
Abstract
Background
A major challenge in downstream processing is the separation and purification of a target biomolecule from the fermentation broth which is a cocktail of various biomolecules as impurities. Aqueous two phase system (ATPS) can address this issue to a great extent so that the separation and partial purification of a target biomolecule can be integrated into a single step. In the food industry, starch production is carried out using thermostable glucoamylase. Humicola grisea serves as an attractive source for extracellular production of glucoamylase.
Results
In the present investigation, the possibility of using polyethylene glycol (PEG)/salt-based ATPS for the partitioning of glucoamylase from H. grisea was investigated for the first time. Experiments were conducted based on one variable at a time approach in which independent parameters like PEG molecular weight, type of phase-forming salt, tie line length, phase volume ratio, and neutral salt concentration were optimized. It has been found that the PEG 4000/potassium phosphate system was suitable for the extraction of glucoamylase from the fermentation broth. From the results, it was observed that, at a phase composition of 22 % w/w PEG 4000 and 12 % w/w phosphate in the presence of 2 % w/w NaCl and at pH 8, glucoamylase was partitioned into the salt-rich phase with a maximum yield of 85.81 %.
Conclusions
A range of parameters had a significant influence on aqueous two-phase extraction of glucoamylase from H. grisea. The feasibility of using aqueous two-phase extraction (ATPE) as a preliminary step for the partial purification of glucoamylase was clearly proven.
Similar content being viewed by others

Background
Glucoamylase (EC 3.2.1.3) is a hydrolytic enzyme that degrades starch and related oligosaccharides, leading to the production of β-d-glucose. Other sectors that benefit from glucoamylase include brewing, textile, food, paper, and pharmaceutical industries [1]. Glucoamylase is sourced from different microbial specimens like bacteria, yeasts, and fungi. The commercial production of glucoamylase has been mainly carried out using the genera Aspergillus and Rhizopus [2]. For the manufacture of high-fructose corn syrups, starch needs to be first converted to glucose by high-temperature liquefaction and saccharification [3]. A lot of focus is currently made on the high thermostability of glucoamylase used in the starch processing. Hence, a highly thermostable and environmentally compatible glucoamylase is very essential for industrial purposes [4]. The main benefits of using thermostable enzymes in the starch processing industry include increased reaction rates, decreased contamination risk and cost-reduction in terms of cooling system [5, 6]. The thermophilic fungus, Humicola grisea possesses an efficient hydrolytic system for the production of glucoamylase. Moreover, the enzyme is stable when exposed to high temperature for a longer duration. With regard to these advantages, glucoamylase derived from the thermophilic fungus, H. grisea MTCC 352 has been used in the current study [3].
A variety of downstream processing techniques such ion exchange chromatography, hydrophobic interaction chromatography, and gel filtration chromatography have been exploited for the purification of glucoamylase [1, 7–11]. But the flipside of these procedures is that they are expensive, time consuming, and are often multistep low-yield protocols, not suitable for large scale production. In this regard, the use of aqueous two phase systems (ATPSs) for extraction and purification of glucoamylase has been attempted in the present investigation. Aqueous two-phase extraction (ATPE) has been widely used as a rapid and economic method for the separation and partial purification of many intracellular and extracellular enzymes [12–15].
ATPS can be formulated by mixing appropriate quantity of two hydrophilic polymers or a hydrophilic polymer and a salt. However, the use of ATPS based on hydrophilic polymer and a salt has attracted many researchers because of the following advantages: ease of separation, low cost, ease of scale-up and operation, biocompatibility, and high water content. Moreover, ATPE has high capacity and yield [16]. The protein partitioning in any ATPS depends on many factors such as hydrophobic interactions, hydrogen bonding, ionic interactions, and van der Waals forces. Therefore, with respect to the type of polymer, polymer molecular weight and concentration, type of salt and concentration, tie line length (TLL), phase volume ratio (V R), and other processing parameters such as pH, temperature, and presence of neutral salt concentration, and the partitioning behavior varies [17, 18].
Over the years, ATPSs are widely used in the purification of monoclonal antibodies, extractive fermentation, and recovery of industrial enzymes [18]. Recent studies have employed the use of ATPS (polyethylene glycol (PEG)/potassium phosphate) for biomolecule extraction and primary purification, to a great extent. Nandini and Rastogi [19] dealt with the partitioning of lactoperoxidase from milk whey and studied the effect of phase-forming salt, PEG molecular weight, pH, TLL and V R, resulting in a purification-fold (PF) of 2.31. Ratanapongleka [20] studied the partitioning behavior of laccase from Lentinus polychrous Lev., to study the effect of PEG molecular weight and concentration, salt concentration, pH, and NaCl, leading to 99 % yield and PF of 3. Babu et al. [21] studied the extraction of polyphenol oxidase from pineapple and studied PEG molecular weight and concentration, salt concentration, and pH, which gave 90 % recovery and a PF of 2.7. Naganagouda and Mulimani [22] carried out ATPE of α-galactosidase from Aspergillus oryzae and studied the effect of PEG molecular weight, salt concentration, pH, and NaCl, resulting in a PF of 3.6 and recovery of 87.71 %. The portioning of glucoamylase from Aspergillus awamori NRRL 3112 was studied by Minami and Kilikian [23] using a two-step ATPE consisted of PEG/phosphate system and achieved a threefold PF. Glucoamylase from the same organism was partitioned using bioaffinity extraction with starch as a free bioligand by de Gouveia and Kilikian [24]. To the best of our knowledge, there are no available studies based on the ATPE for glucoamylase from any known thermophilic fungi.
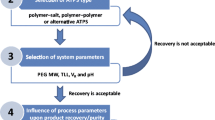
The present investigation was done to comprehend and augment the partition of glucoamylase. Accordingly, studies were systematically carried out by varying the stated parameters, through the one-variable-at-a-time approach. In the current study, the choice of the phase-forming salt was first done, followed by the molecular weight of PEG (fixing the concentration of PEG and salt at a constant level). Next, the influence of process parameters such as tie line length, phase volume ratio, and pH were investigated. Finally, the effect of the presence of a neutral salt (sodium chloride) on the partitioning behavior of glucoamylase was studied.
Methods
Materials
Polyethylene glycol (molecular weight (MW) 1000, 2000, 4000, and 6000), dipotassium hydrogen orthophosphate, potassium dihydrogen orthophosphate, trisodium citrate, tripotassium citrate, magnesium sulfate, magnesium sulfate heptahydrate, sodium chloride, and calcium chloride were obtained from Merck (India). Potato dextrose agar, yeast extract, and soluble starch were obtained from Hi Media Laboratories Pvt. Ltd (India). Glucose oxidase/peroxidase (GOD-POD) assay kit used was obtained from Agappe diagnostics Ltd (India). All chemicals were of analytical grade. The fungi H. grisea MTCC 352 was obtained from Microbial Type Culture Collection, Chandigarh, India.
Enzyme production and preparation of crude enzyme
The microorganism was maintained on potato dextrose agar (PDA) slant, grown at 45 °C for 10 days before being stored at 4 °C. Glucoamylase was produced through submerged cultivation in a chemically defined medium. The medium consisted of 2.84 g soluble starch, 0.96 g yeast extract, 0.05 g KH2PO4, 0.24 g K2HPO4, 0.05 g NaCl, 0.05 g CaCl2, 0.19 g MgSO4.7H2O, and 0.1 mL of Vogel’s trace elements solution. The pH of the medium was adjusted to 6 [3]. Cultures were incubated with agitation at 150 rpm at 45 °C for 4 days. The fermented broth was further subjected to filtration using Whatman No. 1 filter paper. After the filtrate was centrifuged at 10,000 rpm for 10 min, the fungal mycelia were removed. The cell-free supernatant was referred to as crude enzyme and was used throughout the experiments.
Partitioning studies in aqueous two-phase system
Aqueous two-phase systems were prepared by mixing the requisite amounts of PEG and the various salts (trisodium citrate, tripotassium citrate, magnesium sulfate, and mono/dibasic potassium phosphate). The total weight of the systems was 10 g, and the crude enzyme amount was 10 % of the total system. The tubes were vigorously vortexed and centrifuged at 3000 rpm for 10 min to speed up the separation process. The phase equilibration was achieved by overnight incubation of the tubes, and the samples were withdrawn from the individual phase and then analyzed for total protein and glucoamylase activity. Without the incorporation of the enzyme, the samples were analyzed against blanks containing similar composition, to avoid interference of the phase components.
Glucoamylase activity
An appropriate amount of the crude enzyme was allowed to react with 1 % (w/v) soluble starch solution in 50 mM citrate buffer (pH 5.5), at 60 °C for 10 min. The concentration of the glucose produced was estimated by GOD-POD method using a standard glucose curve prepared under similar conditions. One unit of glucoamylase activity was defined as the amount of enzyme that releases 1 μmol of glucose from soluble starch per minute under assay conditions.
Protein assay
The total protein was estimated, as described by Bradford [25], using bovine serum albumin as a standard.
Estimation of partition parameters
The partitioning parameters in ATPS were calculated as follows.
The phase volume ratio (V R) was defined as the ratio of volume in the top phase (V T) and bottom phase (V B).
The partition coefficient for glucoamylase (K GA) was defined as the ratio of glucoamylase activity in the top phase (A T) to that in the bottom phase (A B).
The partition coefficient for total protein (K TP) was defined as the ratio of protein concentration in the top phase (C T) to that in the bottom phase (C B).
The specific activity (SA) was defined as the ratio of glucoamylase activity (A) to protein concentration (C) in the respective phases.
The purification factor (PF) was calculated by the ratio of the specific activity in the bottom phase (SAB) to the specific activity in the crude extract (SAF).
The glucoamylase yield in the bottom phase is given by the following equation.
The TLL is defined as
where C PT and C PB are the PEG concentrations (% w/w) in the top and bottom phases, respectively, and C ST and C SB are the salt concentrations (% w/w) in the top and bottom phases, respectively.
Results and discussion
The essence of ATPE lies in the differential partitioning of the target biomolecule to one phase and the contaminants to the other phase. It is this mechanism that leads to the purification of a target biomolecule. Extraction of biomolecules using ATPS could be tougher using theoretical predictions, primarily due to the fact that a complex set of parameters decide the extent of partitioning in an ATPS. They include the properties of the biomolecule (size, charge, and hydrophobicity) and the properties of the system like (i) type and concentration of phase-forming salt, (ii) concentration and molecular weight of phase-forming polymer, (iii) tie line length, (iv) phase volume ratio, (v) pH of the system, and (vi) concentration of neutral salts. Details of the selection of each of these parameters and their effect on partitioning of glucoamylase have been presented in the following sections.
Effect of phase-forming salts
Due to the significant influence of the phase-forming salt on system environment, its selection has a direct consequence on separation, concentration, and purification of a given biomolecule in ATPE [26]. In order to identify the most appropriate salt for the recovery of glucoamylase and ensure its efficient extraction, ATPE experiments were performed by incorporating a phase system of PEG (MW 4000) with four different phase-forming salts such as trisodium citrate, tripotassium citrate, magnesium sulfate, and mono/dibasic potassium phosphate. The partition coefficients of H. grisea-derived glucoamylase and total protein using 15 % (w/w) PEG 4000 + 15 % (w/w) salt are shown in Fig. 1. In all the phase systems studied, it was observed that the values of K GA and K TP obtained from all of the systems were lesser than 1. Glucoamylase was preferentially partitioned to the bottom phase that indicated a strong preference of glucoamylase to the bottom phase. It resulted in low partition coefficients in the range of 0.28–0.78. This is in agreement with Minami and Kilikian [23]. ATPE studies on glucoamylase from A. awamori. The disparities in the values of K GA in the partitioning process are caused by the non-uniform distribution of the salt ions in the top and bottom phases. It is also due to the difference in the electric potential that improves protein mobility to the other phase by electrostatic repulsion/attraction, hydrophobicity, and size of the salt ions [18, 27]. The specific activity of the top and bottom phases and yield and purification factor for the bottom phase in the systems with different phase-forming salts are portrayed in Table 1. It was noted that the yield was higher in the bottom phase (69.73–82.48 %) for all the phase-forming salts. Except for the magnesium salt, it was observed that the specific activity (U/mg) was higher in the bottom phase and so was the purification factor, for rest of the salts. Trisodium citrate and potassium phosphate exhibited relatively higher yield of 82.23 & 82.48% respectively. However, based on the purification factor, potassium phosphate resulted in a higher PF (1.46). Potassium phosphate system was recognized to be more effective for lactoperoxidase [19], laccase [20], polyphenol oxidase [21], and α-galactosidase [22]. Based on the preliminary results, PEG/potassium phosphate (K2HPO4 and KH2PO4) system was used for further studies.

Effect of phase-forming salt on the partitioning of glucoamylase
Effect of PEG molecular weight
The molecular weight of PEG decides the extent of partition of the target biomolecules and the other molecules in the extract. As the chain length of PEG increases, the volume exclusion effect generally follows an increasing trend. In the presence of salt, the hydrophobicity of the polymer-rich top phase increases with a rise in chain length [18]. The extraction efficiency is influenced by the composition of phases and the number of polymer–protein interactions. These factors are governed by the polymer, possessing different degrees of polymerization [28]. In order to attain the most appropriate molecular weight of PEG for the recovery of glucoamylase, partitioning studies were carried out by employing PEG/KH2PO4 K2HPO4 system with different molecular weights of PEG (1000, 2000, 4000, and 6000). The concentration of the phase composition and pH was maintained at a constant value throughout (15 % (w/w) PEG 4000 + 15 % (w/w) potassium phosphate, pH 7). The partition coefficients of glucoamylase and total protein are shown in Fig. 2. The partition coefficients of glucoamylase (K e) and total protein (K p) were found to decrease with an increase in PEG molecular weight. The decrease in partition coefficient of glucoamylase and total proteins could be ascribed to the effect of volume exclusion, which increases with an increase in molecular weight of the polymer. As a result, the biomolecules will selectively partition to the bottom phase. Similar results were observed by Nandini and Rastogi [26], Priyanka et al. [29], and Lakshmi et al. [30]. The specific activity of the top and bottom phases and yield and purification factor for the bottom phase in the systems with different PEGs (MW 1000, 2000, 4000, and 6000) are shown in Table 2. From the experimental runs, we observed that the specific activity of the bottom phase was greater than that of the top phase, irrespective of the molecular weight of the polymer. With a rise in the molecular weight of PEG, the yield of glucoamylase in the bottom phase was on the increasing mode. This trend can be explained due to the increase of the top phase hydrophobicity. As there is an increase in the chain length of PEG, it experiences a deficiency in hydroxyl groups for the same concentration of the polymer [12]. The specific activity of the enzyme in the bottom phase rose up to the stage when the molecular weight increased from 1000 to 4000. Thereafter, with PEG 6000, it took a dip; so was the case with the purification factor. This behavior is due to the fact that the bottom phase may be reaching the solubility limit with respect to glucoamylase. The resulting salting-out effect, therefore, tends to push the enzyme to the top phase. Similar results were observed by Yuzugullu and Duman [31] and Madhusudhan and Raghavarao [32]. Thus, it was observed that the transition of yield and purification factor towards a higher value was observed at PEG 4000. Based on this fact, PEG 4000 was chosen for further studies.

Effect of PEG molecular weight on the partitioning of glucoamylase
Effect of TLL
The effect of TLL (22.91–31.61 %) on glucoamylase partitioning was investigated in PEG 4000/potassium phosphate systems. The composition of the PEG-salt system within the specified TLL range was obtained from the liquid-liquid equilibrium data, as provided by Carvalho et al. [33]. The phase volume ratio was maintained at 1 for these set of experiments. It was observed that the partition coefficient values of both glucoamylase and total protein increased with an increase in TLL (Fig. 3). This could be because of the decrease in the relative free volume in the bottom phase and subsequent decrease in the solubility of the biomolecules [32, 34]. As depicted in the Fig. 4, the increase in the partition coefficient of glucoamylase with the increase in TLL at constant volume ratio results in the decrease in glucoamylase yield (Eq. 6). The purification factor increased and reached a maximum value of 1.72 at TLL of 30.62 %. A decrease in the purification factor value was observed for further increase in TLL, and this may be due to the high salt concentration at the bottom phase which affects the solubility of glucoamylase [30].

Effect of TLL on the partitioning of glucoamylase

Effect of TLL on the recovery of glucoamylase
Effect of phase volume ratio
In order to further purify the enzyme, various volume ratios (0.41–1.57) were selected on the TLL of 30.62 % and the consecutive effect of this on PF and yield was investigated. It can be evidenced from Fig. 5 that the increase in phase volume ratio increases the yield owing to the reduction in the bottom phase volume [32]. A lower PF was observed at lower V R due to the fact that a larger volume of bottom phase at the lower V R promotes the partitioning of the contaminant proteins to the bottom phase. A maximum PF of 1.84 was observed at a V R of 1.37 and further increment resulted in a decrease in PF. In contrast to this, the yield decreased with the increase in phase volume ratio. Similar result was observed by Chethana et al. [35].

Effect of V R on the recovery of glucoamylase
Effect of pH
One of the significant factors that govern the partition behavior of biomolecules in an ATPS is the pH at which the process is carried out. Any change in pH has the ability to influence the charge of the solute or the ratio of the charged molecules. The phase system selected from the previous step (22 % (w/w) PEG 4000 and 12 % (w/w) potassium phosphate) was further subjected to pH changes from 6 to 9. The variation of partition coefficients with respect to the pH of the system is shown in Fig. 6. The increase in pH improved the migration of contaminant proteins to the top phase and consequently the partition coefficient of glucoamylase was found to decrease. This phenomenon enhanced the purification factor and yield at the bottom phase. The results are in accordance with reported literature (Nandini and Rastogi [19] and Naganagouda and Mulimani [22]). It can be visualized from Fig. 7 that the increase in pH has a positive effect on glucoamylase yield and it reached a maximum yield of 82.62 % at a pH of 9. But the PF reached a maximum of 2.61 at a pH of 8 and decreased further. The low stability of glucoamylase at higher pH could be a possible reason for this reduction [4]. It is a well-known fact that the pH of the ATPS has a profound effect on the partitioning of biomolecules since it may change the charge of the biomolecule or the ratio of the charged biomolecules. The partitioning depends on the system pH and the isoelectric point of glucoamylase. The literature reveals that the isoelectric point of glucoamylase from H. grisea is greater than 8 [8, 36, 37]. The decrease in pH makes the glucoamylase more positively charged and leads to stronger interaction between glucoamylase and polymer which migrated more enzymes to the PEG-rich phase. Similar results were observed by Nandini and Rastogi [19] and Ratanapongleka [20].

Effect of pH on the partitioning of glucoamylase

Effect of pH on the recovery of glucoamylase
Effect of NaCl
One of the definitive methods to arrive at an optimum value of the selectivity and yield has been the addition of neutral salts to the ATPS [16]. With a view to examine the effect of a neutral salt on the partitioning of the enzyme, the concentration NaCl was varied from 0 to 5 % w/w in the selected system from the previous step (22 % w/w PEG 4000 12 % w/w phosphate system, pH 8.0). In general, the addition of neutral salts to the ATPS changes the partitioning behavior of protein by changing the electrostatic potential difference between the phases or by increasing the hydrophobic interactions [38]. Because of the changes in the electrostatic potential difference, the increase in NaCl concentration promoted more partitioning of glucoamylase to the bottom phase and a lowest partition coefficient of 0.126 was obtained at 2 % NaCl concentration (Fig. 8). This system resulted in a PF of 2.68 and a yield of 85.81 %. As shown in Fig. 8, the further increase in NaCl concentration caused the reduction of PF which could be as a consequence of increase in hydrophobic interactions between the protein and PEG in the top phase [39].

Effect of NaCl on the recovery of glucoamylase
Based on the above observations, it is clear that PEG 4000 and KH2PO4/K2HPO4 phase system can be used as a potential technique for the separation and partial purification of glucoamylase.
Conclusion
The recovery of glucoamylase from thermophilic fungal sources using aqueous two-phase extraction was reported for the first time. The influence of various parameters on separation and partial purification of glucoamylase from H. grisea in aqueous two-phase systems was revealed. The PEG 4000/potassium phosphate phase system was found to be the most efficient for the extraction of glucoamylase, when compared to other salt systems. It was noted that glucoamylase preferentially partitioned to the salt-rich bottom phase. The optimized conditions of tie line length were at 30.62 %, phase volume ratio 0.53, pH 8, and 2 % w/w NaCl. The said conditions provided a maximum yield of 85.81 % and purity of 2.68-fold compared to crude extract. Overall, the results demonstrated the feasibility of using ATPE as a preliminary step for the partial purification of glucoamylase.
References
Riaz M, Perveen R, Javed MR, Nadeem HU, Rashid MH (2007) Kinetic and thermodynamic properties of novel glucoamylase from Humicola sp. Enzyme Microb Tech 41:558–564
Pandey A (1995) Glucoamylase research: an overview. Starch 47:439–445
Ramesh V, Murty VR (2014) Sequential statistical optimization of media components for the production of glucoamylase by thermophilic fungus Humicola grisea MTCC 352. Enzyme Res. http://www.hindawi.com/journals/er/2014/317940/
Gomes E, Souza SR, Grandi RP, Da Silva R (2005) Production of thermostable glucoamylase by Aspergillus flavus A 1.1 and Thermomyces Lanuginosus A 13.37. Braz J Microbiol 36:75–82
Kaur P, Satyanarayana T (2004) Production and starch saccharification by a thermostable and neutral glucoamylase of a thermophilic mould Thermomucor indicae-seudaticae. World J Microbiol Biotechnol 20:419–425
Koç O, Metin K (2010) Purification and characterization of a thermostable glucoamylase produced by Aspergillus flavus HBF34. African J Biotechnol 9(23):3414–3424
Ferreira-Nozawa MS, Rezende JL, Guimarães LHS, Terenzi HF, Jorge JA, Polizeli MLTM (2008) Mycelial glucoamylases produced by the thermophilic fungus Scytalidium thermophilum strains 15.1 and 15.8. Purification and biochemical characterization. Braz J Microbiol 39(2):344–352
Campos L, Felix CR (1995) Purification and characterization of a glucoamylase from Humicola grisea. Appl Env Microbiol 61(6):2436–2438
Nguyen QD, Rezessy-Szabó JM, Claeyssens M, Stals I, Hoschke A (2002) Purification and characterization of amylolytic enzymes from thermophilic fungus Thermomyces lanuginosus strain ATCC 34626. Enzyme Microb Tech 31:345–352
Thorsen TS, Johnsen AH, Josefsen K, Jensen B (2006) Identification and characterization of glucoamylase from the fungus Thermomyces lanuginosus. Biochim Biophys Acta 1764(4):671–676
Negi S, Gupta S, Banerjee R (2011) Extraction and purification of glucoamylase and protease produced by Aspergillus awamori in a single-stage fermentation. Food Technol Biotechnol 49:310–315
Gautam S, Simon L (2006) Partitioning of β-glucosidase from Trichoderma reesei in poly(ethylene glycol) and potassium phosphate aqueous two-phase systems: influence of pH and temperature. Biochem Eng J 30:104–108
Madhusudhan MC, Raghavarao KSMS, Nene S (2008) Integrated process for extraction and purification of alcohol dehydrogenase from baker’s yeast involving precipitation and aqueous two phase extraction. Biochem Eng J 38:414–420
Kammoun R, Chouayekh H, Abid H, Naili B, Bejar S (2009) Purification of CBS 819.72 -amylase by aqueous two-phase systems: modelling using response surface methodology. Biochem Eng J 46:306–312
Kianmehr A, Pooraskari M, Mousavikoodehi B, Mostafavi SS (2014) Recombinant D-galactose dehydrogenase partitioning in aqueous two-phase systems: effect of pH and concentration of PEG and ammonium sulfate. Bioresource Bioprocess 1:6
Albertsson PA (1987) Partitioning of cell particles and macromolecules, 3rd edn. New York, John Wiley and Sons
Benavides J, Rito-Palomares M (2008) Practical experiences from the development of aqueous two-phase processes for the recovery of high value biological products. J Chem Technol Biotechnol 83:133–142
Raja S, Murty VR, Thivaharan V, Rajasekar V, Ramesh V (2011) Aqueous two phase systems for the recovery of biomolecules—a review. Science Technol 1:7–16
Nandini KE, Rastogi NK (2011) Integrated downstream processing of lactoperoxidase from milk whey involving aqueous two-phase extraction and ultrasound-assisted ultrafiltration. Appl Biochem Biotechnol 163:173–185
Ratanapongleka K (2012) Partitioning behavior of laccase from Lentinus polychrous Lev in aqueous two phase systems. Songklanakarin J Sci Technol 34(1):69–76
Babu BR, Rastogi NK, Raghavarao KSMS (2008) Liquid–liquid extraction of bromelain and polyphenol oxidase using aqueous two-phase system. Chem Eng Process 47:83–89
Naganagouda K, Mulimani VH (2008) Aqueous two-phase extraction (ATPE): an attractive and economically viable technology for downstream processing of Aspergillus oryzae α-galactosidase. Process Biochem 43:1293–1299
Minami NM, Kilikian BV (1998) Separation and purification of glucoamylase in aqueous two-phase systems by a two-step extraction. J Chromatogr B 711:309–312
de Gouveia T, Kilikian BV (2000) Bioaffinity extraction of glucoamylase in aqueous two-phase systems using starch as free bioligand. J Chromatogr B 743:241–246
Bradford MM (1976) A rapid and sensitive for the quantitation of microgram quantitites of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Nandini KE, Rastogi NK (2011) Liquid–liquid extraction of lipase using aqueous two-phase system. Food Bioprocess Technol 4:295–303
Nagaraja VH, Iyyaswami R (2015) Aqueous two phase partitioning of fish proteins: partitioning studies and ATPS evaluation. J Food Sci Technol 52(6):3539–3548. http://www.ncbi.nlm.nih.gov/pubmed/26028736
Mohamadi HS, Omidinia E (2007) Purification of recombinant phenylalanine dehydrogenase by partitioning in aqueous two-phase systems. J Chromatogr B 854:273–278
Priyanka BS, Rastogi NK, Raghavarao KSMS, Thakur MS (2012) Downstream processing of luciferase from fireflies (Photinus pyralis) using aqueous two-phase extraction. Process Biochem 47:1358–1363
Lakshmi MC, Madhusudhan MC, Raghavarao KSMS (2012) Extraction and purification of lipoxygenase from soybean using aqueous two-phase system. Food Bioprocess Technol 5:193–199
Yuzugullu Y, Duman YA (2015) Aqueous two-phase (PEG4000/Na2SO4) extraction and characterization of an acid invertase from potato tuber (Solanum tuberosum). Prep Biochem Biotechnol 45(7):696–711. http://www.ncbi.nlm.nih.gov/pubmed/25127162
Madhusudhan MC, Raghavarao KSMS (2011) Aqueous two phase extraction of invertase from baker’s yeast: effect of process parameters on partitioning. Process Biochem 46:2014–2020
Carvalho CP, Coimbra JSR, Costa IAF, Minim LA, Silva LHM, Maffia MC (2007) Equilibrium data for PEG 4000 plus salt plus water systems from (278.15 to 318.15) K. J Chem Eng Data 52:351–356
Selvakumar P, Ling TC, Walker S, Lyddiatt A (2012) Recovery of glyceraldehyde 3-phosphate dehydrogenase from an unclarified disrupted yeast using aqueous two-phase systems facilitated by distribution analysis of radiolabelled analytes. Sep Purif Technol 85:28–34
Chethana S, Nayak CA, Raghavarao KSMS (2007) Aqueous two phase extraction for purification and concentration of betalains. J Food Eng 81:679–687
Cereia M, Guimaraes LHS, Nogueira SCP, Jorge JA, Terenzi HF, Greene LJ, Polieli MLTM (2006) Glucoamylase isoform (GAII) purified from a thermophilic fungus Scytalidium thermophilum 15.8 with biotechnological potential. African J Biotechnol 5(12):1239–1245
Aquino ACMM, Jorge JA, Terenzi HF, Polizeli MLTM (2001) Thermostable glucose-tolerant glucoamylase produced by thermophilic fungus Scytalidiuyem thermophilum. Folia Microbiol 46(1):11–16
Kavakçıoğlu B, Tarhan L (2013) Initial purification of catalase from Phanerochaete chrysosporium by partitioning in poly(ethylene glycol)/salt aqueous two phase systems. Sep Purif Technol 105:8–14
Raja S, Murty VR (2013) Optimization of aqueous two-phase systems for the recovery of soluble proteins from tannery wastewater using response surface methodology. J Eng. http://www.hindawi.com/journals/je/2013/217483/
Acknowledgment
The authors gratefully acknowledge the Department of Biotechnology, MIT, Manipal University for providing the facilities to carry out the research work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
Both the authors have active participation in the implementation and analysis of the present study. Ramesh performed the research protocols and wrote the manuscript. Both the authors have read and approved the final version of the manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0), which permits use, duplication, adaptation, distribution, and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Ramesh, V., Murty, V.R. Partitioning of thermostable glucoamylase in polyethyleneglycol/salt aqueous two-phase system. Bioresour. Bioprocess. 2, 25 (2015). https://doi.org/10.1186/s40643-015-0056-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40643-015-0056-6