
Abstract
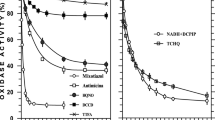
In a previous work (Kömen et al. 1991) it has been concluded that membrane fragments isolated from autotrophically grown Alcaligenes eutrophus H16 contain several iron-sulphur centres along with haems of a-, b-, c-, and d-type. These redox components have been proposed to be part of a branched respiratory chain leading to multiple membrane bound oxidases. Here, some of the respiratory activities catalyzed by membrane fragments from wild type cells of A. eutrophus (H16) and, for comparison, Paracoccus denitrificans, have been investigated through the use of electron transport inhibitors. Cyanide (CN-) titration curves indicated that in A. eutrophus H16 oxidation of succinate and H2 preferentially proceeds via the cytochrome c oxidase(s) branch (I 50=2 · 10-5 M) whereas the NADH dependent respiration started being inhibited at higher CN- concentrations (I 50=5 · 10-4 M). In membranes isolated from both, cells harvested at late growth-phase (OD 12) and from a mutant deficient in cytochrome c oxidase activity (A. eutrophus RK1), respiration was insensitive to low CN- concentrations (< 10-4 M), and it was sustained by the high catalytic activities of two quinol oxidases. These alternative oxidases of b- (formally o-) and d-type showed different sensitivities to KCN (I 50=10-3 M and 10-2 M, respectively). Interestingly, the cytochrome c oxidase(s) dependent respiration of H16 membranes was insensitive to antimycin A but largely inhibited by myxothiazol (10-6 M). This, and previous work (Kömen et al. 1991), suggest that although the respiratory chain of A. eutrophus is endowed with a putative bc 1 complex, its biochemical nature and role in respiration of this organism are apparently different from those of P. denitrificans. The peculiarity of the respiratory chain of A. eutrophus is confirmed by the rotenone insensitivity of the NADH oxidation in both protoplasts and membrane fragments from wild type and soluble hydrogenase deficient cells (HF14 and HF160). A tentative model of the respiratory chain of autotrophically grown A. eutrophus is presented.
Similar content being viewed by others
Abbreviations
- CTAB:
-
Cetyltrimethylammonium bromide
- cytochrome c-551:
-
number: wavelength in nm; cytochrome c 270, number: mid-point potential in mV
- DCIP:
-
2,6-dichlorophenol-indophenol
- DMSO:
-
dimethyl sulfoxide
- E m, 7 :
-
mid-point potential of an oxidation-reduction couple at pH 7.0
- I 50 :
-
concentration of the inhibitor at which 50% inhibition of the activity is observed
- KP-buffer:
-
potassium phosphate-buffer
- OD:
-
optical density at 436 nm, 1 cm light path
- PMS:
-
phenazine methosulfate
- Q:
-
ubiquinone
- Qi :
-
Q-reducing side of the bc 1 complex
- Qo :
-
Q-oxidizing side of the bc 1 complex
- TMPD:
-
N,N,N′,N′-tetramethyl-p-phenylenediamine
References
Ackrell BAC, Kearney EB, Singer TP (1978) Mammalian succinate dehydrogenase. Methods Enzymol 53:466–483
Beatrice MC, Chappell JB (1979) The respiratory chain of Alcaligenes eutrophus H16. Biochem J 178:15–22
Dooijewaard G, Slater EC (1976a) Steady-state kinetics of high molecular weight (type-I) NADH dehydrogenase. Biochim Biophys Acta 440:1–15
Dooijewaard G, Slater EC (1976b) Steady-state kinetics of low molecular weight (type-II) NADH dehydrogenase. Biochim Biophys Acta 440:16–35
Dry JB, Moore AL, Day DA, Wiskich JT (1989) Regulation of alternative pathway activity in plant mitochondria: Nonlinear relationship between electron flux and the redox poise of the quinone pool. Arch Biochem Biophys 273:148–157
Harmon HJ, Hall JD, Crane FL (1974) Structure of mitochondrial christae membranes. Biochim Biophys Acta 344:119–155
Hata-Tanaka A, Matsuura K, Itoh S, Anraku Y (1987) Electron flow and heme-heme interaction between cytochromes b-558, b-595 and d in a terminal oxidase in Escherichia coli. Biochim Biophys Acta 893:289–295
Jagow G von, Ljungdahl PO, Graf P, Onishi T, Trumpower BL (1984) An inhibitor of mitochondrial respiration which binds to cytochrome b and displaces quinone from the iron-sulfur protein of the cytochrome bc 1 complex. J Biol Chem 259: 6318–6326
Keilin D (1966) The history of cell respiration and cytochrome. Cambridge University Press, Cambridge
King MT, Drews G (1973) The function and localization of ubiquinone in the NADH-succinate-oxidase system of Rhodopseudomonas palustris. Biochim Biophys Acta 305:230–248
Kömen R, Zannoni D, Ingledew WJ, Schmidt K (1991) The electron transport system of Alcaligenes eutrophus H16. I. Spectroscopic and thermodynamic properties. Arch Microbiol 155:382–390
Lorence RM, Koland JG, Gennis RB (1986) Coulometric and spectroscopic analysis of the purified cytochrome d complex in Escherichia coli. Evidence for the identification of “cytochrome a 1” as cytochrome b-595. Biochemistry 25:2314–2321
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with Folin phenol reagent. J Biol Chem 193:265–275
Marrs B, Gest H (1973) Genetic mutations affecting the respiratory electron transport system of the photosynthetic bacterium Rhodopseudomonas capsulata. J Bacteriol 114:1045–1051
Meijer EM, Wever R, Stouthamer AH (1977) The role of iron-sulfur center 2 in electron transport and energy conservation in the NADH-ubiquinone segment of the respiratory chain in Paracoccus denitrificans. Eur J Biochem 81:267–275
Meinhardt SW, Yang X, Trumpower BL, Onishi T (1987) Identification of a stable ubisemiquinone and characterization of the effects of ubiquinone oxidation-reduction status on the Rieske iron-sulfur protein in the three-subunit ubiquinol-cytochrome c oxido-reductase of Paracoccus denitrificans. J Biol Chem 262:8702–8706
Miller MJ (1985) The cytochrome d complex is a coupling site in the aerobic respiration chain of Escherichia coli. J Biol Chem 260:14003–14008
Mitchell P (1975) Proton motive redox mechanism of the b/c complex in the respiratory chain: proton motive ubiquinone cycle. FEBS Lett 56:1–6
Moore AL, Dry IB, Wiskitch JT (1988) Measurement of the redox state of the ubiquinone pool in plant mitochondria. FEBS Lett 235:76–80
Onishi T (1973) Mechanism of electron transport and energy conservation in the site I region of the respiratory chain. Biochim Biophys Acta 301:105–128
Onishi T, Meinhardt SW, Matsushita K (1987) Studies of bacterial NADH-ubiquinone (or menaquinone) oxidoreductase systems. In: Papa S, Chance B, Ernster L (eds) Cytochrome systems. Molecular biology and bioenergetics. Plenum Press, New York, pp 443–450
Poole RK, Williams HD (1987) Proposal that the function of the membrane-bound a 1-like haemoprotein (cytochrome b 595) in Escherichia coli is a direct electron donation to cytochrome d. FEBS Lett 217:49–52
Porte F, Vignais PM (1980) Electron transport chain and energy transduction in Paracoccus denitrificans under autotrophic growth conditions. Arch Microbiol 127:1–10
Puustinen A, Finel M, Virkki M, Wikström M (1989) Cytochrome o (bo) is a proton pump in Paracoccus denitrificans and Escherichia coli. FEBS Lett 249:163–167
Rich P (1984) Electron and proton transfer through quinones and cytochrome bc 1 complexes. Biochim Biophys Acta 768:53–79
Salerno JC, Bolgiano B, Poole RK, Gennis RB, Ingledew WJ (1990) Heme-copper and heme-heme interactions in the cytochrome bo-containing quinol oxidase of Escherichia coli. J Biol Chem 265:4364–4368
Schink B, Schlegel HG (1978) Hydrogen metabolism in aerobic hydrogen oxidizing bacteria. Biochimie 60:279–305
Schneider K, Schlegel HG (1976) Purification and properties of soluble hydrogenase from Alcaligenes eutrophus H16. Biochim Biophys Acta 452:66–80
Simon R, Priefer U, Pühler A (1983a) A broad host range mobilization system for in vivo genetic engineering: transposon mutagenesis in gram-negative bacteria. Biotechnology 1:784–791
Simon R, Priefer U, Pühler A (1983b) Vector plasmids for in vivo manipulation of gram-negative bacteria. In: Pühler A (ed) Molecular genetics of the bacteria-plant interaction. Springer, Berlin Heidelberg New York, pp 98–106
Stouthamer AH (1980) Bioenergetic studies on Paracoccus denitrificans. Trends Biochem Sci 5:164–166
Venturoli G, Fernández-Velasco JG, Crofts AR, Melandri BA (1987) Demonstration of a collisional interaction of ubiquinol with the ubiquinol-cytochrome c 2 oxidoreductase complex in chromatophores from Rhodobacter sphaeroides. Biochim Biophys Acta 851:340–352
Williams HD, Poole RK (1987) The cytochromes of Acetobacter pasteurianus NCIB 6428. Evidence of a role for a cytochrome a 1-like haemoprotein in electron transfer to cytochrome oxidase a. J Gen Microbiol 133:2461–2472
Zannoni D (1985) Mefloquine: an antimalarial drug interacting with the b/c region of bacterial respiratory chains. FEBS Lett 183:340–344
Zannoni D (1987) The interplay between photosynthesis and respiration in facultative anoxygenic phototrophic bacteria. In: Papa S, Chance B, Ernster L (eds) Cytochrome systems. Molecular biology and bioenergetics. Plenum Press, New York, pp 575–583
Zannoni D (1989) The respiratory chains of pathogenic pseudomonads. Biochim Biophys Acta 975:299–316
Zannoni D, Moore AL (1990) Measurements of the redox state of the ubiquinone pool in Rhodobacter capsulatus membrane fragments. FEBS Lett 271:123–127
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Kömen, R., Zannoni, D. & Schmidt, K. The electron transport system of Alcaligenes eutrophus H16. Arch. Microbiol. 155, 436–443 (1991). https://doi.org/10.1007/BF00244958
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00244958