.57, p = 0.001), explaining 9.3% of the overall variation (Fig. 2). No species was significantly associated with fish-containing ponds. Of the most common aquatic beetles, the species significantly correlated (p < 0.05) with fish-free ponds were M. appendiculata (r = 0.392), G. austriacus (r = 0.331), R. latitans (r = 0.284) and Agabus undulatus (r = 0.266). Dissolved oxygen concentration only approached significance (λa = 0.15, F = 1.81, p = 0.052). The other environmental factors were insignificant (all λa ≤ 0.12, F ≤ 1.47, p > 0.125). The first axis of the CCA of insect-environment relationships was defined by the fish status of the pond (correlation with fish-free ponds, r = 0.762), gradients of dissolved oxygen concentration (r = − 0.570) and electrical conductivity (r = − 0.610), while the area of emergent vegetation in the pond (r = 0.579) and total suspended solids (r = – 0.382) were associated with the second axis. Both the first CCA axis (F = 3.56, p = 0.004, eigenvalue = 0.342, Monte Carlo permutation test) and the entire ordination (F = 1.95, p = 0.001, trace value = 0.795) accounted for more variation than expected by chance.

Aquatic insect species-environmental variable biplots (Heteroptera represented as circles, Coleoptera as triangles). The environmental variables used in the model explained 24% of the variation in the insect assemblage. The first axis explained 10.3% of the variation in insect relative abundance, the second 4.4%. Significant variables (p ≤ 0.05) are shown in bold. Species names are abbreviated using three letters each of genus and species, defined in Table S4
Macroinvertebrate abundances and assemblage structure
The GLMMs showed that the total relative densities and species richness of the three macroinvertebrate groups varied in their responses to the fish presence and mesohabitat. The densities and species richness of water mites were higher in fish-containing ponds, but did not differ between littoral habitat and open water (Tables 1 and 2; Fig. 3A). Aquatic bugs were non-significantly more numerous in fish-containing ponds (Table 1; Fig. 3B) and preferred the littoral habitat; no differences in their species richness with respect to the fish presence or mesohabitat were found (Table 2). Aquatic beetles were less abundant in fish-containing ponds than in fish-free ponds. Their abundance and species richness were higher in littoral habitat; some species, such as C. lateralimarginalis and G. austriacus occurred exclusively in littoral habitat (Tables 1 and 2; Fig. 3C). The interactions between fish status in the pond and mesohabitat were all insignificant (p > 0.2). Young stages and adults were pooled for analyses; however, it should be noted that of the total of 161 coleopteran larvae collected, only 7 individuals (4.3%) were found in the open water zone of fish-containing ponds.

Mean (± SE) relative density of a Hydrachnidia, b Heteroptera, c Coleoptera in open water (empty squares) and littoral (solid circles) habitats in fish-free and fish-containing ponds
The assemblages of all three macroinvertebrate groups were significantly nested (NODF range = 23.8–38.0, all Z ≥ 4.1, all p < 0.001); however, nestedness patterns differed among the taxonomic groups with respect to fish presence/absence in the ponds. The water mite assemblages recorded in fish-free ponds represented nested subsets of assemblages from ponds containing fish (t30 = 2.43, p = 0.021), while the reverse pattern was observed for aquatic beetles (t31 = 2.31, p = 0.028), and bug nestedness was independent of fish presence (t35 = 1.15, p = 0.258).
Discussion
Effects of fish on macroinvertebrate groups
The studied groups of macroinvertebrates showed differences in abundance and species composition with respect to the fish status of the ponds, and also in the use of within-pond mesohabitat. It should be noted that CCA and GLMM measured the habitat preferences of pond macroinvertebrates at different spatial levels; GLMMs were based on the total densities within mesohabitats, while CCA described the distribution of species with pond as the statistical unit. Thus, for example, species preferring ponds with extensive cover of emergent vegetation could potentially occur mainly in the open-water mesohabitat. The total densities and species richness of water mites were higher in ponds with fish, while the opposite was observed for aquatic beetles. The densities of aquatic bugs and beetles were distinctly higher in the emergent littoral vegetation than in open water, while water mites were equally distributed between mesohabitats. Given the lack of significant interactions between the effects of fish status and the effects of mesohabitat type, we conclude that the observed dependence of aquatic bugs and beetles on littoral vegetation was a result of their general habitat preferences and not a behavioural response to fish presence.
The higher densities and species richness of water mites in fish-containing ponds than in fishless ones is exceptional among macroinvertebrates; although certain species are adapted to minimize the risk of predation, most macroinvertebrate taxa are considered fish-sensitive (Bendell and McNicol 1995; Binckley and Resetarits 2005). Water mites are eaten by fish, but they are an infrequent and presumably occasionally accidental component of the fish diet (Eriksson et al. 1980; Crivelli 1981; Matveev and Martinez 1990; Kloskowski 2011b). Hence, consumption by fish is generally assumed to have no significant impact on water mite populations (Kerfoot 1982; Proctor et al. 2015; but see Leppä et al. 2003). Their relative invulnerability to fish is usually linked to low palatability. Water mites can be unpalatable to vertebrate predators, and when captured by fish they are usually rejected and often survive the attacks (Proctor and Garga 2004). The bright body colours of many water mite species are believed to function as an aposematic signal (Kerfoot 1982; but see Proctor and Garga 2004). Some species (e.g., P. pusilla and P. dispersa) are simply of small individual size, about 1 mm, and may be negatively selected by large carp (Sibbing 1988; Nieoczym and Kloskowski 2014; Gerecke et al. 2016). Likewise, it has been suggested that aquatic bugs are taken by fish only in small numbers (Scrimshaw and Kerfoot 1987). However, many studies have demonstrated a strong impact of fish on this taxonomic group (Macan 1965; Cook and Streams 1984; Jansson 1986; Bendell and McNicol 1995). Fish presence can enforce mesohabitat shifts by aquatic bugs such as corixids to in-shore vegetation because species inhabiting deeper parts of water (and farther from the shore) are most exposed to fish predation (Jansson 1986; Oscarson 1987). We found no differences in aquatic bug densities or species richness between fish-free and fish-containing ponds (see also Hassall et al. 2011); however, we did not analyse any subtler shifts in habitat use by individual species in the presence of fish. The two most common aquatic bugs, S. falleni and I. cimicoides, are believed to favour open-water habitats, provided that they are rich in submerged vegetation (Savage 1994; Peták et al. 2014; Olosutean et al. 2019). In our study, their abundance was independent of fish status in the pond and of mesohabitat type. Aquatic bugs show various antipredator defences, potentially allowing their co-occurrence with fish. Fast and erratic movements (Kovac and Maschwitz 1991) and the formation of large aggregations (Bailey 1987) by Notonectidae and Corixidae can be considered to be defensive adaptations against vertebrate predators (see also Cook and Streams 1984). Naucoridae, here represented by I. cimicoides, as well as Notonectidae, can inflict painful bites, while corixids eject defensive secretions when attacked (reviewed by Dettner 2019). Moreover, fish may facilitate water mite or aquatic bug abundance by suppressing their potential invertebrate predators (Eriksson et al. 1980; Punčochář and Hrbacek 1991; see also Batzer et al. 2000). In addition, bioturbation caused by benthivorous fish increases the availability of food (mainly chironomid larvae) for water mites and possibly also for other predatory macroinvertebrates (Paterson 1970; ten Winkel and Davids 1985).
No aquatic beetle species were found to be positively associated with fish presence, and total beetle densities were generally higher in littoral vegetation. As the use of littoral vegetation did not increase in the presence of fish, the higher abundances in littoral vegetation cannot be explained by shift to better cover. This was surprising, given that coleopteran densities and species richness were negatively related to fish presence, and that shifts to complex-structured (micro)habitats as a response to fish predation risk were documented in fish-vulnerable amphibians and in their large-sized insect predators in a parallel study (Kloskowski et al. 2020; see also Rennie and Jackson 2005). In addition, in the absence of the top predator, a more even distribution across shallow ponds should reduce competition among macroinvertebrates. The coleopterans collected in the present study were mainly small- and medium-sized species, whose movements are not limited by densely structured emergent plants. In the complex-structured littoral habitat, they may find protection against wave action and refuge from many predators (Hejný and Husák 1978; Peckarsky 1984). The phytophagous M. appendiculata (Kölsch and Kubiak 2011) is an example of a poor swimmer with little chance of survival following an encounter with a fish predator in open water, restricted to densely vegetated areas where fish are less numerous and their movements are hampered (Wirsing et al. 2010). We found almost no coleopteran larvae in the open water zone of fish-containing ponds, indicating that larvae, typically less agile than adults, could not persist in the presence of fish without protective cover provided by emergent vegetation. Large predatory invertebrates are typically more exposed to fish predation (Cook and Streams 1984; Diehl 1992; Tolonen et al. 2003). However, the relatively large-sized C. lateralimarginalis, considered to be a fast swimmer adapted to living in open water (Ribera and Nilsson 1995), was caught exclusively in a littoral vegetation habitat. It is possible that predatory beetles find more prey in the littoral zone (Kajgrova et al. 2021). Some of the phytophilous coleopterans occurring in the study ponds, including small crawling species, such as Haliplidae, or Noteridae, which burrow in the bottom substrate and in root mats (Yee and Kehl 2015), show a ‘natural’ propensity for vegetated habitat, not directly related to a need for protection from predators. In addition, littoral vegetation provides adult beetles with a reproductive habitat largely inaccessible to fish predators.
It is not clear to what degree the differential distribution of taxa among ponds was a result of oviposition habitat selection (Brodin et al. 2006; Šigutová et al. 2021) or of consumptive impact of fish on species vulnerable to predation. Carp is not a pursuit forager of fast-swimming prey, and the contribution of our focal macroinvertebrate groups to its diet is minor (Guziur 1976; Michel and Oberdorff 1995; García-Berthou 2001; Kloskowski 2011b), but this does not rule out strong behavioural effects (Werner and Peacor 2003; Kloskowski 2011a). Aquatic beetle species richness was distinctly lower in ponds containing fish, indicating that aquatic insects with highly mobile adult stages may avoid water bodies with fish when choosing oviposition sites (Resetarits 2001; Brodin et al. 2006; Trekels and Vanschoenwinkel 2017), even though the ponds contained abundant littoral vegetation (i.e., a habitat relatively safe from fish predators). On the other hand, some aquatic beetles and bugs do not exhibit habitat selection against fish presence (Åbjörnsson et al. 2002; Trekels and Vanschoenwinkel 2017). Fish trophic effects can be more complex in speciose communities in which juvenile and small-sized fishes can occupy vegetated areas to escape predation by larger piscivorous fish, with multi-level consequences (increased competition with and predation on invertebrates, intra-guild predation) for littoral invertebrates (Foster et al. 1988; Mittelbach 1988). The interpretation of mesohabitat preferences, especially in water mites, which did not exhibit a clear pattern, may be constrained by the use of 24 h activity traps, which provide a time-integrated estimate of macroinvertebrate densities, independent of potential diel changes in their distribution, but may conceal mesohabitat shifts, e.g., from day to night (cf. Hampton and Friedenberg 2002).
The assemblages of all studied macroinvertebrate taxa exhibited a nested pattern of species occurrence; fish-free ponds supported relatively high numbers of coleopteran taxa, including rare species, while fish-containing ponds harboured relatively high numbers of water mite species; in both taxa the other type of pond contained only subsets of the common species. The significant nestedness in species distribution suggests that ponds differing in fish status did not support different macroinvertebrate communities driven by replacement of species (Baselga 2012); aquatic beetle assemblages suffered a loss of species due to fish presence in the ponds, while water mites showed a directionally opposite relationship.
The role of abiotic conditions
According to CCA, abiotic features of ponds affected the distribution of water mites. On the other hand, most of the species dominant in the study ponds are known to inhabit small water bodies (Gerecke et al. 2016), and such species have a wide tolerance to environmental conditions (Smit and van der Hammen 2000). The only species significantly associated with high dissolved oxygen levels, P. pusilla, as well as P. imminuta, which was relatively common in the study ponds, occurs mainly in lakes (Gerecke et al. 2016), i.e., under relatively high-oxygen conditions. However, these species utilize a broad range of aquatic habitats and are fairly resistant to physicochemical disturbances (Smit and van der Hammen 2000). Similarly, most of the aquatic beetles and bugs observed in the study ponds are common, eurytopic species; they prefer shallow, eutrophic macrophyte-rich water bodies, but can populate a broad range of habitats (Eyre et al. 1986; Savage 1994; Nilsson and Holmen 1995; Papáček 2001; Peták et al. 2014). Corixids usually move between deep and inshore parts of water bodies following thermal gradients during the day (Popham 1964); however, thermal (and also physicochemical) gradients do not occur or are weak in shallow waterbodies due to minor depth differences and thorough mixing (Korinek et al. 1987). Although some differences in abiotic characteristics between ponds with and without fish or between mesohabitats were statistically significant, the ranges of individual parameters for each pond category were relatively narrow. Hence we assume that gradients of most of the abiotic conditions across mesohabitats were of secondary importance for macroinvertebrate distribution, except for a few specialized species. In addition, since some abiotic conditions were likely to be affected to a significant degree by carp activities, their effects per se were difficult to distinguish from the trophic or other impact of fish (Lougheed et al. 1998; Parkos III et al. 2003). For example, stirring of bottom sediments by benthivorous fish enhances total suspended solid concentrations, but an associated increase in prey availability is presumably more important for predatory macroinvertebrates (Paterson 1970; ten Winkel and Davids 1985; Kornijów et al. 2001; Hädicke et al. 2017). Higher conductivity in littoral vegetation than in the open water zone can result from more ample detritus deposits, which support favourable nutritional and environmental conditions for water mites and some aquatic bugs, such as the dominant S. falleni (Jansson 1986; Pieczyńska 1990; Savage 1994).
Conclusion
Predation is a critical determinant of the survival and reproductive success of many aquatic macroinvertebrates, and consequently predation risk is assumed to be largely responsible for their distribution among water bodies and mesohabitats (Nilsson 1984; Nicolet et al. 2004; Gioria 2014). Here, we found that aquatic beetle densities and species richness were lower in ponds with fish than in ponds without fish. However, our findings reveal that water mites and aquatic bugs, which play a significant role in the organization of aquatic food webs, especially as parasites or predators (Papáček 2001; Proctor et al. 2015), can persist in waters with high fish densities and possibly even benefit from fish presence (cf. Bendell and McNicol 1995; Carlisle and Hawkins 1998; Pope and Hannelly 2013), and that waters dominated by fish, such as managed fish ponds, include habitats compatible with macroinvertebrate conservation.
Our results stress the importance of littoral habitat for aquatic macroinvertebrates (see also Pierce and Hinrichs 1997; Burks et al. 2002), as aquatic beetles and bugs occurred at higher abundances in emergent littoral vegetation than in the open water area. Although fish were an important factor structuring coleopteran distribution among ponds, we found no evidence that mesohabitat preferences at the taxonomic group level were related to fish presence for any of the macroinvertebrate groups studied. A possible explanation is that some species may occur in emergent vegetation, at least during certain diel periods, for reasons other than predation avoidance (Hampton and Friedenberg 2002). In addition, the spatial distribution of many macroinvertebrates may reflect avoidance of predators other than fish. High densities of aquatic insects in littoral mesohabitat in the absence of fish were unexpected, as large-sized predaceous invertebrates, including cannibalistic congeners, which attack prey from ambush, pose a significant threat to smaller species/individuals (Bennett and Streams 1986; Åbjörnsson et al. 2002; Rennie and Jackson 2005) and benefit from cover provided by habitat structure (Tolonen et al. 2003; Klecka and Boukal 2014). On the other hand, when fish are absent, large-sized invertebrate predators such as large Dytiscidae and larval Anisoptera may shift out of the littoral habitat to feed in open water (Kloskowski et al. 2020). Hence, small-sized macroinvertebrates, unless morphologically or chemically protected from predation, may favour densely vegetated littoral habitat irrespective of which predators (invertebrates or fish) dominate the open water areas.
Data availability
The datasets generated and analysed during the current study are available from the corresponding author on reasonable request.
References
Åbjörnsson K, Brönmark C, Hansson L-A (2002) The relative importance of lethal and non-lethal effects of fish on insect colonisation of ponds. Freshw Biol 47:1489–1495. https://doi.org/10.1046/j.1365-2427.2002.00883.x
Almeida-Neto M, Guimarães P, Guimarães PR Jr, Loyola RD, Ulrich W (2008) A consistent metric for nestedness analysis in ecological systems: reconciling concept and measurement. Oikos 117:1227–1239. https://doi.org/10.1111/j.0030-1299.2008.16644.x
Bailey PC (1987) Abundance and age-specific spatial and temporal distribution in two waterbug species, Anisops deanei (Notonectidae) and Ranatra dispar (Nepidae) in three farm dams in South Australia. Oikos 49:83–90. https://doi.org/10.2307/3565557
Baselga A (2012) The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Global Ecol Biogeogr 21:1223–1232. https://doi.org/10.1111/j.1466-8238.2011.00756.x
Batzer DP, Wissinger SA (1996) Ecology of insect communities in nontidal wetlands. Annu Rev Entomol 41:75–100. https://doi.org/10.1146/annurev.en.41.010196.000451
Batzer DP, Pusateri CR, Vetter R (2000) Impacts of fish predation on marsh invertebrates: direct and indirect effects. Wetlands 20:307–312. https://doi.org/10.1672/0277-5212(2000)020[0307:IOFPOM]2.0.CO;2
Bendell BE, McNicol DK (1995) Lake acidity, fish predation, and the distribution and abundance of some littoral insects. Hydrobiologia 302:133–145. https://doi.org/10.1007/BF00027038
Bennett DV, Streams FA (1986) Effects of vegetation on Notonecta (Hemiptera) distribution in ponds with and without fish. Oikos 46:62–69. https://doi.org/10.2307/3565381
Binckley CA, Resetarits WJ (2005) Habitat selection determines abundance, richness and species composition of beetles in aquatic communities. Biol Lett 1:370–374. https://doi.org/10.1098/rsbl.2005.0310
Bouchard RW, Genet JA, Chirhart JW (2014) Does supplementing dipnet samples with activity traps improve the ability to assess the biological integrity of macroinvertebrate communities in depressional wetlands? Wetlands 34:699–711. https://doi.org/10.1007/s13157-014-0535-0
Brodin T, Johansson F, Bergsten J (2006) Predator related oviposition site selection of aquatic beetles (Hydroporus spp.) and effects on offspring life-history. Freshw Biol 51:1277–1285. https://doi.org/10.1111/j.1365-2427.2006.01563.x
Broyer J, Curtet L (2011) The influence of fish farming intensification on taxonomic richness and biomass density of macrophyte-dwelling invertebrates in french fishponds. Knowl Manag Aquat Ecosyst 400:1–12. https://doi.org/10.1051/kmae/2011017
Buczyńska E, Buczyński P, Lechowski L, Stryjecki R (2007) Fish pond complexes as refugia of aquatic invertebrates (Odonata, Coleoptera, Heteroptera, Trichoptera, Hydrachnidia): a case study of the pond complex in Zalesie Kańskie (Central-East Poland). Nat Conserv 64:39–55
Burks RL, Lodge D, Jeppesen E, Lauridsen T (2002) Diel horizontal migration of zooplankton: costs and benefits of inhabiting the littoral. Freshw Biol 47:343–365. https://doi.org/10.1046/j.1365-2427.2002.00824.x
Carlisle DM, Hawkins CP (1998) Relationships between invertebrate assemblage structure, 2 trout species, and habitat structure in Utah mountain lakes. J N Am Benthol Soc 17:286–300. https://doi.org/10.2307/1468332
Cook WL, Streams FA (1984) Fish predation on Notonecta (Hemiptera): relationship between prey risk and habitat utilization. Oecologia 64:177–183. https://doi.org/10.1007/BF00376868
Crivelli A (1981) The biology of the common carp, Cyprinus carpio L. in the Camargue, southern France. J Fish Biol 18:271–290. https://doi.org/10.1111/j.1095-8649.1981.tb03769.x
de Mendoza G, Rico E, Catalan J (2012) Predation by introduced fish constrains the thermal distribution of aquatic Coleoptera in mountain lakes. Freshw Biol 57:803–814. https://doi.org/10.1111/j.1365-2427.2012.02746.x
Dettner K (2019) Defenses of water insects. In: Del-Claro K, Guillermo R (eds) Aquatic insects. Springer, Cham., pp 191–262. https://doi.org/10.1007/978-3-030-16327-3_9
Di Sabatino A, Gerecke R, Gledhill T, Smit H (2010) Chelicerata: Acari II. In: Gerecke R (ed) Freshwater fauna of Central Europe, vol 7. Spektrum Akademischer Verlag, München
Diehl S (1992) Fish predation and benthic community structure: the role of omnivory and habitat complexity. Ecology 73:1646–1661. https://doi.org/10.2307/1940017
Diehl S, Kornijów R (1998) Influence of submerged macrophytes on trophic interactions among fish and macroinvertebrates. In: Jeppesen E, Søndergaard M, Søndergaard M, Christoffersen K (eds) The structuring role of submerged macrophytes in lakes. Springer, New York, pp 24–46
Eriksson MOG, Henrikson L, Nilsson BI, Nyman G, Oscarson H, Stenson AE (1980) Predator-prey relations important for the biotic changes in acidified lakes. Ambio 9:248–249
Evans RA (1989) Response of limnetic insect populations of two acidic, fishless lakes to liming and brook trout (Salvelinus fontinalis). Can J Fish Aquat Sci 46:342–351. https://doi.org/10.1139/f89-045
Eyre M, Ball S, Foster G (1986) An initial classification of the habitats of aquatic Coleoptera in north-east England. J Appl Ecol 23:841–852. https://doi.org/10.2307/2403938
Foster SA, Garcia VB, Town MY (1988) Cannibalism as the cause of an ontogenetic shift in habitat use by fry of the threespine stickleback. Oecologia 74:577–585. https://doi.org/10.1007/BF00380056
García-Berthou E (2001) Size-and depth-dependent variation in habitat and diet of the common carp (Cyprinus carpio). Aquat Sci 63:466–476. https://doi.org/10.1007/s00027-001-8045-6
Gerecke R, Gledhill T, Pešić V, Smit H (2016) Chelicerata: Acari III. In: Gerecke R (ed) Freshwater fauna of Central Europe, vol 7. Spektrum Akademischer Verlag, München
Gilinsky E (1984) The role of fish predation and spatial heterogeneity in determining benthic community structure. Ecology 65:455–468. https://doi.org/10.2307/1941408
Gioria M (2014) Habitats. In: Yee DA (ed) Ecology, systematics, and the natural history of predaceous diving beetles. Dytiscidae), Coleoptera, pp 307–362
Guziur J (1976) The feeding of two year old carp (Cyprinus·carpio L.) in a vendace lake Klawój. Ekol Pol 24:211–235
Hädicke WC, Rédei D, Kment P (2017) The diversity of feeding habits recorded for water boatmen (Heteroptera: Corixoidea) world-wide with implications for evaluating information on the diet of aquatic insects. Eur J Entomol 114:147–159. https://doi.org/10.14411/eje.2017.020
Hampton SE, Friedenberg NA (2002) Nocturnal increases in the use of near-surface water by pond animals. Hydrobiologia 477:171–179. https://doi.org/10.1023/A:1021059108674
Hanson MA, Riggs MR (1995) Potential effects of fish predation on wetland invertebrates: a comparison of wetlands with and without fathead minnows. Wetlands 15:167–175. https://doi.org/10.1007/BF03160670
Hassall C, Hollinshead J, Hull A (2011) Environmental correlates of plant and invertebrate species richness in ponds. Biodivers Conserv 20:3189–3222. https://doi.org/10.1007/s10531-011-0142-9
Healey M (1984) Fish predation on aquatic insects. In: Resh VH, Rosenberg DM (eds) The ecology of aquatic insects. Praeger, New York, pp 255–288
Hejný S, Husák Å (1978) Higher plant communities. In: Dykyjová D, Kvĕt J (eds) Pond littoral ecosystems. Ecological Studies, vol 28. Springer, Berlin, Heidelberg, pp 23–64
Hilsenhoff WL (1987) Effectiveness of bottle traps for collecting Dytiscidae (Coleoptera). Coleopt Bull 41:377–380
Hyvönen T, Nummi P (2000) Activity traps and the corer: complementary methods for sampling aquatic invertebrates. Hydrobiologia 432:121–125. https://doi.org/10.1023/A:1004038707992
Jäch MA, Balke M (2008) Global diversity of water beetles (Coleoptera) in freshwater. Hydrobiologia 595:419–442. https://doi.org/10.1007/s10750-007-9117-y
Jansson A (1986) The Corixidae (Heteroptera) of Europe and some adjacent regions. Acta Entomol Fenn 47:1–94
Kajgrova L, Adamek Z, Regenda J, Bauer C, Stejskal V, Pecha O, Hlavac D (2021) Macrozoobenthos assemblage patterns in european carp (Cyprinus carpio) ponds – the importance of emersed macrophyte beds. Knowl Manag Aquat Ecosyst 422:9. https://doi.org/10.1051/kmae/2021008
Kerfoot WC (1982) A question of taste: crypsis and warning coloration in freshwater zooplankton communities. Ecology 63:538–554. https://doi.org/10.2307/1938969
Klausnitzer B (1996) Käfer im und am Wasser. Die neue Brehm-Bücherei, 567. Westarp Wissenschaften, Spektrum Akademische Verlag, Magdeburg – Heidelberg – Berlin – Oxford
Klecka J, Boukal DS (2014) The effect of habitat structure on prey mortality depends on predator and prey microhabitat use. Oecologia 176:183–191. https://doi.org/10.1007/s00442-014-3007-6
Kloskowski J (2011a) Consequences of the size structure of fish populations for their effects on a generalist avian predator. Oecologia 166:517–530. https://doi.org/10.1007/s00442-010-1862-3
Kloskowski J (2011b) Differential effects of age-structured common carp (Cyprinus carpio) stocks on pond invertebrate communities: implications for recreational and wildlife use of farm ponds. Aquacult Int 19:1151–1164. https://doi.org/10.1007/s10499-011-9435-y
Kloskowski J, Nieoczym M, Stryjecki R (2020) Between-habitat distributions of pond tadpoles and their insect predators in response to fish presence. Hydrobiologia 847:1343–1356. https://doi.org/10.1007/s10750-020-04190-5
Kölsch G, Kubiak M (2011) The aquatic leaf beetle species Macroplea mutica and M. appendiculata (Coleoptera, Chrysomelidae, Donaciinae) differ in their use of Myriophyllum spicatum as a host plant. Aquat Insect 33:13–26. https://doi.org/10.1080/01650424.2011.572558
Korinek V, Fott J, Fuksa J, Lellák J, Prazakova M (1987) Carp ponds of Central Europe. In: Michael RG (ed) Managed aquatic ecosystems of the world, 29 edn. Elsevier Science Publishing Co, Amsterdam, pp 29–62
Kornijów R, Stryjecki R, Kuoppamäki K, Horppila J, Luokkanen E, Kairesalo T (2001) Do water mites benefit from benthivorous fish in vegetated lake areas? Verh Internat Verein Limnol 27:3291–3294. https://doi.org/10.1080/03680770.1998.11902434
Kottelat M, Freyhof J (2007) Handbook of european freshwater fishes. Publications Kottelat
Kovac D, Maschwitz U (1991) The function of the metathoracic scent gland in corixid bugs (Hemiptera, Corixidae): secretion-grooming on the water surface. J Nat Hist 25:331–340. https://doi.org/10.1080/00222939100770241
Kurek J, Cwynar LC, Weeber RC, Jeffries DS, Smol JP (2010) Ecological distributions of Chaoborus species in small, shallow lakes from the Canadian Boreal Shield ecozone. Hydrobiologia 652:207–221. https://doi.org/10.1007/s10750-010-0333-5
Leppä M, Hämäläinen H, Karjalainen J (2003) The response of benthic macroinvertebrates to whole-lake biomanipulation. Hydrobiologia 498:97–105. https://doi.org/10.1023/A:1026224923481
Liao W, Venn S, Niemelä J (2020) Environmental determinants of diving beetle assemblages (Coleoptera: Dytiscidae) in an urban landscape. Biodivers Conserv 29:2343–2359. https://doi.org/10.1007/s10531-020-01977-9
Lima SL, Dill LM (1990) Behavioral decisions made under the risk of predation: a review and prospectus. Can J Zool 68:619–640. https://doi.org/10.1139/z90-092
Lougheed VL, Crosbie B, Chow-Fraser P (1998) Predictions on the effect of common carp (Cyprinus carpio) exclusion on water quality, zooplankton, and submergent macrophytes in a great lakes wetland. Can J Fish Aquat Sci 55:1189–1197. https://doi.org/10.1139/f97-315
Lürig MD, Best RJ, Dakos V, Matthews B (2021) Submerged macrophytes affect the temporal variability of aquatic ecosystems. Freshw Biol 66:421–435. https://doi.org/10.1111/fwb.13648
Lytle DA (2015) Order Hemiptera. In: Thorp JH, Rogers DC (eds) Ecology and general biology: Thorp and Covich’s freshwater invertebrates, 4th ed., Academic Press, pp. 951–963. https://doi.org/10.1016/B978-0-12-385026-3.00037-1
Macan T (1965) Predation as a factor in the ecology of water bugs. J Anim Ecol 34:691–698. https://doi.org/10.2307/2456
Matveev V, Martinez C (1990) Can water mites control populations of planktonic Cladocera? Hydrobiologia 198:227–231. https://doi.org/10.1007/BF00048637
Michel P, Oberdorff T (1995) Feeding habits of fourteen european freshwater fish species. Cybium 19:5–46
Mittelbach GG (1988) Competition among refuging sunfishes and effects of fish density on littoral zone invertebrates. Ecology 69:614–623. https://doi.org/10.2307/1941010
Morin PJ (1984) Odonate guild composition: experiments with colonization history and fish predation. Ecology 65:1866–1873. https://doi.org/10.2307/1937784
Murkin HR, Abbott PG, Kadlec JA (1983) A comparison of activity traps and sweep nets for sampling nektonic invertebrates in wetlands. Freshw Invertebr Biol 2:99–106. https://doi.org/10.2307/1467114
Nicolet P, Biggs J, Fox G, Hodson MJ, Reynolds C, Whitfield M, Williams P (2004) The wetland plant and macroinvertebrate assemblages of temporary ponds in England and Wales. Biol Conserv 120:261–278. https://doi.org/10.1016/j.biocon.2004.03.010
Nieoczym M, Kloskowski J (2014) The role of body size in the impact of common carp Cyprinus carpio on water quality, zooplankton, and macrobenthos in ponds. Int Rev Hydrobiol 99:212–221. https://doi.org/10.1002/iroh.201301644
Nieoczym M, Kloskowski J (2015) Responses of epibenthic and nektonic macroinvertebrate communities to a gradient of fish size in ponds. J Limnol 74:50–62. https://doi.org/10.4081/jlimnol.2014.981
Nieoczym M, Mencfel R, Gorzel M, Kloskowski J (2020) Reduced abundance but increased diversity of chironomid larvae under higher trophic pressure from fish in semi-permanent ponds. Limnologica 82:125778. https://doi.org/10.1016/j.limno.2020.125778
Nilsson AN (1984) Species richness and succession of aquatic beetles in some kettle-hole ponds in northern Sweden. Ecography 7:149–156. https://doi.org/10.1111/j.1600-0587.1984.tb01115.x
Nilsson AN, Holmen M (1995) The aquatic Adephaga (Coleoptera) of the Fennoscandia and Denmark. II. Dytiscidae. Fauna Ent Scand 32, EJ Brill/Scandinavian Science Press Ltd: Leiden & Copenhagen
Oscarson HG (1987) Habitat segregation in a water boatman (Corixidae) assemblage: the role of predation. Oikos 133–140. https://doi.org/10.2307/3566018
Olosutean H, Bungiac S, Perju M (2019) Are aquatic and semiaquatic true bugs (Hemiptera: Nepomorpha; Gerromorpha) distinct aquatic communities? A case study in homogeneous habitats. Inland Waters 9:513–521. https://doi.org/10.1080/20442041.2019.1600937
Papáček M (2001) Small aquatic and ripicolous bugs (Heteroptera: Nepomorpha) as predators and prey: the question of economic importance. Eur J Entomol 98:1–12. https://doi.org/10.14411/eje.2001.001
Parkos IIIJJ, Santucci VJ Jr, Wahl DH (2003) Effects of adult common carp (Cyprinus carpio) on multiple trophic levels in shallow mesocosms. Can J Fish Aquat Sci 60:182–192. https://doi.org/10.1139/f03-011
Paterson CG (1970) Water mites (Hydracarina) as predators of chironomid larvae (Insectar: Diptera). Can J Zool 48:610–614. https://doi.org/10.1139/z70-112
Peckarsky BL (1982) Aquatic insect predator-prey relations. Bioscience 32:261–266. https://doi.org/10.2307/1308532
Peckarsky BL (1984) Predator-prey interactions among aquatic insects. In: Resh VH, Rosenberg DM (eds) The ecology of aquatic insects. Praeger, New York, pp 196–254
Peták E, Erős T, Bakonyi G (2014) Habitat use and movement activity of two common predatory water bug species, Nepa cinerea L., 1758 and Ilyocoris cimicoides (L., 1758) (Hemiptera: Nepomorpha): field and laboratory observations. Aquat Insect 36:231–243. https://doi.org/10.1080/01650424.2015.1079638
Pieczyńska E (1990) Littoral habitats and communities. In: Jørgensen SE, Löffler H (eds) Guidelines of Lake Management 3, Lake Shore Management, pp 39–71
Pieczyński E (1976) Ecology of water mites (Hydracarina) in lakes. Pol Ecol Stud 2:5–54
Pierce C, Hinrichs B (1997) Response of littoral invertebrates to reduction of fish density: simultaneous experiments in ponds with different fish assemblages. Freshw Biol 37:397–408. https://doi.org/10.1046/j.1365-2427.1997.00169.x
Pope KL, Hannelly EC (2013) Response of benthic macroinvertebrates to whole-lake, non-native fish treatments in mid-elevation lakes of the Trinity Alps, California. Hydrobiologia 714:201–215. https://doi.org/10.1007/s10750-013-1537-2
Popham E (1964) The migration of aquatic bugs with special reference to the Corixidae (Hemiptera Heteroptera). Arch Hydrobiol 60:450–496
Proctor HC, Garga N (2004) Red, distasteful water mites: did fish make them that way? Exp Appl Acarol 34:127–147. https://doi.org/10.1023/B:APPA.0000044444.81413.1a
Proctor HC, Smith IM, Cook DR, Smith BP (2015) Subphylum Chelicerata, Class Arachnida. In: Thorp JH, Rogers DC (eds) Ecology and general biology: Thorp and Covich’s freshwater invertebrates, 4th edn. Academic Press, pp 599–660. https://doi.org/10.1016/B978-0-12-385026-3.00025-5
Punčochář P, Hrbacek J (1991) Water mites in the plankton of Hubenov Reservoir and their relations to fish stock composition. In: Dusbábek F, Bukva V (eds) Modern Acarology, vol 1. Academia Prague and SPB Academic Publishing, The Hague, pp 449–457
Rennie MD, Jackson LJ (2005) The influence of habitat complexity on littoral invertebrate distributions: patterns differ in shallow prairie lakes with and without fish. Can J Fish Aquat Sci 62:2088–2099. https://doi.org/10.1139/f05-123
Resetarits WJ (2001) Colonization under threat of predation: avoidance of fish by an aquatic beetle, Tropisternus lateralis (Coleoptera: Hydrophilidae). Oecologia 129:155–160. https://doi.org/10.1007/s004420100704
Ribera I, Nilsson A (1995) Morphometric patterns among diving beetles (Coleoptera: Noteridae, Hygrobiidae, and Dytiscidae). Can J Zool 73:2343–2360. https://doi.org/10.1139/z95-275
Savage A (1994) The distribution of Corixidae in relation to the water quality of british lakes: a monitoring model. Freshw Forum 4:32–61
Schilling EG, Loftin CS, Degoosh KE, Huryn AD, Webster KE (2008) Predicting the locations of naturally fishless lakes. Freshw Biol 53:1021–1035. https://doi.org/10.1111/j.1365-2427.2007.01949.x
Schilling EG, Loftin CS, Huryn AD (2009) Macroinvertebrates as indicators of fish absence in naturally fishless lakes. Freshw Biol 54:181–202. https://doi.org/10.1111/j.1365-2427.2008.02096.x
Scrimshaw S, Kerfoot WC (1987) Chemical defenses of freshwater organisms: beetles and bugs. In: Kerfoot WC, Sih A (eds) Predation: direct and indirect impacts on aquatic communities. University Press of New England, Hanover, New Hampshire, pp 240–262
Sibbing FA (1988) Specializations and limitations in the utilization of food resources by the carp, Cyprinus carpio: a study of oral food processing. Environ Biol Fish 22:161–178. https://doi.org/10.1007/BF00005379
Šigutová H, Harabiš F, Šigut M, Vojar J, Choleva L, Dolný A (2021) Specialization directs habitat selection responses to a top predator in semiaquatic but not aquatic taxa. Sci Rep 11:18928. https://doi.org/10.1038/s41598-021-98632-2
Smit H, van der Hammen H (2000) Atlas van de nederlandse watermijten (Acari: Hydrachnidia). Ned Faun Med 13:1–272
Specziár A, Tölg L, Bíró P (1997) Feeding strategy and growth of cyprinids in the littoral zone of Lake Balaton. J Fish Biol 51:1109–1124. https://doi.org/10.1111/j.1095-8649.1997.tb01130
Strona G, Galli P, Seveso D, Montano S, Fattorini S (2014) Nestedness for dummies (NeD): a user-friendly web interface for exploratory nestedness analysis. J Stat Softw 59:1–9. https://doi.org/10.18637/jss.v059.c03
Stryjecki R (2007) The role of fish ponds as important habitats for water mites (Hydrachnidia, Acari). Oceanol Hydrobiol Stud 36. Sup. 4:73–79
ten Winkel EH, Davids C (1985) Bioturbation by cyprinid fish affecting the food availability for predatory water mites. Oecologia 67:218–219. https://doi.org/10.1007/BF00384287
ter Braak CJ Šmilauer P (2018) Canoco reference manual and user’s guide: Software for ordination (version 5.10). Biometris. Wageningen University & Research
Tolonen K, Hämäläinen H, Holopainen I, Mikkonen K, Karjalainen J (2003) Body size and substrate association of littoral insects in relation to vegetation structure. Hydrobiologia 499:179–190. https://doi.org/10.1023/A:1026325432000
Toro M, Granados I, Rubio Á, Camacho A, Sabás I, Ventura M, Montes C (2020) Response of the aquatic invertebrate community to the eradication of an exotic invasive fish 30 years after its introduction into an Iberian alpine lake. Aquat Sci 82:55. https://doi.org/10.1007/s00027-020-00728-w
Trekels H, Vanschoenwinkel B (2017) When fear kicks in: predator cues initially do not but eventually do affect insect distribution patterns in a new artificial pond cluster. Hydrobiologia 790:157–166. https://doi.org/10.1007/s10750-016-3027-9
Yee DA, Kehl S (2015) Order Coleoptera. In: Thorp JH, Rogers DC (eds) Ecology and general biology: Thorp and Covich’s freshwater invertebrates, 4th edn. Academic Press, pp 1003–1042. https://doi.org/10.1016/B978-0-12-385026-3.00039-5
Werner EE, Peacor SD (2003) A review of trait-mediated indirect interactions in ecological communities. Ecology 84:1083–1100. https://doi.org/10.1890/0012-9658(2003)084[1083:AROTII]2.0.CO;2
Winfield I (1986) The influence of simulated aquatic macrophytes on the zooplankton consumption rate of juvenile roach, Rutilus rutilus, rudd, Scardinius erythrophthalmus, and perch, Perca fluviatilis. J Fish Biol 29:37–48. https://doi.org/10.1111/j.1095-8649.1986.tb04997.x
Wirsing AJ, Cameron KE, Heithaus MR (2010) Spatial responses to predators vary with prey escape mode. Anim Behav 79:531–537. https://doi.org/10.1016/j.anbehav.2009.12.014
Zimmer KD, Hanson MA, Butler MG (2000) Factors influencing invertebrate communities in prairie wetlands: a multivariate approach. Can J Fish Aquat Sci 57:76–85. https://doi.org/10.1139/f99-180
Zimmer KD, Hanson MA, Butler MG, Duffy WG (2001) Size distribution of aquatic invertebrates in two prairie wetlands, with and without fish, with implications for community production. Freshw Biol 46:1373–1386. https://doi.org/10.1046/j.1365-2427.2001.00759.x
Acknowledgements
We would like to thank the pond managers for their cooperation with this study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Authors declare no conflict of interest.
Ethical approval
This study was conducted under permits DOP-oz. 6401.02.23.2012.JRO.2 and WPN.6401.42.2013.MO from the Regional Directorate for Environmental Protection in Lublin, Poland.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nieoczym, M., Stryjecki, R., Buczyński, P. et al. Differential abundance, composition and mesohabitat use by aquatic macroinvertebrate taxa in ponds with and without fish. Aquat Sci 85, 25 (2023). https://doi.org/10.1007/s00027-022-00922-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-022-00922-y