
Abstract
Background
Mountain pine beetle (MPB) is a native disturbance agent across most pine forests in the western US. Climate changes will directly and indirectly impact frequencies and severities of MPB outbreaks, which can then alter fuel characteristics and wildland fire dynamics via changes in stand structure and composition. To investigate the importance of MPB to past and future landscape dynamics, we used the mechanistic, spatially explicit ecosystem process model FireBGCv2 to quantify interactions among climate, MPB, wildfire, fire suppression, and fuel management under historical and projected future climates for three western US landscapes. We compared simulated FireBGCv2 output from three MPB modules (none, simple empirical, and complex mechanistic) using three focus variables and six exploratory variables to evaluate the importance of MPB to landscape dynamics.
Results
We found that inclusion of MPB (empirical or mechanistic) in the simulations significantly changed past and future landscape dynamics and that the mechanistic MPB module had more cross-scale interactions that increased variability, and perhaps realism, of simulation results. We also evaluated impacts of fire and fuel management on MPB dynamics and found that fire suppression influenced fuel loadings more than MPB disturbance, but at a landscape scale, most fuel treatment programs did little to change fuel loadings, MPB dynamics, and burned area, except under high fire suppression.
Conclusions
Synergistic interactions of climate, MPB, and wildfire catalyzed landscape-scale changes in vegetation distributions, fuels, and fire regimes in FireBGCv2 simulations. Models that simulate climate change on pine-dominated landscapes may be improved by including mechanistic MPB simulations to account for potentially important ecological interactions.
Resumen
Antecedentes
El escarabajo del pino de montaña (MPB) es un agente de disturbio nativo que se extiende a través de la mayoría de los bosques de pino del oeste de los EEUU. El cambio climático impacta directa o indirectamente en la frecuencia e intensidad de los estallidos del MPB, lo que puede alterar las características del combustible y la dinámica de los incendios a través de cambios en la estructura y composición de los rodales. Para investigar la importancia de MPB en el pasado y futuro de la dinámica del paisaje, usamos el modelo mecanístico explícito de procesos ecológicos titulado FireBGCv2, para cuantificar las interacciones entre el clima, MPB, incendios, supresión de fuegos y manejo del combustible bajo climas pasados y proyectados a futuro, para tres paisajes del oeste de los EEUU. Comparamos los resultados simulados obtenidos con el FireBGCv2 en tres módulos de PMB (ninguno, empírico simple y mecanístico completo) usando tres variables focales y seis variables exploratorias para evaluar la importancia de MPB en la dinámica del paisaje.
Resultados
Encontramos que la inclusión de MPB (tanto empírico como mecanístico) en las simulaciones cambiaban significativamente la dinámica pasada y futura de los paisajes y que el módulo mecanístico del MPB tenía más interacciones entre escalas que incrementaban la variabilidad, y tal vez el realismo, de los resultados de la simulación. También evaluamos los impactos del fuego y el manejo de combustibles en la dinámica del MPB y encontramos que la supresión influyó más en la carga de combustible que el disturbio causado por el MPB, aunque, a escala de paisaje, la mayoría de los tratamientos de combustible hicieron poco para cambiar las cargas de combustible, la dinámica del PMB, y área quemada, excepto bajo una fuerte supresión del fuego.
Conclusiones
Las acciones sinérgicas del clima, MPB e incendios catalizaron cambios a escala de paisaje sobre la distribución de la vegetación, los combustibles, y los regímenes de fuego en las simulaciones de FireBGCv2. Los modelos que simulan el cambio climático en paisajes dominados por pinos deben incluir simulaciones mecanísticas para contabilizar interacciones ecológicas potencialmente importantes.
Similar content being viewed by others

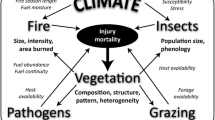

Introduction
Mountain pine beetle (Dendroctonus ponderosae Hopkins) (MPB) and wildland fire are two of the most significant natural disturbances in pine-dominated forests of the western US (Jenkins et al. 2008). As interacting and co-occurring disturbances, MPB outbreaks and wildland fire can cause highly visible, rapidly occurring, and persistent changes in landscape composition and structure (Loehman et al. 2017). For example, bark beetle-caused tree mortality can affect the amount and configuration of surface and canopy fuels, with the potential for increased wildfire intensity and severity and subsequent, fire-induced changes in the availability of host trees for MPB (Meigs et al. 2015; Page et al. 2013). Beetle-caused tree mortality may also affect the amount of light, water, and nutrients available for overstory and understory plants, including exotics (Runyon et al. 2020), thereby altering species composition and stand structure (i.e., fuel characteristics) for years after an outbreak (Audley et al. 2020; Hansen 2014).
Wildland fire and MPB disturbances are responsive to patterns of climate and vegetation (Kane et al. 2017; Scheller et al. 2018). Climate, as a top-down driver, influences frequency and timing of MPB and fire disturbance (Bentz et al. 2010; Heyerdahl et al. 2001; Swetnam and Betancourt 1998), while vegetation and fuel characteristics act as bottom-up drivers, influencing MPB and fire severity and extent (Archibald et al. 2013; Fettig and Hilszczański 2015; Whitlock et al. 2008).
Anthropogenic climate changes—in particular drought and increasing temperatures—have been recognized as key drivers of increased wildfire (Littell et al. 2009) and bark beetle activities (Berner et al. 2017; Fettig et al. 2019). Warming temperatures may facilitate MPB population growth in some forests (Bentz et al. 2016, 2019), and projected moisture deficit may increase drought stress, increasing vulnerability of host trees to MPB attack (Chapman et al. 2012; Kolb et al. 2016). Warming temperatures may also increase wildland fire frequency (Abatzoglou and Williams 2016; Westerling et al. 2006), size (Running 2006; Shive et al. 2014), intensity (Flannigan et al. 2009), and sometimes, severity (Miller et al. 2009). Predictions of warmer temperatures and increased drought suggest that the total area affected by large fires may increase in the coming decades (Williams, 2013). Acting independently or synchronously in space and time, wildland fires and MPB outbreaks can significantly influence forest structure, composition, and function; abruptly reorganize landscapes; and alter biogeochemical processes such as carbon cycling, water supply, and nutrient cycles (Edburg et al. 2012; Fettig et al. 2019; Kurz et al. 2008). Influences of MPB on wildfire occur in the context of other drivers, including topography, forest type, and disturbance history (Hicke et al. 2012). The complexity of these interactions among drivers and responses increases substantially when climate change and land management actions affect these processes, especially in this modern era of fire exclusion when approximately 96% of fires are suppressed each year (Loehman et al. 2017).
The potential for increased wildland fire following MPB outbreaks has been observed since the early twentieth century (Evenden and Gibson 1940; Weaver 1943). Few studies, however, have addressed long-term influences of wildland fires on MPB disturbance regimes, and fewer still have examined reciprocal interactions of MPB and fire through several disturbance cycles (but see Parker et al. (2006)). Flammability and crown fire potential is highest in the first 3 years after MPB attack, when needles of killed pine trees have lower foliar moisture than green trees (Stalling et al. 2017) and are still attached to branches, maintaining aerial fuel continuity (Jenkins et al. 2014). Once standing dead trees have lost their needles and small branches (4 to 10 years after attack), active crown fire potential is much lower than in non-attacked stands, but increased fine surface fuel loads may result in higher surface fire rates of spread, flame lengths, and torching potential (Jenkins et al. 2014; Schoennagel et al. 2012). The highest fire hazard is assumed to occur decades after attack as a result of accumulation of heavy, large-diameter fuels (snags and large branches), dense regeneration, and increased wind speeds and drying of fuels resulting from the loss of pine canopy (Hansen et al. 2015; Jenkins et al. 2008).
Reciprocally, fire effects on MPB population activity are also time-dependent. Although fire-weakened trees are most susceptible to attack in the months after severe fire injury (Davis et al. 2012; Lerch et al. 2016; Parker et al. 2006; Powell et al. 2012), wildland fires can affect beetle activity indirectly over longer time periods by altering forest composition and structure (e.g., removing fire intolerant species) and providing increased water, light, and nutrients for surviving trees (i.e., growth release), thus influencing availability and vigor of suitable host trees (Fettig et al. 2007; Hessburg et al. 2005; Keeling and Sala 2012). In addition, occurrence of stand-replacing fires can reduce the likelihood of MPB attack until regenerating forests have attained a threshold diameter sufficient to attract beetles, especially when beetle populations are at relatively low levels (Kulakowski et al. 2003). Jentsch et al. (2021) found that fire helps dampen MPB outbreaks not only by removing host trees but also by altering the demographic structure of forest stands.
Although MPB was rarely considered a major contributor to post-fire pine mortality historically (Geiszler et al. 1980; McHugh and Kolb 2003; Six and Skov 2009), recent findings indicate that MPB may indeed be attracted to multiple fire-injured pine species and that the likelihood of mass attack and subsequent tree mortality was positively associated with several measures of fire injury, including crown scorch and cambial death (Davis et al. 2012; Lerch et al. 2016). Fires can therefore contribute to maintaining local MPB populations (Elkin and Reid 2004; Powell et al. 2012), but MPB response was found to pulse and recede within two years post-burn, thereby rarely generating sustainable outbreak populations with the potential for spread to adjacent stands (Davis et al. 2012; Powell et al. 2012).
Ecological change as a result of beetle outbreaks and wildfire also has important implications for wildlife populations (Saab et al. 2014; Saab and Powell 2005). Fire- and bark beetle-caused tree mortality provides important habitat for wildlife dependent on snags and downed logs by providing nesting, roosting, and foraging substrate (e.g., several woodpecker species) (Saab et al. 2019). Disturbance-caused tree mortality, however, reduces occupancy of other species dependent on live foliage for nesting and foraging habitat (e.g., several songbird species) (Latif et al. 2021; Mosher et al. 2019).
Predicting landscape responses to MPB, wildfire, and climate is problematic because of the inherent complexity of their interactions (Loehman et al. 2020), the long time periods and broad spatial scales over which the disturbances co-occur (Archibald et al. 2013), and the important influences of historical and current land use on antecedent and future disturbance patterns and responses. Management actions, such as fire suppression and fuel treatments, confound projections of landscape responses to the interactions. Landscape simulation models provide an alternate method for assessing dynamics of large-scale interacting processes at appropriate temporal and spatial scales. Keane et al. (2011) developed the FireBGCv2 simulation modeling platform specifically to explore interactions of climate, vegetation, fire, and other disturbance agents. The original FireBGCv2 module that simulated MPB dynamics was empirical, simplistic, and overly generalized due to a general lack of comprehensive information to create a mechanistic simulation of MPB dynamics (Keane et al. 2011). Initiation of MPB outbreaks and outbreak severity were inferred from landscape conditions and a coarse evaluation of climate conditions, and MPB outbreaks were only held in check by winter temperature thresholds known to result in cold-induced MPB mortality. Tree mortality from MPB was simulated using a series of empirical regression equations dependent on tree diameter and developed over 50 years ago (Amman and Cole 1983, Cole et al. 1985). Recently developed, mechanistic MPB models predict influences of thermal history on MPB cold tolerance (Régnière and Bentz 2007) and developmental timing and adult emergence synchrony (Powell and Bentz 2009, 2014), both important factors in MPB population dynamics. We coupled these models with more up-to-date knowledge of MPB tree attack dynamics and stand susceptibility (Randall et al. 2011) within the FireBGCv2 platform to create a more complex, mechanistic MPB module.
In this study, our two major objectives were to (1) evaluate the importance MPB dynamics and its simulation rigor (no MPB, simple empirical simulation, and complex mechanistic simulation) on simulated landscape responses under future climates and land management actions and (2) assess the impacts of management actions on landscape-scale MPB, fire, and climate interactions. Simulations of these interactions over the entire western US are computationally intractable using the FireBGCv2 landscape model. Therefore, we selected three large, forested landscapes in the western US with varying fire regimes and densities of pine species that are known hosts to MPB. We then implemented a multifactorial simulation design that included three MPB factors (no MPB, simple empirical module, complex mechanistic module), two climate scenarios (observed historical weather, RCP 8.5 future climate), two wildfire suppression levels (no suppression, 96% suppression), and three levels of fuel treatment effort (no treatments, low and moderate intensity treatment).
Methods
Study areas
In our simulation experiment, we selected three western US study areas (Fig. 1) that we felt represented a wide range of climate, vegetation, and fire regimes. The Ochoco watershed (OCH) in central Oregon is a 168,000 ha, dry mixed-conifer ecosystem consisting of low elevation ponderosa pine (Pinus Ponderosa Douglas ex C. Lawson) and Douglas-fir Pseudotsuga menziesii (Mirbel) Franco) forests, middle elevation lodgepole pine (Pinus contorta Douglas) and subalpine fir (Abies lasiocarpa (Hooker) Nuttall) forests, and upper elevation whitebark pine (Pinus albicaulis Engelm.) forests, with historically frequent (25- to 100-year return intervals), low-severity fires. The Elkhorn watershed (ELK) in central Montana is a 140,000 ha, xeric mixed-conifer ecosystem of low elevation ponderosa pine and Douglas-fir forests and high elevation lodgepole pine, whitebark pine, and subalpine fir forests, with moderately frequent (~ 20–50-year return intervals), low severity fires at lower elevations, and infrequent, high severity fires in the upper elevations. This landscape had the least area dominated by the MPB host pine species. The Yellowstone Central Plateau (YCP) in Yellowstone National Park, Wyoming, is a 175,000 ha high elevation landscape used in a previous simulation study (Clark et al. 2017) that consisted mainly of mature lodgepole pine forests with scattered late-successional subalpine fir and whitebark pine stands at the highest elevations. YCP mostly had an infrequent (> 150 year) fire return interval and stand-replacement fire regime.

Map showing the location of the three landscapes (Ochoco-OCH, Elkhorns-ELK, Yellowstone Central Plateau-YCP) (in black) in the western US used in this simulation experiment. All three have pine forests with a history of MPB outbreaks. The Elkhorns have significantly less pines than the other landscapes. The ELK and OCH were initialized and parameterized specifically for this study while Clark et al. (2017) quantified the YCP landscape
FireBGCv2 model
FireBGCv2 is a mechanistic, individual-tree, gap model that is implemented in a spatial domain. The model was developed by integrating empirically derived deterministic functions with stochastically driven algorithms to approximate landscape and ecosystem behavior across time and space (Keane et al. 2011, 1996) (Fig. 2). The executable FireBGCv2 program used for this project is available upon request and Keane et al. (2011) document model algorithms, assumptions, parameters, and validations.

A schematic of the FireBGCv2 model. The right diagram shows important ecological processes and the scale that they are simulated including the mountain pine beetle (MPB), fire, climate, and fuel treatments (Land use). The middle diagram shows the five scales represented in FireBGCv2. Stand-level processes are simulated on a vignette of a stand called a plot. This plot is comprised of various ecosystem components (upper right panel). The flow of energy, carbon, water, and nitrogen is simulated across each of these components using mechanistic algorithms detailed in the Keane et al. (2011) document. Biophysical processes such as evaporation, transpiration, photosynthesis, and respiration are governed by five daily weather variables: precipitation, maximum and minimum temperature, radiation, and humidity
The FireBGCv2 modeling platform integrates models of fire behavior and effects (FIRESUM (Keane et al. 1989), FOFEM (Reinhardt et al. 1997), and FARSITE (Finney 1998)) with the BIOME-BGC ecosystem model (Running and Hunt 1993; Thornton et al. 2002; White et al. 2000) and other modeled processes that describe tree and vegetation growth and mortality (Keane et al. 2011). The FireBGCv2 modules important in this study include simulation of stand-level management actions including timber harvest, tree planting, fuels treatments, and prescribed burning (Keane et al. 2019, 2011); detailed fire algorithms that simulate natural ignitions based on weather, fuels, and topography; and spread fire across the simulation landscape based on vectors of wind and slope (Rothermel 1972), fire suppression, expressed as a proportion of naturally ignited fires that are suppressed at initial attack, an input parameter (Keane et al. 1996), and modeling of fire effects to vegetation based on fire intensity and soil heating (Reinhardt and Keane 1998).
Empirical mountain pine beetle FireBGCv2 module
The empirical MPB module initiated MPB population outbreaks based on percent landscape dominated by host pine species and two cold thresholds (− 20 and − 40 °C) that represent temperatures lethal to developing MPB. MPB-induced mortality of all pines larger than 20 cm DBH was then calculated using Amman and Cole (1983) algorithms as implemented in FVS (Crookston and Dixon 2005) for years with favorable, non-lethal freezing temperatures (see Keane et al. 2011 for further details).
Mechanistic mountain pine beetle module
The mechanistic MPB module implemented in FireBGCv2 was based on a demographic model developed by Powell and Bentz (2009, 2014). This model simulated annual MPB population growth (R) as a function of temperature-dependent phenology as determined from daily maximum and minimum temperatures (Régnière et al. 2012). The demographic model incorporates the important role of tree defense in MPB success by connecting adult emergence timing (i.e., phenology) with population consequences based on emergence timing required for successful mass attacks on trees. High R values (R > 1) signify thermal conditions that support highly synchronized and seasonally appropriate adult emergence, and low R values indicate either unsynchronized emergence or emergence outside of a seasonally appropriate time window.
Cold temperature is considered a significant mortality factor in MPB (Weed et al. 2015). Acclimation of compounds that confer tolerance to extreme cold occurs over an extended time period as temperatures decrease in the fall prior to winter (Bentz and Mullins 1999). Here, we used a previously developed model to estimate annual probabilities of population survival as a function of daily maximum and minimum temperatures that influence larval cold tolerance from autumn to the following spring (Régnière and Bentz 2007).
Results from phenology, demographic, and cold tolerance models were linked by assuming that population growth would be most affected when a cold event results in extremely low offspring survival (also see Bentz et al. 2019). Several user input parameters for the mechanistic MPB model were required, including a threshold value at which cold mortality was introduced in a simulation year. If predicted survival from cold fell below the cold tolerance threshold, population growth predicted from the demographic model was reduced by the probability of cold mortality derived from the cold tolerance model. For FireBGCv2 results reported here, we assumed only extreme cold events would influence population growth. If predicted annual probability of survival from cold was ≤ 5%, population growth predicted from the module was multiplied by the probability of survival, thereby reducing population growth. No effect due to cold was applied if survival was > 5%.
The mechanistic MPB module does not predict initiation of an outbreak, but instead simulates population growth following initiation. To introduce MPB to a FireBGCv2 landscape, a background beetle mortality probability (set at 0.005) was used so every susceptible pine tree (diameter breast height (DBH) > 12.5 cm) in the stand had a 0.5% chance of dying from a random, endemic MPB attack in the current year. At the end of each simulation year, FireBGCv2 looped through all trees on a simulation plot and determined probabilistically whether each tree lives or dies due to beetles based on its probability of death. If a tree is chosen to be killed by beetles, based on this random draw, it was recorded as a “red” tree (i.e., dead tree with red needles) in the subsequent year and treated as a snag on the simulation plot, with its death due to MPB recorded.
Growth of MPB populations in a simulation year, once initiated, was also determined by the MPB mechanistic module, which combines effects of MPB phenology, demography, and cold tolerance models, as described above. Computed R-values represent ratios of new trees that will be killed in the stand in the current year based on the number of “red” trees killed the previous year, and daily maximum and minimum temperatures for that year. Daily weather was simulated at the site level; therefore, each site will have a different predicted R-value each year.
To implement tree mortality, stands with sufficient host tree species of adequate size were initially identified. To do this, we used a hazard rating system developed to estimate stand-level susceptibility of multiple pine species to MPB attack based on (1) average diameter of pines in the stand, (2) total basal area of all trees, and (3) proportion of the stand basal area comprised of pines (Randall et al. 2011). Based on these criteria, a single hazard rating is calculated on a scale from 0 to 27, with values 2 to 17 considered moderate, and values ≥ 18 considered high susceptibility. At the start of each simulation year, a “susceptibility” parameter was calculated using the hazard rating criteria for each stand. Another input parameter was the value above which a stand is considered susceptible. For the model results reported here, stands with a moderate rating (2–17) were considered susceptible to MPB attack for the current simulation year.
When the number of predicted red trees for a given year surpassed background beetle mortality threshold, the predicted number of red trees was then used as the population initiation for the next years’ simulation. At low levels of MPB (< 10% of host trees attacked the previous year), only small host trees (12.5 < DBH < 18 cm) under the most stress (> 5 years of growth stress; see Keane et al. (2011)) were preferentially attacked. Tree stress in FireBGCv2 occurred when diameter growth dropped below a threshold value because of a combination of water stress, crowding, shading, and atypical temperatures. Each simulated tree had a counter recording the number of years that it has been under reduced growing conditions due to stress. At low MPB attack levels (< 10% of host trees attacked the previous year), the probability of death due to MPB assigned to each tree in the simulation plot for a susceptible stand was weighted by (1) total number of trees predicted to be killed according to the R-value and (2) stress level of each small tree as compared to all other available small trees on the stand. With increasing MPB population size (10–25% of total pines in the stand were attacked the previous year), stress was no longer considered an important factor in tree attack. At the highest levels of MPB presence (> 25% of total pines in the stand the previous year were attacked), only the largest host trees (DBH > 20 cm) were targeted. At the end of a simulation year, the fate of each tree on the simulation stand was determined by a random draw compared with its individual probability of death due to MPB. Due to the structure of FireBGCv2 (Fig. 2), probabilities of MPB death for individual trees were determined at the stand level, although random draws determine the fates of individual trees.
Simulation experiment
Our factorial simulation experiment consisted of replicated model runs for all combinations of four factors, with two or three levels for each factor (Table 1). Factors and levels included:
-
(1)
Mountain pine beetle (MPB)—no modeled MPB, MPB simulations using the simple empirical module, and MPB simulations using the complex mechanistic module as discussed in detail above.
-
(2)
Fire suppression level (FSL)—no suppression (historical fire regime with no fires suppressed) and full suppression (96% of fires suppressed which represents the current level for each of the three landscapes).
-
(3)
Fuel treatment effort (FTE)—A “fuel treatment” is simulated by the removal of trees to reduce canopy fuels and a prescribed burn to reduce surface fuels (details discussed later in this section. There were three levels of fuel treatments—none, a passive, low-level fuels program (~ 1% landscape treated annually), and an aggressive, moderate-level fuels program (~ 5% landscape treated annually)
-
(4)
Climate (CLI)—an historical climate and a projected future climate under RCP8.5 scenario.
These last three factors (FSL, FTE, and CLI) were included to evaluate if differences in output from MPB modules were also evident for past and future climates, historical and future fire regimes, and after implementation of fuel treatments. The nine response variables in Table 1 were output at the end of every simulation decade over a 400-year simulation period for five replicates per factorial combination resulting in 200 observations per response variable per scenario. The short 400-year time span relative to some fire return intervals (e.g., YCP) was chosen because of a lack of adequate computing resources; it often took weeks to simulate landscape dynamics over this time span and simulation times for longer time spans increase exponentially.
Historical climate data were derived from a nearby weather station for each simulation landscape (see Table 1) using techniques from previous FireBGCv2 projects (Clark et al. 2017; Keane et al. 2019, 2011). Weather data included maximum and minimum temperatures and precipitation. Temperatures and precipitation, and also solar radiation and humidity, were extrapolated across each landscape using the model MTCLIM (Hungerford et al. 1989). Future climate was derived by offsetting daily, historical climate variables by offsets (Delta method) as described in (Clark et al. 2017; Loehman et al. 2010) (e.g., we added a temperature value computed from climate projections, called an offset, to each daily temperature value). Offsets were derived from CNRM-CM5 (National Centre of Meteorological Research, France) 30-arc-second resolution (~ 800-m) climate model, the highest ranked model overall for the three landscapes (Rupp et al. 2013), for the Coupled Model Intercomparison Project Phase 5. We used the 8.5 Representative Concentration Pathway (RCP85) data, an emissions scenario that predicts the highest average annual temperature increases among various possible emissions scenarios (about 6 °C above pre-industrial levels) because it represents an extreme climate scenario (Peters et al. 2012). The Delta method is commonly used when dealing with daily weather data because future daily projections poorly represent climate variability (Keane et al. 2015b). For simulations using historical climate, we continually repeated the 60–100 + year weather record for the full 400-year simulation, and for future climate simulations, we also cycled the weather years but ramped weather so that all offsets were fully implemented after 100 years, and then kept offsets constant for the remaining simulation time (Holsinger et al. 2014).
A fuel treatment in this study was a combination of simulated silvicultural cuttings to remove all fire-sensitive, shade-tolerant species down to 4.0-cm diameter at breast height followed by a low intensity prescribed burn (~ 400 kW m−1 fireline intensity) to reduce slash and kill understory seedlings and saplings. FireBGCv2 simulated treatments only in stands that were (1) above a minimum basal area (10 m2 ha−1); (2) contained fire-sensitive species (Table 1); and (3) had high fuel loadings (> 5 kg m−2). There were many simulation years when few stands met these criteria so treatment level targets were not reached (Keane et al. 2019).
Nine response variables (Table 1) were output every 10 years for a simulation time series of 41 observations per variable per run, and then performed five replicate runs of each scenario combination. We selected these nine response variables because they were (1) often important in land management, (2) uncorrelated to each other (Keane et al. 2019), (3) and used in past FireBGCv2 projects (Holsinger et al. 2014; Keane et al. 2019, 2018) with success. And, for each landscape and scenario, we initiated simulations with the same tree species list (Table 2), which dictates tree species available to inhabit or migrate into the landscape during simulations.
Statistical analysis
We compared responses among scenarios by fitting generalized linear mixed models that included the fixed effects of MPB, FSL, FTE, CLI, landscape, and year, and a first-order autoregressive covariance structure to account for temporal autocorrelation (Box et al. 1994; Pinheiro et al. 2007). All two-way interactions were included in the model. Separate models were fit for each landscape and response variable. We used the significance of each factor (P ≤ 0.05) and standard boxplots to assess differences between each MPB/FSL/FTE/CLI factorial scenario. All mixed model analyses were performed in the open source statistical language R (Team 2017) using the glmmTMB (Brooks et al. 2017) and car p (Fox and Weisberg 2019) packages.
Of the nine response variables, we focused on the differences across scenarios using three ecological variables: (1) basal area of MPB host pine species (BAP), (2) proportion of landscape in MPB host species (HOSTAREA), and (3) proportion of the simulation landscape burned annually (PLBURN) (Figs. 3, 4, and 5). We selected these response variables because we judged that they were the most important for understanding influences of MPB, fire, and climate interactions. The remaining six variables were used as explanatory variables and their simulated results are shown in supplementary material (Figures S1,S2,S3,S4,S5 and S6).

Boxplots of FireBGCv2 results (n = 200, 5 reps of decade output over 400 years)for basal area of mountain pine beetle (MPB) host pine trees (BAP; m2 ha.−1) showing the influence of the detail of MPB simulation (none = red, empirical = blue, mechanistic = green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP), fuel treatment effort (none, passive, aggressive), and fire suppression level (none, suppression) under A historical climate and B future climate (RCP8.5) (Table 1). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range

Boxplots of FireBGCv2 simulation results for the proportion of the landscape in mountain pine beetle (MPB) host pine species (HOSTAREA; proportion) showing the influence of the detail of MPB simulation (none = red, empirical = blue, mechanistic = green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP), fuel treatment effort (none, passive, aggressive), and fire suppression level (none, suppression) under A historical climate and B future climate (RCP8.5) (Table 1). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range

Boxplots of FireBGCv2 simulation results of the annual percent of landscape burned (PLBURN %) showing the influence of the influence of mountain pine beetle (MPB) simulation (none = red, empirical = blue, mechanistic = green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP), treatment effort (none, passive, aggressive), and fire suppression level (none, suppression) under A current climate and B future climate (RCP8.5). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range
Recognizing that differences across the four factors and their levels might depend on combinations of all nine response variables (Keane et al. 2019), we used principal component analysis (PCA) on the nine response variables in Table 1 to reduce the dimensionality of the data and to evaluate differences among scenarios and levels for each landscape. Only the scores of the first principal component (PC1) were evaluated in our PCA analysis to detect significant differences (P ≤ 0.05) using methods described in the previous paragraph (Fig. 6). The PCA analyses were performed in R using built in functions.

Boxplots of the scores of the first PCA axis (PC1) using all variables in Table 1 in the PCA analysis of FireBGCv2 simulations showing the influence of the influence of mountain pine beetle (MPB) simulation (none = red, empirical = blue, mechanistic = green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP), fuel treatment effort (none, passive, aggressive), and fire suppression level (none, suppression) under A current climate and B future climate (RCP8.5). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range
Results
Sensitivity of ecological responses to the complexity of MPB sub-models
The significance of MPB module (none, empirical, mechanistic) on predictions of the three focus and six explanatory response variables depended on the landscape simulated and FSE, FTE, and CLI scenarios (Table 3, Figs. 3, 4, 5, and 6, and Figures S1,S2,S3,S4,S5 and S6). First and most importantly, MPB module type influenced predicted BAP, HOSTAREA, and PLBURN on the OCH landscape, BAP and HOSTAREA on the YCP, and only BAP on the ELK landscape (Figs. 3, 4, and 5). In both historical and future climates, predicted BAP and HOSTAREA were lower (> 30%) using the more complex mechanistic MPB module compared to simulation runs with no-MPB included (Figs. 3 and 4). Low values for BAP and HOSTAREA were direct consequences of greater predicted MPB-killed BA by the mechanistic MPB module, particularly on the YCP landscape (Fig. S5). Significant differences (> 50%) between the mechanistic and the other two MPB modules for the YCP landscape were supported by values of PC1 (Fig. 6).
Predictions of BAP and HOSTAREA, but not PLBURN, were also the most variable in historical climate simulations using the mechanistic MPB module (see width of box and whiskers in Figs. 3 and 4). In future climates, however, variability in predicted BAP and HOSTAREA was highest when the empirical MPB model was used, especially for YCP (Figs. 3 and 4), which may reflect that future climate was outside the weather information used to create statistical equations.
Historical climate simulations with no-MPB resulted in greater BAP (> 20%) on both the OCH and ELK landscapes, and greater HOSTAREA on the OCH, but no differences between no-MPB and empirical MPB model simulations were observed on the YCP (Figs. 3 and 4). In future climate scenarios, however, the no-MPB simulations predicted greater BAP and HOSTAREA on all three landscapes, although variability in model outcomes was also greater, particularly for the YCP landscape. In the historical climate simulations, PLBURN was greater on the OCH landscape compared to the ELK and YCP, regardless of the MPB model type. This prediction was only found for no suppression treatments in historical climate simulations. In future climate scenarios, FSE, FTE, and MPB model type influenced predicted values of PLBURN.
Influence of management on MPB, fire, and landscape interactions
We found that the aggressiveness of fuel treatments (FTE) had relatively minor effects on most response variables across all three landscapes (Figs. 3, 4, 5, and 6), even though some differences were significant at the FTE factor level (Table 3). Moreover, FTE had little effect on MPB-killed basal area, especially when fires were allowed to burn on the landscape (Figure S5). In future climate scenarios, PLBURN was predicted to be greater on the OCH than both other landscapes and was also greater in no suppression compared to suppression simulations. PLBURN was also greater in future climate scenarios for the YCP and ELK landscapes (~ 25% greater), but only in no suppression simulations (Fig. 5). Simulations with no-MPB and no suppression resulted in less PLBURN on the OCH landscape as the level of FTE became more aggressive (Fig. 5). Neither FSE, FTE, nor MPB model type influenced predicted PLBURN on the YCP and ELK landscapes. Relative to historical simulations, HOSTAREA and BAP were greater in future climate scenarios on the OCH for the mechanistic MPB model and no-MPB simulations, relative to the simulations with MPB included (Figs. 3 and 4). On the YCP landscape, variability in HOSTAREA and BAP increased in the future climate simulation runs that included MPB.
Forest fuels, represented by canopy bulk density (CBD; Figure S2), coarse woody debris (CWD; Figure S2), and fine woody debris (FWD; Figure S3) were similar across all three MPB model scenarios. FTE did not cause major reductions in the wildland fuel output variables. FSE had a much greater influence on fuels and fire dynamics than fuel treatment. In both historical and future climate scenarios, HOSTAREA on the ELK landscape was predicted to be very low, regardless of the MPB model type, FSE or FTE.
Discussion
The key finding from this simulation effort is the importance of including MPB dynamics in landscape models that simulate compositions and structures of current and future western US landscapes that contain MPB pine host species. Stand, landscape, and regional models that ignore impacts of this important disturbance are missing meaningful interactions of a major ecological disturbance (Kurz et al. 2008; Liu et al. 2011). We found significant differences and increased variability in simulated projections of landscape composition and basal area when either the empirical or mechanistic MPB module was used, compared to simulations with no-MPB. However, the mechanistic MPB module consistently predicted less BAP and HOSTAREA in historical and future climate scenarios relative to no-MPB but did not influence the third focus variable PLBURN. The mechanistic MPB module also predicted the greatest MPB-killed basal area. In addition, we found greater variability in predictions of BAP and HOSTAREA in historical simulations when the mechanistic MPB module was used. This high variability is a result of more ecological interactions represented in the mechanistic MPB module as it was integrated within FireBGCv2, including direct factors such as tree stress, climate, fire, and forest structure and composition (Keane et al. 2011). By contrast, variability in BAP and HOSTAREA was greatest in future climates when the empirical MPB module was used, especially for the YCP landscape. The likely reason for odd model behavior is that the empirical MPB module was largely based on statistical algorithms and cold temperature thresholds that were inappropriate because future weather was outside the bounds of data used to create the original equations and estimate important thresholds.
The mechanistic MPB module is likely to provide more realistic projections than the empirical model under future RCP8.5 climates because that module (1) includes an explicit representation of the influence of weather on MPB population success, (2) contains numerous interactions with other ecosystem variables (e.g., tree size and abundance, previous year MPB-caused tree mortality), (3) integrates several interactions in tree mortality algorithms, and (4) is not limited by the narrow ranges of empirical data used in the empirical MPB module. MPB is an important disturbance process in western US pine forests, and our results suggest it is the interactions of MPB population dynamics, climate, and their effects with other ecological processes that dictate landscape model projections (Bockino 2008; Jentsch et al. 2021; Keane et al. 2015a; Loehman et al. 2017).
Another important finding is that there are great differences among landscapes for most response variables. Simulation results for the ELK landscape, for example, differed from the OCH and YCP in terms of predicted proportion of pine on the landscape in both historical and future climates. This could be because the inventory data used to initialize FireBGCv2 for ELK contained mostly Douglas-fir and subalpine fir, and pine (MPB host) species were rare (ponderosa, lodgepole, and whitebark pine). As a result, differences across scenarios for the ELK landscape were difficult to detect because of low pine populations. Results from the YCP landscape often contained greater variation for all nine variables potentially due to the long fire return intervals (> 250 years) and the dominance of colder climates which favored lodgepole pine. Frequent fire regimes as found on the ELK and OCH landscapes tend to create shifting mosaics of vegetation communities that are surprisingly stable (Keane et al. 2019). The OCH landscape had a warmer environment and was dominated by ponderosa pine with higher annual % landscape burned, and a lower proportion of the historical landscape with fire adapted species compared to YCP.
The simulation of MPB dynamics, regardless of the module type, had significant impacts on burned area (PLBURN), but only for the OCH landscape and only under fuel treatments. The OCH landscape had a frequent fire regime that likely can be replicated by fuel treatments. In general, wildfire had a greater effect on wildland fuels in our simulations by decreasing CBD (Figure S2), CWD (Figure S3), and FWD (Figure S4) in both historical and future climates, especially for the OCH and ELK landscapes. The short simulation time span (only 400 years) was not long enough for the long fire return intervals for the YCP landscape (~ 250 years), resulting in fewer fires that probably masked impacts of wildfire on canopy and surface fuels. MPB-caused mortality often results in the deposition of canopy fuel from dead pines on the ground along with the dead canopy fuel killed by fire (Jenkins et al. 2012), but we found little change in simulated surface fuels as a consequence of MPB-caused tree mortality. These results are supported by Stalling et al. (2017) who found little change in forest floor fuels within a decade following MPB outbreaks. Under future climates, fire suppression was predicted to result in greater BAP on the YCP landscape and more MPB-killed basal area compared to no suppression actions in future climates. Interestingly, the opposite effect was seen in the OCH and ELK landscapes where greater MPB-killed basal area was predicted when there was no fire suppression. Fire suppression on the YCP allowed established pines to grow into trees of sufficient size to be vulnerable to MPB, thereby resulting in landscape homogenization favoring pine and, in turn, creating the potential for high severity outbreaks of MPB.
Another surprising result was minor influence of fuel treatments on most simulated response variables, especially their low impact on tree mortality due to MPB or fire. Aggressive fuel treatments reduced total and pine basal area and percent landscape burned on the OCH without MPB, but this was the only case. Keane et al. (2019) found that fuel treatments are only effective at restoring landscapes if implemented under high fire suppression levels, which was validated in this effort using different landscapes. Wildland fire, whether as a controlled or uncontrolled wildfires, tends to do most of the work of fuel treatments but with significantly less resources (Keane et al. 2019). Fuel treatments, regardless of level of effort, didn't indicate any effect on MPB-caused tree mortality except for the OCH landscape without MPB simulation. This is partially a result of wildfire effects mitigating the limited influence of fuel treatments over long time periods.
Limitations
The current implementation of MPB modules within the FireBGCv2 simulation platform did not incorporate dispersal of MPB outbreaks across stands because of the additional computational power required to integrate MPB dispersal models (Powell and Bentz 2014). Our simulations took over 7 months of computing time on a Cray supercomputer and the inclusion of a MPB spread simulation might have doubled or tripled that time. The consequence is that MPB outbreaks are limited to stands and not landscapes. Any excess of trees predicted to be killed by MPB disappear when the availability of trees at the stand level is exhausted. However, this side-effect is somewhat mitigated by having shared values for predicted R-values across sites. Neighboring stands within a site do not share access to individual trees; however, they experience the same bioclimatic conditions that are used to predict MPB population growth. Altering background MPB mortality rate could help offset the consequences of the lack of dispersal among stands (e.g., by ensuring that enough initiation red trees are present on the landscape to overcome stand-level sample size issues); however, it would be difficult to decide on a credible value. As a result, the magnitude and variability of MPB impacts on the simulated landscapes may have been much greater if dispersal effects had been included.
Results of our statistical analysis of the simulation output also have shortcomings. Our analyzed output was generated every 10 years over the 400-year time span for five replications giving us a large 200 observations’ data set. This large number of observations provides great statistical power, but as White et al. (2014) note, this statistical power is easily created in simulation projects by simply outputting more data. Because of this, it may be best to de-emphasize the importance of the p-values in Table 3 and put more confidence in the visual comparison of the box and whisker plots (Figs. 1, 2, 3, 4, 5, and 6 and S1, S2, S3, S4, S5 and S6). The variabilities of output variables over the 200 observations are also important. When employing mostly deterministic models, such as FireBGCv2 (the stochasticity in FireBGCv2 is only 3–5% of the variance) (Keane et al. 2011), the magnitude of the variance becomes an important indicator of the realism of simulated results. In general, increased variability often results from increased interactions causing non-linear model behaviors (Keane 2019). Obviously, the simulated variability is only a rough approximation of the actual ecological variability, but its comparison across scenarios provides an indication of possible realism in model results. For example, the same amount of variability across the box and whisker plots may indicate that the model may not be simulating alternate interaction pathways.
Summary and conclusions
Provided here are simulation results for three landscapes using FireBGCv2 integrated with empirical and mechanistic MPB modules that demonstrated the importance of including MPB when simulating landscape dynamics. As expected, adding MPB effects decreased total and pine basal area, regardless of fire suppression in both historical and a future climate, although there was little effect on the annual percent of landscape burned. Fire suppression increased large pine abundance and reduced annual percent of landscape burned but had varying effects on MPB killed basal area depending on the landscape and climate scenario. Simulations using integrated MPB models underscore the significant influence of MPB disturbances in these landscapes and highlights future stability in species composition with and without management. We anticipate integrating wildlife population models with these simulations to evaluate competing risks of MPB, wildfire, and fuels treatments on future habitat quality (based on forest structure and composition) and the distributions of selected wildlife species (e.g., (Saab et al. 2019; Saab and Powell 2005).
Availability of data and materials
All data are available from the authors.
References
Abatzoglou, J.T., and A.P. Williams. 2016. Impact of anthropogenic climate change on wildfire across western US forests. Proceedings of the National Academy of Sciences 113: 11770–11775. https://doi.org/10.1073/pnas.1607171113.
Archibald, S., C.E.R. Lehmann, J.L. Gómez-Dans, and R.A. Bradstock. 2013. Defining pyromes and global syndromes of fire regimes. Proceedings of the National Academy of Sciences 110: 6442–6447. https://doi.org/10.1073/pnas.1211466110.
Bentz, B.J., et al. 2010. Climate change and bark beetles of the Western United States and Canada: Direct and indirect effects. BioScience 60: 602–613. https://doi.org/10.1525/bio.2010.60.8.6.
Bentz, B.J., J.P. Duncan, and J.A. Powell. 2016. Elevational shifts in thermal suitability for mountain pine beetle population growth in a changing climate. Forestry 89: 271–283.
Bentz, B.J., A.M. Jönsson, M. Schroeder, A. Weed, R.A.I. Wilcke, and K. Larsson. 2019. Ips typographus and Dendroctonus ponderosae models project thermal suitability for intra-and inter-continental establishment in a changing climate Frontiers in Forests and Global. Change 2: 1.
Chapman, T.B., T.T. Veblen, and T. Schoennagel. 2012. Spatiotemporal patterns of mountain pine beetle activity in the southern Rocky Mountains. Ecology 93: 2175–2185.
Davis, R.S., S. Hood, and B.J. Bentz. 2012. Fire-injured ponderosa pine provide a pulsed resource for bark beetles. Canadian Journal of Forest Research 42: 2022–2036.
Edburg, S.L., et al. 2012. Cascading impacts of bark beetle-caused tree mortality on coupled biogeophysical and biogeochemical processes. Frontiers in Ecology and the Environment 10: 416–424. https://doi.org/10.1890/110173.
Evenden, J.C., and A.L. Gibson. 1940. A destructive infestation in lodgepole pine stands by the mountain pine beetle. Journal of Forestry 38: 271–275.
Fettig, C.J., K.D. Klepzig, R.F. Billings, A.S. Munson, T.E. Nebeker, J.F. Negrón, and J.T. Nowak. 2007. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. Forest Ecology and Management 238: 24–53.
Flannigan, M.D., M.A. Krawchuk, W.J. de Groot, B.M. Wotton, and L.M. Gowman. 2009. Implications of changing climate for global wildland fire. International Journal of Wildland Fire 18: 483–507. https://doi.org/10.1071/WF08187.
Hansen, E.M. 2014. Forest development and carbon dynamics after mountain pine beetle outbreaks. Forest Science 60: 476–488.
Hessburg, P.F., J.K. Agee, and J.F. Franklin. 2005. Dry forests and wildland fires of the inland Northwest USA: Contrasting the landscape ecology of the pre-settlement and modern eras. Forest Ecology and Management 211: 117–139. https://doi.org/10.1016/j.foreco.2005.02.016.
Heyerdahl, E.K., L.B. Brubaker, and J.K. Agee. 2001. Spatial controls of historical fire regimes: A multiscale example from the Interior West. USA Ecology 82: 660–678.
Hicke, J.A., M.C. Johnson, J.L. Hayes, and H.K. Preisler. 2012. Effects of bark beetle-caused tree mortality on wildfire. Forest Ecology and Management 271: 81–90. https://doi.org/10.1016/j.foreco.2012.02.005.
Holsinger, L., R.E. Keane, D.J. Isaak, L. Eby, and M.K. Young. 2014. Relative effects of climate change and wildfires on stream temperatures: A simulation modeling approach in a Rocky Mountain watershed. Climatic Change 124: 191–206. https://doi.org/10.1007/s10584-014-1092-5.
Hungerford, R.D., R.R. Nemani, S.W. Running, and J.C. Coughlan. 1989. MTCLIM: A mountain microclimate simulation model. Intermountain Research Station, Ogden, UT: USDA Forest Service.
Jenkins, M.J., W.G. Page, E.G. Hebertson, and M.E. Alexander. 2012. Fuels and fire behavior dynamics in bark beetle-attacked forests in Western North America and implications for fire management. Forest Ecology and Management 275: 23–34. https://doi.org/10.1016/j.foreco.2012.02.036.
Jentsch, P.C., C.T. Bauch, and M. Anand. 2021. Fire mitigates bark beetle outbreaks in serotinous forests. Theoretical Ecology 14: 611–621. https://doi.org/10.1007/s12080-021-00520-y.
Kane, J.M., J.M. Varner, M.R. Metz, and P.J. van Mantgem. 2017. Characterizing interactions between fire and other disturbances and their impacts on tree mortality in western U.S. Forests Forest Ecology and Management 405: 188–199. https://doi.org/10.1016/j.foreco.2017.09.037.
Keane, R.E. 2019. Applying ecosystem and landscape models in natural resource management. Taylor and Francis Group, Baco Raton, Florida USA: CRC Press.
Keane, R.E., R. Loehman, J. Clark, E. Smithwick, and C. Miller. 2015a. Exploring interactions among multiple disturbance agents in forest landscapes: Simulating effects of fire, beetles, and disease under climate change. In Modeling and Mapping Forest Landscape Patterns, ed. A.H. Perera, T.K. Remmel, and L.J. Buse, 201–231. New York, USA: Springer.
Keane, R.E., R.A. Loehman, L.M. Holsinger, D.A. Falk, P. Higuera, S.M. Hood, and P.F. Hessburg. 2018. Use of Landscape Simulation Modeling to Quantify Resilience for Ecological Applications Ecosphere 9: e02414.
Keane, R.E., K. Gray, B. Davis, L.M. Holsinger, and R. Loehman. 2019. Evaluating ecological resilience across wildfire suppression levels under climate and fuel treatment scenarios using landscape simulation modelling. International Journal of Wildland Fire 28: 533–549. https://doi.org/10.1071/WF19015.
Keeling, E.G., and A. Sala. 2012. Changing growth response to wildfire in old-growth ponderosa pine trees in montane forests of north central I daho. Global Change Biology 18: 1117–1126.
Kolb, T.E., et al. 2016. Observed and anticipated impacts of drought on forest insects and diseases in the United States. Forest Ecology and Management 380: 321–334. https://doi.org/10.1016/j.foreco.2016.04.051.
Kulakowski, D., T.T. Veblen, and P. Bebi. 2003. Effects of fire and spruce beetle outbreak legacies on the disturbance regime of a subalpine forest in Colorado. Journal of Biogeography 30: 1–12.
Kurz, W.A., et al. 2008. Mountain pine beetle and forest carbon feedback to climate change. Nature 452: 987–992.
Latif, Q.S., V.A. Saab, and J.G. Dudley. 2021. Prescribed fire limits wildfire severity without altering ecological importance for birds Fire. Ecology 17: 1–22.
Lerch, A.P., J.A. Pfammatter, B.J. Bentz, and K.F. Raffa. 2016. Mountain Pine Beetle Dynamics and Reproductive Success in Post-Fire Lodgepole and Ponderosa Pine Forests in Northeastern Utah PloS One 11: e0164738.
McHugh, C.W., and T.E. Kolb. 2003. Ponderosa pine mortality following fire in northern Arizona. International Journal of Wildland Fire 12: 7–22.
Meigs, G.W., J.L. Campbell, H.S.J. Zald, J.D. Bailey, D.C. Shaw, and R.E. Kennedy. 2015. Does wildfire likelihood increase following insect outbreaks in conifer forests? Ecosphere 6: 1–24. https://doi.org/10.1890/es15-00037.1.
Miller, J.D., H.D. Safford, M. Crimmins, and A.E. Thode. 2009. Quantitative evidence for increasing forest fire severity in the Sierra Nevada and Southern Cascade Mountains. California and Nevada, USA Ecosystems 12: 16–32.
Mosher, B.A., V.A. Saab, M.D. Lerch, M.M. Ellis, and J.J. Rotella. 2019. Forest Birds Exhibit Variable Changes in Occurrence during a Mountain Pine Beetle Epidemic Ecosphere 10: e02935.
Page, W.G., M.E. Alexander, and M.J. Jenkins. 2013. Wildfire’s resistance to control in mountain pine beetle-attacked lodgepole pine forests. The Forestry Chronicle 89: 783–794. https://doi.org/10.5558/tfc2013-141.
Pinheiro J, Bates D, DebRoy S, Sarkar D, Team RC. 2007. Linear and Nonlinear Mixed Effects Models R Package Version 3: 1–89.
Powell, E.N., P.A. Townsend, and K.F. Raffa. 2012. Wildfire provides refuge from local extinction but is an unlikely driver of outbreaks by mountain pine beetle. Ecological Monographs 82: 69–84.
Régnière, J., and B. Bentz. 2007. Modeling cold tolerance in the mountain pine beetle. Dendroctonus Ponderosae Journal of Insect Physiology 53: 559–572.
Reinhardt, E., and R.E. Keane. 1998. FOFEM - a First Order Fire Effects Model Fire Management Notes 58: 25–28.
Runyon, J.B., et al. 2020. Changes in understory vegetation including invasive weeds following mountain pine beetle outbreaks Trees. Forests and People 2: 100038.
Saab, V.A., et al. 2014. Ecological consequences of mountain pine beetle outbreaks for wildlife in western North American forests. Forest Science 60: 539–559.
Scheller, R.M., A.M. Kretchun, E.L. Loudermilk, M.D. Hurteau, P.J. Weisberg, and C. Skinner. 2018. Interactions Among Fuel Management, Species Composition, Bark Beetles, and Climate Change and the Potential Effects on Forests of the Lake Tahoe Basin. Ecosystems 21: 643–656. https://doi.org/10.1007/s10021-017-0175-3.
Schoennagel, T., T.T. Veblen, J.F. Negron, and J.M. Smith. 2012. Effects of mountain pine beetle on fuels and expected fire behavior in lodgepole pine forests, Colorado. USA PLoS ONE 7: e30002.
Shive, K.L., P.Z. Fulé, C.H. Sieg, B.A. Strom, and M.E. Hunter. 2014. Managing burned landscapes: Evaluating future management strategies for resilient forests under a warming climate. International Journal of Wildland Fire 23: 915–928. https://doi.org/10.1071/WF13184.
Six, D.L., and K. Skov. 2009. Response of bark beetles and their natural enemies to fire and fire surrogate treatments in mixed-conifer forests in western Montana. Forest Ecology and Management 258: 761–772.
Stalling, C., R.E. Keane, and M. Retzlaff. 2017. Surface fuel changes after severe disturbances in northern Rocky Mountain ecosystems. Forest Ecology and Management 400: 38–47. https://doi.org/10.1016/j.foreco.2017.05.020.
Swetnam, T.W., and J.L. Betancourt. 1998. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. Journal of Climate 11: 3128–3147.
Weaver, H. 1943. Fire as an ecological and silvicultural factor in the ponderosa pine region of the pacific slope. Journal of Forestry 41: 7–14.
Weed, A.S., B.J. Bentz, M.P. Ayres, and T.P. Holmes. 2015. Geographically variable response of Dendroctonus ponderosae to winter warming in the western United States. Landscape Ecology 30: 1075–1093.
White, J.W., A. Rassweiler, J.F. Samhouri, A.C. Stier, and C. White. 2014. Ecologists should not use statistical significance tests to interpret simulation model results. Oikos 123: 385–388. https://doi.org/10.1111/j.1600-0706.2013.01073.x.
Amman GD, Cole WE (1983) Mountain pine beetle dynamics in lodgepole pine forests. Part II: Population dynamics. USDA Forest Service Intermountain Forest and Range Experiment Station, Ogden, Utah USA
Audley JP et al. (2020) Impacts of mountain pine beetle outbreaks on lodgepole pine forests in the Intermountain West, U.S., 2004–2019 Forest Ecology and Management 475:118403 doi:https://doi.org/10.1016/j.foreco.2020.118403
Bentz B, Mullins D (1999) Ecology of mountain pine beetle (Coleoptera: Scolytidae) cold hardening in the intermountain west Environmental Entomology 28:577–587
Berner LT, Law BE, Meddens AJ, Hicke JA (2017) Tree mortality from fires, bark beetles, and timber harvest during a hot and dry decade in the western United States (2003–2012) Environmental Research Letters 12:065005
Bockino NK (2008) Interactions of white pine blister rust, host species, and mountain pine beetle in whitebark pine ecosystems in the Greater Yellowstone. M.S. Thesis, Univeristy of Wyoming
Box G, Jenkins G, Reinsel G (1994) Time series analysis, forecasting and control. Englewood Clifs. NJ: Prentice Hall,
Brooks ME et al. (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling The R journal 9:378–400
Clark JA, Loehman RA, Keane RE (2017) Climate changes and wildfire alter vegetation of Yellowstone National Park, but forest cover persists Ecosphere 8:e01636-n/a doi:https://doi.org/10.1002/ecs2.1636
Crookston NL, Dixon GE (2005) The forest vegetation simulator: a review of its structure, content, and applications Computers and Electronics in Agriculture 49:60–80
Elkin CM, Reid ML (2004) Attack and reproductive success of mountain pine beetles (Coleoptera: Scolytidae) in fire-damaged lodgepole pines Environmental Entomology 33:1070–1080
Fettig CJ, Hilszczański J (2015) Chapter 14 - Management strategies for bark beetles in conifer forests. In: Vega FE, Hofstetter RW (eds) Bark Beetles. Academic Press, San Diego, pp 555–584. doi:https://doi.org/10.1016/B978-0-12-417156-5.00014-9
Fettig CJ, Mortenson LA, Bulaon BM, Foulk PB (2019) Tree mortality following drought in the central and southern Sierra Nevada, California, U.S Forest Ecology and Management 432:164–178 doi:https://doi.org/10.1016/j.foreco.2018.09.006
Finney MA (1998) FARSITE: Fire Area Simulator -- model development and evaluation. United States Department of Agriculture, Forest Service Rocky Mountain Research Station, Ft. Collins, CO USA
Fox J, Weisberg S (2019) An R companion to applied regression (Third) Thousand Oaks CA: Sage
Geiszler DR, Gara RI, Driver CH, Gallucci VF, Martin RE (1980) Fire, fungi, and beetle influences on a lodgepole pine ecosystem of south-central Oregon Oecologia 46:239–243
Hansen EM, Johnson MC, Bentz BJ, Vandygriff JC, Munson AS (2015) Fuel loads and simulated fire behavior in “old-stage” beetle-infested Ponderosa Pine of the Colorado Plateau Forest Science 61:644–664
Jenkins MA, Hebertson E, Page W, Jorgensen CA (2008) Bark beetles, fuels, fire and implications for forest management in the Intermountain West Forest Ecology and Management 254:16–34
Jenkins MJ, Runyon JB, Fettig CJ, Page WG, Bentz BJ (2014) Interactions among the mountain pine beetle, fires, and fuels Forest Science 60:489–501
Keane RE, Loehman RA, Holsinger LM (2011) The FireBGCv2 landscape fire and succession model: a research simulation platform for exploring fire and vegetation dynamics. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO USA
Keane RE, McKenzie D, Falk DA, Smithwick EAH, Miller C, Kellogg L-KB (2015b) Representing climate, disturbance, and vegetation interactions in landscape models Ecological Modelling 309–310:33–47
Keane RE, Morgan P, Running SW (1996) FIRE-BGC - a mechanistic ecological process model for simulating fire succession on coniferous forest landscapes of the northern Rocky Mountains. United States Department of Agriculture, Forest Service Intermountain Forest and Range Experiment Station, Ogden, UT USA
Keane RE, Stephen F. Arno, Brown JK (1989) FIRESUM -- an ecological process model for fire succession in western conifer forests. USDA Forest Service,
Littell JS, McKenzie D, Peterson DL, Westerling AL (2009) Climate and wildfire area burned in western U.S. ecoprovinces, 1916–2003 Ecological Applications 19:1003–1021 doi:https://doi.org/10.1890/07-1183.1
Liu SG et al. (2011) Simulating the impacts of disturbances on forest carbon cycling in North America: Processes, data, models, and challenges Journal of Geophysical Research-Biogeosciences 116 doi:Artn G00k08: Doi https://doi.org/10.1029/2010jg001585
Loehman et al., 2010 RA Loehman JA Clark RE Keane 2010 Modeling effects of climate change and fire management on western white pine (Pinus monticola) in the northern Rocky Mountains USA Forests 2 https://doi.org/10.3390/f2040832
Loehman RA, Keane RE, Holsinger LM (2020) Simulation modeling of complex climate, wildfire, and vegetation dynamics to address wicked problems in land management frontiers in forests and global change 3 doi:https://doi.org/10.3389/ffgc.2020.00003
Loehman RA, Keane RE, Holsinger LM, Wu Z (2017) Interactions of landscape disturbances and climate change dictate ecological pattern and process: spatial modeling of wildfire, insect, and disease dynamics under future climates Landscape Ecology 32:1447–1459 doi:https://doi.org/10.1007/s10980-016-0414-6
Parker TJ, Clancy KM, Mathiasen RL (2006) Interactions among fire, insects, and pathogens in coniferous forests of the interior western United States and Canada Agricultural and Forest Entomology 8:167–189
Peters GP et al. (2012) The challenge to keep global warming below 2 °C Nature Climate Change 3:4 doi:https://doi.org/10.1038/nclimate1783. https://www.nature.com/articles/nclimate1783#supplementary-information
Powell JA, Bentz BJ (2009) Connecting phenological predictions with population growth rates for mountain pine beetle, an outbreak insect Landscape Ecology 24:657–672
Powell JA, Bentz BJ (2014) Phenology and density-dependent dispersal predict patterns of mountain pine beetle (Dendroctonus ponderosae) impact Ecological Modelling 273:173–185 doi:https://doi.org/10.1016/j.ecolmodel.2013.10.034
Randall CB, Steed BE, Bush R (2011) Revised R1 forest insect hazard rating system user guide for use with inventory data stored in FSVeg and/or analyzed with the forest vegetation simulator
Régnière J, Powell J, Bentz B, Nealis V (2012) Effects of temperature on development, survival and reproduction of insects: experimental design, data analysis and modeling Journal of Insect Physiology 58:634–647
Reinhardt E, R.E. Keane, Brown JK (1997) First Order Fire Effects Model: FOFEM 4.0 User's Guide. USDA Forest Service,
Rothermel RC (1972) A mathematical model for predicting fire spread in wildland fuels. Ogden, Utah
Running SW (2006) Is global warming causing more, larger wildfires Science 313:927–928
Running SW, Hunt ER (1993) Generalization of a forest ecosystem process model for other biomes, BIOME-BGC, and an application for global-scale models. In: Scaling physiological processes: leaf to globe. Academic Press, Inc., pp 141–157
Rupp DE, Abatzoglou JT, Hegewisch KC, Mote PW (2013) Evaluation of CMIP5 20th century climate simulations for the Pacific Northwest USA Journal of Geophysical Research: Atmospheres 118:10,884–810,906 doi:https://doi.org/10.1002/jgrd.50843
Saab VA, Latif QS, Dresser MA, Dudley JG (2019) Woodpecker nest survival, density, and a pine beetle outbreak The Journal of Wildlife Management 83:1387–1400
Saab VA, Powell HD (2005) Fire and avian ecology in North America: process influencing pattern In: Saab, V; Powell, H, eds Fire and Avian Ecology in North America Studies in Avian Biology 30: 1–13:1–13
Team RC (2017) R: A Language and Environment for Statistical Computing
Thornton P, E. et al. (2002) Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests Agricultural and Forest Meteorology 113:185–222
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase western US forest wildfire activity Science 313 doi:https://doi.org/10.1126/science.1128834
White M, A., Thornton P, E., Running SW, Nemani R (2000) Parameterization and sensitivity analysis of the Biome-BGC terrestrial ecosystem model: Net primary production controls Earth Interactions 4:1–85
Whitlock C, Marlon J, Briles C, Brunelle A, Long CJ, Bartlein PJ (2008) Long-term relations among fire, fuel, and climate in the north-western US based on lake-sediment studies International Journal of Wildland Fire 17:72–83
Williams PA et al. (2013) Temperature as a potent driver of regional forest drought stress and tree mortality Nature Clim Change 3:292–297
Acknowledgements
We thank Martha Ellis for coding, testing, and documenting the MPB algorithms for the mechanistic MPB model; Chris Stalling of the USFS Rocky Mountain Research Station for extensive reviews; and the Western Wildland Threat Assessment Center for funding. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Funding
This study was funded by the National Fire Plan, US Forest Service Rocky Mountain Research Station, and the US Forest Service Western Wildland Threat Assessment Center.
Author information
Authors and Affiliations
Contributions
REK helped designed the study, interpreted results, and wrote the majority of the paper; BB wrote the MPB model, provided the code for implementation of the model, redesigned the model to incorporate into FireBGCv2, interpreted the data, and wrote many parts of the paper; LH did all the simulations and results’ analysis and also helped in the writing of the paper; VS helped design the study and edited the final paper; RAL participated in the development of the study and edited the paper. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study did not involve human participants, tissue, or data.
Consent for publication
All figures and tables were created by the authors and all authors give their consent to publish.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government. The findings and conclusions in this publication are those of the author(s) and should not be construed to represent any official USDA determination or policy. This paper was partly written and prepared by US Government employees on official time and therefore is in the public domain and not subject to copyright.
Supplementary Information
Additional file 1:
Figure S1. Basal area of all trees (BA; m2 ha-1) boxplots of FireBGCv2 results showing the influence of the detail of mountain pine beetle (MPB) simulation (none=red, empirical=blue, mechanistic =green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP) and fire suppression level (none, suppression) under (A) historical climate and (B) future climate (RCP8.5). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range. Figure S2. Canopy bulk density (CBD; kg m-3) boxplots of FireBGCv2 results showing the influence of the detail of mountain pine beetle (MPB) simulation (none=red, empirical=blue, mechanistic=green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP) and fire suppression level (none, suppression) under (A) historical climate and (B) future climate (RCP8.5). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range. Figure S3. Coarse woody debris loading (CWD; kg m-2) boxplots of FireBGCv2 results showing the influence of the detail of mountain pine beetle (MPB) simulation (none=red, empirical=blue, mechanistic=green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP) and fire suppression level (none, suppression) under (A) historical climate and (B) future climate (RCP8.5). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range. Figure S4. Fine woody debris loading (FWD; kg m-2) boxplots of FireBGCv2 results showing the influence of the detail of mountain pine beetle (MPB) simulation (none=red, empirical=blue, mechanistic=green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP) and fire suppression level (none, suppression) under (A) historical climate and (B) future climate (RCP8.5). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range. Figure S5. Basal area of all trees killed by mountain pine beetle (BAMPB; m2 ha-1) boxplots of FireBGCv2 results showing the influence of the detail of mountain pine beetle (MPB) simulation (empirical=blue, mechanistic=green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP) and fire suppression level (none, suppression) under (A) current climate and (B) future climate (RCP8.5). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range. Figure S6. Proportion of the landscape composed of fire adapted species (PLFAS; proportion) boxplots of FireBGCv2 results showing the influence of the detail of mountain pine beetle (MPB) simulation (none=red, empirical=blue, mechanistic=green) stratified by landscape (Ochoco-OCH, Elkhorns-ELK, and Yellowstone Central Plateau-YCP) and fire suppression level (none, suppression) under (A) historical climate and (B) future climate (RCP8.5). The horizontal bold line in each boxplot is the median, the box top and bottom are the 25th and 75th percentiles respectively, and the whiskers extend to the smallest or largest values but no further than 1.5*Interquartile range. Figure S7. PCA results for YCP landscape from FireBGCv2 simulations of historical climate, full fire suppression, mechanistic mountain pine beetle module, and no fuel treatments. Circle shows the range that contains 68% of the variation. Names with arrows indicate variable importance in PC1 and PC2 scores. Variable acronyms are defined in Table 1.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Keane, R.E., Bentz, B., Holsinger, L.M. et al. Modeled interactions of mountain pine beetle and wildland fire under future climate and management scenarios for three western US landscapes. fire ecol 18, 12 (2022). https://doi.org/10.1186/s42408-022-00137-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-022-00137-4