
Abstract
Background
Central nervous system (CNS) astrocytes have various functions in the central nervous system (CNS). Many neurodegenerative diseases are associated with astrocyte dysfunction.
Main body of the abstract
Undoubtedly, astrocytes play a crucial role in neurogenesis and synaptogenesis by controlling the intercellular permeability of the blood–brain barrier and maintaining the homeostasis of the extracellular space. Regarding nerve damage, mature astrocytes are divided into A1 and A2 astrocytes. The supportive patterns of reactive astrocytes can be converted into toxic patterns and eventually lead to the development of neurological diseases. Alterations of neurotransmitters, cell communication, receptors, and signaling pathways, especially in the site of inflammation, secretion of inflammatory factors, secretion of growth factors, protein deposition, ion homeostasis, and finally, changes in the size and number of astrocytes are among the most important pathogenic alterations in astrocytes. Astrocytes also exhibit considerable heterogeneity due to the developmental mechanisms they follow and stimulus-specific cellular responses influenced by CNS location, cell–cell interactions, and other factors.
Short conclusion
In recent years, biomolecular advances have led to a better understanding of astrocyte function, allowing them to be considered a therapeutic target in healthy and diseased individuals. Understanding the interactions between astrocytes and other cells will improve our knowledge of the regulation of astrocyte function in homeostasis and new therapeutic targets in future studies.
Similar content being viewed by others


Background
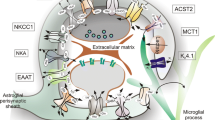
Astrocytes or astroglia are glial cells that resemble a star and are the most abundant glial cells in the central nervous system (CNS) (Hanani and Verkhratsky 2021). The characteristics of adult astrocytes can be classified based on their functional and morphological characteristics and their anatomical location. In terms of morphology, astrocytes are divided into four types (Zhou et al.2019). Fibrous astrocytes are among the most abundant astrocytes in the white matter of CNS, probably supporting glial cells. Protoplasmic astrocytes are the most common astrocytes found in the gray matter of CNS. They are found in the deeper layers of the cerebral cortex and play a crucial role in synapse development and function (Verkhratsky et al. 2018). Polar astrocytes are found in deep layers near the white matter. The density of these cells reflects individual diversity of perception; their appearance is affected by aging, and they have never been observed in the brain of a human infant. The distal processes of protoplasmic and polar astrocytes also make connections with blood vessels and create perivascular end-feet. Despite composing a significant fraction of the human cortex astrocytes, the function of interlaminar astrocytes is unclear (Liu et al. 2017).
Glial fibrillary acidic protein (GFAP) is a major component of astrocyte intermediate filaments, and antibodies to this protein are the most extensively used immunohistochemical marker for astrocyte detection in rat and human tissue (Escartin et al. 2021). Connexins, which maintain the normal morphology and function of astrocytes and allow the passage of molecules between cells and the extracellular matrix, are found in gap junctions and hemichannels. Connexin-43 (Cx43) is a functional entity capable of affecting metabolic gradients that is highly expressed by astrocytes, particularly at the blood–brain barrier (Pekny et al. 2016). Aquaporin-4 (AQP4) is a water channel protein that is expressed by astrocytic end-feet surrounding capillaries and is linked with water transfer into and out of the brain parenchyma (Tang and Yang 2016). The glutamate transporter 1 (GLT-1)/excitatory amino acid transporter 2 (EAAT-2) is the primary transporter responsible for removing glutamate from neuronal synapses and the extracellular environment, and it is expressed particularly by astrocytes (Williams et al. 2005).
Astrocytes play essential roles in CNS growth, homeostasis, and response to injury. These cells can change their functions in response to the needs of the cells in their surroundings. In addition to the regulation of synaptogenesis, synaptic signaling, formation of the blood–brain barrier, control of cerebral blood flow, energy provision, metabolism, ion and pH homeostasis, antioxidant response, inflammation, and the development, homeostasis, and reconstruction of the white matter, astrocytes are also responsible for regulating a variety of other functions (Molina-Gonzalez and Miron 2019).
Researchers have shown that astrocytes are involved in synaptic, circuit, and behavioral functions; in fact, these cells are active communication elements of the central nervous system capable of releasing various regulatory signals. It is estimated that an astrocyte contacts with more than 100,000 synapses at once and plays a role in synaptic signaling through the release and uptake of neurotransmitters (Farmer and Murai 2017). Astrocytes play a remarkable role in enhancing synaptic strength and function by expressing neurotransmitter receptors and releasing neurotransmitters, such as glutamate, GABA (Gamma-aminobutyric acid), adenosine triphosphate, and serine D (Filous and silver 2016; Ahadiat and Hosseinian 2022a, b).
Astrocytes are structurally attached to synapses via their outgrowths, and this structural connection contributes to synaptic signaling via neurotransmitter release and absorption. Astrocytes play an important role in increasing synaptic strength and function by expressing neurotransmitter receptors and releasing neurotransmitters like glutamate, GABA, adenosine triphosphate, and serine D.Nerve activity causes the release of neurotransmitters, which cause the generation of calcium signals in these cells via neurotransmitter receptors expressed on astrocytes, a well-known phenomenon known as astrocyte excitability. Eventually, this transmission causes the production of different transmitters like prostaglandins, which allow them to modify synaptic function. (Emirandetti et al. 2006; Bolton and Eroglu 2009; Varela-Echevarría et al. 2017).
Regulation of neurotransmitter homeostasis, including glutamate, is one of the important functions of astrocytes since they should quickly clear these neurotransmitters from the extracellular space. Animal studies show that reactive astrocytes lose their supportive role after activation and become toxic in developing neurological diseases (Dutta et al. 2018).
The goal of this review is to provide comprehensive and innovative knowledge about the role of activated astrocytes in the development of neurological disorders such as Alzheimer's, Parkinson's, amyotrophic lateral sclerosis, multiple sclerosis, and Huntington's disease. Finally, we discuss the role of astrocytes in oxidative stress and neuroinflammation briefly.
Main text
Astrocyte markers in tissue
Astrocytes are identified by protein antibodies including (glial fibrillary acidic protein (GFAP), gap junction proteins such as connexin 40 and protein S100, water channel aquaporin 4 (AQP4), glutamate markers (EAAT2, EAAT1) and glutamine synthetase) in brain tissue (O'Leary and Mechawar 2021; Chierzi et al. 2023).
Astrocytes in Alzheimer's disease
Alzheimer's disease is the most common CNS disorder characterized by impairment of memory and cognitive function (Zenaro et al. 2017). Pathologically, the aggregation of beta-amyloid protein and tau protein with abnormal phosphorylation is evident in the brain tissue of patients with Alzheimer's disease (Mohammadsadeghi 2021). Alzheimer's disease is believed to occur due to the impaired relationship between neural and astrocytic functions, particularly in the hippocampus (Sapkota et al. 2022). Astrocytes clear amyloid-beta in the normal brain; however, in patients with Alzheimer's disease, impaired amyloid-beta clearance may exacerbate amyloid plaque formation (Nadkarni and Pillai 2022).
On the other hand, abnormal calcium and glutamate homeostasis in astrocytes has also been suggested to contribute to Alzheimer's disease pathogenesis. In a biophysical model of the Alzheimer's disease brain, astrocyte calcium signaling and gliotransmitter secretions can be disrupted by amyloid plaques, indicating that neuronal dysfunction can be involved in the first neurological defects of Alzheimer's disease. Nevertheless, the mechanisms that link Aβ signaling, astrocytic calcium dynamics, and gliotransmission remain unknown (Mattson and Chan 2003). But briefly, the accumulation of amyloid-β (Aβ) in the brain impairs the uptake of glutamate in astrocytes by inhibiting the glutamate-stimulated amino acid transporter 2 (EAAT2), which leads to the overactivation of glutamate receptors which causes intracellular Ca2 increase and nerve dysfunction. An increase in intracellular calcium causes an increase in the release of γ-aminobutyric acid (GABA), which, as a result, increases the absorption of GABA by neurons, disrupting memory and synaptic plasticity (Lee et al. 2022).
Excessive production of GABA by responsive astrocytes can lead to tonic inhibition of granule cells in the hippocampus. Surprisingly, inhibiting GABA synthesis restores animal models' synaptic plasticity and memory defects. The incremental elevation of GFAP is a feature of the astrocyte response in the brain of patients with Alzheimer's disease (Dejakaisaya et al. 2021; Chun et al. 2018). Also, the increase in the number of reactive astrocytes is often associated with cognitive impairment. Under normal conditions, reactive astrocytes are among the most important brain inflammation regulators; however, they can cause neurotoxicity by producing inflammatory cytokines and reactive oxygen species in response to an inflammation under pathological conditions (Markiewicz and Lukomska 2006). According to one study, IL-6, TGF, IL-1, and interferon all contribute to the formation of amyloid plaques in Alzheimer's disease. However, these cytokines significantly increase amyloid beta production in astrocytes. (Wilkaniec et al. 2018).
In addition to this, an abnormal energy metabolism has been found in the reactive astrocytes of Alzheimer's disease patients. Beta-amyloid deposition can be decreased in these astrocytes by inhibiting energy metabolism and oxidative stress. It, in turn, leads to an improvement in memory and a delay in the progression of the disease (Allaman et al. 2010).
Interestingly, brain damage has long been associated with an increased risk of Alzheimer's disease. Evidence suggests that acute brain injury can induce the expression of amyloid beta and persilinin 1 in reactive astrocytes. Nonetheless, it is still unclear whether reactive astrocytes may play a neuroprotective function at different stages of Alzheimer's disease, and further research is required in this regard (Frost and Li 2017).
Astrocytes in Parkinson's disease
This disorder is characterized by the loss of dopaminergic neurons, movement disorders, and other non-motor disorders, such as cognitive impairment and sleep disorders. The underlying molecular mechanisms in Parkinson's are unclear, though some evidence suggests the role of pro-inflammatory glial responses in the pathogenesis of Parkinson's disease (Kouli et al. 2018).
Animal studies show that intravenous injection of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) can cause Parkinson's disease in mice since the monoamine oxidase enzyme converts MPTP to 1-Methyl-4-phenylpyridinium (MPP) in the cytoplasm of astrocytes, thereby destroying dopaminergic cells (Meredith and Rademacher 2011).
Moreover, at the onset of Parkinson's disease, α-synuclein accumulates in astrocytes, which attracts phagocytic microglia. Phagocytes attack the neurons of the brain leading to the clinical symptoms of Parkinson's disease (Angelopoulou et al. 2020).
Pathological studies have shown that in the early stages of Parkinson's disease, α-synuclein activates astrocytes and leads to astrocyte aggregation; thus, increased number of astrocytes and elevated expression of GFAP expression are also other findings of the brain tissue of patients with Parkinson's disease (Gu et al. 2010).
Interestingly, increased accumulation of α-synuclein is also observed before the onset of Parkinson's symptoms, which is directly related to the expansion of reactive astrocytes. Note that the accumulation of intracellular proteins may disturb the ability of astrocytes in homeostasis and regulation of the blood–brain barrier leading to neuronal damage. These findings indicate that the interaction between astrocytes and neurons is very important in the health of dopaminergic neurons (Lieu et al. 2013; Udovin et al. 2020).
NURR1 is an orphan ligand-activated transcription factor that is essential for dopaminergic neuron growth and maintenance. The receptor NURR1 (Nuclear receptor-related factor 1), one of the major genetic risk factors for Parkinson's disease, plays an important role in controlling microglia and astrocyte neurotoxicity and exerts anti-inflammatory effects in astrocytes. Thus, selective reduction in this receptor in astrocytes leads to increased production of neurotoxic factors such as NO and ROS. Also, a glucagon-like peptide 1 receptor (GLP1R) agonist, which is a powerful neuroprotective agent in neurodegenerative diseases by stopping the microglia-driven differentiation of C3+ neurotoxic astrocytes, prevents dopaminergic neuron loss and behavioral deficits in preclinical PD mouse models. Indeed, astrocytes contribute to Parkinson's disease pathophysiology by controlling dopaminergic cell mortality via glutamate-mediated excitotoxicity, K+ buffering, and Ca2+ homeostasis. (Halliday and Stevens 2011; Al-Nusaif et al. 2022; Lee et al. 2022).Genetic mutations involved in PD are significantly expressed by α-Synuclein, Parkin, PINK1, DJ-1 FBXO7, and GBA genes in human astrocyte biology, during PD pathogenesis, and in addition, EAAT2 gene mutations and AQP4 cause severe loss of dopaminergic neurons (Hindeya Gebreyesus and Gebrehiwot Gebremichael 2020).
Astrocytes in multiple sclerosis
Multiple sclerosis (MS) is an autoimmune disease that is associated with demyelination and axon damage in the CNS (Bando 2020; Ahadiat et al. 2022a, 2022b). Despite the variable etiology of MS, a body of evidence suggests the important role of autoimmunity in the pathogenesis of this disorder. Studies have shown the increased activity of astrocytes around inflammatory lesions in patients suffering from the disease (Perriard et al. 2015). The researchers found that mice whose activated astrocytes lack MHC activator were less prone to MS by inhibiting the activity of CD4 T helper cells because CD4 T helper activity is inhibited. An inflammatory response in the CNS of MS patients is usually initiated by impaired BBB permeability and immune cell attacks, leading to the activation of astrocytes due to proinflammatory cytokines and pathogen-associated molecular pattern molecules (PAMPs) (Stüve et al. 2002). Reactive astrocytes, the main source of cytokines, such as macrophage inflammatory proteins (MIP-1α) and IL-22, IL-17A, and IL-10, do harm by uncontrolled inflammatory reactions and destroy the blood–brain barrier. IL-10, produced by regulatory T cells, limits the activity of astrocytes and reduces neuroinflammation (Blaževski et al. 2013). Furthermore, GM-CSF (Granulocyte–macrophage colony-stimulating factor) is an important cytokine in the pathogenic activity of T cells; GM-CSF signaling also limits the anti-inflammatory functions of astrocytes by reducing the expression of TRAIL (Tumor necrosis factor-related apoptosis-inducing ligand) (Lee et al. 2022). These findings indicate the vital role of this cytokine in the regulation of the balance of pro-inflammatory and anti-inflammatory astrocyte subsets in MS and other neurological disorders (Aharoni et al. 2021).
Astrocytes in amyotrophic lateral sclerosis (ALS)
ALS is an incurable, progressive motor neuron disorder characterized by the degeneration of motor neurons in the CNS, which eventually leads to muscle atrophy and respiratory failure (Cicardi et al. 2021; Ahadiat and Hosseinian 2022a, b). Even though the pathology of ALS is still being studied, existing evidence suggests the accumulation of the superoxide dismutase enzyme in neurons and astrocytes in this condition (Van Harten et al. 2021).
An experimental animal study showed that transplantation of astrocytes with superoxide dismutase 1 mutation into the spinal cord might lead to the degeneration of the motor neurons. Research has shown that reactive astrocytes are found in susceptible areas, and their receptive response depends on the extent of nerve damage in these patients, while on the other hand, these astrocytes are probably involved in the degeneration of motor neurons (Brandebura et al. 2023).
Reactive astrocytes are associated with various abnormalities in signaling pathways, including alterations in the neuronal expression of the Glutamate Receptor Ionotropic (GluR2) subunit of α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors (They play an important role in excitatory glutamatergic neurotransmission), defects in lactate transportation and activation of the P75 receptor signaling pathway in motor neurons, decreased expression of Glutamate transporter (GLT-I), Ca + release, and apoptosis due to metabotropic glutamate receptor (mGluR)-mediated glutamate signaling (Momeni et al. 2021; Baloh et al. 2022).
One of the most important functions of astrocytes is to control and reduce the extracellular concentration of glutamate in the synapse, primarily through the glutamate transporter EAAT2.
According to studies, astrocytes in ALS diseases lose the ability to control the extracellular concentration of glutamate in synapses due to the loss of the EAAT2 transporter, as well as their mitochondrial function is impaired and lactate production and calcium homeostasis in ALS astrocytes are reduced (Rosenblum and Trotti 2017). Astrocytes as well secrete neurotrophic factors such as glial-derived neurotrophic factor (GDNF), brain-derived neurotrophic factor (BDNF), and ciliary neurotrophic factor (CNTF) (CNF). Increasing their levels at ALS causes more neuroprotection. In a study, the change of the internal mechanisms controlling the production of astrocyte GDNF in the treatment of ALS was investigated. According to this study, TNFR1 receptor (TNF receptor superfamily 1) signaling is a major driver of GDNF synthesis in astrocytes, and TNFa/TNFR1 signaling is required to generate the neurotrophic factor in the central nervous system. The TNFR1-GDNF pathway may be a novel therapeutic strategy for ALS, as deletion of this receptor caused further neuronal damage (Brambilla et al. 2016; Yamanaka and Komine 2018).
Astrocytes in Huntington's disease (HD)
Huntington's disease is a progressive, lethal neurological disorder caused by mutations in the HD gene and CAG repeat. The presence of mutated HD genes in astrocytes is detrimental to neural health (Petkau et al.2019a, b). Characteristics of Huntington's disease include the accumulation of mHTT (Mutant HTT protein) in the brain, leading to astrocyte dysfunction and neurodegeneration through impaired glutamate uptake, K+ homeostasis, Ca2+ signaling, and metabolic reprogramming. Interestingly, mutations in the huntingtin gene of astrocytes cause age-related neurological disorders (Bezprozvanny and Hayden 2004; Lee et al. 2022).
A single study showed that non-mHTT-expressing microglia were found to have increased expression and transcriptional activity of PU1 and CCAT-binding protein/enhancer compared to mHTT-expressing microglia. They postulated that this leads to an increase in the transcription of basal pro-inflammatory genes; in turn, the increased levels of pro-inflammatory cytokines can perpetuate further inflammation and tissue damage (Fig. 1) (Palpagama et al.2019).

The contribution of microglia and astrocytes in Huntington's disease (HD). Various stimulatory molecules stimulate surveillance microglias to proliferate through NFkB signaling, which promotes the upregulation of PU1 (as a transcription factor that regulates hematopoiesis and macrophage differentiation) and CCAT binding. Several molecular processes can cause the death of neurons when activated microglia and reactive astrocytes produce reactive oxygen species (ROS) and neurotoxic molecules (such as quinolinic acid). Note that stimulatory molecules induce reactive astrogliosis, which results in the upregulation of pro-inflammatory cytokine production, glutamate excitotoxicity, and hyperexcitability of neurons. Image created in BioRender.com
A recent study demonstrated that mHTT-expressing microglia show significant elevations in NF-κB when stimulated with IL-6. As a result, it is suggested that mHTT may alter immune responses in microglia in a stimuli-dependent manner (Erta et al. 2012).
A recent study has questioned the extent to which these microglial changes affect HD pathology. This study showed that mHTT depletion in microglia did not significantly improve behavioral performance. In contrast, mHTT depletion in all other cells except microglia resulted in behavioral recovery. Nevertheless, even though mHTT-induced changes in microglia may not be sufficient to induce the HD phenotype, they may contribute to the pathology and other brain changes (Petkau et al. 2019a, b).
Simultaneous changes caused by the huntingtin gene in neurons and astrocytes also cause more severe neurological symptoms and premature death. There is also evidence pointing to the role of the astrocyte junction in HD, as an increase in connexin expression has been observed in the brains of patients with HD (Sánchez et al. 2020).
Similar to other neurological disorders, astrocytic inflammatory signaling pathways can be involved in the pathogenesis of HD. HD symptoms are associated with defects in ascorbic acid transport between astrocytes and neurons in clinical models. Ascorbic acid, for example, is a key antioxidant molecule in the brain that is highly concentrated in the brain, and extracellular ascorbic acid concentrations vary according to brain activity. In nerve cells, ascorbic acid can inhibit glucose consumption. As a result, the concentration of pyruvate and protons decreases, which is associated with an increase in the (nicotinamide adenine dinucleotide (NAD) + hydrogen (NAD + / NADH ratio), and Lactate dehydrogenase (LDH-1) can catalyze lactate oxidation. In fact, ascorbic acid stimulates lactate uptake in cortical neurons.
Glutamate can cause ascorbic acid efflux from astrocytes and neurons that express SVCT2. When neuronal ascorbic acid uptake via SVCT2 is impaired resulting in neurometabolic failure and neurodegeneration in HD. When extracellular ascorbic acid concentrations increase in experiments using immortalized striatal neurons expressing STHdhQ cells (represent a widely used cell culture model for studying cellular and molecular aspects of HD), SVCT2 fails to translocate to the plasma membrane, resulting in impaired ascorbic acid uptake in HD neurons.
As a result, modulating neuronal metabolism is independent of this molecule's antioxidant properties, and changes in ascorbic acid concentrations in these tissues can cause metabolic changes in the cells of various tissues (Acuña et al. 2015; Salazar et al.2021).
Furthermore, cannabinoid receptors on the surface of astrocytes play a role in modulating inflammation associated with HD by increasing signaling of this receptor by decreasing TNF-alpha, thereby alleviating inflammation in HD (Kim et al. 2022).
Astrocytes in oxidative stress
In recent years, as research has progressed, cumulative evidence has emerged demonstrating the critical role of astrocytes in the regulation of oxidative stress levels in the CNS (Ongnok et al.2021; Ahadiat et al. 2022a, 2022b). The antioxidant response in astrocytes protects the CNS from oxidative stress damage through scavenging free radicals produced by neurons and other CNS cells (Ahadiat and Hosseinian 2022a, b; Shandilya et al. 2022; Ahadiat and Hosseinian 2023). On the other hand, astrocytes can act as one of the main sources of reactive oxygen species (ROS) and reactive nitrogen species (RNS) under certain pathological conditions, and these excess free radicals can promote microglia activation or directly damage the neural tissue (Lee et al. 2021). Glutamate neurotransmitter is one of the major excitatory neurotransmitters in the CNS, released primarily by excitatory neurons that deliver excitatory signals. However, excess glutamate in synaptic clefts can contribute to calcium overload by overactivation of N-methyl-D-aspartate (NMDA) receptors or AMPA receptors, or kainic acid (KA) receptors, which produce large amounts of ROS (Parsons et al.2007; Di Meo 2016; Peng et al.2019).
The Nrf2keap1ARE pathway is an important endogenous antioxidant system in the CNS. Some researchers found that endogenous H2O2, produced in the astrocytes under certain conditions, can be neuroprotective against oxidative stress by activating the Nrf2 protein, which in turn stimulates the antioxidant stress response (Wang et al. 2022) (Fig. 2).

In a situation with high levels of reactive oxygen species (ROS), the transcription factor Nrf2 is released from binding to the protein Keap1, transported to the nucleus, and then binds to ARE gene sequences which allow for the activation of antioxidant genes such as HO-1 and NQO1. Image created in BioRender.com.
Abundant evidence suggests that astrocytes have a greater capacity to produce and store GSH than neurons, and they optimally protect neurons from oxidative damage by producing GSH (boosting Sxc expression) in the pericellular environment. Considering the increased GSH synthesis in astrocytes, some antioxidants such as Mirapexin, N-acetylcysteine, and Zonegran are useful in treating CNS degenerative diseases (Asanuma and Miyazaki 2021; Chen et al. 2020).
It has been reported that the pathogenesis of various neurological disorders, including stroke, trauma, infection, and neurodegenerative diseases, is intimately linked to redox homeostasis in astrocytes. Similarly, reactive astrogliosis can develop due to excess free radicals in the CNS, leading to inflammation and the formation of glial scar tissue (Ranjit et al. 2018). The cumulative evidence suggesting that astrocytes can modulate oxidative stress to some degree might be useful in future therapies for oxidative stress-related neurodegenerative diseases.
Astrocytes in neuroinflammation
Neuroinflammation is the brain's response to infection, disease, and destructive injury. Two groups of immune cells are involved in neuroinflammatory responses; the first group includes lymphocytes, monocytes, and macrophages in the immune system, and the second group includes microglia and astrocytes in the CNS (Lee et al. 2012). Mature astrocytes are activated during neuroinflammation-induced damage to the CNS. Astrocyte activity is believed to be vital to stabilize and continuing immune responses, rebuilding the blood–brain barrier, and reducing neuronal death.
Initially, neuroinflammation is accompanied by astrocytic hypertrophy, followed by the formation of two types of reactive astrocytes, known as A1 and A2. The A1 astrocytes induced by neuroinflammation release neurotoxins that cause rapid death of neurons and oligodendrocytes. On the other hand, the ischemia-induced A2 astrocytes promote neuronal survival and tissue regeneration. Following nervous system damage, activation of Toll-like receptors (TLR2-TLR4 and TLR3) receptors on astrocytes leads to the secretion of pro-inflammatory cytokines. (Carson et al. 2006; Ransohoff et al. 2015; Giovannon and Quintana 2020) The A1 reactive astrocytes can produce large amounts of pro-inflammatory factors and neurotoxins, which can lead to neuronal death. Activated A2 astrocytes regulate anti-inflammatory factors and many neurotrophic factors that promote neuron survival and growth (Fig. 3; Table 1) (Li et al. 2019a, 2019b, 2021).

Changes in morphology and function of astrocytes after neuroinflammation. A section: Based on the phenotypic changes of astrocytes, astrocytes are converted into reactive astrocytes, and their function is changed. Functional changes: (1) Atrophy with loss of function; (2) Pathological reconstruction; and (3) Reactive astrogliosis. B, E section: Activated astrocytes can be classified as A1 and A2 astrocytes. Astrocyte phenotypic differentiation may result in either a pro-inflammatory phenotype (A1) contributing to inflammation and neuronal death or an anti-inflammatory phenotype (A2) promoting neuronal survival and functional recovery. C, D section: These stimuli will likely promote divergent microglia phenotypes, such as the pro-inflammatory phenotype M1, which can have neurotoxic effects. M1 microglia secrete several pro-inflammatory cytokines such as IL-1β, IL-6, IFN-γ, TNF-α, complement proteins, inducible nitric oxide synthase (iNOS), and reactive oxygen species (ROS). Anti-inflammatory M2 Microglia is believed to be neuroprotective in nature. They secrete several cytokines and growth factors, such as IL-10, TGF-β, brain-derived neurotrophic factor (BDNF), and arginase-1 (Arg-1). Image created in BioRender.com.
Evidence suggests that A1-neuroinflammatory astrocytes are stimulated by NFk-B signaling, while A2-proliferative astrocytes are stimulated by STAT3-mediated signaling, with TNF-alpha and IL-1 converting quiescent astrocytes to type A2 (Khodadadei et al. 2022). TNF-alfa and IL-1 can modulate the glial scaring process in the brain by stimulating IL-6 secretion from astrocytes; they can also modulate scar formation by activating the STAT3 signaling pathway in A2-reactive astrocytes In other words, tumor necrosis factor-alpha (TNF) and IL-6 induces an inflammatory response in astrocytes via activated STAT3 signaling. Therefore, the modulation of IL-6 in inflammation may play an important role in disease management (Tanabe et al. 2010; Matsumoto et al. 2018).
In neurodegenerative diseases, the accumulating proteins can activate receptors for advanced glycation end products (RAGE2) receptors; this stimulation produces various pro-inflammatory factors in astrocytes (Derk et al. 2018). Cytokines altering the astrocyte biology and anti-inflammatory behavior, including IL-17, IL-10, IL-6, IL-1, and -INF, are expressed by T lymphocytes, such as Treg, Th17, Th1, and TRL, as well as monocytes, macrophages, and resident microglia.
The secreted chemokines into the blood by astrocytes disrupt the blood–brain barrier and attract different groups of immune cells to the inflammation site; in addition to chemokines, expression of the nitric oxide synthase enzyme is increased during inflammation leading to NO production in the astrocytes. The produced NO can also induce inflammation in the CNS by inhibiting Stromal cell-derived factor 1 (CXCL12-Protein Coding gene) (Rothhammer and Quintana 2015).
Some studies have pointed to the opposing roles of astrocytes in inflammation. At the onset of injury, gliosis limits neuronal damage and inflammation. On the other hand, some studies have also shown that reducing astrocytes in the chronic stage of MS disease has ameliorating effects by reducing leukocyte infiltration in the neural tissue (Brambilla 2019). Other studies have shown that astrocytes derived from induced pluripotent stem cells reduced the extent of inflammation and damage in epilepsy (Sajadian et al. 2014; Alayli et al. 2023).
Hence, astrogliosis is regulated in specific and time-dependent ways; the stimuli convert beneficial astrocytes to harmful astrocytes during neuroinflammation (Pamies et al. 2021).
The destruction of structural proteins also directly affects glial inflammatory signaling. Therefore, in developing treatment methods for inflammatory neurodegenerative diseases, the identification and selective targeting of astrocyte functions should be considered (Valori et al. 2021; Mohamed et al. 2023).
Thus, it is clear that astrocytes are somehow regulators of tissue inflammation since they can increase or reduce nerve damage, demyelination, and inflammation depending on the nature of the stimuli present in the inflammatory environment (Nair et al. 2008).
Conclusions
A review of the studies showed that the activity of astrocytes could have favorable or unfavorable effects through various mechanisms.
Astrocytes play an important role in normal CNS function. They play a role in potassium ion buffering, glutamate clearance, the brain's antioxidant defenses, and the modulation of neuronal excitability. In several pathological conditions, such as AD, PD, ALS, and ME, astrocytes play an active role in both the exacerbation of damage and the neuroprotective mechanisms. The balance between proinflammatory and neuroprotective glial cells may be critical in the progression of neurodegenerative diseases. Considering the complexity of microglia and astrocyte phenotypes and the different types of action of drugs and stages of neurodegenerative diseases (proinflammatory to neuroprotective) and with all the studies mentioned above, how to regulate the function of active astrocytes and their development to obtain their effective properties may become an important point in increasing further understanding in the treatment of neurological diseases.
Some strategies that target astrocytes can be intrathecal poly(lactic-co-glycolic acid) nanoparticles and Miglustat (Zavesca) and Venglustat (Ibiglustat) may be a useful tool to modulate astrocytic responses (Wang et al. 2008). Another metabolism-based approach to modulate astrocytic responses could be to target mitochondrial dysfunction. For example, in HD, low brain glucose levels induce metabolic reprogramming of striatal astrocytes, increasing ROS production and neurotoxicity. XJB-5-131, a mitochondria-targeted antioxidant, limits astrocyte-mediated ROS-induced neuronal damage (Polyzos et al. 2016). Glutamate transporters EAAT1 and EAAT2, which are expressed by glial cells (mainly astrocytes), are important for neuronal survival and function in AD, HD, and PD. For example, treatment with the antibiotic ceftriaxone can increase EAAT2 expression in astrocytes and in the result identifies EAAT2 as a promising drug target for various neurodegenerative disorders (Rothstein et al. 2005).
Availability of data and materials
Not applicable.
Abbreviations
- AMPA:
-
α-Amino-3-hydroxy-5-methyl-4-isoxazole propionic acid
- ARE:
-
Antioxidant response element
- CXCL12:
-
Stromal cell-derived factor 1
- EAAT:
-
Excitatory amino acid transporter 1
- Fzd1:
-
Frizzled1
- GABA:
-
Gamma-aminobutyric acid
- GFAP:
-
Glial fibrillary acidic protein
- GluR2:
-
Glutamate receptor ionotropic
- GLT-1:
-
Glutamate transporter
- GM-CSF:
-
Granulocyte–macrophage colony-stimulating factor
- MGluRs:
-
Metabotropic glutamate receptors
- MIP-1 alpha:
-
Macrophage inflammatory proteins-1 alpha
- MPTP:
-
1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine
- MPP:
-
1-Methyl-4-phenylpyridinium
- NAD-H:
-
Nicotinamide adenine dinucleotide-hydrogen
- NURR1:
-
Nuclear receptor-related factor 1
- PAMPs:
-
Pathogen-associated molecular pattern molecules
- p-Nrf2:
-
Phosphorylated Nrf2
- RAGE2:
-
Receptor for advanced glycation end products
- RNS:
-
Reactive nitrogen species
- ROS:
-
Reactive oxygen species
- STAT3:
-
Signal transducer and activator of transcription 3
- SVCT2:
-
Sodium-dependent vitamin C transporter 2
- TGF:
-
Transforming growth factor
- TNFR1:
-
TNF receptor superfamily 1
- TRAIL:
-
Tumor necrosis factor-related apoptosis-inducing ligand
- Wnt1:
-
Wingless-type MMTV integration site
References
Acuña AI, Beltrán A, Castro MA (2015) Old things new view: ascorbic acid protects the brain in neurodegenerative disorders. Int J Mol Sci 16(12):28194–28217
Ahadiat SA, Hosseinian Z (2022a) The role of biomaterials in degenerative diseases. JSKUMS 24(4):153–154
Ahadiat SA, Hosseinian Z (2022b) The role of biomaterials in treating Lou Gehrig’s disease. Arch Neurosci 9(4):e131625
Ahadiat SA, Hosseinian Z (2023) Diabetes ketoacidosis and COVID-19: an insight into the pathophysiology. Int J Endocrinol 21(2)
Ahadiat SA, Masoud SA, Khorshidi MS, Hosseinian Z, Namvari A (2022a) Helicobacter Pylori specific antibodies among multiple sclerosis patients. Neurol Lett 1(1):28–31
Ahadiat SA, Barati R, Moghadam NS, Samami E, Ghiabi S, Alyari M et al (2022b) Role of oxidative stress and antioxidants in malignancies. Kindle 2(1):1–122
Aharoni R, Eilam R, Arnon R (2021) Astrocytes in multiple sclerosis—essential constituents with diverse multifaceted functions. Int J Mol Sci 22(11):5904
Alayli A, Lockard G, Gordon J, Connolly J, Monsour M, Schimmel S (2023) Stem cells: recent developments redefining epilepsy therapy. Cell Transpl 32:09636897231158967
Allaman I, Gavillet M, Bélanger M, Laroche T, Viertl D, Lashuel HA, Magistretti PJ (2010) Amyloid-β aggregates cause alterations of astrocytic metabolic phenotype: impact on neuronal viability. J Neurosci 30(9):3326–3338
Al-Nusaif M, Yang Y, Li S, Cheng C, Le W (2022) The role of NURR1 in metabolic abnormalities of Parkinson’s disease. Mol Neurodegener 17(1):1–15
Angelopoulou E, Paudel YN, Shaikh MF, Piperi C (2020) Fractalkine (CX3CL1) signaling and neuroinflammation in Parkinson’s disease: potential clinical and therapeutic implications. Pharmacol Res 158:104–930
Asanuma M, Miyazaki I (2021) Glutathione and related molecules in Parkinsonism. Int J Mol Sci 22(16):8689
Baloh RH, Johnson JP, Avalos P, Allred P, Svendsen S, Gowing G, Svendsen CN (2022) Transplantation of human neural progenitor cells secreting GDNF into the spinal cord of patients with ALS: a phase 1/2a trial. Nat Med 289:1813–1822
Bando Y (2020) Mechanism of demyelination and remyelination in multiple sclerosis. Clin Exp Neuroimmunol 11:14–21
Bezprozvanny I, Hayden MR (2004) Deranged neuronal calcium signaling and Huntington disease. Biochem Biophys Res Commun 322(4):1310–1317
Blaževski J, Petković F, Momčilović M, Jevtic B, Miljković D, Stojković MM (2013) High interleukin-10 expression within the central nervous system may be important for initiation of recovery of Dark Agouti rats from experimental autoimmune encephalomyelitis. Immunobiology 218(9):1192–1199
Bolton MM, Eroglu C (2009) Look who is weaving the neural web: glial control of synapse formation. Curr Opin Neurobiol 19(5):491–497
Brambilla R (2019) The contribution of astrocytes to the neuroinflammatory response in multiple sclerosis and experimental autoimmune encephalomyelitis. Acta Neuropathol 137(5):757–783
Brambilla L, Guidotti G, Martorana F, Iyer AM, Aronica E, Valori CF, Rossi D (2016) Disruption of the astrocytic TNFR1-GDNF axis accelerates motor neuron degeneration and disease progression in amyotrophic lateral sclerosis. Hum Mol Genet 25(14):3080–3095
Brandebura AN, Paumier A, Onur TS, Allen NJ (2023) Astrocyte contribution to dysfunction, risk and progression in neurodegenerative disorders. Nat Rev Neurosci 24(1):23–39
Carson MJ, Thrash JC, Walter B (2006) The cellular response in neuroinflammation: the role of leukocytes, microglia and astrocytes in neuronal death and survival. Clin Neurosci Res 65:237–245
Chen Y, Qin C, Huang J, Tang X, Liu C, Huang K et al (2020) The role of astrocytes in oxidative stress of central nervous system: a mixed blessing. Cell Prolif 53(3):e12781
Chierzi S, Kacerovsky JB, Fok AH, Lahaie S, Rosen AS, Farmer WT, Murai KK (2023) Astrocytes transplanted during early postnatal development integrate, mature, and survive long term in mouse cortex. J Neurosci 43(9):1509–1529
Chun H, An H, Lim J, Woo J, Lee J, Ryu H, Lee CJ (2018) Astrocytic proBDNF and tonic GABA distinguish active versus reactive astrocytes in hippocampusExp. Neurobiol 27(3):155
Cicardi ME, Marrone L, Azzouz M, Trotti D (2021) Proteostatic imbalance and protein spreading in amyotrophic lateral sclerosis. EMBO J 4010:106–389
Dejakaisaya H, Kwan P, Jones NC (2021) Astrocyte and glutamate involvement in the pathogenesis of epilepsy in Alzheimer’s disease. Epilepsia 627:1485–1493
Derk J, MacLean M, Juranek J, Schmidt AM (2018) The receptor for advanced glycation endproducts (RAGE) and mediation of inflammatory neurodegeneration. J Alzheimers Dis 8(1):421
Dutta DJ, Woo DH, Lee PR, Pajevic S, Bukalo O, Huffman WC et al (2018) Regulation of myelin structure and conduction velocity by perinodal astrocytes. Proc Natl Acad Sci 115(46):11832–11837
Emirandetti A, Zanon RG, Sabha M Jr, de Oliveira ALR (2006) Astrocyte reactivity influences the number of presynaptic terminals apposed to spinal motoneurons after axotomy. Brain Res 10951:35–42
Erta M, Quintana A, Hidalgo J (2012) Interleukin-6, a major cytokine in the central nervous system. Int J Biol Sci 8(9):1254
Escartin C, Galea E, Lakatos A, O’Callaghan JP, Petzold GC, Serrano-Pozo A et al (2021) Reactive astrocyte nomenclature, definitions, and future directions. Nat Neurosci 24(3):312–325
Farmer WT, Murai K (2017) Resolving astrocyte heterogeneity in the CNS. Front Cell Neurosci 11:300
Filous AR, Silver J (2016) Targeting astrocytes in CNS injury and disease: a translational research approach. Prog Neurobiol 144:173–187
Frost GR, Li YM (2017) The role of astrocytes in amyloid production and Alzheimer’s disease. Open Biol 712:170–228
Giovannoni F, Quintana FJ (2020) The role of astrocytes in CNS inflammation. Trends Immunol 41(9):805–819
Gu XL, Long CX, Sun L, Xie C, Lin X, Cai H (2010) Astrocytic expression of Parkinson’s disease-related A53T α-synuclein causes neurodegeneration in mice. Mol Brain 31:1–16
Halliday GM, Stevens CH (2011) Glia: initiators and progressors of pathology in Parkinson’s disease. Mov Disord 26(1):6–17
Hanani M, Verkhratsky A (2021) Satellite glial cells and astrocytes, a comparative review. Neurochem Res 46(10):2525–2537
Hindeya Gebreyesus H, Gebrehiwot Gebremichael T (2020) The potential role of astrocytes in Parkinson’s disease (PD). Med Sci 8(1):7
Khodadadei F, Arshad R, Morales DM, Gluski J, Marupudi NI, McAllister JP, Harris CA (2022) The effect of A1 and A2 reactive astrocyte expression on hydrocephalus shunt failure. Fluids Barriers CNS 19(1):78
Kim H, Leng K, Park J, Sorets A G, Kim S, Shostak A, et al (2022) Reactive astrocytes transduce blood-brain barrier dysfunction through a TNFα-STAT3 signaling axis and secretion of alpha 1-antichymotrypsin. bioRxiv, 2022–02
Kouli A, Torsney KM, Kuan WL (2018) Parkinson’s disease: etiology, neuropathology, and pathogenesis. Exon Publications, Brisbane City, pp 3–26
Lee DH, Gold R, Linker RA (2012) Mechanisms of oxidative damage in multiple sclerosis and neurodegenerative diseases: therapeutic modulation via fumaric acid esters. Int J Mol Sci 13(9):11783–11803
Lee KH, Cha M, Lee BH (2021) Crosstalk between neuron and glial cells in oxidative injury and neuroprotection. Int J Mol Sci 22(24):13315
Lee HG, Wheeler MA, Quintana FJ (2022) Function and therapeutic value of astrocytes in neurological diseases. Nat Rev Drug Discov 21(5):339–358
Li K, Li J, Zheng J, Qin S (2019a) Reactive astrocytes in neurodegenerative diseases. Aging Dis 10(3):664
Li T, Chen X, Zhang C, Zhang Y, Yao W (2019b) An update on reactive astrocytes in chronic pain. J Neuroinflamm 16:1–13
Li L, Acioglu C, Heary RF, Elkabes S (2021) Role of astroglial toll-like receptors (TLRs) in central nervous system infections, injury and neurodegenerative diseases. Brain Behav Immun 91:740–755
Lieu CA, Chinta SJ, Rane A, Andersen JK (2013) Age-related behavioral phenotype of an astrocytic monoamine oxidase-B transgenic mouse model of Parkinson’s disease. PLoS ONE 8(1):e54200
Liu B, Teschemacher AG, Kasparov S (2017) Neuroprotective potential of astroglia. J Neurosci Res 95(11):2126–2139
Markiewicz I, Lukomska B (2006) The role of astrocytes in the physiology and pathology of the central nervous system. Acta Neurobiol Exp 66(4):343–358
Matsumoto J, Dohgu S, Takata F, Machida T, Hatip FFB, Hatip-Al-Khatib I et al (2018) TNF-α-sensitive brain pericytes activate microglia by releasing IL-6 through cooperation between IκB-NFκB and JAK-STAT3 pathways. Brain Res 1692:34–44
Mattson MP, Chan SL (2003) Neuronal and glial calcium signaling in Alzheimer’s disease. Cell Calcium 34(4–5):385–397
Di Meo S, Reed TT, Venditti P, Victor VM (2016) Role of ROS and RNS sources in physiological and pathological conditions. Oxid Med Cell Longev
Meredith GE, Rademacher DJ (2011) MPTP mouse models of Parkinson’s disease: an update. J Parkinsons Dis 1(1):19–33
Mohamed W, Kumar J, Alghamdi BS, Soliman A, Toshihide Y (2023) Neurodegeneration and inflammation crosstalk: therapeutic targets and perspectives. IBRO Rep 14:95–110
Mohammadsadeghi H (2021) The role of astrocytes in the central nervous system: physiological and pathophysiological conditions. Neurosci J Shefaye Khatam 9(2):119–139
Molina-Gonzalez I, Miron VE (2019) Astrocytes in myelination and remyelination. Neurosci Lett 713:134532
Momeni Z, Bautista M, Neapetung J, Urban R, Yamamoto Y, Krishnan A, Campanucci VA (2021) RAGE signaling is required for AMPA receptor dysfunction in the hippocampus of hyperglycemic mice. Physiol Behav 229:113255
Nadkarni S, Pillai AG (2022) Amyloid pathology disrupts gliotransmitter release in astrocytes. PLoS Comput Biol 18(8):e1010334
Nair A, Frederick TJ, Miller SD (2008) Astrocytes in multiple sclerosis: a product of their environment. CMLS 65:2702–2720
O’Leary LA, Mechawar N (2021) Implication of cerebral astrocytes in major depression: a review of fine neuroanatomical evidence in humans. Glia 69(9):2077–2099
Ongnok B, Khuanjing T, Chunchai T, Pantiya P, Kerdphoo S, Arunsak B, Chattipakorn SC (2021) Donepezil protects against doxorubicin-induced chemobrain in rats via attenuation of inflammation and oxidative stress without interfering with doxorubicin efficacy. Neurotherapeutics 18:2107–2125
Palpagama TH, Waldvogel HJ, Faull RL, Kwakowsky A (2019) The role of microglia and astrocytes in Huntington’s disease. Front Mol Neurosci 12:258
Pamies D, Sartori C, Schvartz D, González-Ruiz V, Pellerin L, Nunes C et al (2021) Neuroinflammatory response to TNFα and IL1β cytokines is accompanied by an increase in glycolysis in human astrocytes in vitroInt. J Mol Sci 22(8):4065
Parsons CG, Stöffler A, Danysz W (2007) Memantine: a NMDA receptor antagonist that improves memory by restoration of homeostasis in the glutamatergic system-too little activation is bad, too much is even worse. Neuropharmacology 53(6):699–723
Pekny M, Pekna M, Messing A, Steinhäuser C, Lee JM, Parpura V et al (2016) Astrocytes: a central element in neurological diseases. Acta Neuropathol 131:323–345
Peng JJ, Lin SH, Liu YT, Lin HC, Li TN, Yao CK (2019) A circuit-dependent ROS feedback loop mediates glutamate excitotoxicity to sculpt the Drosophila motor system. Elife 8:e47372
Perriard G, Mathias A, Enz L, Canales M, Schluep M, Gentner M et al (2015) Interleukin-22 is increased in multiple sclerosis patients and targets astrocytes. J Neuroinflammation 12(1):1–18
Petkau TL, Hil A, Connolly C, Lu G, Wagner P, Kosior N et al (2019a) Mutant huntingtin expression in microglia is neither required nor sufficient to cause the Huntington’s disease-like phenotype in BACHD mice. Hum Mol Genet 28(10):1661–1670
Petkau TL, Hill A, Connolly C, Lu G, Wagner P, Kosior N (2019b) Mutant huntingtin expression in microglia is neither required nor sufficient to cause the Huntington’s disease-like phenotype in BACHD mice. Hum Mol Genet 28:1661–1670
Polyzos A, Holt A, Brown C, Cosme C, Wipf P, Gomez-Marin A et al (2016) Mitochondrial targeting of XJB-5-131 attenuates or improves pathophysiology in HdhQ150 animals with well-developed disease phenotypes. Hum Mol Genet 25(9):1792–1802
Ranjit S, Patters BJ, Gerth KA, Haque S, Choudhary S, Kumar S (2018) Potential neuroprotective role of astroglial exosomes against smoking-induced oxidative stress and HIV-1 replication in the central nervous system. Expert Opin Ther Pat 22(8):703–714
Ransohoff RM, Schafer D, Vincent A, Blachère NE, Bar-Or A (2015) Neuroinflammation: ways in which the immune system affects the brain. Neurotherapeutics 12:896–909
Rosenblum LT, Trotti D (2017) Trotti D (2017) EAAT2 and the molecular signature of amyotrophic lateral sclerosis. Adv Neurobiol 16:117–136
Rothhammer V, Quintana FJ (2015) Control of autoimmune CNS inflammation by astrocytes. Semin Immunopathol 37:625–638
Rothstein JD, Patel S, Regan MR, Haenggeli C, Huang YH, Bergles DE et al (2005) β-Lactam antibiotics offer neuroprotection by increasing glutamate transporter expression. Nature 433(7021):73–77
Sajadian A, Jafarian M, Khodaie B, Mohammad SS, Ghaemi A (2014) Reduction of neuroinflammation in epilepsy by using induced pluripotent stem (iPS) cells-derived astrocytes. Neurosci J Shefaye Khatam 2(2):56–64
Salazar K, Espinoza F, Cerda-Gallardo G, Ferrada L, Magdalena R, Ramírez E et al (2021) SVCT2 overexpression and ascorbic acid uptake increase cortical neuron differentiation, which is dependent on vitamin c recycling between neurons and astrocytes. Antioxidants 10(9):1413
Sánchez OF, Rodríguez AV, Velasco-España JM, Murillo LC, Sutachan JJ, Albarracin SL (2020) Role of connexins 30, 36, and 43 in brain tumors, neurodegenerative diseases, and neuroprotection. Cells 9(4):846
Sapkota D, Florian C, Doherty BM, White KM, Reardon KM, Ge X et al (2022) Aqp4 stop codon readthrough facilitates amyloid-β clearance from the brain. Brain 145(9):2982–2990
Shandilya S, Kumar S, Jha NK, Kesari KK, Ruokolainen J (2022) Interplay of gut microbiota and oxidative stress: Perspective on neurodegeneration and neuroprotection. J Adv Res 38:223–244
Stuve O, Youssef S, Slavin AJ, King CL, Patarroyo JC, Hirschberg DL et al (2002) The role of the MHC class II transactivator in class II expression and antigen presentation by astrocytes and in susceptibility to central nervous system autoimmune disease. J Immun 169(12):6720–6732
Tanabe K, Matsushima-Nishiwaki R, Yamaguchi S, Iida H, Dohi S, Kozawa O (2010) Mechanisms of tumor necrosis factor-α-induced interleukin-6 synthesis in glioma cells. J Neuroinflamm 7(1):1–8
Tang G, Yang GY (2016) Aquaporin-4: a potential therapeutic target for cerebral edemaInt. J Mol Sci 17(10):1413
Udovin L, Quarracino C, Herrera MI, Capani F, Otero-Losada M, Perez-Lloret S (2020) Role of astrocytic dysfunction in the pathogenesis of Parkinson’s disease animal models from a molecular signaling perspective. Neural Plast
Valori CF, Possenti A, Brambilla L, Rossi D (2021) Challenges and opportunities of targeting astrocytes to halt neurodegenerative disorders. Cells 10(8):2019
Van Harten AC, Phatnani H, Przedborski S (2021) Non-cell-autonomous pathogenic mechanisms in amyotrophic lateral sclerosis. TINS 44(8):658–668
Varela-Echevarría A, Vargas-Barroso V, Lozano-Flores C, Larriva-Sahd J (2017) Is there evidence for myelin modeling by astrocytes in the normal adult brain? Front Neuroanat 11:75
Verkhratsky A, Bush NAO, Nedergaard M, Butt A (2018) The special case of human astrocytes. Neuroglia 1(1):21–29
Wang YC, Wu YT, Huang HY, Lin HI, Lo L, Tzeng SF, Yang CS (2008) Sustained intraspinal delivery of neurotrophic factor encapsulated in biodegradable nanoparticles following contusive spinal cord injury. Biomater 29(34):4546–4553
Wang L, Zhang X, Xiong X, Zhu H, Chen R, Zhang S et al (2022) Nrf2 regulates oxidative stress and its role in cerebral ischemic stroke. Antioxidants 11(12):2377
Wilkaniec A, Gąssowska-Dobrowolska M, Strawski M, Adamczyk A, Czapski GA (2018) Inhibition of cyclin-dependent kinase 5 affects early neuroinflammatory signalling in murine model of amyloid beta toxicity. J Neuroinflamm 15(1):1–18
Williams SM, Sullivan RK, Scott HL, Finkelstein DI, Colditz PB, Lingwood BE et al (2005) Glial glutamate transporter expression patterns in brains from multiple mammalian species. Glia 49(4):520–541
Yamanaka K, Komine O (2018) The multi-dimensional roles of astrocytes in ALS. Neurosci Res 126:31–38
Zenaro E, Piacentino G, Constantin G (2017) The blood-brain barrier in Alzheimer’s disease. Neurobiol Dis 1(107):41–56
Zhou B, Zuo YX, Jiang RT (2019) Astrocyte morphology: diversity, plasticity, and role in neurological diseases. CNS Neurosci Ther 25(6):665–673
Acknowledgements
Not applicable.
Funding
No funding was obtained for this study.
Author information
Authors and Affiliations
Contributions
SAAA and ZH approved the manuscript topic, both initiated the search and began writing the initial manuscript, and together edited and drafted the final manuscript. All authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ahadiat, SA., Hosseinian, Z. Astrocytes’ innate role in neurodegenerative disorders. Bull Natl Res Cent 47, 105 (2023). https://doi.org/10.1186/s42269-023-01083-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42269-023-01083-0