
Abstract
Background
Tree-related microhabitats (hereafter, “TreMs”) are key components of forest biodiversity but they are still poorly known in North American hardwood forests. The spatial patterns of living trees bearing TreMs (hereafter, “TreM-trees”) also remain to be determined. As logging practices can lead to a loss of TreM-trees and of their associated biodiversity, it is essential to identify the factors explaining TreM occurrence to better integrate them into forest management. We therefore inventoried TreMs in 4 0.5-ha survey strips in northern hardwood forests in Quebec, Canada, while recording the spatial location of each tree. Two strips were located in unmanaged old-growth forests, and 2 were in forests managed under selection cutting. All 4 stands were dominated by sugar maple (Acer saccharum Marsh.) and American beech (Fagus grandifolia Ehrn.). Beech bark disease, an exotic pathology, was observed in all the strips.
Results
Large diameter at breast height and low tree vigor were the main characteristics explaining the presence of TreMs at the tree scale. TreM-trees presented slight spatial aggregation patterns. These aggregates, however, were not well-defined and were generally constituted by a large number of trees bearing few different types of TreMs. Two TreM classes (broken branch or top and woodpecker lodge) also presented a spatial aggregation. Logging practices had no significant effect on TreM occurrence. Beech bark disease increased the frequency of senescent beeches. The impact of this pathology on TreMs was however mitigated by the small size of infected trees and probably by the short time elapsed since its appearance.
Conclusion
The factors explaining the presence and abundance of TreMs on trees has so far been little studied in North American hardwood forests. Our results highlight that TreM-tree characteristics in the surveyed forests are consistent with those of previous studies conducted in other forest types and regions (e.g., Europe or Northwestern America). To our knowledge, this study is also the first to identify a spatial aggregation of TreM-trees and of specific TreM classes. It will be nevertheless necessary to determine whether the small impact of logging activities we observed results from current or past management practices.
Similar content being viewed by others

Background
Finding a balance between wood production and other ecosystem services, such as biodiversity, carbon sequestration or aesthetic values, is now a common objective of forest management (Sarr and Puettmann 2008; Puettmann et al. 2009; Watson et al. 2018). For this purpose, many silvicultural practices that mimic natural forest dynamics have been proposed. These strategies for sustainable forestry include maintaining continuous forest cover, complex horizontal and vertical forest structure, large trees, as well as standing or fallen deadwood (Bauhus et al. 2009; Raymond et al. 2009; Eyvindson et al. 2021). However, evaluating the capacity of actively managed forest landscapes to maintain structural attributes and biodiversity close to that of natural forests is challenging. Exhaustive biodiversity surveys are often hard to implement, expensive and generally concentrated on a few taxa (Puumalainen et al. 2003; Burrascano et al. 2018; Larrieu et al. 2019). Accordingly, proxies of forest biodiversity are generally preferred in lieu of exhaustive surveys (Burrascano et al. 2018; Larrieu et al. 2018a; Larrieu et al. 2018b; Barrette et al. 2020).
Tree-related microhabitats (hereafter “TreMs”) are among the proposed indicators that can help to better identify the structural elements supporting forest biodiversity (Larrieu et al. 2018a; Asbeck et al. 2021). They are defined as “all distinct and well-delineated structures occurring on living or standing dead trees, that constitute a particular and essential substrates or life site for species or species communities during at least a part of their life cycle to develop, feed, shelter or breed” (Larrieu et al. 2018a). Examples of TreMs include cavities, broken branches or trunks, and fruiting bodies of saproxylic fungi that harbor specific animal, vegetal and fungal species (Bouget et al. 2013; Regnery et al. 2013; Larrieu et al. 2018a; Paillet et al. 2018; Basile et al. 2020). TreM surveys are easier to conduct than complete censuses of species richness and diversity (Larrieu et al. 2018a). For example, Martin and Raymond (2019) showed that they could be easily identified at the same time as tree-defect inventories, which are common in temperate managed forests.
Research considering the concept of TreMs as structural and functional resources for a wide range of taxa is scarce in North America, and most of the current knowledge is built on studies from temperate, Mediterranean and mountain forests of Europe (Larrieu et al. 2018a). The majority of the existing research in North America comes from the West Coast (Stevenson et al. 2006; Michel and Winter 2009; Michel et al. 2011; Pritchard et al. 2017; Asbeck et al. 2020a). On the East Coast, the few existing studies on TreMs were performed either on urban trees compared to trees in natural hardwood forests (Großmann et al. 2020), in the conifer-dominated boreal forest (Martin M, Fenton NJ, Morin H: Tree-related microhabitats and deadwood dynamics form a diverse and constantly changing mosaic of habitats in boreal old-growth forests, submitted) or in the temperate mixedwood forest (Martin and Raymond 2019), the latter marking the transition from boreal to temperate forests. In terms of management guidelines, TreM-trees are almost exclusively those with cavities and other TreMs are rarely considered (Ontario Ministry of Natural Resources (OMNR) 2004; Ministère des Forêts de la Faune et des Parcs (MFFP) 2017). Therefore, better knowledge of the full range of tree microhabitats in temperate hardwood forests is needed to set the standards for maintaining their diversity and abundance in managed forests.
In Europe and western North America, studies highlighted that a larger diameter at breast height (DBH) and a lower tree vigor are the main characteristics explaining the presence of numerous and/or large TreMs (Vuidot et al. 2011; Winter et al. 2015). It is thus likely that the same factors will drive TreM formation in temperate forests of eastern North America. Yet, little is known about the spatial arrangement of TreM-trees (i.e., tree bearing at least one TreM), for example, if they are spatially clumped or, on the contrary, dispersed. As TreMs often result from abiotic or biotic damages (e.g., wind damage, fall of a dead tree or branch, fungal infection, insect epidemics), a distinct spatial pattern of TreM-trees has been expected (Kozák et al. 2018; Asbeck et al. 2020b), but has been observed only for woodpeckers cavities (Puverel et al. 2019). Understanding the spatial structure of TreMs is, nonetheless, a key factor in developing sustainable silvicultural practices aiming to maintain biodiversity in managed forests. Innovative silvicultural systems for example aim to increase the horizontal complexity of forest stands by combining the creation of various-sized gaps and the selection of individual trees (Kane et al. 2011; Bédard et al. 2014; Raymond et al. 2018). Knowing the spatial patterns of TreM-trees would help to maintain patches of habitat trees and to ensure that they act as effective ecological corridors for taxa with low dispersal capacity. Yet, silvicultural practices can have a negative impact on TreMs. According to European studies carried out in hardwood or mixedwood temperate forests, silvicultural practices generally cause a loss of TreM abundance and diversity, because (i) trees considered as senescent or with lower wood quality are progressively removed, and (ii) trees are often harvested before they can reach a diameter that favor the development of TreMs (Winter and Möller 2008; Larrieu et al. 2012; Larrieu et al. 2014; Paillet et al. 2017). The impacts of silviculture on TreMs must therefore be evaluated in terms of density, diversity and general composition.
In this study, we aim to identify the factors promoting TreM development on living trees in hardwood temperate forests of eastern North America. Selection cutting, i.e. the harvest of trees of multiple sizes to create gaps, is a silvicultural practice common in these forests. We predicted that in unmanaged old-growth forests, a combination of tree characteristics (large diameter at breast height, low tree vigor) and of spatial variables (spatial aggregation) increases the probability that a tree develops TreMs. Moreover, we expect that selection cutting will create a more uniform spatial arrangement of TreM-trees, as these would be logged first to promote healthy and/or valuable trees at a more or less regular spacing to optimize wood production.
Material and methods
Study area
The study was conducted in the Papineau-Labelle Wildlife Reserve (45°59′ N, 75°20′ W), a 162,800 ha territory landscape in the sugar maple (Acer saccharum Marshall)–yellow birch (Betula alleghaniensis Britton) bioclimatic ecoregion (Saucier et al. 2009) in western Quebec, Canada (Fig. 1). Mean annual temperatures range from 2.5 °C to 5 °C, precipitations from 900 to 1100 mm, and the growing season lasts from 170 to 180 days (Gosselin 2002). The topography is essentially characterized by hills with gentle slopes. Sugar maple and yellow birch are the dominant tree species. Other species such as American beech (Fagus grandifolia Ehrh.), white birch (Betula papyrifera Marshall), red maple (Acer rubrum L.), or balsam fir (Abies balsamea (L.) Mill.) can also be found (Gosselin 2002). Most of the forests in this region have been actively managed since the early twentieth century (Gaffield 1994). The forest was harvested by selective cutting practices such as diameter-limit cutting, i.e., the harvest of healthy trees that reached a specific diameter, until selection cutting, i.e., the harvest of trees of multiple sizes to create small gaps, became prevalent in the years 1990s (Bédard et al. 2014; Lussier and Meek 2014; Nyland 2016). The most notable exception is the Lac-de-l’Écluse forest (45°52′ N–75°24′ W), one of the largest landscapes (7.74 km2) of hardwood old-growth forest found in Quebec. This protected area is considered as an old-growth forest because (i) it has not been severely affected by natural disturbances since several centuries and (ii) present no traces of forest management since the European settlement in this region (Villeneuve and Brisson 2003). The influence of First Nations on North American forests prior to this time was essentially forest fires (Munoz and Gajewski 2010; Blarquez et al. 2018). Beech bark disease (BBD) is an exotic pathology that has been present in the study territory at least since 2013 and currently causes a marked senescence and mortality of beech trees (Morin et al. 2007; Ministère des Forêts de la Faune et des Parcs (MFFP) 2020).

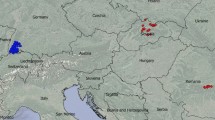
Location of the study area in eastern North America (a) and location of the survey strips in the Papineau-Labelle Wildlife Reserve (b). “Sel”: selection cutting, “OG”: unmanaged, old-growth forests
Sampling design and strip characteristics
In 2014, we set up two survey strips (0.5 ha each, 500 m-long, 10 m-wide) in managed forests (named “Sel1” and “Sel2”) and 2 others of the same size in the Lac-de-l’Écluse old-growth forest (named “OG1” and “OG2”; Fig. 1). Their location was determined using stratified random sampling, based on five variables defining forest tree species composition, successional stage and history: (i) potential vegetation (namely, the theoretical tree composition at the end of the succession, here a sugar maple–yellow birch forest); (ii) stand structure (irregular and older than 80 years based on the provincial forest survey age and structure typology (Ministère des Ressources Naturelles et de la Faune du Québec (MRNF) 2008)); (iii) logging history (logged or unlogged, that is to say, within the Lac-de-l’Écluse old-growth forests); (iv) distance from forest road (at most, 125 m to avoid any edge effect); and (v) size sufficient to survey a 500 m-long strip following a cardinal point without encountering any change in potential vegetation or edge effect.
Each strip was delimited using hip chain and measuring tape, and by aiming a compass at 1 of the 4 cardinal points. When we crossed a tree with a DBH ≥ 19 cm within the strip limits (i.e., at most 5 m from the center of the strip), we recorded its species, DBH, vigor class (vigorous, senescent (with a low probability of survival according the tree vigor classification scheme of Boulet (2005)) or dead (snag)), location within the strip and the occurrence of TreMs (presence/absence, on living trees only). When these were present, we noted the TreM classes (Table 1) adapted from Emberger et al. (2013). We considered the centroid of the trunk at breast height along the north/south and east/west axes as the exact location of the tree within the strip. We sampled a total of 366 living trees and 58 snags.
Statistical analyses
To assess our prediction that a combination of environmental and spatial variables increases the probability of a tree developing TreMs, we first performed a Bayesian generalized linear mixed effects model (GLMM) with a Poisson distribution for count data (hereafter, “Poisson GLMM”), based on the attributes of surveyed living trees and using the bglmer() function of the blme package (Chung et al. 2013) in R software (R Core Team 2019). We decided to use a mixed-effect model to limit the influence of the spatial links that might exist between trees in a same transect and that can influence models’ reliability (Dormann et al. 2007). The random effect consisted therefore of the nested transect with the forest management status (managed under selection cutting or old-growth; hereafter “Management”), similarly as previous research studying TreMs (e.g., Paillet et al. 2019; Asbeck et al. 2020b). The Bayesian approach was chosen to avoid model singularity (i.e., variances of one or more linear combinations of effects are close to zero), as suggested by Bates et al. (2015), using a covariance matrix of the random effects based on the Wishart distribution (Chung et al. 2013). The dependent variable was the total number of TreM-classes on the tree, and the independent variables were tree DBH, tree vigor, management and number of TreM-trees among the five closest living trees (“TreM-trees aggregation”). We used the function step() in R software to perform a stepwise selection to obtain the most parsimonious model. The validity of the model (overdispersion, zero-inflation) was assessed using the DHARMa package (Harting 2019). Then, we performed for each TreM class a Bayesian GLMM with a logistic distribution for binary data (hereafter, “Logistic GLMM”), using the same random effects and covariance matrix than the Poisson GLMM. The dependant variable was the presence/absence of the TreM class studied on each tree, and the independent variables the same as the Poisson GLMM. For each model, we also performed a stepwise selection to obtain the most parsimonious model. Validity of the models were verified similarly as before. For each logistic GLMM, we determined its predictive ability using the area under the receiver operating characteristic curve (AUC, Zweig and Campbell 1993), which ranges from 0.5 (uninformative model) to 1 (perfect model; Fielding and Belll 1997).
Tree species was not included in the GLMMs models, as preliminary analyses indicated that this variable was strongly associated with tree vigor because of the BBD. To better understand the influence of tree species in our results, we first analyzed the frequency of TreM-trees (living trees bearing at least one TreM class), TreM classes and tree vigor classes by tree species, using pairwise Fisher tests with a Bonferroni adjustment (Mangiafico 2016). Less common tree species were grouped in a same category (“Other species”). It was sometimes impossible to clearly determine the species of dead trees. Therefore, trees of unknown species were not considered in the Fisher analysis of tree vigor classes. Second, we compared the DBH of living sugar maple and American beech trees using Kruskal-Wallis test, also taking into account the presence/absence of TreMs (sugar maples with TreMs, sugar maples without TreMs, American beeches with TreMs, American beeches without TreMs). If this test result was significant, we then performed Dunn’s post hoc test (Dunn 1964) with a Bonferroni correction. Finally, to determine if specific TreM classes tended to be more closely associated with other classes on the sampled trees and if selection cutting maintained specific TreM assemblages, we performed a non-metric dimensional scaling analysis (NMDS) based on the presence/absence of the different TreM classes. The NMDS was followed by an analysis of similarities (ANOSIM; Clarke 1993), with management as the dependent variable, using Jaccard distance and 9999 permutations. Only living trees bearing at least 2 different TreM classes were considered in this analysis.
The analyses were performed on R software, version 3.6.1 (R Core Team 2019), using the blme (Chung et al. 2013), DHARMa (Harting 2019), ROCR (Sing et al. 2005), Desctools (Signorell 2017), emmeans (Russel 2018), FactoMiner (Lê et al. 2008), rcompanion (Mangiafico 2019), FSA (Ogle et al. 2019) and vegan (Oksanen et al. 2018) packages.
Results
Strip characteristics
The studied strips were dominated by sugar maple, with American beech as the second dominant species (Table 2). Other tree species were present to a lesser, except in the OG1 strip, where the presence of a few large eastern hemlocks (Tsuga canadensis (L.) Carrière) and yellow birches explains greater basal area values. The mean DBH of living trees and snag was equal to 34.3 ± 12.8 cm and 41.9 ± 11.0 cm, respectively. The DBH range in the studied strips was equal to 19–84 cm for the trees and 22–74 cm for the snags.
TreM composition in northern hardwoods
We found that 40% of living trees bore at least one TreM, for a mean density of 72.5 TreMs·ha− 1 (Table 3). TreM-trees were observed in each transects but few trees contained at least 3 different TreM classes (Fig. 2). On average, TreM-trees had 1.49 TreM classes and a mean DBH of 40.4 cm (Table 3). The most abundant TreM classes were bark loss (30.5 trees·ha− 1) and rot holes (mean density: 26 trees·ha− 1). In contrast, the least abundant TreM classes were cracks (mean density: 3.5 trees·ha− 1) and trunk base rot holes (mean density: 4 trees·ha− 1).

Location, DBH, tree vigor and number of TreM classes of the studied trees in the managed under selection cutting (“Sel”) and unmanaged, old-growth (“OG”) transect strips
Factors driving TreM occurrence
The Poisson GLMM was significant (p < 0.001) and contained three variables, all significant: DBH (p < 0.001), tree vigor (p < 0.001) and TreM-trees aggregation (p = 0.006) (Table 4). The number of different TreM classes observed on the same tree increased significantly with DBH (Fig. 3a). Senescence and a higher agglomeration of TreM-trees also had a positive, although more moderate, effect (Fig. 3b and c). All logistic GLMMs were significant except for bark loss (p = 0.084), saproxylic fungi (p = 0.168) and trunk base rot hole (p = 0.172) (Table 5). For the significant models, we observed high and relatively homogeneous AUC values (0.74–0.84; Table 4). DBH had a significant positive influence in all final models (Fig. 4a). Tree vigor significantly increased the probability of occurrence of crown deadwood and woodpecker lodge (Fig. 4b). Management had no significant influence on the occurrence of the different TreM classes. We however observed a trend (p = 0.081) for crown deadwood, where the occurrence of this class was lower in forests managed under selection cutting (Fig. 4c). Finally, the TreM-trees aggregation positively influenced the occurrence of broken branch or top, as well as woodpecker lodge (Fig. 4d). One tree was removed from these analyses due to unknown tree vigor (number of trees considered in the analyses: 365).

Number of TreM classes predicted based on the explanatory variables of the Poisson regression: a) DBH, b) tree vigor, and c) TreM-trees aggregation. #: Number of TreM-trees in the 5 closest living trees. Details of the model are presented in Table 4

Estimate, standard deviation and significance of the independent variables of the logistic regressions estimating the probability of occurrence of the different TreM classes (details of the models are presented in Table 5). No results indicate that the variable was not included in the final model. The brackets indicate for discrete variables the class to which these results apply. “***”: variable significant at p < 0.001, “**”: variable significant at p < 0.01, “*”: variable significant at p < 0.05. Details of the results are provided in Supplementary Table S1
The occurrence of TreMs was not different among species. However, we observed significant differences in the frequency of TreMs and tree vigor classes between sugar maple and American beech (Table 6). Most TreM classes were observed on sugar maple. On American beech, only crown deadwood and saproxylic fungi were very frequent (observed on 40% and 56% of the TreM-trees, respectively), while other classes were infrequent (observed on less than 16% of the TreM trees). Similarly, American beech accounted for 75% of the senescent trees, whereas the majority of the vigorous trees were sugar maples (83% of the vigorous trees). Overall, other tree species presented few differences with sugar maple and American beech. The only significant difference concerned tree vigor, as a higher proportion of beeches were classified as senescent, compared to other species.
The DBH of sugar maple and American beech stems differed significantly and was associated with the occurrence of TreMs (Fig. 5). Sugar maples bearing at least one TreM had the largest DBH (41.7 cm), followed by sugar maples without TreMs (32.1 cm). American beeches without TreMs had the smallest DBH (25.1 cm), while those bearing at least one TreM presented intermediate values (31 cm).

Boxplot of sugar maple and American beech DBH, divided according to the occurrence of TreMs (TreM: presence of at least one TreM class; no TreM: no TreM class). Red dots indicate mean values, and letters indicate significant differences between the species/TreM classes
For trees bearing at least 2 different TreM classes, the NMDS separated the classes into 4 groups (Fig. 6). We observed positive values on both NMDS axes for broken branches or tops as well as for woodpecker lodges, but negative values on both axes for bark loss. Trunk base rot holes, fungi and crown deadwood had negative values on the first axis of the NMDS and positive values in the second. In contrast, we observed positive values on the first axis and negative values on the second for cracks and trunk rot holes. Hence, TreM classes were recurrently grouped on a same tree according to the following associations: trunk base rot holes/fungi/crown deadwood; broken branches or tops/woodpecker lodges; crack/trunk rot holes; fungi. However, the ANOSIM underscores that forest management had no influence on these assemblages (R = 0.024, p = 0.189).

Biplot of the NMDS performed on living trees bearing at least 2 different TreM classes, and results of the ANOSIM test between managed and unmanaged strips. n.s.: non-significant
Discussion
This study provides a better understanding of the attributes at the tree- and stand-scales that drive TreM occurrence in the hardwood forests in eastern North America, where this knowledge is still scarce. It is also one of the few studies currently examining the spatial patterns of TreM-trees (Kozák et al. 2018; Puverel et al. 2019; Asbeck et al. 2020b). The occurrence of TreMs in sugar maple–American beech forests resulted both from individual tree characteristics (large DBH and low tree vigor) and, to a lower extent, from the spatial arrangement of the trees in the forest. These spatial clumps were generally composed of several trees bearing a few TreM classes rather than by a few trees bearing several classes. Finally, we observed no marked influence of logging history on TreM-tree characteristics. Trees in unmanaged stands and those in treated stands under selection cutting and former diameter-limit cuts had thus similar characteristics.
Tree size and vigor are the primary drivers of TreM occurrence in sugar maple–American beech forests
In the sugar maple–American beech forests of our study, tree DBH and vigor were the main factors influencing the probability of TreM occurrence. A higher DBH favored the presence of several TreM classes on a tree and, individually, the occurrence of all the TreM classes studied except bark loss, saproxylic fungi and trunk base rot hole. A lower tree vigor also increased the number of TreM classes on the tree and the occurrence of crown deadwood and woodpecker lodges. These positive effects of a larger DBH and of a lower tree vigor on TreMs are consistent with previous studies conducted in the coniferous or mixed forests of North America (Michel and Winter 2009; Michel et al. 2011; Martin and Raymond 2019), as well as in the mixed or hardwood forests of Europe (Larrieu and Cabanettes 2012; Courbaud et al. 2017; Paillet et al. 2019) and Middle East (Jahed et al. 2020). Indeed, a larger diameter often implies a greater age, and hence, an increased risk of biotic or abiotic damages that lead to TreM formation (Paillet et al. 2019). A larger DBH also favors the development of TreMs that are large enough to be useful for biodiversity while not threatening tree survival (Courbaud et al. 2017; Larrieu et al. 2018a). Furthermore, many TreM types are wounds that can eventually lead to tree senescence or that can be interpreted as signs of senescence (Angers et al. 2005; Martin and Raymond 2019). This implies that larger senescence marks are more likely to occur in old and large trees.
In this study, we also observed a weak association between TreM classes that can be considered as indicators of tree senescence (crown deadwood, fungi and trunk base rot holes; Boulet (2005)). In contrast, other TreM classes can be considered as wounds that do not necessarily threaten short-term tree survival (broken branches or tops, woodpecker lodges, cracks or trunk rot holes), although they can significantly decrease stem mechanical properties and economic value (Boulet 2005; Havreljuk et al. 2014). TreM classes indicating senescence were more often observed on American beech while wounds were more frequent on sugar maple. These results are congruent with those of Guillemette et al. (2008), who noted that sugar maples, including large individuals, are highly resistant to injuries caused by both logging operations and natural agents (e.g., bark ripped off by the fall of a neighboring tree). Beeches, on the other hand, lose vigor more easily, although the influence of BBD in this result remained unclear. The association between certain TreM classes in the studied forests is therefore probably partly species-specific.
The co-occurrence of different TreM classes observed in our transect strips was also globally consistent with previous studies, for example broken branches or top with woodpecker cavities, or cracks with rot-holes, as well as the absence of some co-occurrences, for example between cracks and fungi (Larrieu and Cabanettes 2012; Regnery et al. 2013; Winter et al. 2015). The use of different TreM typologies between these studies however limits the comparison of the results, underlining the benefits of the homogenized typology proposed by Larrieu et al. (2018a). Overall, the factors driving TreM formation in the sugar maple–American beech forests of our study shared many common features with forests with other species compositions or in other biomes. The management of TreMs in North American forests can therefore draw on suggestions made for forests from other continents.
TreM-trees and specific TreM classes present slight spatial aggregation patterns
This is the first time, to our knowledge, that a spatial link between TreM-trees is identified. In particular, the higher the number of different TreMs classes observed, the more likely the surrounding trees were also carrying TreMs. Instead of well-delineated spatial patterns (in other words, a few trees bearing a high number TreM classes surrounded by trees without TreMs in the studied strips), we however observed a relatively slight spatial aggregation of TreM-trees. These results may be partly explained by the low number of TreM classes per tree (mean of 1.49 ± 0.76 classes per tree and a maximum of 4 classes observed on a same living tree). In uneven-aged stands, most TreM-trees indeed carry a few TreM classes, while only a few bear several TreM classes (Michel et al. 2011; Larrieu and Cabanettes 2012). As a result, the presence of TreM-trees bearing several TreM classes was generally diluted among trees bearing no TreMs or a few TreM classes in the stands. We observed a spatial aggregation only for broken branch or top and woodpecker lodge when the TreM classes were analyzed separately. The occurrence of broken branch or top often depends on relatively localized drivers (e.g., treefall or windthrow), which can explain this result. Dufour-Pelletier et al. (2020) pointed out that woodpeckers prefer to feed on spatially aggregated snags and senescent trees, even if they are relatively small in diameter. The proximity of woodpecker lodges to other TreMs trees, and thus potentially a source of food, is consistent with this behavior. It is also common for woodpeckers to forage cavities in dead branches on living trees, as the wood is softer (Martin et al. 2004; Nappi et al. 2015). The association observed between broken branch or top and woodpecker lodge can explain why the latter also presented a significant spatial pattern.
In our study, TreM-trees aggregation was never the main variable explaining the occurrence and diversity of TreMs. The relatively weak spatial links observed between TreM classes and TreM-trees can partially explain why Kozák et al. (2018) and Asbeck et al. (2020b) were unable to observe clear spatial patterns among TreM-trees in European forests. Only a selection of large trees (mean DBH > 50 cm) were however studied by Asbeck et al. (2020b), while we surveyed all trees with a DBH > 19 cm. Our methods might also be limited by the use of the TreM typology of Emberger et al. (2013). It indeed relies on 8 TreM classes, which may explain the low number of classes observed per tree. More detailed TreMs classification, such as the one defined by Larrieu et al. (2018a), where TreMs are divided in 7 forms, 15 groups and 47 types, could eventually help to better distinguish TreM spatial patterns in forest stands. Our sampling was also based on relatively narrow transects, exploring only a limited part of the immediate surroundings. Further studies, with more replications and partly based on large circular or rectangular plots (e.g., Kozák et al. 2018; Asbeck et al. 2020b) could therefore help to identify in greater detail the subtle spatial patterns highlighted in this study.
Limited impact of logging history on the characteristics of TreM-trees
Since we found no significant influence of logging history on the drivers of TreM occurrence in the studied strips, the characteristics of TreMs in the managed stand appear similar to those in the old-growth forest. We only observed a trend for crown deadwood, with an occurrence that was negatively although not significantly influenced by management under selection cutting. The death of the canopy can indeed be considered as an indicator of impending tree death (Guillemette et al. 2008), explaining why these trees are logged in priority. It is thus possible that TreMs perceived as defects could be considered of low urgency, while defects justifying the rapid harvesting of trees were not classified as TreMs (Martin and Raymond 2019). The first symptoms of BDD (spots and then small cracks in the bark), for example, cannot be considered as TreM but may justify the urgent harvest of the tree. These results are consistent with those of Vuidot et al. (2011) in mixed French forests that were either managed or left unmanaged for 10 to more than 150 years. These authors observed no influence of forest practices on TreMs at the tree scale, but a lower diversity and density at the stand scale. This negative impact of logging practices on TreMs has long been known in Europe (Winter and Möller 2008; Larrieu et al. 2012; Regnery et al. 2013) and to a lesser extent, in eastern North America (Stevenson et al. 2006; Michel and Winter 2009). Due to the limited number of strips in our study, we cannot determine whether managed stands have a lower TreM density and diversity than natural stands in the studied territory. Many forests of eastern North America are nonetheless characterized by a smaller human impact than European forests, due to their relatively recent occidental colonization (Potapov et al. 2008; Watson et al. 2018). Over time, past diameter-limit cut practices could also have increased the proportion of poor-quality trees—most likely to bear TreMs—in managed stands (Kenefic et al. 2005; Nyland 2016). Guidelines for the protection of habitat trees have also developed in recent decades (Ontario Ministry of Natural Resources (OMNR) 2004; Ministère des Forêts de la Faune et des Parcs (MFFP) 2017), which may lead to better conservation of TreM-trees in the studied transects. Martin and Raymond (2019) hence highlighted that silvicultural practices maintaining a continuous forest cover in mixed forests presented a TreM density and diversity similar to that observed in stands characterized by a small human footprint. These authors nevertheless underscored that the cumulative impact of logging over time could lead to a loss in TreM richness similar to what is observed in Europe. As such, Stevenson et al. (2006) and Michel and Winter (2009) already observed a lower density of TreM-trees in managed forests of western North America. In the study area, the protection of habitat trees also focuses mainly on snags and little on living trees (Ministère des Forêts de la Faune et des Parcs (MFFP) 2017). For these reasons, complementary research will be necessary to better evaluate the impacts of logging practices on TreMs in the forests of North America.
Limited influence of beech bark disease on TreM formation
BBD has been reported as an important driver of tree senescence and mortality in the study area (Ministère des Forêts de la Faune et des Parcs (MFFP) 2020). It could have thus influenced TreM formation as well as spatial patterns in the strips. More than 55% of the living American beeches were classified as senescent, as opposed to less than 5% of the living sugar maples in the studied strips. Similarly, crown deadwood and fungi, two indicators of BBD, were prevalent TreM classes in American beech (Savard et al. 2005; Taylor et al. 2013). These results suggest that BBD, which probably infected the study area around 2013 (Ministère des Forêts de la Faune et des Parcs (MFFP) 2020), may have influenced TreM development on American beeches in the studied stands. In forests where this disease has been present for several decades, Kahler and Anderson (2006) identified woodpecker lodges and broken branches as common in affected beeches. These two TreM classes were rare in the American beeches we sampled, but this could be because the disease killed the larger trees first (Tubbs and Houston 1990) or, conversely, because the arrival of the disease in the study area is too recent. The threshold we used to define broken branches or tops (diameter ≥ 20 cm at the broken point) requires large trees, and woodpeckers generally favor the largest trees to excavate their cavities (Swallow et al. 1988; Remm et al. 2006; Vaillancourt et al. 2008). In our study area, beech stems were generally small: even those with TreMs had a significantly smaller DBH than sugar maples without TreMs. Beeches affected by BBD were hence most likely to present TreM classes that depend little on tree size, such as fungi and crown deadwood. It is nevertheless difficult to estimate if the trend observed will change with the progression of the disease in the forests studied. Moreover, beeches that die because of BDD generally form clumps (Senécal et al. 2018). Yet, the proportion of beeches among snags or TreM-trees (26.4% and 14.8%, respectively) was relatively close to their abundance within the living trees (22.3%). This suggests that the presence of BBD in the studied strips did not create an abnormal number of TreM-trees or large snags at that stage. The relatively recent arrival of the disease as well as the scarcity of large beeches in the strips can explain these results. Moreover, the study area is at the northern range limit of American beech (Tubbs and Houston 1990), where their growth and survival could be limited. For all these reasons, it seems unlikely that BBD markedly influenced our results.
Conclusion
TreMs are still little known in northeastern North America, and this study provides a better understanding of the factors explaining their occurrence in temperate forests. Our results consistently link TreM characteristics in the studied forest with previous research conducted in other biomes and continents. We also both highlighted the slight spatial aggregation of TreM-trees and of specific TreM classes (broken branch and top, woodpecker lodge). Overall, TreMs were more frequent and diversified on large and/or senescent trees. Sustainable forestry practices must therefore maintain enough of these trees in managed stands to maintain viable populations of forest-dwelling species in comparison to natural forests. Such practices are coherent with those aiming to conserve old-growth forest attributes, such as deadwood or very large trees, in managed forests. Maintaining unharvested patches within managed forests to promote the development of TreMs and old-growth attributes would be congruent with the possible spatial aggregation of TreM-trees observed in this study. Moreover, the BDD had little influence on the occurrence of TreMs on living trees at the time of the survey. Harvesting infected trees to control the disease could, however, limit the supply of snags, which also provide many TreMs. It would be necessary to accurately evaluate the cost/benefice ratio of this practice, on particular on deadwood-dependant species. Finally, uncertainties remain regarding how the management history of the studied stands (an initial diameter-limit cutting followed by selection cutting) affects our results. Further research considering a greater variety of natural forests and stands with different management histories will therefore be necessary to better understand anthropogenic impacts on TreMs in North American hardwood forests. Similarly, more diverse and complete sampling designs could help to better understand the spatial patterns of TreM-trees.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- BDD:
-
Beech bark disease
- DBH:
-
Diameter at breast height
- OG:
-
Unmanaged, old-growth forest
- SEL:
-
Forest under selection cutting
- TreM:
-
Tree-related microhabitat
References
Angers VA, Messier C, Beaudet M, Leduc A (2005) Comparing composition and structure in old-growth and harvested (selection and diameter-limit cuts) northern hardwood stands in Quebec. For Ecol Manag 217(2–3):275–293. https://doi.org/10.1016/j.foreco.2005.06.008
Asbeck T, Basile M, Stitt J, Bauhus J, Storch I, Vierling KT (2020a) Tree-related microhabitats are similar in mountain forests of Europe and North America and their occurrence may be explained by tree functional groups. Trees-Struct Funct 34(6):1453–1466. https://doi.org/10.1007/s00468-020-02017-3
Asbeck T, Großmann J, Paillet Y, Winiger N, Bauhus J (2021) The use of tree-related microhabitats as forest biodiversity indicators and to guide integrated forest management. Curr For Reports 7(1):59–68. https://doi.org/10.1007/s40725-020-00132-5
Asbeck T, Messier C, Bauhus J (2020b) Retention of tree-related microhabitats is more dependent on selection of habitat trees than their spatial distribution. Eur J For Res 139(6):1015–1028. https://doi.org/10.1007/s10342-020-01303-6
Barrette M, Dumais D, Auger I, Boucher Y, Bouchard M, Bouliane J (2020) Naturalness assessment performed using forestry maps to validate forest management sustainability. Ecol Indic 119:106832. https://doi.org/10.1016/j.ecolind.2020.106832
Basile M, Asbeck T, Jonker M, Knuff AK, Bauhus J, Braunisch V, Mikusiński G, Storch I (2020) What do tree-related microhabitats tell us about the abundance of forest-dwelling bats, birds, and insects? J Environ Manage 264. https://doi.org/10.1016/j.jenvman.2020.110401
Bates D, Mächler M, Bolker BM, Walker SC (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48. https://doi.org/10.18637/jss.v067.i01
Bauhus J, Puettmann K, Messier C (2009) Silviculture for old-growth attributes. For Ecol Manag 258(4):525–537. https://doi.org/10.1016/j.foreco.2009.01.053
Bédard S, Guillemette F, Raymond P, Tremblay S, Larouche C, DeBlois J (2014) Rehabilitation of northern hardwood stands using multicohort silvicultural scenarios in Québec. J For 112(3):276–286. https://doi.org/10.5849/jof.13-035
Blarquez O, Talbot J, Paillard J, Lapointe-Elmrabti L, Pelletier N, Gates St-Pierre C (2018) Late Holocene influence of societies on the fire regime in southern Québec temperate forests. Quat Sci Rev 180:63–74. https://doi.org/10.1016/j.quascirev.2017.11.022
Bouget C, Larrieu L, Nusillard B, Parmain G (2013) In search of the best local habitat drivers for saproxylic beetle diversity in temperate deciduous forests. Biodivers Conserv 22(9):2111–2130. https://doi.org/10.1007/s10531-013-0531-3
Boulet B (2005) Défauts externes et indices de la carie des arbres. Ministère des Ressources Naturelles et de la Faune du Québec, Québec, p 308
Burrascano S, de Andrade RB, Paillet Y, Ódor P, Antonini G, Bouget C, Campagnaro T, Gosselin F, Janssen P, Persiani AM, Nascimbene J, Sabatini FM, Sitzia T, Blasi C (2018) Congruence across taxa and spatial scales: are we asking too much of species data? Glob Ecol Biogeogr 27(8):980–990. https://doi.org/10.1111/geb.12766
Chung Y, Rabe-Hesketh S, Dorie V, Gelman A, Liu J (2013) A nondegenerate penalized likelihood estimator for variance parameters in multilevel models. Psychometrika 78(4):685–709. https://doi.org/10.1007/s11336-013-9328-2
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18(1):117–143. https://doi.org/10.1111/j.1442-9993.1993.tb00438.x
Courbaud B, Pupin C, Letort A, Cabanettes A, Larrieu L (2017) Modelling the probability of microhabitat formation on trees using cross-sectional data. Methods Ecol Evol 8(10):1347–1359. https://doi.org/10.1111/2041-210X.12773
Dormann CF, McPherson JM, Araújo MB, Bivand R, Bolliger J, Carl G, Davies GR, Hirzel A, Jetz W, Kissling WD, Kuhn I, Ohlemuller R, Peres-Neto PR, Reineking B, Schroder B, Schurr FM, Wilson R (2007) Methods to account for spatial autocorrelation in the analysis of species distributional data: a review. Ecography (Cop) 30(5):609–628. https://doi.org/10.1111/j.2007.0906-7590.05171.x
Dufour-Pelletier S, Tremblay JA, Hébert C, Lachat T, Ibarzabal J (2020) Testing the effect of snag and cavity supply on deadwood-associated species in a managed boreal forest. Forests 11(424):1–17
Dunn OJ (1964) Multiple comparisons using rank sums. Technometrics 6(3):241–252. https://doi.org/10.1080/00401706.1964.10490181
Emberger C, Larrieu L, Gonin P (2013) Dix facteurs clés pour la diversité des espèces en forêt. Comprendre l’Indice de Biodiversité Potentielle (IBP). Document technique. Institut pour le développement forestier, Paris, p 56
Eyvindson K, Du R, Triviño M, Blattert C, Potterf M (2021) High boreal forest multifunctionality requires continuous cover forestry as a dominant management. Land Use Policy 100:104918. https://doi.org/10.1016/j.landusepol.2020.104918
Fielding AH, Belll JF (1997) A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ Conserv 24(1):38–49. https://doi.org/10.1017/S0376892900021214
Gaffield C (1994) Histoire de l’Outaouais. Institut québécois de recherche sur la culture, Québec, p 876
Gosselin J (2002) Guide de reconnaissance des types écologiques des régions écologiques 3a – Collines de l’Outaouais et du Témiscamingue et 3b – Collines du lac Nominingue, Ministère des Ressources naturelles du Québec, Forêt Québec, Direction des inventaires forestiers, Division de la classification écologique et de la productivité des stations, p 188
Großmann J, Pyttel P, Bauhus J, Lecigne B, Messier C (2020) The benefits of tree wounds: microhabitat development in urban trees as affected by intensive tree maintenance. Sci Total Environ 55:126817. https://doi.org/10.1016/j.ufug.2020.126817
Guillemette F, Bédard S, Fortin M (2008) Evaluation of a tree classification system in relation to mortality risk in Québec northern hardwoods. For Chron 84(6):886–899. https://doi.org/10.5558/tfc84886-6
Harting F (2019) DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. https://cran.r-project.org/package=DHARMa. Accessed 24 Aug 2020
Havreljuk F, Achim A, Auty D, Bédard S, Pothier D (2014) Integrating standing value estimations into tree marking guidelines to meet wood supply objectives. Can J For Res 44(7):750–759. https://doi.org/10.1139/cjfr-2013-0407
Jahed RR, Kavousi MR, Farashiani ME (2020) A comparison of the formation rates and composition of tree-related microhabitats in beech-dominated primeval Carpathian and Hyrcanian forests. Forests 11(144):1–13. https://doi.org/10.3390/f11020144
Kahler HA, Anderson JT (2006) Tree cavity resources for dependent cavity- using wildlife in West Virginia forests. North J Appl For 23(2):114–121. https://doi.org/10.1093/njaf/23.2.114
Kane VR, Gersonde RF, Lutz JA, McGaughey RJ, Bakker JD, Franklin JF (2011) Patch dynamics and the development of structural and spatial heterogeneity in Pacific northwest forests. Can J For Res 41(12):2276–2291. https://doi.org/10.1139/x11-128
Kenefic LS, Sendak PE, Brissette JC (2005) Comparison of fixed diameter-limit and selection cutting in northern conifers. North J Appl For 22(2):77–84. https://doi.org/10.1093/njaf/22.2.77
Kozák D, Mikoláš M, Svitok M, Bače R, Paillet Y, Larrieu L, Nagel TA, Begovič K, Čada V, Diku A, Frankovic M, Janda P, Kameniar O, Keren S, Kjucukov P, Labusova J, Langbehn T, Malek J, Mikac S, Morrissey RC, Novakova MH, Schurrman JS, Svobodov K, Synek M, Teodosiu M, Toromani E, Trotsiuk V, Vitkova L, Svoboda M (2018) Profile of tree-related microhabitats in European primary beech-dominated forests. For Ecol Manag 429(July):363–374. https://doi.org/10.1016/j.foreco.2018.07.021
Larrieu L, Cabanettes A (2012) Species, live status, and diameter are important tree features for diversity and abundance of tree microhabitats in subnatural montane beech-fir forests. Can J For Res 42(8):1433–1445. https://doi.org/10.1139/X2012-077
Larrieu L, Cabanettes A, Brin A, Bouget C, Deconchat M (2014) Tree microhabitats at the stand scale in montane beech-fir forests: practical information for taxa conservation in forestry. Eur J For Res 133(2):355–367. https://doi.org/10.1007/s10342-013-0767-1
Larrieu L, Cabanettes A, Delarue A (2012) Impact of silviculture on dead wood and on the distribution and frequency of tree microhabitats in montane beech-fir forests of the Pyrenees. Eur J For Res 131(3):773–786. https://doi.org/10.1007/s10342-011-0551-z
Larrieu L, Gosselin F, Archaux F, Chevalier R, Corriol G, Dauffy-Richard E, Deconchat M, Gosselin M, Ladet S, Savoie JM, Tillon L, Bouget C (2018b) Cost-efficiency of cross-taxon surrogates in temperate forests. Ecol Indic 87:56–65. https://doi.org/10.1016/j.ecolind.2017.12.044
Larrieu L, Gosselin F, Archaux F, Chevalier R, Corriol G, Dauffy-Richard E, Deconchat M, Gosselin M, Ladet S, Savoie JM, Tillon L, Bouget C (2019) Assessing the potential of routine stand variables from multi-taxon data as habitat surrogates in European temperate forests. Ecol Indic 104:116–126. https://doi.org/10.1016/j.ecolind.2019.04.085
Larrieu L, Paillet Y, Winter S, Bütler R, Kraus D, Krumm F, Lachat T, Michel AK, Regnery B, Vandekerkhove K (2018a) Tree related microhabitats in temperate and Mediterranean European forests: a hierarchical typology for inventory standardization. Ecol Indic 84:194–207. https://doi.org/10.1016/j.ecolind.2017.08.051
Lê S, Josse J, Husson F (2008) FactoMineR: an R package for multivariate analysis. J Stat Softw 25(1):1–18. https://doi.org/10.18637/jss.v025.i01
Lussier J-M, Meek P (2014) Multiple-treatment irregular shelterwood method. J For 112(3):287–295. https://doi.org/10.5849/jof.13-041
Mangiafico S (2016) Summary and analysis of extension program evalution in R version 1.18.1. New Brunswick: Rutgers cooperative extension
Mangiafico S (2019) Rcompanion: functions to support extension education program evaluation. R package version 2.3.7. https://cran.r-project.org/package=rcompanion. Accessed 24 Aug 2020
Martin K, Aitken KEH, Wiebe KL (2004) Nest sites and nest webs for cavity-nesting communities in interior British Columbia, Canada: nest characteristics and niche partitioning. Condor 106(1):5. https://doi.org/10.1650/7482
Martin M, Raymond P (2019) Assessing tree-related microhabitat retention according to a harvest gradient using tree-defect surveys as proxies in eastern Canadian mixedwood forests. For Chron 95(3):157–170
Michel AK, Winter S (2009) Tree microhabitat structures as indicators of biodiversity in Douglas-fir forests of different stand ages and management histories in the Pacific northwest, U.S.A. For Ecol Manag 257(6):1453–1464. https://doi.org/10.1016/j.foreco.2008.11.027
Michel AK, Winter S, Linde A (2011) The effect of tree dimension on the diversity of bark microhabitat structures and bark use in Douglas-fir (Pseudotsuga menziesii var. menziesii). Can J For Res 41:300–308. https://doi.org/10.1139/X10-207
Ministère des Forêts de la Faune et des Parcs (MFFP) (2020) Évolution de la maladie corticale du hêtre au Québec. Direction de la protection des forêts, Québec, Gouvernement du Québec, p 14
Ministère des Forêts de la Faune et des Parcs (MFFP) (2017) Intégration des enjeux écologiques dans les plans d’aménagement forestier intégré de 2018–2023, Cahier 5.1 - Enjeux liés aux attributs de structure interne des peuplements et au bois mort. Direction de l’aménagement et de l’environnement forestiers, Québec, p 66
Ministère des Ressources Naturelles et de la Faune du Québec (MRNF) (2008) Norme de stratification écoforestière. Quatrième inventaire forestier. Direction des inventaires forestiers, Québec, p 101
Morin RS, Liebhold AM, Tobin PC, Gottschalk KW, Luzader E (2007) Spread of beech bark disease in the eastern United States and its relationship to regional forest composition. Can J For Res 37(4):726–736. https://doi.org/10.1139/X06-281
Munoz SE, Gajewski K (2010) Distinguishing prehistoric human influence on late-Holocene forests in southern Ontario, Canada. Holocene 20(6):967–981. https://doi.org/10.1177/0959683610362815
Nappi A, Drapeau P, Leduc A (2015) How important is dead wood for woodpeckers foraging in eastern north American boreal forests? For Ecol Manag 346:10–21. https://doi.org/10.1016/j.foreco.2015.02.028
Nyland RD (2016) Silviculture: concepts and applications. 3rd edit. Waveland 680
Ogle DH, Wheeler P, Dinno A (2019) FSA: fisheries stock analysis. R package version 0.8.26. https://github.com/droglenc/FSA. Accessed 24 Aug 2020
Oksanen J, Blanchet G, Friendly M, Kindt R, Legendre P, MCGlinn D, Minchin PRB, O’Hara B, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2018) Vegan: community ecology package. R package version 2.4.6. https://cran.r-project.org/package=vegan. Accessed 24 Aug 2020
Ontario Ministry of Natural Resources (OMNR) (2004) Ontario tree marking guide, version 1.1. Ontario Ministry of Natural Ressources, Toronto, p 252
Paillet Y, Archaux F, Boulanger V, Debaive N, Fuhr M, Gilg O, Gosselin F, Guilbert E (2017) Snags and large trees drive higher tree microhabitat densities in strict forest reserves. For Ecol Manag 389:176–186. https://doi.org/10.1016/j.foreco.2016.12.014
Paillet Y, Archaux F, du Puy S, Bouget C, Boulanger V, Debaive N, Gilg O, Gosselin F, Guilbert E (2018) The indicator side of tree microhabitats: a multi-taxon approach based on bats, birds and saproxylic beetles. J Appl Ecol 55(5):2147–2159. https://doi.org/10.1111/1365-2664.13181
Paillet Y, Debaive N, Archaux F, Cateau E, Gilg O, Guilbert E (2019) Nothing else matters ? Tree diameter and living status have more effects than biogeoclimatic context on microhabitat number and occurrence: an analysis in French forest reserves. PLoS One 14(5):1–18
Potapov P, Yaroshenko A, Turubanova S, Dubinin M, Laestadius L, Thies C, Aksenov D, Egorov A, Yesipova Y, Glushkov I, Karpachevskiy M, Kostikova A, Manisha A, Tsybikova E, Zhuravleva I (2008) Mapping the World’s intact forest landscapes by remote sensing. Ecol Soc 13(2):16
Pritchard KR, Hagar JC, Shaw DC (2017) Oak mistletoe (Phoradendron villosum) is linked to microhabitat availability and avian diversity in Oregon white oak (Quercus garryana) woodlands. Botany 95(3):283–294. https://doi.org/10.1139/cjb-2016-0249
Puettmann KJ, Coates KD, Messier C (2009) A critique of Silviculture: managing for complexity. Island Press, Washington D.C., p 190
Puumalainen J, Kennedy P, Folving S (2003) Monitoring forest biodiversity: a European perspective with reference to temperate and boreal forest zone. J Environ Manag 67(1):5–14. https://doi.org/10.1016/S0301-4797(02)00183-4
Puverel C, Abourachid A, Böhmer C, Leban JM, Svoboda M, Paillet Y (2019) This is my spot: what are the characteristics of the trees excavated by the black woodpecker? A case study in two managed French forests. For Ecol Manag 453:117621. https://doi.org/10.1016/j.foreco.2019.117621
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.r-project.org/. Accessed 24 Aug 2020
Raymond P, Bédard S, Roy V, Larouche C, Tremblay S (2009) The irregular shelterwood system: review, classification, and potential application to forests affected by partial disturbances. J for 405–413
Raymond P, Royo AA, Prévost M, Dumais D (2018) Assessing the single-tree and small group selection cutting system as intermediate disturbance to promote regeneration and diversity in temperate mixedwood stands. For Ecol Manag 430:21–32. https://doi.org/10.1016/j.foreco.2018.07.054
Regnery B, Paillet Y, Couvet D, Kerbiriou C (2013) Which factors influence the occurrence and density of tree microhabitats in Mediterranean oak forests? For Ecol Manag 295:118–125. https://doi.org/10.1016/j.foreco.2013.01.009
Remm J, Lõhmus A, Remm K (2006) Tree cavities in riverine forests: what determines their occurrence and use by hole-nesting passerines? For Ecol Manag 221(1–3):267–277. https://doi.org/10.1016/j.foreco.2005.10.015
Russel L (2018) Emmeans: estimated marginal means, aka least-squares means. R package version 1.3.0. https://cran.r-project.org/package=emmeans. Accessed 24 Aug 2020
Sarr DA, Puettmann K (2008) Forest management , restoration , and designer ecosystems : Integrating strategies for a crowded planet. Ecoscience 15(1):17–26, DOI: https://doi.org/10.2980/1195-6860(2008)15[17:FMRADE]2.0.CO;2
Saucier J-P, Grondin P, Brisson J, Gauthier S, Gosselin J, Lavoie C, Leduc A, Morin H, Morneau C, Payette S (2009) Écologie forestière. In: Ordre des ingénieurs forestiers du Québec (ed) Manuel de foresterie, édition revue et augmentée. Éditions Multimondes, Québec, pp 165–315
Savard S, Leblanc F, Parent A, Lesquir O (2005) Développement d’une méthode de caractérisation, de classification et de conservation d ‘arbres à vocation écologique applicable aux aménagements forestiers. Société sylvicole de la Haute-Gatineau, Messines, p 94
Senécal JF, Doyon F, Messier C (2018) Tree death not resulting in gap creation: an investigation of canopy dynamics of northern temperate deciduous forests. Remote Sens 10(1). https://doi.org/10.3390/rs10010121
Signorell A (2017) DescTools: tools for descriptive statistics. R package version 0.99.23. https://cran.r-project.org/package=DescTools. Accessed 24 Aug 2020
Sing T, Sander O, Beerenwinkel N, Lengauer T (2005) ROCR: visualizing classifier performance in R. Bioinformatics 21(20):78881. http://rocr.bioinf.mpi-sb.mpg.de. Accessed 24 Aug 2020
Stevenson SK, Jull MJ, Rogers BJ (2006) Abundance and attributes of wildlife trees and coarse woody debris at three silvicultural systems study areas in the interior cedar-hemlock zone, British Columbia. For Ecol Manag 233(1):176–191. https://doi.org/10.1016/j.foreco.2006.06.023
Swallow S, Howard R, Gutierrez R (1988) Snag preferences of woodpeckers foraging in a northeastern hardwood forest. Wilson Bull 100(2):236–246
Taylor AR, McPhee DA, Loo JA (2013) Incidence of beech bark disease resistance in the eastern Acadian forest of North America. For Chron 89(5):690–695. https://doi.org/10.5558/tfc2013-122
Tubbs CH, Houston DR (1990) Fagus grandifolia, American beech. U.S. Department of Agriculture, Forest Service, Washington, pp 325–332
Vaillancourt M-A, Drapeau P, Gauthier S, Robert M (2008) Availability of standing trees for large cavity-nesting birds in the eastern boreal forest of Québec, Canada. For Ecol Manag 255(7):2272–2285. https://doi.org/10.1016/j.foreco.2007.12.036
Villeneuve N, Brisson J (2003) Old-growth forests in the temperate deciduous zone of Quebec: identification and evaluation for conservation and research purposes. For Chron 79(3):559–569. https://doi.org/10.5558/tfc79559-3
Vuidot A, Paillet Y, Archaux F, Gosselin F (2011) Influence of tree characteristics and forest management on tree microhabitats. Biol Conserv 144(1):441–450. https://doi.org/10.1016/j.biocon.2010.09.030
Watson JEM, Evans T, Venter O, Williams B, Tulloch A, Stewart C, Thompson I, Ray JC, Murray K, Salazar A, McAlpine C, Potapov P, Walston J, Robinson JG, Painter M, Wilkie D, Filardi C, Laurance WF, Houghton RA, Maxwell S, Grantham H, Samper C, Wang S, Laestadius L, Runting RK, Silva-Chavez GA, Ervin J, Lindenmayer D (2018) The exceptional value of intact forest ecosystems. Nat Ecol Evol 2(4):599–610. https://doi.org/10.1038/s41559-018-0490-x
Winter S, Höfler J, Michel AK, Böck A, Ankerst DP (2015) Association of tree and plot characteristics with microhabitat formation in European beech and Douglas-fir forests. Eur J For Res 134(2):335–347. https://doi.org/10.1007/s10342-014-0855-x
Winter S, Möller GC (2008) Microhabitats in lowland beech forests as monitoring tool for nature conservation. For Ecol Manag 255(3–4):1251–1261. https://doi.org/10.1016/j.foreco.2007.10.029
Zweig MH, Campbell G (1993) Receiver-operating characteristic (ROC) plots: a fundamental evaluation tool in clinical medicine. Clin Chem 39(4):561–577. https://doi.org/10.1093/clinchem/39.4.561
Acknowledgments
We would like to thank Guillaume Plante for his precious help during field sampling, as well as Denise Tousignant for the linguistic revision of this manuscript. We would also like to thank the two anonymous reviewers and the associate editor, who provided accurate and relevant comments that greatly improved the quality of the manuscript.
Funding
This study was funded by the Ministère des Forêts, de la Faune et des Parcs (Quebec, Canada), project #142332185.
Author information
Authors and Affiliations
Contributions
MM, YB and PR defined the study and the sampling protocol. MM collected and prepared the data, performed the analyses and wrote the first draft of the manuscript. YB and PR discussed the results and commented on the manuscript. MM wrote the final draft of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Martin, M., Raymond, P. & Boucher, Y. Influence of individual tree characteristics, spatial structure and logging history on tree-related microhabitat occurrence in North American hardwood forests. For. Ecosyst. 8, 27 (2021). https://doi.org/10.1186/s40663-021-00305-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40663-021-00305-z