
Abstract
Background
Biodiesel is a long chain of fatty acid alkyl esters derived from renewable feedstock and it was used as a partial or full substitute for petrol diesel.
Results
Among 36 Egyptian filamentous fungi that were screened for their abilities to produce lipid by using Nile red staining assay, Fusarium oxysporum NRC2017 was selected and identified based on the molecular method. Optimization of culture conditions for F. oxysporum NRC2017 maximum lipid production were investigated by using a basal liquid medium with initial pH 5.0, incubation temperature 30 °C, and after 9 days of incubation at a static condition. Six types of agro-industrial by-products were used as components of semi-solid-state fermentation, namely breaking rice, rice straw, wheat bran, corn stalk, sweet sorghum, and bagasse. The highest lipid accumulation was obtained when baggase 98.3 mg/g was used. Gas chromatography (GC) analysis of F. oxysporum NRC 2017 biodiesel showed the presence of 2.2% palmitoleic acid, 11.1% oleic acid, 5.6% linoleic acid, 3.0% α-linoleic acid, 2.5% eicosatrienoic acid, 6.8% eicosapentaenoic acid, 16.1% erucic acid, 4.2% margaric acid, and 3.9% behenic acid. Biodiesel from F. oxysporum NRC2017 was blended with petroleum diesel in 5% “B5” and the physical properties results were density 0.84 g/ml, viscosity 2.55 mm2/s, cloud point 0 °C, iodine value 92.71 gI2/100 g, acid value 1.6 mg/g, and pour point − 1.3 °C.
Conclusion
This work revealed the possibility of biodiesel production from the Egyptian promising fungal strain F. oxysporum NRC2017.
Similar content being viewed by others

Explore related subjects
Find the latest articles, discoveries, and news in related topics.Background
Biodiesel was a biofuel produced from renewable sources, like vegetable oil or animal fat by the triglycerides transesterification to produce fatty acid alkyl esters (Gunston 2001; Rottig et al. 2010). Biodiesel was used as a partial or full substitute for petrol diesel in unmodified combustion engines used for power generation as well as engines for road and waterway transport (Gavrilescu and Chisti 2005). Biodiesel fuels were increasingly attracting worldwide attention due to the environmental benefits such as biodegradability; a decrease of sulfur and aromatic hydrocarbons content, which reduced their emission during fuel combustion and lower emission of CO, CO2, and particulate materials (Demirbas 2008; Kotasthan 2017). The main material sources for biodiesel production were vegetable oils and animal fats (Hanna 1991). Animal fat origin often contained large free fatty acids amounts, which made its conversion into biodiesel of good quality very difficult (Canakci and Van 2001; Demirbas 2003). Concerns had been expressed about the competition of vegetable oil with food supply by the use of arable land (Brennan and Owende 2010). The use of microorganisms which could be easily cultivated and yield large lipid amounts had been regarded as one of the most promising solutions to solve the negative aspects of the production of biodiesel from animal or agricultural sources (Tao et al. 2006). Lipids accumulated in microorganisms like fungi, algae, and bacteria are mainly in the triacylglycerols (TAG) form utilized as metabolites for energy storage. Most of these lipids which were composed of long chain fatty acids were similar to conventional vegetable oils (Rude and Schirmer 2009). Compared with animal fats and vegetable oils, the microbial oil production had many advantages, such as: (1) fasting growing, (2) recovery of cells to extract oil required less work than the harvesting of oleaginous plants, (3) no influence of environmental conditions in lipid accumulation when microbial growth was performed in closed systems (bioreactors), and (4) easy growth in large scale (Li et al. 2008). This study concerned with studying oleaginous fungal strains isolated, screened, identified, and optimized the culture conditions for biodiesel production from Fusarium oxysporum NRC2017, and at the same time, the harmful effects of fossil fuel in the environment will be reduced.
Methods
Samples collection and fungi isolation
Five soil samples were collected from several districts in Egypt. The soil serial dilution methodology was employed to isolate oleaginousfungi on basal medium according to Li et al. (2011). The typical formula was (g/l): xylose 100.0, yeast extract 1.0, KH2PO4 2.0, MgSO4.7H2O 0.75, Na2HPO4 1.0, CaCl2.2H2O 0.2, FeCl3 0.01, and ZnCl2 0.1; then, the pH was adjusted at 7.0 (± 0.1), 10 ml/l of Rose Bengal (4, 5, 6-Tetrachlorofluorescein) was added at concentration 50 mg/l.
Screening for oleaginous fungi isolates by using Nile red stain
The mycelia were mixed with 0.5 ml phosphate buffer solution (PBS) solution and 0.05 ml Nile red solution at concentration 10 mg/1000 ml acetone under aseptic condition (Greenspan et al. 1985).The Nile red was used for staining intracellular lipid droplets after the storage of the mixture of Nile red and mycelia for 30 min in a dark place (Lim et al. 2003), stained lipid bodies were photographed using fluorescence microscope (HBO50/ac Axiostar plus, Zeiss, Jena, Germany) equipped with a CCD camera (105 colar Axiocam, Zeiss, Jena, Germany).
Molecular identification of the promising fungal isolate
The molecular identification of the promising fungal isolate was carried out by Lab Technology Company including the preparation of the fungal biomass, then extraction of fungal genomic DNA, and finally PCR amplification for fungal DNA.
Preparation of the fungal biomass
The spores of 4-day-old culture of tested fungi were collected by addition 5 ml of sterile saline solution into the fungal slant and the obtained suspension was inoculated into 100 ml of Czapek Dox’s broth in a 250-ml Erlenmeyer flask. After incubation at 30 °C for 4 days, the culture fungal growth was filtered and the mats biomass were collected and washed three times with distilled water.
Genomic DNA extraction
Genomic DNA of fungi was extracted as the follows; 50 to 100 mg fresh fungal mycelia were homogenized by using a mortar. Five hundred microliters of DNA extraction buffer (200 mM Tris-HCl pH 8, 240 mM NaCl, 25 mM ethylene diamine tetra acetic acid (EDTA), and 1%Sodium dodecyl sulfate (SDS)) were then added to the homogenized fungal materials. One volume of phenol/chloroform, at a ratio of 1:1 (v/v), was added to a mixture and mixed gently for 10 min on a shaker at 150 rpm followed by centrifugation at 38324 rpm for 10 min. The upper phase was transferred to a new tube and 0.1 ml of 3 M sodium-acetate buffer at pH 5.2 and 2 ml of ethanol (96%) were added and mixed well, incubated for 30 min at − 20 °C followed by centrifugation (38,324 rpm /4 °C/20 min). The resulting pellets were washed 3 times with 700 μl of 70% ethanol; air dried and re-suspended in 100 μl of sterile bi-distilled water; then, the PCR amplification for DNA pellets was carried out as follows.
PCR amplification
The primers ITS1 5′-TCCGTAGGTGAACCTGCGG-3′ and ITS4 5′-TCCTCCGCTTAT TGATATGC-3′ were used for the PCR. The PCR reaction was performed with 20 ng of genomic DNA as the template in a 30-μl reaction mixture by using an EF-Taq (Sol Gent, Korea) as follows: Activation of Taq polymerase at 95 °C for 2 min, 35 cycles of 95 °C for 1 min, 55 °C and 72 °C for 1 min each were performed, finishing with a 10 min step at 72 °C. The amplification products were purified with a multiscreen filter plate (Millipore Corp., Bedford, MA, USA). Sequencing reaction was performed using a PRISM Big Dye Terminator v3.1 Cycle sequencing Kit. The DNA samples containing the extension products were added to Hi-Di formamide (Applied Biosystems, Foster City, CA). The mixture was incubated at 95 °C for 5 min, followed by 5 min on ice and then analyzed by ABI Prism 3730XL DNA analyzer (Applied Biosystems, Foster City, CA).
Optimization of the culture conditions for the best fungal lipid production
Effect of nutritional and environmental parameters on lipid production was carried out. Basal medium was used for isolation and purification of oleaginous fungi (Sergeeva et al. 2008). The typical formula was (g/l): glucose5.0, yeast extract 2.0, KH2PO4 0.05, K2HPO4 0.05, MgSO4.7H2O 0.05, MnSO4 0.1, CaCl2 10, FeCl3 0.2, asparagine 0.05, leucine 0.05, glycine 0.05, and glutamic acid 0.05; then, the pH was adjusted at 7.0 ± 0.1 The prepared inoculum size of Fusarium oxysporum NRC2017 was 6.1 × 105 spores/ml for all the following experiments.
Effect of initial medium pH on fungal lipid production
This experiment was carried out to evaluate the effect of initial pH medium on the lipid production of F. oxysporum NRC2017. The pH of the culture medium was adjusted with 1 N HCl or 1 N KOH after autoclaving at different values (4.0, 4.5, 5.0, 5.5, 6.0, 7.0, and 8.0).
Effect of incubation temperature on fungal lipid production
This experiment was carried out to evaluate the effect of different incubation temperatures on lipid production of F. oxysporum NRC2017 at different temperatures 20, 25, 30, 35, and 40 °C.
Effect of incubation period on fungal lipid production
This experiment was carried out to evaluate the effect of different incubation periods on the lipid production of F. oxysporum NRC2017 at different periods 1, 2, 3, 5, 7, 9, and 11 days.
Effect of inoculum size on fungal lipid production
This experiment was carried out to evaluate the effect of inoculum size on the lipid production of F. oxysporum NRC2017. Spore suspension inoculation size of the culture medium was adjusted at different values 0.50, 0.75, 1.00, 1.25, and 1.50 ml in 100 ml of liquid production medium.
Effect of shaking and static conditions on fungal lipid production
This experiment was carried out to evaluate the effect of shaking at 80 rpm and static condition on the lipid production of F. oxysporum NRC2017.
Effect of different carbon sources on fungal lipid production
This experiment was carried out to evaluate the effect of different carbon sources on the lipid production F. oxysporum NRC2017. An equimolar amount of five different carbon sources were used as follows: xylose, glucose, sucrose, lactose, and raffinose.
Effect of different nitrogen sources on fungal lipid production
This experiment was carried out to evaluate the effect of different nitrogen sources on the lipid production of F. oxysporum NRC2017. An equimolar amount of six different nitrogen sources were used as follows: yeast extract, peptone, casein, ammonium sulphate, sodium nitrate, and ammonium nitrate were individually added to the production medium.
Effect of agro-industrial by-products as feedstock for fungal lipid production
Six agro-industrial by-products represent only a component of semi-solid fermentation medium such as breaking rice, wheat bran, rice straw, corn stalk, sweet sorghum, and bagasse tested by F. oxysporum NRC2017 as semi-solid fermentation media for lipid production.
Lipid extraction and determination
The optimum conditions increased the amount of lipid in F. oxysporum NRC2017. Indirect transesterification procedure by initial extraction of lipids from biomass followed by transesterification was carried out (Vicente et al. 2010). The common lipid extraction method, the culture biomass was collected by using Sigma 3–18 KS centrifuge (5000 rpm/4 °C/10 min) then washed 3 times with distilled water to remove the medium residues. The mycelia were dried in an oven at 60 °C until a constant dry weight then the lipid in the biomass was estimated by lipid extraction and determination (Bligh and Dyer 1959; Lewis et al. 2000).
Extraction of lipid
Lipid was extracted from the fungal biomass with a solvent mixture of chloroform and methanol (2:1 v/v); 200 mg of the dried biomass washed three times with 10 ml of the solvent mixture for 10 min each together with ultrasonication to favor cell membrane disruption (Folch et al. 1957; Vicente et al. 2009). The solvent mixture containing extracted lipids was separated from residual biomass by centrifugation using Sigma 3-18 KS centrifuge (5000 rpm/4 °C/10 min) and all the fractions from each stage were collected and the solvent was evaporated.
Determination of lipid
Lipid was determined according to Mishra et al. (2014) using sulfo-phospho-vanillin (SPV) reagent. It was prepared by initially dissolving 0.6 g vanillin in 10 ml absolute ethanol and 90 ml deionized water and stirred continuously. Subsequently, 400 ml of concentrated H3PO4 was added to the mixture, and the resulting reagent was stored in a dark bottle. To ensure high activity, the freshly prepared phospho-vanillin reagent was recommended. For SPV reaction of the fungal lipid quantification, a known amount of fungal lipid was used for routine assay, 2.0 ml of concentrated H2SO4 (98%) was added to the fungal lipid sample and heated for 10 min at 100 °C, then cooled for 5 min in an ice bath. Five milliliters of freshly prepared SPV reagent was added, and the sample was incubated for 15 min at 37 °C in an incubator shaker (200 rpm), and the absorbency was measured at 530 nm by using JASCO V-630 spectrophotometer in order to quantify the lipid concentration within the fungal lipid samples.
Biodiesel production
Biodiesel production according to Vicente et al. (2009) experiments was planned to use HCl as an acid catalyst and the reaction temperature 25 °C. Reactions of extracted microbial lipids were performed in 15-ml glass closed vessels with magnetic stirring (900 rpm) using a methanol to oil ratio of 60:1 M and a catalyst concentration of 8 wt% relative to microbial oil. The reactor was then immersed in a thermostatic bath at the reaction temperature for 8 h. The fatty acid methyl esters (FAMEs) layer was collected and the crude glycerol was washed five times with petroleum ether: diethyl ether (80:20%) as well as the same volume of water. The upper organic layers were put together with the first FAME layer and the solvent was removed on a rotary evaporator leaving the residue containing the FAMEs, which was used in measuring the reaction yield relative to the dry microbial biomass.
Gas chromatographic analysis
Gas chromatographic analysis for of fatty acids methyl esters (FAMEs) for the high producer isolate F. oxysporum NRC2017 was performed in Central Laboratories in National Research Centre (NRC) using Agilent Technologies 6890 N (Net Work GC system) the USA. The oven was held at initial temperature 50 °C and maintained for 2 min. At rate 10, 8, 5, 6 °C/min, raised to 70, 170, 200, and 240 °C, at the rate of 2, 9, 5, 15 min and run time 60.17 min. Injector temperature was held at 250 °C splitless. A capillary column HP-5MS (5% phenyl methyl siloxane) has a dimension of 30 m × 320 μm, film thickness 0.25 μm. The flame ionization detector temperature was 280 °C and the flow rate was 1.5 ml/min. The carrier gas was nitrogen, with a flow rate of 30 ml/min. Hydrogen flow rate was 30 ml/min and air flow rate 300 ml/min.
Physical properties of biodiesel
The physical properties analysis for fatty acid methyl esters (FAMEs) were performed at the Engineering development and consulting unit in National Research Centre (NRC). FAMEs were blended with petroleum diesel at concentration 5% and determined density was (ASTM D1298), the viscosity was ASTM D445, cloud point was ASTM D2500, pour point was ASTM D97, iodine value was ASTM D5768, and the acid value was ASTM D664.
Results
Qualitative screening for fungal isolates
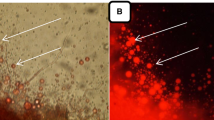
The present study carried out on 36 fungal locally isolates from different Egyptian soil samples. All fungal isolates exhibited strong fluorescence signals when using Nile red assay. Figure 1 indicated the lipid particles under a fluorescence microscope.

Fluorescence microscopy of Nile-red stained F. oxysporum NRC2017
Identification of the promising fungal isolate
The best selected fungal isolate for lipid production was identified F. oxysporum NRC2017. Figure 2 showed the phylogenetic tree structure for F. oxysporum NRC2017 from the Gene bank database (http://www.ncbi.nlm.nih.gov/). The 18S rRNA sequences of F. oxysporum NRC2017 was deposited to Gene bank under accession number MF62208.

Phylogenetic tree showing the relationships among the selected isolate F. oxysporum NRC 2017 and other closely related sequences collected from the GeneBank
Optimization of the culture conditions for the best fungal lipid production
The effect of nutritional and environmental parameters for the best lipid production of the identified strain F. oxysporum NRC2017 was indicated as follows.
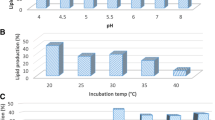
Effect of initial medium pH on fungal lipid production
The medium pH was adjusted for F. oxysporum NRC2017 at different values. Figure 3 showed the highest lipid accumulation by F. oxysporum NRC2017reached to 64.59% at initial pH 5.0.

Effect of initial pH on F. oxysporum NRC 2017 lipid production
Effect of incubation temperature on fungal lipid production
F. oxysporum NRC2017 was incubated at different incubation temperatures at pH 5.0. Figure 4 indicated the best value of lipid production 55.27% at incubation temperature 30 °C.

Effect of incubation temperatures on F. oxysporum NRC 2017
Effect of incubation period on fungal lipid production
F. oxysporum NRC2017 strain was inoculated at pH 5.0 and incubated at 30 °C for different tested periods. Figure 5 showed the best lipid production 55.22% after incubation period 9 days.

Effect of incubation period on F. oxysporum NRC 2017
Effect of inoculum size on fungal lipid production
The cultures of F. oxysporum NRC2017was adjusted at different values at the previously mentioned optimum conditions. Figure 6 indicated the best lipid production 59.88% by using inoculum size 1%.

Effect of inoculum size on F. oxysporum NRC 2017 lipid production
Effect of static and shaking conditions on fungal lipid production
Static and shaking condition at 80 rpm was evaluated by the growth of F. oxysporum NRC2017 strain at the previously mentioned optimum conditions. Figure 7 indicated the highest values of lipid production 50.32% higher than that produced under shaking condition.

Effect of static and shaking conditions on F. oxysporum NRC 2017 lipid production
Effect of different carbon sources on fungal lipid production
An equimolar amount of five different carbon sources the inoculum size of was at 1% broth medium, pH 5.0 then incubated for 9 days under static condition. The results in Fig. 8 indicated the best lipid production 53.32% in presence of xylose.

Effect of different carbon sources on F. oxysporum NRC 2017 lipid production
Effect of different nitrogen sources on fungal lipid production
An equimolar amount of six different nitrogen sources at the previously mentioned optimum conditions. The results in Fig. 9 indicated the best lipid production 52.42% in the presence of yeast extract while inorganic nitrogen sources were good for cell growth but not for lipid accumulation.

Effect of different nitrogen sources on F. oxysporum NRC 2017 lipid production
Agro-industrial by-products as feedstock to produce fungal lipid
The results in Fig. 10 indicated that F. oxysporum NRC2017 appeared lipid production on all six agro-industrial by-products. The highest value 98.27 mg/g was obtained with baggase.

Lipid production by F. oxysporum NRC 2017 using different by products as semi solid state
Biodiesel production
The extracted lipid from F. oxysporum NRC2017 was estimated by Sulfo-phospho-vanillin reagent (SPV) technique. After lipid was extracted from the tested isolate indirect transesterification process using acid as a catalyst used for transfer the extracted lipid into biodiesel. After biodiesel production gas chromatography and physical properties occur on it.
Analysis of biodiesel by gas chromatography
The composition of fatty acid methyl esters with the profiles for F. oxysporum NRC2017 was analyzed by GC. The single-cell oils were found to contain a high fraction of mono and polyunsaturated fatty acids, mainly 2.03% palmitoleic acid (C16:1), 11.13% oleic acid (C18:1), 5.57% linoleic acid (C18:2), 3.04% α-linoleic acid (C18:3), 2.52% eicosatrienoic acid (C20:3), 6.81% eicosapentaenoic acid (C20:5), and 16.12% erucic acid (C22:1), and a limited percentage of saturated fatty acids mainly 4.24% margaric acid (C17), 0.33% stearic acid (C18), and 3.86% behenic acid (C22).
Biodiesel physical properties
In this work, the amount of biodiesel produced by the tested fungal identified isolate was blended with diesel at a ratio of 5%. Table 1 showed the biodiesel physical properties obtained from blending biodiesel F. oxysporum NRC2017 as compared with standard diesel ASTM, standard biodiesel ASTM, and ASTM D975 (standard biodiesel for B5).
Discussion
Several fungal species are attractive microorganisms because they have high oil content (Hoffmeister and Keller 2007). In this study, all fungal isolates exhibited strong fluorescence signals when using Nile red assay. Nile red stain emitting strongly red fluorescence positive signals only with hydrophobic compounds such as any lipid particles inside the cells and could be detected by fluorescence spectroscopy (Beopoulos et al. 2008). The best selected fungal isolate for lipid production was identified Fusariumoxysporum NRC2017 that agreement with fungal taxonomy based on molecular techniques exhibiting high specificity and sensitivity for identifying microorganisms (Sette et al. 2006). The 18S rRNA sequences of F. oxysporum NRC2017 was deposited to Gene bank under accession number MF62208.The effect of nutritional and environmental parameters for the best lipid production of the identified strain F. oxysporum NRC2017 was indicated as following. Lipid content varied according to the nature of microorganism and culture conditions like pH of the medium, incubation temperature, incubation period, static and shaking conditions, carbon sources, and nitrogen sources (Alvarez and Steinbuchel 2002; Papanikolaou et al. 2004; Abdelhamid et al. 2019). The highest lipid accumulation by F. oxysporum NRC2017 reached 64.59% at initial pH 5.0. It was known that the function of the plasma membrane was to regulate the transport of substances from in and out the cells (Amanullah et al. 2001). Comparable results were obtained by Ali and El-Ghonemy (2014) and Jiru et al. (2017); they recorded that pH range was found to be the suitable pH for most fungal growth. F. oxysporum NRC2017 was incubated at different incubation temperatures at pH 5.0. The best value of lipid production 55.27% at incubation temperature 30 °C this result was congruent with Venkata and Venkata (2014); they reported that optimum temperature for Aspergillus awamori was 30 °C; however, lipid production was drastically reduced gradually when the fungus was incubated at a higher temperature (35–40 °C). F. oxysporum NRC2017 strain showed the best lipid production 55.22% after incubation period 9 days, while Peng et al. (2011) studied the mutant strain Mortierella alpine (MAI502–8) grown on mixture of glucose and xylose and found high accumulation of biomass and lipid, after 11 days of incubation time. The highest values of lipid production were 50.32% by F. oxysporum NRC2017 under static condition. These results were similar to Ali and El-Ghonemy (2014). The best lipid production was 53.32% in presence of xylose. These results were congruent with Li et al. (2011); they studied the oleaginous fungi that lived in the Qinghai plateau and reported that fungal isolates were able to use xylose in the industrial application of biodiesel while the majority of oil-producing microorganisms are known to use glucose to produce oil (Loffhagen et al. 2006; Li et al. 2007). The best lipids production 52.42% in presence of yeast extract while inorganic nitrogen sources were good for cell growth but not for lipid accumulation. These results were almost on the same line with Huang et al. (1998); they reported that inorganic nitrogen sources were good for cell growth but not suitable for oil production. F. oxysporum NRC2017 appeared lipid production on all six agro-industrial by-products. The highest value 98.27 mg/g was obtained with baggase. This result was almost accepted with Pandey et al. (2000) who reported that fungi could grow well under solid-state conditions and their hyphal penetration into solid substrates. The extracted lipid from F. oxysporum NRC2017 was estimated by Sulfo-phospho-vanillin reagent (SPV) technique. Also, Mishra et al. (2014) used SPV reaction to rapidly obtain an overview of lipid accumulation in microalgae. After lipid was extracted from the tested isolate indirect transesterification process using acid as catalyst used to transfer the extracted lipid into biodiesel. This technique was confirmed by Lewis et al. (2000) and Ren et al. (2017); they reported that indirect transesterification was the popular method for lipid extraction. After biodiesel production, gas chromatography and physical properties occurred on it. The composition of fatty acid methyl esters with the profiles for F. oxysporum NRC2017 was analysis. The single-cell oils were found to contain a high fraction of mono and polyunsaturated fatty acids mainly 2.03% palmitoleic acid (C16:1), 11.13% oleic acid (C18:1), 5.57% linoleic acid (C18:2), 3.04% α-linoleic acid (C18:3), 2.52% eicosatrienoic acid (C20:3), 6.81% eicosapentaenoic acid (C20:5), and 16.12% erucic acid (C22:1) and a limited percentage of saturated fatty acids mainly 4.24% margaric acid (C17), 0.33% stearic acid (C18), and 3.86% behenic acid (C22). Fatty acid profiles of the lipid were in agreement with Subramanian et al. (2010) and Fei et al. (2011) those found in plants. These include palmitic acid (C16:0), stearic acid (C18:0), oleic acid (C18:1), linoleic acid (C18:2), and linolenic acid (C18:3). In this work, the amount of biodiesel produced by the tested fungal identified isolate was blended with diesel at ratio 5%. The obtained blending biodiesel B5 was accepted by Pimentel et al. (2006) and Knothe (2006), they reported that all engine manufacturers provided certain warranties on the engines powered with biodiesel while B5 was accepted by all engine manufacturers.
Conclusion
The promising fungal identified isolate F. oxysporum NRC2017 designated for biodiesel production was isolated from Egyptian soils. Optimum conditions for maximum lipid production 55.2% obtained when the basal liquid medium was used with xylose as a carbon source and yeast extract as a nitrogen source, with initial pH 5.0 after 9 days in a static condition. When agro-industrial by-products were used as semi-solid fermentation the highest lipid accumulation for F. oxysporum NRC2017 was 98.3 mg/g in the presence of baggase. Gas chromatography analysis of the tested fungal species revealed that the fatty acids suitable for biodiesel production. The physical properties of blending biodiesel that was produced from the tested fungal identified isolate were in accordance with biodiesel standards. Therefore, this study revealed the possibility of using the promising fungus F. oxysporum NRC2017 for biodiesel production.
Availability of data and materials
All data and materials are available
Abbreviations
- ASTM:
-
American Society for Testing Materials
- FAME:
-
Fatty acid methyl ester
- GC:
-
Gas chromatography
- mm:
-
Millimeter
- mM:
-
Millimolar
- NRC:
-
National Research Centre
- PCR:
-
Polymerase chain reaction
- rpm:
-
Revolutions per minute
- SPV:
-
Sulfo-phospho-vanillin
- TAG:
-
Triacylglycerols
References
Abdelhamid SA, Hussein AA, El Sayed OH, Asker MS, Mohamed SS (2019) Optimization of culture conditions for biodiesel production from Egyptian isolate Penicillium commune NRC2016. BNRC 43:15–23
Ali TH, El-Ghonemy DH (2014) Optimization of culture conditions for the highest lipid production from some oleaginous fungi for biodiesel preparation. Asian J Appl Sci 2:600–609
Alvarez HM, Steinbuchel A (2002) Triacylglycerols in prokaryotic microorganisms. Appl Microbiol Biotechnol 60:367–376
Amanullah A, McFarlane CM, Emery AN, Nienow AW (2001) Scale-down model to simulate spatial pH variations in large-scale bioreactors. Bioethanol Bioeng 73:390–399
Beopoulos A, Mrozova Z, Thevenieau F, Le Dall MT, Hapala I, Papanikolaou S, Chardot T, Nicaud JM (2008) Control of lipid accumulation in the yeast Yarrowia lipolytica. Appl Environ Microbiol 74:7779–7789
Bligh EG, Dyer WJ (1959) A rapid method for total lipid extraction and purification. Can J Biochem Physiol 37:911–917
Brennan L, Owende P (2010) Biofuels from microalgae - a review of technologies for production, processing and extractions of biofuels and co-products. Renew Sust Energ Rev 14:557–577
Canakci M, Van GJ (2001) Biodiesel production from oils and fats with high free fatty acids. ASABE 44:1429–1436
Demirbas A (2003) Biodiesel fuels from vegetable oils via catalytic and non-catalytic supercritical alcohol transesterifications and other methods: a survey. Energy Conv Manag 44:2093–2109
Demirbas A (2008) Biodiesel. In: Demirbas A (ed) A realistic fuel alternative for diesel engines. Springer, London, pp 39–64
Fei Q, Chang HN, Shang L, Choi JDR, Kim N, Kang J (2011) The effect of volatile fatty acids as a sole carbon source on lipid accumulation by Cryptococcus albidus for biodiesel production. Bioresour Technol 102:2695–2701
Folch J, Lees M, Stanley GHS (1957) A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226:497–509
Gavrilescu M, Chisti Y (2005) A sustainable alternative for chemical industry. Biotechnol Adv 23:471–499
Greenspan P, Mayer EP, Fowler SD (1985) Nile red: a selective fluorescent stain for intracellular lipid droplets. JCB 100:965–973
Gunston FD (2001) Basic oleochemicals, oleochemical products, and new industrial oils. In: Gunstone FD, Hamilton RJ (eds) Oleochemical manufacture and applications. Sheffield Academic Press, England, pp 1–23
Hanna MA (1991) Biodiesel production: a review. Bioresour Technol 70:1–15
Hoffmeister D, Keller NP (2007) Natural products of filamentous fungi: enzymes, genes, and their regulation. Nat Prod Rep 24:393–416
Huang J, Shi Q, Zhou X, Lin Y, Xie B, Wu S (1998) Studies on the breeding of Mortierella isabellina mutant high producing lipid and its fermentation condition. Microbiology 25:187–191
Jiru TM, Groenewald M, Pohl C, Steyn L, Kiggundu N, Abate D (2017) Optimization of cultivation conditions for biotechnological production of lipid by Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 for biodiesel preparation. 3 Biotech 7:145–155
Knothe G (2006) Analyzing biodiesel: standards and other methods. J Amer Oil Chem Soc 83:823–833
Kotasthan T (2017) Potential of microalgae for sustainable biofuel production. JMSRD 7:223–232
Lewis T, Nichols PD, McMeekin TA (2000) Evaluation of extraction methods for recovery of fatty acids from lipid producing microheterotrophs. J Microbiol Methods 43:107–116
Li YH, Zhao ZB, Bai FW (2007) High-density cultivation of oleaginous yeast Rhodosporidium toruloides Y4 in fed batch culture. Enzym Microb Technol 41:312–317
Li Q, Du W, Liu D (2008) Perspectives of microbial oils for biodiesel production. Appl Biochem Microbiol 80:749–756
Li SL, Feng SL, Li ZT, Xu H, Yu YP, Qiao DR, Ao Y (2011) Isolation, identification and characterization of oleaginous fungi from the soil of Qinghai Plateau that utilize D-xylose. Afr J Microbiol Res 5:2075–2081
Lim SH, Ming H, Park EY, Choi JS (2003) Improvement of riboflavin production using mineral support in the culture of Ashbya gossypii. Food Tech Biotec 41:137–144
Loffhagen N, Hartig C, Harms H (2006) Impact of membrane fatty acid composition on the uncoupling sensitivity of the energy conservation of Comamonas testosterone TCC 17454. Appl Microbiol Biotechnol 70:618–624
Mishra SK, Suh WI, Farooq W, Moon M, Shrivastav A, Park MS, Ji WY (2014) Rapid quantification of microalgal lipids in aqueous medium by a simple colorimetric method. Bioresour Technol 155:330–333
Pandey A, Soccol CR, Mitchell D (2000) New developments in solid state fermentation: I-bioprocesses and products. Process Biochem 35:1153–1169
Papanikolaou S, Komaitis M, Aggelis G (2004) Single cell oil (SCO) production by Mortierella isabellina grown on high-sugar content media. Bioresour Technol 37:287–291
Peng W, Lamei Z, Zhiming Z, Li W, Hui W, Chengling Y, Guohong G (2011) Microbial lipid production by co-fermentation with Mortierella alpina obtained by ion beam implantation. Chem Eng Technol 34:422–428
Pimentel MF, Ribeiro GM, da Cruz RS, Stragevitch L, Pacheco Filho JGA, Teixeira LSG (2006) Determination of biodiesel content when blended with mineral diesel fuel using infrared spectroscopy and multivariate calibration. Microchem J 82:201–206
Ren X, Zhao X, Turcotte F, Deschênes JS, Tremblay R, Jolicoeur M (2017) Current lipid extraction methods are significantly enhanced adding a water treatment step in Chlorella protothecoides. Microbial Cell Fact 16:26–39
Rottig A, Wenning L, Broker D, Steinbuchel A (2010) Fatty acid alkyl esters: perspectives for production of alternative biofuels. Appl Microbiol Biotechnol 44:1713–1733
Rude MA, Schirmer A (2009) New microbial fuels: a biotech perspective. Curr Opin Microbiol 12:274–281
Sergeeva YE, Galanina LA, Andrianova DA, Feofilova EP (2008) Lipids of filamentous fungi as a material for producing biodiesel fuel. Appl Biochem Microbiol 44:523–527
Sette LD, Passarini MRZ, Delarmelina C, Salati F, Duarte MCT (2006) Molecular characterization and antimicrobial activity of endophytic fungi from coffee plants. WJMB 22:1185–1195
Subramanian R, Dufreche S, Zappi M, Bajpai R (2010) Microbial lipids from renewable resources: production and characterization. J Indus Microb Biotech 37:1271–1287
Tao J, Dai CC, Dai Q (2006) The conversion efficiency and economic feasibility of microbial energy. Chin J Microbiol 26:48–54
Venkata SG, Venkata MS (2014) Lipid accumulation for biodiesel production by oleaginous fungus Aspergillus awamori: influence of critical factors. Fuel 116:509–515
Vicente G, Bautista LF, Rodriguez R, Gutierrez FJ, Sadaba I, Ruiz VRM, Torres MS, Garre V (2009) Biodiesel production from biomass of an oleaginous fungus. Biochem Eng J 48:22–27
Vicente G, Bautista LF, Gutiérrez FJ, Rodríguez R, Martínez V, Rodríguez FRA, Ruiz VRM, Torres MS, Garre V (2010) Direct transformation of fungal biomass from submerged cultures into biodiesel. Energy Fuel 24:3173–3178
Acknowledgements
National Research Centre and Ain Shams University
Funding
All funding from the National Research Centre
Author information
Authors and Affiliations
Contributions
All the participant researchers contribute to do this work and this research was from the thesis of Abdelhamid, S.A. (2018), Biochemical studies on the production of biodiesel from some species of fungi, MSc, Ain Shams University, Egypt. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
All the participant researchers consent for publication
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sayeda, A.A., Mohsen, S.A., Osama, H.E.S. et al. Biodiesel production from Egyptian isolate Fusarium oxysporum NRC2017. Bull Natl Res Cent 43, 210 (2019). https://doi.org/10.1186/s42269-019-0254-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42269-019-0254-z