
Abstract
Root-associated symbiotic fungi can enhance the host plant’s ability to tolerate adverse environmental conditions, but it’s unclear whether and how they enhance waterlogged peach plants. This study aimed to investigate the effects of Funneliformis mosseae (Fm), an arbuscular mycorrhizal fungus, and Serendipita indica (Si), an endophytic fungus, on plant growth, root development, reactive oxygen species (ROS) levels, total flavonoids, ROS scavenging activity of flavonoids, and activities and gene expression of antioxidant enzymes in roots of waterlogged peach seedlings. Although waterlogging treatment had little effect on root development, Fm and Si, particularly Fm, improved root development variables to varying degrees. Waterlogging significantly promoted superoxide anion radicals (O2•−) and hydrogen peroxide (H2O2) levels, whereas Fm and Si distinctly reduced O2•− and H2O2 levels under waterlogging. Waterlogging triggered an increase in total flavonoids in fungi-inoculated seedlings, and the two fungi also significantly increased total flavonoid concentrations and the scavenging activity of total flavonoids to hydroxyl radical, O2•−, and 2,2-diphenyl-1-picrylhydrazyl radical. In addition, root fungal colonization rate was negatively and significantly correlated with O2•− and H2O2, but positively with total flavonoids. Under waterlogging conditions, Fm significantly raised root superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPX) activities, and Si likewise elevated root SOD, CAT, GPX, and peroxidase (POD) activities, compared with uninoculated treatment. Although the expression of various antioxidant enzyme genes in roots remained mostly unchanged or were inhibited by the waterlogging, Fm up-regulated the expression of PpSOD3 and PpSOD7, whereas Si up-regulated the expression of PpSOD3, PpCAT1, PpGPX6, PpGPX8, and PpPOD1. In conclusion, low oxidative bursts in Fm- and Si-inoculated peach plants exposed to waterlogging were associated with an increase in total flavonoids as well as activities and gene expression of antioxidant enzymes.
Graphical Abstract

Similar content being viewed by others
Introduction
Peach (Prunus persica) is a high-value fruit tree, and its fruits are abundant in phenolic and other compounds with free radical scavenging and antioxidant properties [1]. Peaches are sensitive to soil waterlogging because of their shallow roots and high oxygen (O2) consumption [2]. As a result of inadequate drainage, improper irrigation, and heavy rainfall in the orchard, peaches are vulnerable to soil waterlogging [3]. Therefore, it is essential and pressing the key to enhance peach waterlogging tolerance.
Waterlogging stress has an impact on hindering plant growth and development and even death [4]. Under soil waterlogging conditions, plants switch from aerobic respiration to anaerobic respiration due to a lack of O2 supply, leading to a dramatic decrease in energy production and over accumulation of toxic substances [5]. Waterlogging hampered intracellular electrolyte transport and metabolism of reactive oxygen species (ROS), thus inhibiting the healthy growth of plants [6]. Waterlogging stress resulted in elevated superoxide anion radical (O2•−) and hydrogen peroxide (H2O2) levels in leaves and roots of six peach rootstocks, thereby, triggering the oxidative damage to peaches [2, 7]. In fact, antioxidant defense systems are a crucial mechanism for plants to resist ROS bursts under stress conditions [8]. Antioxidant defense systems consist of antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPX), and peroxidase (POD), as well as non-enzymatic antioxidants like glutathione, ascorbic acid, tocopherol, and flavonoids [9]. Among them, flavonoids, which are abundant in nature, have gained substantial interest due to their ability to effectively eliminate excessive ROS in stressed plants [10, 11]. A study conducted by Su et al. [12] demonstrated that total flavonoids extracted from Rosa laevigata plants exhibited excellent scavenging activity against hydroxyl radicals and O2•−. Liu et al. [13] reported that inoculation with Funneliformis mosseae (Fm) increased the scavenging activity of total flavonoids to hydroxyl radicals and O2•− in roots of trifoliate orange under drought, thus maintaining a low oxidative burst in inoculated plants. It can thus gain a better understanding of the total flavonoid role in stress-triggered oxidative burst by analyzing the change in total flavonoids and their scavenging activity to ROS.
Some fungi, such as arbuscular mycorrhizal fungi (AMF), can establish symbiotic associations with plant roots [14]. AMF are beneficial fungi widely inhabiting in the soil and can form symbioses with over 80% of terrestrial plants [15, 16]. This symbiosis helps host plants absorb nutrients and water to promote plant growth through extraradical hyphae [17]. Additionally, it can accelerate photosynthetic capacity, accumulation of osmotic substances, and enhancement of antioxidant defense systems, thus improving the resistance of host plants to stressful environments [18]. AMF populations are discovered in soil waterlogging environments, including wetlands [19]. Transcriptome sequencing investigations have shown that AMF inoculation under waterlogging conditions up-regulated the expression of genes associated with antioxidant defense systems in Phragmites australis plants, thus enhancing their resistance to waterlogging [20]. Nevertheless, in Citrus junos, AMF elevated CAT activity only in leaves, not roots, under waterlogging conditions [21]. In Vigna unguiculata plants, inoculating Glomus intraradices under drought stress led to significant increases in flavonoids, total phenolic compounds, sugars, and 2,2-diphenyl-1-picrylhydrazyl radical (DPPH) scavenging activity, therefore enhancing the host plant’s antioxidant defenses [22].
In addition, Serendipita indica (Sav. Verma, Aj. Varma, Rexer, G. Kost & P. Franken) M. Weiß, Waller, A. Zuccaro & Selosse (Si) is a root-associated symbiotic fungus that shares similar functions, such as enhanced stress tolerance, with AMF [23]. However, unlike AMF, Si can be propagated in vitro without the need for plants [24]. Walnut plants colonized by Si showed better drought tolerance than non-Si-colonized plants [25]. This could be attributed to the fact that Si-inoculated plants had enhanced antioxidant enzyme activities, which helped mitigate oxidative damage caused by ROS [9]. Another study demonstrated that Si enhanced waterlogging tolerance in Chinese cabbage plants by increasing POD and CAT activities [26]. However, Si hampered CAT activity in wheat and both CAT and APX activities in maize under soil drought conditions [27, 28]. These findings suggest that the effect of Si on host stress resistance may vary depending on the specific stress and plant species. It is unknown whether and how Si enhances waterlogging tolerance in peach plants.
Based on these previous findings, we, thus, hypothesized that inoculation with root-associated symbiotic fungi, such as AMF and Si, could mitigate oxidative damage caused by ROS in peach roots by promoting activities and gene expression of antioxidant enzymes and the accumulation of flavonoids, ultimately enhancing the plants’ resistance to waterlogging. To test this hypothesis, we conducted a potted experiment where peach seedlings were inoculated with AMF and Si and then subjected to waterlogging stress. Activities and gene expression of antioxidant enzymes, total flavonoid levels, and their ROS scavenging activity were analyzed.
Materials and methods
Plant culture, fungal inoculation, and waterlogging treatment
Seeds of peach [Prunus persica (L.) Batsch cv. Maotao] were provided by Hubei Academy of Agricultural Sciences (Wuhan, China). After soaking the seeds at room temperature for 1 week, they were subjected to low-temperature stratification treatment at 4 °C in autoclaved sand for 3 months. Five-leaf peach seedlings were transplanted into plastic pots filled with 2.2 kg of autoclaved soil and sand combination by a volume ratio of 3:1 (pH 5.8, NH4+-N 41.9 mg/kg, NO3−-N 74.3 mg/kg, Olsen-P 46.2 mg/kg, and available K 497.5 mg/kg). Each pot contained two peach seedlings. At the time of transplanting, fungal inoculations were carried out. The pots received 100 g of Fm inoculums as AMF treatment (+ Fm) and 100 mL spore suspension of Si (+ Si) as endophytic fungal treatment. Among them, the biological characteristics of the mycorrhizal fungus Fm had been described by Zhang and Wang [29]. The Fm was trapped using white clover under potted conditions for 10 weeks, and the mixture of harvested root segments and growth substrate was used as mycorrhizal inoculums, along with 51 spores/g inoculum. The biological characteristics of Si had been described by Yang et al. [30]. The Si was propagated in vitro according to the method described by Yang et al. [31], and the concentration of Si spore suspension was 3.61 × 109 CFU/mL. With regard to the non-fungi treatment (-F), 100 g of autoclaved Fm inoculums and 100 mL of autoclaved spore suspension of Si were collectively inoculated into the pot [32].
During the 11 weeks of fungal inoculations, all seedlings were uniformly watered with 75% of the maximum water capacity of the substrate by weighing. For the following 2 weeks, half of the potted seedlings were subjected to waterlogging treatment (W) by placing them in a water-storage box (90 × 40 × 23 cm) with a 3 cm water level higher than the pot [32]. The remaining half of the potted seedlings were placed in the same box without water as the non-waterlogging treatment (N). During the 2 weeks of waterlogging treatment, the water level of the waterlogged box was checked twice a day at 8:00 am and 6:30 pm, and any lost water in the box was replenished to maintain the designed waterlogging strength.
Experimental design
The experiment consisted of two factors: water treatments (W and N) and fungal inoculations (-F, + Fm, and + Si). There were a total of six treatments, each with eight replicates.
Measurement of root fungal colonization, plant growth, and root development
Plant height was measured before and after waterlogging stress, and plant height growing rate was estimated during waterlogging stress. Half of the harvested plants were frozen in liquid nitrogen for gene expression analysis (n = 4), while the other half were used for physiological variable determination (n = 4). The total biomass was weighed, and the roots were immediately collected and scanned using an Epson scanner (J221A). Root development variables were analyzed using the WinRHIZO software (Regent Ltd., Quebec, Canada).
Root fungal colonization was observed under an optical microscope after staining with 0.05% trypan blue [33]. A formula described by Yang et al. [31] was used to calculate root fungal colonization rates.
Measurement of root total flavonoids and indoleacetic acid (IAA)
The extraction and determination of total flavonoids from roots were performed as per the method described by Liu et al. [13] with minor modifications. Dry root samples (0.1 g) were extracted with 10 mL 70% ethanol solution using ultrasound for 0.5 h, followed by 2 h in a water bath at 55 °C. The extraction solution was then filtered through a filter membrane, and 70% ethanol was added to bring the volume to 25 mL. The 5.4 mL of 70% ethanol and 0.3 mL of 5% NaNO2 were mixed with 2 mL of extract solution for 6 min. Subsequently, 0.3 mL 10% Al(NO3)3 and 2 mL 4% NaOH were supplemented for 27 min. The mixture was colorimetrically determined at 510 nm, using rutin as a control.
Fresh root samples (50 mg) were ground with 1.0 mL 10 mM phosphate buffer (pH 7.4), and the homogenate was centrifuged at 1000 × g for 20 min at 4 °C. The supernatant was used to analyze the root IAA concentration. Root IAA concentration was assayed as per the Enzyme-linked Immunosorbent Assay kit (Shanghai Enzyme-linked Biotechnology Co., Ltd., Shanghai, China) based on the user’s manual, and the absorbance was measured at 450 nm.
Measurement of root O2 •− and H2O2 levels and free radical scavenging activity of total flavonoids
Root O2•− concentrations were measured according to the protocol of Wang and Luo [34] with the reaction of 10 mM hydroxylamine chloride. Root H2O2 concentrations were determined using the method of Velikova et al. [35] with the reaction of 1 M potassium iodide. Fresh root samples (0.1 g) were mixed with 100 mL 70% ethanol, extracted in the dark for 24 h, and then filtered. The scavenging activity of the extracts to hydroxyl radical (•OH), O2•−, and DPPH was determined as per the protocol developed by Liu et al. [13]. Among them, the reaction solution for the determination of the •OH scavenging activity consisted of 6 mM salicylic acid, 2 mM FeSO4, the extracts, and 6 mM H2O2 at 37 °C for 30 min; for the determination of the O2•− scavenging activity, the reaction solution consisted of Tris–HCl buffers, the extracts, and 7 mM catechol solution for 5 min; in the determination of the DPPH scavenging activity of total flavonoids, the reaction solution consisting of 120 µM DPPH and the extracts was placed in the dark for 30 min.
Measurement of root antioxidant enzyme activities
Fresh root samples (0.3 g) were ground with 5 mL of 0.05 M phosphate buffer (pH 7.8) in an ice bath to form a homogenate, and then centrifuged at 10000 × g for 15 min at 4 °C. The supernatant was collected for the analysis of antioxidant enzyme activities. SOD activity was determined using the nitrogen blue tetrazole method described by Wu [36], where the reaction system consisted of 0.05 mL supernatants, 1.5 mL 0.05 M phosphate buffer (pH 7.8), 0.3 mL 130 mM methionine, 0.3 mL 750 mM nitrogen blue tetrazole, 0.3 mL 100 μM EDTA-Na, 0.3 mL 20 μM riboflavin, and 0.25 mL distilled water at 20000 Lux for 10 min. POD activity was determined using the guaiacol method described by Wu [36], in which the reaction system consisted of 0.1 mL supernatants, 2.9 mL 0.05 M phosphate buffer (pH 5.5), 1.0 mL 2% H2O2, and 1.0 mL 0.05 M guaiacol. CAT activity was determined by the colorimetric assay outlined by Wu [36], where the reaction solution consisted of 0.2 mL supernatants, 1.5 mL 0.05 M phosphate buffer (pH 7.8), 0.3 mL 0.1 M H2O2, and 1.0 mL distilled water. GPX activity was determined using the protocol of Drotar et al. [37], where the reactive solution contained 0.05 M potassium phosphate, 2 mM EDTA, 2.0 mM glutathione, 0.1 mM NADPH, 2.5 units of glutathione reductase, and 0.09 mM H2O2.
Measurement of antioxidant enzyme gene expression
According to the findings of Liu et al. [38] and Huan et al. [39], two CAT genes (PpCAT1 and PpCAT2), one POD gene (PpPOD1), two GPX genes (PpGPX6 and PpGPX8, the most abundantly expressed genes in peaches), and eight SOD genes (PpSOD1 − 8) were chosen for analysis. Primer sequences (Additional file 1: Table S1) for these genes were designed using the Primer-BLAST online resource (https://www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi). Total RNA was extracted from the roots using a Plant Total RNA Isolation kit (Polysaccharides & Polyphenolics-rich) (RK02004; Biomarker Technologies Co., Ltd., Beijing, China) and reverse transcribed to cDNA using a PrimeScript™ RT reagent kit with a gDNA eraser (PK02006; Biomarker Technologies Co., Ltd., Beijing, China). The quantity of RNA and cDNA was assessed by a spectrophotometer at 260 nm and 280 nm, respectively, and their quality was determined using 1% agarose gel electrophoresis. The qRT-PCR reaction was carried out in a CFX96 Real Time PCR Detection System (Bio-Rad, Hercules, USA) with Taq Pro Universal SYBR qPCR Master Mix (Q712; Vazyme Biotech Co., Ltd., Nanjing, China). Each assay had four biological replicates (n = 4), and TEF2 was employed as the house-keeping gene [40]. The N-F treatment (= 1) was used to normalize the relative gene expression, as described by Livak and Schmittgen [41].
Statistical analysis
Data were presented as means ± standard deviation (SD) (n = 4). Data analysis was carried out using SAS software (SAS Institute, Inc., Cary, NC, USA), including an analysis of variance (Duncan’s Multiple Range test at P = 0.05), interaction analysis, and Pearson’s correlation analysis. Histograms were generated using Sigmaplot 14.0 (Systat Software Inc., Point Richmond, CA, USA), and a heat map and a principal component analysis (PCA) were created by Origin 2022 (OriginLab Inc., Northampton, MA, USA).
Results
Changes in root fungal colonization
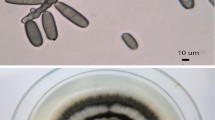
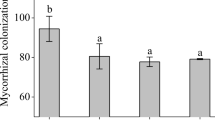
No fungal colonization was observed in the roots of the no-fungi-inoculated seedlings. However, both Fm and Si could colonize peach roots (Fig. 1a, b), with a colonization rate of 96.8–97.0% under Fm inoculation and 53.6–92.9% under Si inoculation, respectively (Table 1). Among them, compared with the N treatment, the W treatment did not have a significant impact on the Fm colonization rate, but significantly increased the Si colonization rate by 73.32%.

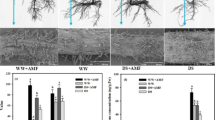
Root colonization of Funneliformis mosseae (a) and Serendipita indica (b) and root development (c) of different treated peach seedlings. -F inoculation without symbiotic fungi, + Fm inoculation with F. mosseae, + Si inoculation with S. indica, N non-waterlogging stress, W waterlogging stress
Changes in plant growth and root development
After 2 weeks of W treatment, peach plants showed distinct changes in vegetative growth performance (Additional file 1: Fig. S1), along with a 24.47% inhibition of plant height growing rate in uninoculated peach seedlings as well as no change in total biomass (Table 1). Compared with uninoculated seedlings, height growing rate was observably promoted by inoculating Fm by 26.60% under N treatment and inoculating Fm and Si by 61.97 and 52.11% respectively under W treatment. Inoculation with Fm and Si significantly increased total biomass by 42.78 and 21.84% under N conditions and 58.68 and 38.11% under W conditions, respectively, compared with no-fungi inoculation. In addition, the 2 week W treatment caused considerable alterations in root development (Fig. 1c), with uninoculated seedlings’ root volume decreasing and Si-inoculated seedlings’ root surface area and volume increasing. The inoculated seedlings developed better root development than the uninoculated treatment. Inoculating Fm significantly increased root surface area and volume by 37.82 and 69.64%, respectively, under N treatment, while it significantly increased root length, surface area, and volume by 36.9%, 77.49, and 129.11%, respectively, under W treatment. In contrast, the improvement effect of Si inoculation on root development was only under the conditions of W, as evidenced by significant increases in total length, surface area, and volume by 29.90, 62.32, and 102.53%, respectively.
Changes in root IAA levels
Compared with the N treatment, the W treatment had no significant effect on root IAA levels in uninoculated seedlings, while it significantly decreased root IAA levels in Fm-inoculated seedlings by 44.90% and increased root IAA levels in Si-inoculated seedlings by 55.17% (Fig. 2). Inoculation with Fm significantly raised root IAA levels by 48.48% under N conditions, but reduced them by 20.59% under W conditions, compared with uninoculated treatments. Inoculation with Si had no significant effect on root IAA levels under N conditions, while increased by 32.35% under W conditions, compared with uninoculated treatments.

Effects of Funneliformis mosseae and Serendipita indica on root IAA levels of peach seedlings under waterlogging stress and non-waterlogging stress. Data (means ± SD, n = 4) with different letters above the bars indicated the significant difference among treatments at 0.05 levels. See Fig. 1 for abbreviations
Changes in root O 2 •− and H 2 O 2 levels
The 2 week W treatment significantly increased root O2•− levels in uninoculated and Si-inoculated seedlings by 58.38% and 90.63%, respectively, as well as root H2O2 levels in uninoculated, Fm-inoculated, and Si-inoculated seedlings by 161.59, 35.50, and 46.55%, respectively (Fig. 3a, b). In comparison to no-fungi inoculation, inoculation with Fm and Si significantly reduced root O2•− levels by 53.66 and 41.36% under N conditions, and by 53.06 and 29.42% under W conditions, respectively. Similarly, inoculation with Fm and Si significantly reduced root H2O2 levels by 25.29 and 43.79% under N conditions, respectively, and by 61.30 and 68.51% under W conditions, respectively.

Effects of Funneliformis mosseae and Serendipita indica on root O2•− (a) and H2O2 (b) concentrations of peach seedlings under waterlogging stress and non-waterlogging stress. Data (means ± SD, n = 4) with different letters above the bars indicated the significant difference among treatments at 0.05 levels. See Fig. 1 for abbreviations
Changes in total flavonoid concentrations
Compared with the N treatment, the W treatment did not have a significant effect on root total flavonoid concentrations in uninoculated seedlings, while it significantly increased root total flavonoid concentrations in Fm- and Si-inoculated seedlings by 34.02 and 31.70%, respectively (Fig. 4). Inoculation with both Fm and Si resulted in a significant increase in root flavonoid concentrations by 16.49 and 17.84% under N conditions and by 41.07 and 40.24% under W conditions, respectively, compared with no-fungi inoculation.

Effects of Funneliformis mosseae and Serendipita indica on root total flavonoid concentrations of peach seedlings under waterlogging stress and non-waterlogging stress. Data (means ± SD, n = 4) with different letters above the bars indicated the significant difference among treatments at 0.05 levels. See Fig. 1 for abbreviations
Changes in ROS scavenging activity of total flavonoids
Compared with the N treatment, the W treatment increased the scavenging activity of ROS by root total flavonoids, especially in seedlings inoculated with symbiotic fungi at significant levels (Fig. 5a–c). In addition, inoculation with Fm and Si significantly increased the scavenging activity of ROS by total flavonoids under N conditions, as evidenced by the increase of 46.68 and 45.55% in the scavenging activity of •OH by total flavonoids, the increase of 56.02 and 52.15% in the scavenging activity of O2•− by total flavonoids, and the increase of 80.42 and 75.29% in the scavenging activity of DPPH by total flavonoids, respectively. Similarly, under W conditions, inoculation with Fm and Si led to a significant increase in the scavenging activity of •OH by total flavonoids by 42.26 and 25.57% respectively, the scavenging activity of O2•− by total flavonoids by 149.23 and 145.16% respectively, and the scavenging activity of DPPH by total flavonoids by 185.30 and 168.36% respectively.

Effects of Funneliformis mosseae and Serendipita indica on •OH (a), O2•− (b), and DPPH (c) scavenging activity of root total flavonoids under waterlogging stress and non-waterlogging stress. Data (means ± SD, n = 4) with different letters above the bars indicated the significant difference among treatments at 0.05 levels. See Fig. 1 for abbreviations
Changes in root antioxidant enzyme activities
Compared with the N treatment, the W treatment significantly reduced root SOD activities in uninoculated seedlings by 14.07%, accompanied by no significant change in Fm- and Si-inoculated seedlings (Fig. 6a). Compared with the uninoculated treatment, inoculation with Fm and Si significantly increased root SOD activities by 13.12 and 10.76% under N and by 35.35 and 29.44% under W, respectively.

Effects of Funneliformis mosseae and Serendipita indica on SOD (a), CAT (b), GPX (c), and POD (d) activities in peach roots under waterlogging stress and non-waterlogging stress. Data (means ± SD, n = 4) with different letters above the bars indicated the significant difference among treatments at 0.05 levels. See Fig. 1 for abbreviations
The W treatment resulted in a significant decrease in root CAT activities in uninoculated seedlings, whereas it significantly increased root CAT activities in Fm- and Si-inoculated seedlings by 77.32 and 41.01%, respectively (Fig. 6b). Under N conditions, inoculation with Fm significantly decreased root CAT activities by 47.93% compared with uninoculated treatment, with no significant change in Si-inoculated seedlings; under W conditions, inoculation of both Fm and Si significantly increased root CAT activities by 253.20 and 386.73%, respectively.
Root GPX activities underwent significant changes under W versus N conditions: a decrease in uninoculated and Si-inoculated seedlings and no change in Fm-inoculated seedlings (Fig. 6c). Under N conditions, inoculation with Fm significantly decreased root GPX activities by 39.52%, while inoculation with Si significantly increased root GPX activities by 26.95%, compared with the uninoculated treatment. Under W conditions, inoculation with both Fm and Si significantly increased root GPX activities by 73.01 and 99.20%, respectively, compared with the uninoculated treatment.
W stress did not affect root POD activities in both uninoculated and Fm-inoculated seedlings, but increased root POD activities in Si-inoculated seedlings by 48.75%, compared with the N treatment (Fig. 6d). Only Si inoculation significantly increased root POD activities under both N and W conditions by 2400.60 and 2233.32%, respectively, compared with the uninoculated treatment.
Changes in root antioxidant enzyme gene expression
Expression levels of eight PpSOD homologous genes were examined (Fig. 7a). Compared with the N treatment, the W treatment significantly decreased the expression of PpSOD2, PpSOD4, PpSOD5, PpSOD6, and PpSOD7 in roots of uninoculated seedlings by 97.14, 97.44, 97.17, 98.11, and 55.88%, respectively. However, in Fm-inoculated seedlings, PpSOD3 and PpSOD7 gene expression was up-regulated by 1690.00 and 266.67% under W versus N conditions, respectively, along with a 441.67% increase in PpSOD3 gene in Si-inoculated seedlings. Inoculation of Fm under N conditions resulted in a significant up-regulation of PpSOD8 expression by 314.56%, compared with no-fungi control, while all other PpSOD1–7 expression levels were significantly down-regulated by 55.88–92.38%. Similarly, inoculation with Si significantly down-regulated PpSOD1–8 expression by 49.06–81.90% under N conditions, compared with no-fungi control. On the other hand, under W conditions, compared with the uninoculated treatment, the expression of PpSOD3 and PpSOD7 was also significantly up-regulated by inoculation with Fm, by 341.98–266.67%, respectively, accompanied by no significant or down-regulated change in the other PpSODs. Inoculation with Si significantly induced an up-regulation of PpSOD3 gene expression under W treatment by 60.49%, compared with the uninoculated treatment, while also suppressed PpSOD1 expression by 91.57%, plus no significant effect on the other PpSODs.

Effects of Funneliformis mosseae and Serendipita indica on expression levels of PpSODs (a), PpCATs (b), PpGPXs (c), and PpPOD1 (d) genes in peach roots under waterlogging stress and non-waterlogging stress. Data (means ± SD, n = 4) with different letters above the bars indicated the significant difference among treatments at 0.05 levels. Here, relative expression of genes were normalized to N-F treatment (= 1). See Fig. 1 for abbreviations
The W treatment resulted in a significant 90.20% down-regulation of PpCAT1 expression in uninoculated peach roots, as well as a 990.91% up-regulation of PpCAT1 expression in Si-inoculated plants, compared with the N treatment (Fig. 7b). PpCAT2 expression was down-regulated in uninoculated and Si-inoculated seedlings respectively by 97.09 and 82.09% under W versus N conditions. Inoculation with Fm significantly reduced the expression of PpCAT1 and PpCAT2 under N conditions by 90.20 and 67.00%, respectively, plus no significant change under W conditions. Inoculation with Si also significantly decreased the expression of PpCAT1 and PpCAT2 by 89.22 and 34.95%, respectively, under N conditions, but up-regulated PpCAT1 expression by 1100% under W conditions.
The W treatment inhibited the expression of PpGPX6 and PpGPX8 in no-fungi-inoculated seedlings, but it did not significantly affect the Fm-inoculated seedlings, compared with the N treatment (Fig. 7c). In addition, PpGPX6 expression was down-regulated in Si-inoculated plants under W versus N conditions, while PpGPX8 expression was up-regulated. Meanwhile, Fm also significantly decreased the expression of PpGPX6 and PpGPX8 by 84.00% and 88.24% under N conditions, but no significant effect was observed under W conditions. Si treatment significantly promoted the expression of PpGPX6 by 202.00 and 582.35% respectively under N and W conditions, and also increased the expression of PpGPX8 by 1850.00% under W conditions, along with 89.22% reduction in PpGPX8 in Si-inoculated seedling under N conditions.
The W treatment significantly promoted the expression of PpPOD1 only in Si-inoculated seedlings (Fig. 7d). Similarly, in the two symbiotic fungi, only Si significantly up-regulated the expression of PpPOD1 by 8316.67 and 1203320.00% under N and W conditions, respectively.
Correlation analysis
Root fungal colonization rate showed a significantly negative correlation with O2•−, H2O2, PpSOD1, PpSOD4, PpSOD5, PpSOD6, and PpGPX8, while exhibiting a positive correlation with total flavonoids and SOD activities (Fig. 8). Furthermore, O2•− was significantly positively correlated with PpSOD1, while negatively correlated with SOD activities and PpSOD8. H2O2 showed a significantly negative correlation with SOD, CAT, GPX, PpCAT2, and PpGPX6, along with a significantly positive correlation with PpSOD1.

The correlation between selected parameters presented with a heat map. Here, * and ** mean significant correlation at 0.05 and 0.01 levels, respectively
PCA studies
The PCA of the treatments as well as the antioxidant variables showed that PC1 (53.7%) distinguished inoculated and non-inoculated treatments and PC2 (20.6%) separated the N and W treatments (Fig. 9). Moreover, in PC2, total flavonoids, root fungal colonization rate, and ROS scavenging activity of total flavonoids were clustered together with ROS levels, showing a close relationship.

Changes in PCA of treatments and physiological variables
Discussion
In this study, a 2-week W treatment did not have a significant effect on the root colonization rate of the AM fungus Fm, indicating a potential waterlogging tolerance in this Fm strain. Several AMF species have been observed to survive under waterlogged soil conditions [42]. Similarly, a short-term (e.g., 11 and 31 days) W treatment had little effect on the root colonization rate of mycorrhizal mixture (Gigaspora margarita and G. rosae) in beans [43]. In contrast, a 10 day W treatment suppressed the root colonization of Fm in trifoliate orange [44], suggesting that AMF colonization under W conditions is dependent on host species. Interestingly, the W treatment significantly increased the colonization rate of the endophytic fungus Si in peach roots, which may be attributable to the strong adaptability of Si to adverse environments [45] and its capacity to colonize primarily apoptotic root cells [46, 47]. In addition, this fungus Si is able to produce IAA [48], which in turn offloads to the host roots, thereby promoting root growth, as shown in this study. Higher root IAA levels in Si-inoculated seedlings under W versus N conditions were consistent with increased root colonization of Si. The increase in IAA levels in host plants by Si was reported to tolerate abiotic stress [49, 50]. This provides Si with more roots for their colonization under W, thus causing an increase in root colonization of Si, whereas it remains to be confirmed by further studies.
Root development determines the ability of roots to absorb soil water and nutrients [51]. In this study, a 2 week W treatment did not significantly inhibit peach root architecture, likely due to the short duration of the W treatment. However, inoculation with Fm or Si improved root development to varying degrees, particularly under W conditions, where both fungi considerably promoted root development, along with Fm exhibiting more pronounced effects than Si. Similar results were observed in citrus seedlings inoculated with an arbuscular mycorrhizal fungus, Diversispora spurca, after a 37 day W treatment [21]. The improvement in root development is closely related to the promotion of root auxins and cytokinins by these root-associated symbiotic fungi [48, 52]. These large roots aid in the colonization of plants by symbiotic fungi, enabling them to withstand W stress. More attention needs to determine whether endophytic fungal colonization affects changes in root anatomical structure under W stress.
Prolonged oxygen deficiency, such as soil waterlogging, can cause a burst of ROS in plants, thus resulting in the peroxidation of membrane lipids and oxidative damage to DNA and proteins [53]. In this study, the 2 week W treatment dramatically triggered an increase in root O2•− and H2O2 levels in both inoculated and uninoculated peach, except no change in root O2•− levels in Fm-inoculated seedlings. The magnitude of this increase was higher in uninoculated seedlings than in fungi-inoculated seedlings, implying that fungi-inoculated peach seedlings were less affected by W than uninoculated seedlings. On the other hand, Fm and Si significantly inhibited root O2•− and H2O2 levels under both N and W conditions. Correlation analysis demonstrated a significantly negative correlation between root fungal colonization rate and root O2•− and H2O2 levels. Similar results were presented in trifoliate orange seedlings inoculated with Si under drought stress conditions [47] and lavender plants inoculated with Fm under W conditions [54]. These results indicate that Fm and Si have significantly inhibitory effects on ROS burst in waterlogged peach.
Flavonoids are a typical class of phenolic compounds that are seen as an antioxidant due to their capacity to chelate metal ions and scavenge ROS [11]. In this study, the W treatment led to significant increases in root total flavonoid concentrations of inoculated seedlings, but not uninoculated seedlings, indicating that W treatment increased the production of flavonoids in roots of fungi-colonized plants. In addition, inoculation with Fm and Si collectively raised root total flavonoid concentrations, with the favorable effect being stronger under W versus N conditions. Total flavonoid concentrations were also significantly and positively correlated with root fungal colonization rate. The results are in agreement with the findings of Liu et al. [13] in trifoliate orange inoculated with Fm under drought stress conditions, as well as the findings of Cheng et al. [55] in Dimocarpus longan seedlings inoculated with Si. Nevertheless, in geranium, Fm triggered a significant increase in total flavonoids under well-watered conditions, but not under moderate and severe drought stress [56]. In Aloe vera plants, Si led to an increase in total flavonoids in leaves, but not roots, one and two months after 200 and 300 mM NaCl application [57]. These findings imply that the effect of root-associated symbiotic fungi on host flavonoids depends on plant organs and the stressful environment. The increase in total flavonoids under root-associated endophytic fungal inoculation conditions may be derived from the activation of associated enzymes and gene expression in flavonoid synthesis pathways [13, 58, 59], while the underlying mechanisms remain unknown and require further investigation.
The present results showed that the W treatment only elicited an increase in the •OH scavenging by root total flavonoids in uninoculated seedlings, with no significant effect on the O2•− and DPPH scavenging. The W treatment, however, led to an increase in the scavenging of •OH, O2•−, and DPPH in roots of fungi-inoculated seedlings, suggesting that symbiotic fungi-colonized plants exhibited a greater ROS scavenging capacity via total flavonoids than uncolonized plants. Similar findings were also observed in trifoliate orange seedlings inoculated with Fm under drought stress conditions, where Fm-inoculated trifoliate orange seedlings exhibited stronger •OH and O2•− scavenging activity through root total flavonoids than the uninoculated control [13]. Furthermore, both Fm and Si inoculation significantly increased the scavenging of •OH, O2•−, and DPPH by root total flavonoids, irrespective of waterlogging or not, which is consistent with the results of Liu et al. [13] under Fm inoculation conditions. This suggests that root-associated symbiotic fungi like Fm and Si boost ROS scavenging activities of total flavonoids, thus allowing the fungus-colonized plants to suffer less oxidative damage under stressful conditions.
In response to excessive ROS accumulation caused by adverse stresses, plants also possess antioxidant enzymes to maintain ROS balance [60]. Peach plants can regulate antioxidant enzyme activities at the beginning of W stress, but antioxidant enzyme activities decrease sharply with increasing duration of W stress, suggesting a limited ability of peach plants to cope with prolonged W [61]. In this study, the W treatment almost inhibited root antioxidant enzyme activities in uninoculated seedlings, which is in agreement with the findings of Zhang et al. [61]. Nevertheless, increased CAT and POD activities in Si-inoculated plants under W versus N conditions as well as elevated CAT activities in Fm-inoculated plants, implied that fungi-inoculated plants maintained a higher activity of a specific antioxidant enzyme under prolonged W, thus presenting a higher W tolerance than uninoculated plants. Inoculation with Fm significantly increased only root SOD activities under N conditions, whereas it increased root SOD, CAT, and GPX activities under W conditions, suggesting that the regulation of antioxidant enzyme activities by Fm is influenced by the W environment. On the other hand, Si inoculation resulted in a significant increase in root SOD, GPX, and POD activities under both N and W conditions, with a significant increase in root CAT activities observed under W conditions only. This suggests that Si has a stronger ability to stimulate antioxidant enzyme activities under W conditions than Fm.
In addition, a 2 week W treatment did not affect or inhibited the expression of PpSODs, PpPODs, PpCATs, and PpGPXs genes in the roots of uninoculated plants. Xu et al. [2] also found inhibitive activities of CAT and POD in peach after 3 days of W conditions. However, PpSOD3 and PpSOD7 expression was up-regulated in Fm-inoculated seedlings under W versus N conditions, and PpSOD3, PpCAT1, PpGPX8, and PpPOD1 expression was up-regulated in Si-inoculated seedlings, with little change or inhibition of other antioxidant enzyme gene expression. This suggests that plants inoculated with symbiotic fungi presented a greater antioxidant enzyme defense system than uninoculated plants. On the other hand, Fm inoculation increased the expression of PpSOD3 and PpSOD7 under W conditions, indicating that the expression of PpSOD3 and PpSOD7 was specific to both Fm inoculation and W treatment. As SODs primarily catalyze O2•− to H2O2, Fm-inoculated plants exhibited lower O2•− levels under W conditions, but not H2O2. However, in trifoliate orange seedlings, Fm inoculation could up-regulate the expression of PtCAT1, PtPOD, and PtMn-SOD under drought stress [62, 63]. These findings demonstrate that the regulation of plant antioxidant enzyme genes by Fm is variable and dependent on environmental conditions and host species. However, the endophytic fungus Si up-regulated the expression of PpSOD3, PpCAT1, PpGPX6, PpGPX8, and PpPOD1 under W conditions, implying a stronger ability of Si to activate antioxidant enzyme gene expression than Fm in response to W stress. Fm inoculation did not have any impact on PpCAT1, PpCAT2, PpGPX6, PpGPX8, and PpPOD1 expression under W conditions, while it increased CAT, GPX, and POD activities under W conditions. Similar results have been reported in symbiotic fungi-inoculated trifoliate orange seedlings under drought [47]. Nevertheless, changes in the expression of antioxidant enzyme genes did not correspond completely to changes in antioxidant enzyme activity under Fm inoculation conditions. This inconsistency is due to the fact that gene expression is followed by transcription and translation, as well as phosphorylation and ubiquitination. GPX, CAT, and POD are involved in the decomposition of H2O2, with GPX additionally catalyzing the decomposition of phospholipid hydroperoxides [64]. Thus, Si-inoculated plants maintained lower O2•− and H2O2 levels in response to W treatment. As a result, it is concluded that the antioxidant enzyme defense mechanisms of Fm and Si in peach against waterlogging are variable. Further evidence, including transcriptome analysis, is required to validate these findings.
Conclusion
Overall, inoculation with the AM fungus Fm and the endophytic fungus Si improved the plant growth and root development and alleviated ROS burst in peach under waterlogging conditions to varying degrees. The reduction in ROS levels caused by Fm inoculation under W was associated with an increase in total flavonoids, three antioxidant enzymes, and the up-regulated expression of two PpSODs in roots. The reduction in ROS levels by Si was linked to an increase in total flavonoids, four antioxidant enzymes, and up-regulated expression of five antioxidant enzyme genes in roots. Based on these findings, an involved mechanism was illustrated in Fig. 10. However, it remains unclear how the two fungi initiate the expression of respective antioxidant enzyme genes and promote total flavonoid synthesis, which has yet to be explored. In addition, this study only analyzed changes in roots, and it is unclear whether the results of leaves are the same as those of roots. Transcriptome sequencing can potentially reveal the underlying molecular mechanism behind the enhanced W tolerance of peach plants by antioxidant defense systems under Fm and Si inoculation conditions. This study also highlights the potential of root-associated symbiotic fungi, particularly the culturable endophytic fungus Si, as a pathway to enhance peach W tolerance.

A mechanism regarding enhanced waterlogging tolerance in peaches caused by Fm and Si inoculation. Here, ↑ means an increase in the variables; ↓ means a decrease in the variables. CAT catalase, DPPH 2,2-diphenyl-1-picrylhydrazyl radical, Fm Funneliformis mosseae, GPX glutathione peroxidase, GSSG oxidized glutathione, GSH glutathione, H2O2 hydrogen peroxide, O2•− superoxide anion radicals, POD peroxidase, Si Serendipita indica, SOD superoxide dismutase
Availability of data and materials
Not application.
References
Bento C, Gonçalves AC, Silva B, Silva LR. Peach (Prunus Persica): phytochemicals and health benefits. Food Rev Int. 2020;38:1703–34.
Xu FJ, Cai HQ, Zhang XA, Su MS, Zhou HJ, Li XW, Zhang MH, Hu Y, Gu C, Du JH, Ye ZW. Comparison of waterlogging tolerance of three peach rootstock seedlings based on physiological, anatomical and ultra-structural changes. Horticulturae. 2022;8:720.
Xiao YS, Wu XL, Sun MX, Peng FT. Hydrogen sulfide alleviates waterlogging-induced damage in peach seedlings via enhancing antioxidative system and inhibiting ethylene synthesis. Front Plant Sci. 2020;11:696.
Kreuzwieser J, Rennenberg H. Molecular and physiological responses of trees to waterlogging stress. Plant Cell Environ. 2014;37(10):2245–59.
Stasnik P, Grobkinsky DK, Jonak C. Physiological and phenotypic characterization of diverse Camelina sativa lines in response to waterlogging. Plant Physiol Biochem. 2022;183:120–7.
Hasanuzzaman M, Mahmud JA, Nahar K, Anee TI, Inafuku M, Oku H, Fujita M. Responses, adaptation, and ROS metabolism in plants exposed to waterlogging stress. In: Khan MIR, Khan NA, editors. Reactive oxygen species and antioxidant systems in plants: role and regulation under abiotic stress. Singapore: Springer; 2017.
McGee T, Shahid MA, Beckman TG, Chaparro JX, Schaffer B, Sarkhosh A. Physiological and biochemical characterization of six Prunus rootstocks in response to flooding. Environ Exp Bot. 2021;183:104368.
Inzé D, Montagu MV. Oxidative stress in plants. Curr Opin Biotechnol. 1995;6(2):153–8.
Liu Y, Qing YL, Jing LC, Zou WT, Guo R. Platinum-copper bimetallic colloid nanoparticle cluster nanozymes with multiple enzyme-like activities for scavenging reactive oxygen species. Langmuir. 2021;37(24):7364–72.
Agati G, Azzarello E, Pollastri S, Tattini M. Flavonoids as antioxidants in plants: location and functional significance. Plant Sci. 2012;196:67–76.
Shen N, Wang TF, Gan Q, Liu S, Wang L, Jin B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022;383:132531.
Su SG, Wei YL, Huang YJ, Huang YQ, Luo GX. Study on antioxidant and free radical scavenging effects of total flavonoids in Rosa laevigata. J Guangxi Univ Chin Med. 2015;18(1):47–8.
Liu XQ, Cheng S, Aroca R, Zou YN, Wu QS. Arbuscular mycorrhizal fungi induce flavonoid synthesis for mitigating oxidative damage of trifoliate orange under water stress. Environ Exp Bot. 2022;204:105089.
Bidartondo MI, Read DJ, Trappe JM, Merckx V, Ligrone R, Duckett JG. The dawn of symbiosis between plants and fungi. Biol Lett. 2011;7(4):574–7.
Bonfante P, Genre A. Mechanisms underlying beneficial plant–fungus interactions in mycorrhizal symbiosis. Nat Commun. 2010;1(1):48.
Wang Y, Zou YN, Shu B, Wu QS. Deciphering molecular mechanisms regarding enhanced drought tolerance in plants by arbuscular mycorrhizal fungi. Sci Hortic. 2023;308:111591.
Dhalaria R, Kumar D, Kumar H, Nepovimova E, Kuča K, Islam MT, Verma R. Arbuscular mycorrhizal fungi as potential agents in ameliorating heavy metal stress in plants. Agronomy. 2020;10(6):815.
Cheng S, Zou YN, Kuča K, Hashem A, Abd Allah EF, Wu QS. Elucidating the mechanisms underlying enhanced drought tolerance in plants mediated by arbuscular mycorrhizal fungi. Front Microbiol. 2021;12:809473.
Fusconi A, Mucciarelli M. How important is arbuscular mycorrhizal colonization in wetland and aquatic habitats? Environ Exp Bot. 2018;155:128–41.
Ban YH, Tan JY, Xiong Y, Mo XT, Jiang YH, Xu ZY. Transcriptome analysis reveals the molecular mechanisms of Phragmites australis tolerance to CuO-nanoparticles and/or flood stress induced by arbuscular mycorrhizal fungi. J Hazard Mater. 2022;442:130118.
Wu QS, Zou YN, Huang YM. The arbuscular mycorrhizal fungus Diversispora spurca ameliorates effects of waterlogging on growth, root system architecture and antioxidant enzyme activities of citrus seedlings. Fungal Ecol. 2013;6:37–43.
Stancheva I, Nedyalkova K, Geneva M, Donkova R, Hristozkova M, Perfanova I, Sichanova M, Petkova G, Djonova E, Valchovski H. Nutritional value of cowpea (Vigna Unguiculata L. Walp) grain grown under different soil moisture as affected to the dual inoculation with nitrogen fixing bacteria and arbuscular mycorrhizal fungi. Bulg J Soil Sci. 2016;1(2):112–21.
Weiß M, Waller F, Zuccaro A, Selosse MA. Sebacinales–one thousand and one interactions with land plants. New Phytol. 2016;211:20–40.
Barrio-Duque A, Ley J, Samad A, Antonielli L, Sessitsch A, Compant S. Beneficial endophytic bacteria-Serendipita indica interaction for crop enhancement and resistance to phytopathogens. Front Microbial. 2019;10:2888.
Liu BH, Jing DW, Liu FC, Ma HL, Liu XH, Peng L. Serendipita indica alleviates drought stress responses in walnut (Juglans regia L.) seedlings by stimulating osmotic adjustment and antioxidant defense system. Appl Microbiol Biot. 2021;105:8951–68.
Yang Y, Zhang J, Zhang Y, Zhu J, Dong S. Effects of Piriformospora indica on Chinese cabbage production and resistance to waterlogged stress. J Chem Pharm Res. 2013;5:520–4.
Hosseini F, Mosaddeghi MR, Dexter AR. Effect of the fungus Piriformospora indica on physiological characteristics and root morphology of wheat under combined drought and mechanical stresses. Plant Physiol Biochem. 2017;118:107–20.
Hosseini F, Mosaddeghi MR, Dexter AR, Sepehri M. Maize water status and physiological traits as affected by root endophytic fungus Piriformospora indica under combined drought and mechanical stresses. Planta. 2018;247:1229–45.
Zhang MQ, Wang YS. Seven species of VA mycorrhizal fungi from northern China. Acta Mycol Sin. 1191;10(1):13–21.
Yang L, Cao JL, Zou YN, Wu QS, Kuča K. Piriformospora indica: a root endophytic fungus and its roles in plants. Not Bot Horti Agrobota. 2020;48:1–13.
Yang L, Zou YN, Tian ZH, Wu QS, Kuča K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci Hortic. 2021;277:109815.
Liang SM, Zou YN, Shu B, Wu QS. Arbuscular mycorrhizal fungi and endophytic fungi differentially modulate polyamines or proline of peach in response to soil flooding. Pedosphere. 2023. https://doi.org/10.1016/j.pedsph.2023.05.002.
Phillips JM, Hayman DS. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc. 1970;55:158–61.
Wang AG, Luo GH. Quantitative relation between the reaction of hydroxylamine and superoxide anion radicals in plants. Plant Physiol Commun. 1990;6:55–7.
Velikova V, Yordanov I, Edreva A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines. Plant Sci. 2000;151:59–66.
Wu QS. Experimental supervision of plant physiology. Beijing: China Agriculture Press; 2018.
Drotar A, Phelps P, Fall R. Evidence for glutathione peroxidase activities in cultured plant cells. Plant Sci. 1985;42:35–40.
Liu J, Macarisin D, Wisniewski M, Sui Y, Droby S, Norelli J, Hershkovitz V. Production of hydrogen peroxide and expression of ROS-generating genes in peach flower petals in response to host and non-host fungal pathogens. Plant Pathol. 2013;62(4):820–8.
Huan C, Jiang L, An XJ, Yu ML, Xu Y, Ma RJ, Yu ZF. Potential role of reactive oxygen species and antioxidant genes in the regulation of peach fruit development and ripening. Plant Physiol Bioch. 2016;104:294–303.
Tong Z, Gao Z, Wang F, Zhou J, Zhang Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol Biol. 2019;10:71.
Livak KJB, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and 2−ΔΔCt method. Methods. 2001;25:402–8.
Huang GM, Srivastava AK, Zou YN, Wu QS, Kuča K. Exploring arbuscular mycorrhizal symbiosis in wetland plants with a focus on human impacts. Symbiosis. 2021;84:311–20.
Sah S, Reed S, Jayachandran K, Dunn C, Fisher JB. The effect of repeated short-term flooding on mycorrhizal survival in snap bean roots. HortScience. 2006;41:598–602.
Cheng XF, Wu HH, Zou YN, Wu QS, Kuča K. Mycorrhizal response strategies of trifoliate orange under well-watered, salt stress, and waterlogging stress by regulating leaf aquaporin expression. Plant Physiol Biochem. 2021;162:27–35.
Pham GH, Kumari R, Singh A, Malla R, Prasad R, Sachdev M, Kaldorf M, Buscot F, Oelmüller R, Hampp R, Saxena AK, Rexer KH, Kost G, Varma A. Axenic culture of symbiotic fungus Piriformospora indica. In: Varma A, Abbott L, Werner D, Hampp R, editors. Plant surface microbiology. New York: Springer-Verlag; 2008. p. 593–613.
Jungk AO. Dynamics of nutrient movement at the soil-root interface. In: Jungk AO, editor. Plant roots: the hidden half. Boca Raton: CRC Press; 2002. p. 919–1016.
Cao JL, He WX, Zou YN, Wu QS. An endophytic fungus, Piriformospora indica, enhances drought tolerance of trifoliate orange by modulating the antioxidant defense system and composition of fatty acids. Tree Physiol. 2023;43(3):452–66.
Sirrenberg A, Göbel C, Grond S, Czempinski N, Ratzinger A, Karlovsky P, Santos P, Feussner I, Pawlowski K. Piriformospora indica affects plant growth by auxin production. Physiol Plant. 2007;131:581–9.
Boorboori MR, Zhang HY. The role of Serendipita indica (Piriformospora indica) in improving plant resistance to drought and salinity stresses. Biology. 2022;11:952.
Rong ZY, Lei AQ, Wu QS, Srivastava AK, Hashem A, Abd Allah EF, Kuča K, Yang T. Serendipita indica promotes P acquisition and growth in tea seedlings under P deficit conditions by increasing cytokinins and indoleacetic acid and phosphate transporter gene expression. Front Plant Sci. 2023;14:1146182.
Singh Gahoonia T, Nielsen NE. Root traits as tools for creating phosphorus efficient crop varieties. Plant Soil. 2004;260:47–57.
Liu RC, Yang L, Zou YN, Wu QS. Root-associated endophytic fungi modulate endogenous auxin and cytokinin levels to improve plant biomass and root morphology of trifoliate orange. Hortic Plant J. 2023;9(3):463–72.
Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Bioch. 2010;48(12):909–30.
Rasouli F, Hassanpouraghdam MB, Pirsarandib Y, Aazami MA, Asadi M, Ercisli S, Mehrabani LV, Puglisi I, Baglieri A. Improvements in the biochemical responses and Pb and Ni phytoremediation of lavender (Lavandula angustifolia L.) plants through Funneliformis mosseae inoculation. BMC Plant Biol. 2023;23:252.
Cheng CZ, Li D, Wang B, Liao B, Qu PY, Liu W, Zhang YY, Lü PT. Piriformospora indica colonization promotes the root growth of Dimocarpus longan seedlings. Sci Hortic. 2022;301:111137.
Amiri R, Nikbakht A, Rahimmalek M, Hosseini H. Variation in the essential oil composition, antioxidant capacity, and physiological characteristics of Pelargonium graveolens L. inoculated with two species of mycorrhizal fungi under water deficit conditions. J Plant Growth Regul. 2017;36:502–15.
Sharma P, Kharkwal AC, Abdin MZ, Varma A. Piriformospora indica-mediated salinity tolerance in Aloe vera plantlets. Symbiosis. 2017;72:103–15.
Khalid M, Hassani D, Bilal M, Liao JL, Huang DF. Elevation of secondary metabolites synthesis in Brassica campestris ssp. chinensis L. via exogenous inoculation of Piriformospora indica with appropriate fertilizer. PLoS ONE. 2017;12(5):e0177185.
Aseel DG, Rashad YM, Hammad SM. Arbuscular mycorrhizal fungi trigger transcriptional expression of flavonoid and chlorogenic acid biosynthetic pathways genes in tomato against tomato mosaic virus. Sci Rep. 2019;9:9692.
Dumanović J, Nepovimova E, Natić M, Kuča K, Jaćević V. The significance of reactive oxygen species and antioxidant defense system in plants: a concise overview. Front Plant Sci. 2021;11:552969.
Zhang BB, Sun MX, Liu WX, Lian M, Yang SK, Peng FT, Xiao YS. Waterlogging resistance and evaluation of physiological mechanism of three peach (Prunus persica) rootstocks. Protoplasma. 2023;260:1375–88.
He JD, Zou YN, Wu QS, Kuča K. Mycorrhizas enhance drought tolerance of trifoliate orange by enhancing activities and gene expression of antioxidant enzymes. Sci Hortic. 2020;262:108745.
Ding YE, Zou YN, Wu QS, Kuča K. Mycorrhizal fungi regulate daily rhythm of circadian clock in trifoliate orange under drought stress. Tree Physiol. 2022;42:616–28.
Ma XY, Deng D, Chen WD. Inhibitors and activators of SOD, GSH-Px, and CAT. Enzym Inhib Act. 2017;29:207–24.
Funding
This study was supported by the Open Fund of Engineering Research Center of Ecology and Agricultural Use of Wetland, Ministry of Education (KFT202005). The authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2023R356), King Saud University, Riyadh, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
SML designed the experiment. SML prepared the materials for the experiment. SML analyzed the data. SML wrote the manuscript. QSW, AH, and EFA revised the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no potential competing interests regarding the publication of this work.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
: Table S1. Primer sequences of genes used in peach for qRT-PCR. Fig S1. The growth performance of potted peach seedlings inoculated with Funneliformis mosseae (Fm), Serendipita indica (Si), and no-fungi (-F) under waterlogging (W) and non-waterlogging (N) conditions.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liang, SM., Hashem, A., Abd-Allah, E.F. et al. Root-associated symbiotic fungi enhance waterlogging tolerance of peach seedlings by increasing flavonoids and activities and gene expression of antioxidant enzymes. Chem. Biol. Technol. Agric. 10, 124 (2023). https://doi.org/10.1186/s40538-023-00500-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-023-00500-w