
Abstract
Background
Previous studies have suggested that the targeted application of biochars in agricultural soils may benefit soil health and crop production. Physico-chemical properties of soils after biochar addition have been explored, but less is known about how microbial parameters respond. Therefore, impact of biochar (NB), mineral fertilizer-activated biochar (AB), or mineral fertilizer (MF) application on selected chemical and microbial parameters of lettuce-planted soil was evaluated in a pot experiment.
Results
In comparison to the control, soil enzymes activities, related to carbon (C), nitrogen (N), and phosphorus (P) cycling, and their content in plant biomass, were significantly increased by the addition of mineral fertilizer with or without biochar (MF, NB + MF). Conversely, microbial respiration (basal and substrate induced) was highly responsive to the activated biochar amendment (AB) as compared to other treatments. N, P, and potassium (K) concentrations in soil pore water were stabilized by the mineral fertilizer-activated biochar, indicating reduced leaching and the likelihood of increased longevity of these nutrients in soils. Enhanced carbon acquisition and mitigated nitrogen acquisition in soil of the most experimental treatments were coupled with higher crop (lettuce) biomass.
Conclusions
Our study demonstrates that the application of biochar both with and without mineral fertilizer has the potential to enhance microbial activity and fertility in the tested agricultural soil, but that leaching of fertilizer-borne nutrients may be mitigated by the activation process.
Graphical Abstract

Similar content being viewed by others


Explore related subjects
Find the latest articles, discoveries, and news in related topics.Background
Intensive agricultural practices to meet the rising population demand currently rely on the injudicious use of mineral fertilizers. This has been driven by a need to ensure food security even at the expense of soil quality [1, 2]. While the application of chemical fertilizers boosts crop production, the continuous use of these amendments poses serious threats to soil quality and microbial processes which may result in a depletion in soil quality over time. Strategies are required to simultaneously maintain or increase crop production and conserve soil health. One alternative to chemical fertilizers is the use of organic amendments such as farmyard manure, compost, digestate and biochar. When applied to soil as amendments, they enhance the soil physical, chemical and biological properties (microbial activities) resulting in increased nutrient cycling in support of crop production [3,4,5,6].
Amongst these organic soil amendments, biochar is a carbon-rich solid material produced by thermo-chemical decomposition of various feedstocks under low oxygen environment. It has been established that the application of biochar can improve soil physical conditions [7, 8] soil chemical properties such pH and cation exchange capacity [9,10,11], soil microbial activity [12, 13], soil nutrients availability and retention [14,15,16]. An increase in certain crops yield and production has also been reported under biochar application [17,18,19]. Furthermore, the combined application of biochar with other amendments such as chemical fertilizers, manures and humic substances has also been proven to enhance soil fertility, microbial soil activity and crop [20,21,22]. For instance, a study reported to increase soil nutrient contents and C and N cycling enzymes in response to combined application of biochar and mineral fertilizers [13]. Another study described enhanced soil basal respiration and C, N and P cycling enzyme activities under the combined application of biochar, manure and humic substances [4]. While biochar has the potential to improve soil health and crop yields, it is often applied with mineral fertilizers due to a low content of readily available nutrients present in biochar. However, the impact of biochar may vary depending on the soil properties, biochar rate, and the amount of fertilizer applied [23,24,25].
Soil microorganisms and their activities are essential indicators of soil quality as they regulate organic matter transformation and nutrient cycles [26]. Exogeneous addition of organic materials results in a shift in microbial diversity and functioning which eventually alters microbial activity in the soil [24, 27]. Biochar application has been shown to affect microbial population both directly and indirectly through altering soil physico-chemical properties such as pH, cation exchange capacity and by increasing the nutrients availability [28, 29]. In addition, biochar application improves soil physical properties which is also linked with the enhanced microbial population and diversity [30]. A study proposed that biochar addition enhanced microbial biomass by providing a niche for microbial growth along with increased nutrients supply [31]. Another study described a positive relationship between pH and soil microbial activity with applied biochar [32]. In general, enhanced microbial population and activities are associated with increased enzymatic activity in the soil. Biochar application additionally enhance these microbial enzymes which are the key drivers regulating nutrient cycles [33]. However, the negative and insignificant impacts of biochar on microbial activity and abundance were found as well [34]. The inconsistent impact of biochar on microbial activity remains a key point to be addressed by an aimed research.
Currently, the effects of biochar in conjunction with various soil amendments remain inconclusive; its efficacy has been found to range from positive in low fertility, acidic soils to neutral or negative in highly fertile soils [23, 30]. Thus, the general effects of biochar on soil health and quality are two-sided [35]. Therefore, the consumers (farmers) increase a demand for manufactured biochar with favourable effects in a variety of soils. Biochars produced through various activation processes may promise to meet these expectations to maximize potential benefits of biochar for enhancing soil health and crop production. Activation provides enhanced surface properties such as pore area and surface area of the resultant activated biochar (AB). A wealth of literature is available on the role of activated biochar in improving soil health, however little is known about the effects of biochar pre-treated with mineral fertilizer on stimulation of microbial activity and soil nutrient status. In addition, how microbial respiration (basal as well as substrate induced) and enzyme activities responds to biochar additions is uncertain.
The objectives of this study were to: (1) measure soil respiration and enzyme activity in agricultural soil amended with biochar, mineral fertilizer, mineral fertilizer + biochar, and mineral fertilizer-activated biochar; (2) determine plant biomass in response to the respective soil amendments; and (3) evaluate the impact of various biochar treatments on the improvement of soil health and crop productivity.
Materials and methods
Preparation of biochars
The biochar obtained for this study was a commercial product prepared by pyrolysis of agricultural waste (cereal husks, sunflower peels, and fruit processing waste) at 650 °C (Sonnenerde GmbH, Austria). According to the manufacturer specification, the properties of the biochar were as follows: 41% dry matter (DM), 866 g·kg−1 carbon (C), 3.0 g·kg−1 N, 11.7% Ash550 °C, 0.42% salts, 0.38 H/C, 2.65 mS cm−1 electric conductivity, 8.5 pH(CaCl2), 288.5 m2 g−1 Brunauer–Emmett–Teller (BET).
An activated biochar was produced by 7-day incubation in mineral fertilizer solution. This activation solution (33 L) consisted of 208.3 g of phosphate fertilizer FOSMAG (with a content 5.46% phosphorus, 25.7% calcium, 1.2% magnesium, 7% sulphur), 72.5 g of urea (46% N), 20.8 g of potassium chloride (48.8% potassium), and 250.0 g of beet molasses (42% sugar). FOSMAG comprised citrate-soluble P (40%), water-soluble P (20%) and storage P in micromilled phosphate (40%). The final concentration of nutrient elements (and microbial energy source) were (in w/v) 345 mg L−1 P, 1.62 g L−1 Ca, 75.7 mg L−1 Mg, 442 mg L−1 S, 1 g·L−1 N, 308 g·L−1 K, 3.2 g·L−1 sugar. The activation solution was added to 5 kg of biochar and the whole suspension was aerated for the entire duration of activation. After 7 days, the biochar was separated from water solution by filtration on a 42-µm-mesh sieve, and added to the soil substrate as described in the following section.
Pot experimental set-up
The 8-week pot experiment with the lettuce (Lactuca sativa L.) was carried out to evaluate the effect of biochar, mineral fertilizers and their combinations on soil properties and plant biomass yield. The growth substrate was prepared by mixing a silty clay loam (USDA Textural Triangle) Haplic Luvisol (WRB soil classification) collected from a farm near the village of Troubsko (Czech Rep., 49°10′28″N 16°29′32″E) with a fine quartz sand (0.1–1.0 mm; ≥ 95% SiO2) in a weight ratio of 1:1 to ensure uniform drainage and prevent anaerobic conditions from developing. The Haplic Luvisol soil properties were as follows: 14.0 g·kg−1 total C, 1.60 g·kg−1 total N, 0.10 g·kg−1 total P, 0.15 g·kg−1 total S, 3.26 g·kg−1 total Ca, 0.24 g·kg−1 total Mg, 0.23 g·kg−1 total K; 7.3 pH (CaCl2).
One kilogram was thoroughly mixed with the amendments displayed in Table 1 and placed into a pot of volume 1 L. All experimental treatments were prepared in 4 replications. Experimental pots were placed randomly into a growth chamber Climacell EVO (BMT, Czech Rep.) where controlled conditions (full-spectrum LED lighting, intensity 20 000 lx; photoperiod 12 h; temperature 18/22 °C night/day; relative air humidity 70%) were maintained. Lettuce seeds were sprouted on wet filter paper for 2 days and then five of them were sown to the depth of approximately 2 mm in each pot. After sowing, soil moisture was maintained at approx. 60% of water holding capacity. The 10-day-old seedlings were reduced to one (the most robust plant) per pot.
After 8 weeks, lettuce seedlings were harvested, washed with water, tapped dry and cut apart at hypocotyl to upper part = aboveground biomass (AGB) and lower part = root. Both, AGB and roots, were weighed on a laboratory scale to determine fresh AGB and root biomass and then dried at 60 °C to the constant weight in laboratory drier to determine dry AGB and root biomass by weighting on laboratory scales.
Soil chemical and microbial analysis
Following harvest of the plants, soil samples were collected from three of the four replicate pots, avoiding any root material, for the determination of soil chemical properties. A 50:50 mixture of soil and water was added to 50-mL Falcon tubes. One rhizon pore-water sampler (Eijkelkamp Agrisearch equipment, NED) was placed into each tube. Pore-water was extracted according to [36] by removable syringes after a 24-h incubation period under ambient temperature conditions (⁓18 °C). To ensure sufficient volume for analysis, two extractions were performed resulting essentially in six replicate samples were treatment. Pore waters were tested for pH and electrical conductivity (EC) using a Multimeter 3420 (WTW Germany). The amount of total nitrogen in the pore water was detected on Vario MACRO GHN elemental analysis unit (GHNS Elementar Total N in Analytical Systems GmbH, Germany). Dissolved organic carbon (DOC) was determined using a total organic carbon analyser TOC-L a (CPH/CPN, Shimadzu), TNM-L segment flow analyzer (Shimadzu). The macronutrients P, K, Ca, Mg and S were measured using ICP-OES (720 ES, Varian Inc., CA, USA).
Dehydrogenase activity (DHA) was determined in 4 °C-stored soil samples by standard method based on triphenyltetrazolium chloride (TTC, values were expresses as µg TPF·g−1·h−1) [37]. The 4 °C-stored soil samples were used to measure a basal (BR) and substrate-induced soil respiration [38]—Glc-IR (D-glucose), Tre-IR (D-trehalose), NAG-IR (N-acetyl-β-D-glucosamine), Man-IR (D-mannose), Ala-IR (L-alanine) using MicroResp® device (The James Hutton Institute, Scotland) and colorimetric indication of CO2 emission.
Enzyme activities—arylsulfatase (ARS), N-acetyl-β-D-glucosaminidase (NAG), urease (Ure), β-glucosidase (GLU), phosphatase (Phos) were determined according to [39]. Nitrophenyl-derivates of natural substrates were used for measurement of GLU, PHOS, ARS, NAG (at emission wavelength 405 nm) and urea was a substrate for URE (measured at wavelength 650 nm), the values were expressed in nmol NH3·g−1·min−1 (urease) and in nmol (p-nitrophenol) PNP·g−1·min−1.
Using enzyme activities, nutrient acquisition ratios were calculated, based on the formulae presented in the study [40]:
Carbon (nitrogen) acquisition ratio directly depends on the secretion rate of carbon-utilizing (nitrogen-utilizing) enzymes. Additionally, microbial resource limitation was estimated through computation of vector length and angle in accordance with the theory of enzymatic stoichiometry proposed by the authors of the study [41]. Microbial C limitation aggravates with the increase in the vector length. The vector angle of < 45° indicates microbial N limitation, whereas the vector angle > 45° indicates microbial P limitation. Vector length and angles were computed using the formulae based on [41], ARCTG2 refers to arcus tangens:
Plant nutrient content
Analyses of the lettuce’s nutritive value were done as described by [36] after mineralization or incineration of individual samples of dry plant material (description at the end of section Pot experimental set-up). The nitrogen content was determined according to [42], using the Vario Macro Cube (Elementar Analysensysteme GmbH, Langenselbold, Germany). The phosphorus content was determined using spectrophotometry (Spectrophotometer: Onda VIS V-10 Plus, Giorgio Bormac, ITA) according to [43]. The potassium content was determined according to [44], and Ca and Mg determination was performed on the basis of the study [45]. All analyses and all measurements were performed using atomic absorption spectrometry (AAS; Agilent 55B AA; Agilent Technologies, CA, USA) according to [46].
Statistical analysis and data presentation
Data processing and statistical analyses were performed using software R, version 3.6.1. [47]. For characterization the relationship between the treatments and selected soil properties it was used principal component analysis (PCA), one-way analysis of variance (ANOVA) type I (sequential) sum of squares at 5% significance level [48]. Tukey's HSD (honestly significant difference) test and “treatment contrast” were utilized to detect statistically significant differences among factor level means and calculate factor level means for each treatment, respectively. The results were also graphically represented with help of Rohlf biplot for standardized PCA. Pearson correlation analysis was performed for measuring the linear dependence between soil properties. Pearson correlation coefficient was interpreted as follows: 0.0 < r < 0.3 (negligible correlation), 0.5 < r < 0.7 (low correlation), 0.5 < r < 0.7 (moderate correlation) and 0.7 < r < 0.9 (high correlation), 0.9 < r < 1.0 (very high correlation) [49].
Results
Soil chemical properties measured via pore water
Considerable differences were observed among the treatments for determined parameters. Soil pH differed little amongst treatment (⁓ pH 7.3), except of significantly lowered value in NB + MB (⁓ pH 7.2), whilst electrical conductivity was greatest in the case where mineral fertilizer was added to soil alone (MF; ~ 2400 mS cm−1) or in combination with biochar (NB + MF; > 3000 mS cm−1) (Fig. 1a, b). All other treatments were insignificantly different to the control (⁓ 800 mS cm−1).

Soil pore water parameters. Soil treatments: control, NB non-activated biochar, MF mineral fertilizers, NB + MF non-activated biochar + mineral fertilizers, AB activated biochar; properties—a pH, b electric conductivity (EC), c dissolved organic carbon (DOC), d total nitrogen (TN), e phosphorus (P), f potassium (K), g calcium (Ca), h magnesium (Mg), i sulfur (S). Mean values (n = 6) ± standard error of mean, different letter indicate differences among treatments at the statistical level of significance p ≤ 0.05
Total nitrogen, phosphorus, potassium, calcium, magnesium, and sulphur (TN, P, K, Ca, Mg, S) differed both within and between treatments, though a general trend was notable whereby the greatest median concentration or widest range was recorded from those treatments with mineral fertilizer addition but without activation (MF; NB + MF), especially so for N, Ca, Mg, S (Fig. 1d and g–i). The sorption capacity of the biochar was documented by the lower content of TN, Ca (significantly less in NB + MF than MF) and Mg, S (insignificant decrease in NB + MF compared to MF) in the pore water (Fig. 1d, g–i). Although at the same time a significant amount of nutrients was removed with the highest yield of plant biomass (Sect. 3.4, Fig. 4) after the application of a mixture of biochar and mineral fertilizers (NB + MF). Biochar without further additions (NB) introduced some significant amount of P, K, and S to the soil pore water, compared to control (Fig. 1e, f). Activation (AB) appeared to stabilize nutrients and hinder leaching of N, K, Ca, Mg and S (Fig. 1d, f). With a comparable production of aboveground biomass after the application of mineral fertilizer (MF) and the application of activated biochar (AB), the amount of N, K, Ca, Mg and S in the pore water after the lettuce harvest was minimal and similar as it was after the application of biochar alone.
Soil enzymes
Dehydrogenase activity (DHA) significantly greater than the control was observed only with the application of non-activated biochar (NB). While all other amendments remained statistically non-significant as compared to control (Fig. 2a). In contrast, all other enzyme activities of NB treatment differed from the control insignificantly (mostly comparable), Fig. 2b–f. In case of β-glucosidase (GLU), the highest activity was observed for MF treated soil followed by NB + MF and AB, which were significantly higher as compared to control and NB treatments (Fig. 2b). The same trend can be observed for phosphatase (Phos) activity, but MF, NB + MF, and AB values were all decreasing significantly, down to the lowest control and NB (Fig. 2c). Likewise, the soil amended with MF showed the significantly highest N-acetyl-β-D-glucosaminidase (NAG) activity followed by NB + MF, significantly more active compared to control (Fig. 2d). On the contrary, soil urease (Ure) values of MF and NB + MF were the only ones statistically lower than control (Fig. 2e). In case of arylsulfatase (ARS), MF treated soil was again the only statistically higher in comparison to control (Fig. 2f).

Soil enzyme activities. Properties—a dehydrogenase, b β-glucosidase, c phosphatase, d N-acetyl-β-D-glucosaminidase, e urease, f arylsulfatase. Mean values (n = 6) ± standard error of mean, different letters indicate differences among treatments at the statistical level of significance p ≤ 0.05
The enzyme activities were used to calculate nutrient calculation ratios, which showed following results: whereas mineral fertilizer-based treatments (MF, NB + MF, AB) showed increased C acquisition ratios compared to the control (with the highest value in MF), N acquisition ratio of the treatments MF and NB + MF (the smallest) were markedly lowered (Table 2). Vector parameter which indicates limitation by carbon was the highest in MF, lower (but increased compared to the control) in NB + MF, AB. All treatments showed limitation by P (based on the angle parameter values), the highest again in the MF treatment (Table 2).
Basal and substrate-induced microbial respirations
The soil microbial activity in terms of basal respiration (BR) and various substrates-induced respiration (-IR) was the most affected parameters under all the applied amendments (Fig. 3). Specifically, the soil BR was significantly affected by all amendments as compared to control treatment, whereby the significantly highest BR values were recorded for AB and NB + MF, respectively, relative to control (Fig. 3a). Similarly, in case of D-glucose-induced respiration (Glc-IR), AB amendment resulted in significantly highest respiration value as compared to control (Fig. 3b), while all other amendments significantly reduced the Glc-IR relative to control (Fig. 3b). The same trend was observed for N-acetyl-β-D-glucosamine induced respiration (NAG-IR) and D-mannose induced respiration (Man-IR), where AB resulted in significantly increased values as compared to control, while all other treatments reduced these activities relative to control (Fig. 3c–d). In case of D-trehalose induced respiration, the soil amendment with AB and NB + MF yielded significantly higher values of Tre-IR as compared to control and other treatments (Fig. 3e). The same treatments resulted in significantly highest L-alanine-induced respiration (Ala-IR) values as compared to control, however their effect remained statistically non-significant to each other (Fig. 3f). The strong loading of basal and substrate-induced respiration under the application of AB as shown by PCA (Additional file 1: Figure S1) further supports our obtained results (Fig. 3).

Soil respiration types. a Basal respiration and respirations induced by b D-glucose, c N-acetyl-β-D-glucosamine, d D-mannose, e L-alanine, f D-trehalose. Mean values (n = 4) ± standard error of mean, different letters indicate differences among treatments at the statistical level of significance p ≤ 0.05
Plant biomass
The aboveground (AGB) and root plant fresh and dry biomass were differently affected with the applied amendments. In all cases (except root dry biomass), the application of NB + MF showed significantly the highest values of plant biomass as compared to control treatment (Fig. 4). This trend was followed by the sole application of AB and sole MF, however their effect remained statistically non-significant with each other. In case of root fresh biomass, the same treatment NB + MF resulted in enhanced root fresh biomass as compared to other treatments and control, respectively (Fig. 4c). However, the sole application of NB showed significantly highest values for root dry weight (Root_dry) as compared to other treatments and control (Fig. 4d). Furthermore, the Pearson correlation showed that the plant biomass parameters were significantly positively correlated with Phos enzyme activity (Additional file 2: Figure S2).

Plant (lettuce) biomass. Fresh plant aboveground (colour columns above the axis X) and root (colour columns below the axis X) biomass, dry aboveground and root biomass expressed as the shaded parts of the respective fresh biomass value columns. Mean values ± standard error of mean, different letters indicate differences among treatments at the statistical level of significance p ≤ 0.05
Plant biomass quality
The determination of plant biomass quality showed that all three mineral fertilizer-based treatments (MF, NB + MF, AB) had significantly increased (compared to the control and NB) and similar content of nitrogen and magnesium (Fig. 5a, e). In contrast, the NB treatment had no significant positive effect on the content of plant nutrients (N, P, K, Ca, Mg); in fact, there was a decrease in N content (Fig. 5a). All soil amendments decreased lettuce P content in comparison to the control (Fig. 5b). Potassium was insignificantly (compared to the control) impacted by all amendments except of significantly increased value under NB + MF (Fig. 5c). The significantly highest Ca was in MF, other values were comparable (Fig. 5d).

Nutrient content in aboveground biomass. a Nitrogen, b phosphorus, c potassium, d calcium and e magnesium. Mean values ± standard error of mean, different letters indicate differences among treatments at the statistical level of significance p ≤ 0.05
Discussion
Effect of amendments on soil chemical properties
Biochar application into most experimental treatments did not significantly increase soil pH, despite its generally alkaline nature (Fig. 1a). The only treatment with a significant decrease in pH was NB + MF, the lowest soil pH value could be caused by an combination of slightly acidic MF mixture and the acidified activated biochar (which pH was probably lowered due to the interaction with acidic activation solution). The main component of MF was FOSMAG fertilizer based mainly on acidic phosphates. The lowest pH was coupled with significantly increased activity of NAG (Fig. 2d). Furthermore, NB + MF-derived slight acidification could be related to enhanced cation exchange capacity, which was the second highest (lower compared to MF but higher than the other treatments) (Fig. 1b). Biochar is a material abundant with cation exchange sites which may enhance the adsorption of acidifying ions (e.g. HPO42−, H2PO4.−) produced through applied fertilizer dissolution and hydrolysis [8, 50, 51]
The amount of DOC in pore water was significantly higher after application of mineral fertilizers, alone or combined with biochar, Fig. 1c. The significant increase in DOC after application of mineral fertilizers or their mixture with biochar may be related to the priming effect [52]. The MF supply of available nutrients was used by microorganisms to decompose soil organic matter more rapidly and release simpler and soluble forms of organic carbon (e.g. fulvic acids). This presumption was corroborated by Pearson’s correlation results (p ≤ 0.001): DOC correlated highly positively (p ≤ 0.001) with TN, Ca, Mg, S, and moderately with NAG, Phos (Additional file 2: Figure S2). On the contrary, co-amendment biochar (prepared at moderately high temperatures) comprises carbon fractions with various recalcitrance which prevents short-termed markable dissolution of DOC into the pore water [53].
This study demonstrates that the addition of soluble salts of nitrogen, phosphorus, and sulfur to soil with mineral fertilizers, as seen in the pore water enhanced electrical conductivity under the sole application of MF (either alone or combined with NB, Fig. 1b), resulted in the highest element content (excluding potassium) of all experimental treatments. One of the key biochar’s properties is their large surface area–volume ratio, orders of magnitude greater than the standard mineral components of soils. In this regard, they have been proven to act as sorbents, regulating the leaching of added nutrients and trace elements in soils by the combined action of physical retention of moisture, and sorption of ions to their surfaces [54]. This presumption corresponded to the concurrently increased content of all nutrients (except of K) in the respective soil treatment NB + MF and to increased P content in the AB treatment. Altogether these ions induced extremely high EC (supraoptimal to most of crops, which prefer EC ≤ 2500 mS cm−1) which EC hinders nutrient uptake by increasing the osmotic pressure of the nutrient solution. The discharge of nutrients into the environment [55] has been reported to impede mineral uptake by soil microorganisms, and a high electrical conductivity (EC) has been associated with a negative impact on microbial carbon mineralization in soil [56]. In contrast to this, biochar was reported to increased EC values in amended soil [57, 58], albeit low pyrolysis temperatures (350–600 °C) was referred to promote lower EC values [59].
Biochar itself is a source of phosphorus and potassium [60], which was confirmed in the treatment NB (Fig. 1e, f). The addition of biochar resulted in higher P and K contents in extracted pore water. Moreover, biochar provides reactive surfaces where P and K ions are retained in soil microbial biomass and in exchange sites, leading to increasing the availability of P and K to plants uptake [60]. In particular, phosphorus content is higher in pore water on all biochar-based treatments (Fig. 1), reflecting the described ability of biochar to sorb phosphate due to its porous structure, large surface area and abundant surface groups [61]. Phosphorus adsorption can be influenced by various process factors such as solution pH, adsorbent dosage, co-existing ions, and reaction temperature [61]. The comparatively higher concentration of phosphorus in pore water after application of mineral fertilizer and mineral fertilizer with biochar (Fig. 1) was also due to the form of P in the applied P fertilizer, where 40% of the P was in insoluble micromilled phosphate.
Effect of amendments on soil enzymes
Biochar addition to soil has been recognized to stimulate soil enzyme activities related to C, N and P cycling. We found differential responses of C, N and P cycling enzymes subjected to various amendments (Fig. 2). Specifically, the NB application demonstrated the highest putative dehydrogenase activity (DHA), which is a measure of microbial degradation of SOM (Fig. 2a). We ascribe this to be associated with enhanced microbial population related to carbon cycling under NB application. This is in accordance with the findings of other authors [34, 62]. They found increased activity of DHA under applied biochar amendment. The β-glucosidase (GLU) activity was found highest in MF treated soils followed by NB + MF and AB (Fig. 2b). Similar results were reported by [13, 24], the authors ascribed the increased activity of GLU due to the more rapid microbial turnover with the addition of NPK, a pattern similar to the applied (NB + MF) treatment in our study. The carbon acquisition ratio values (Table 2) were markedly increased over the control value in all MF-based treatments (the highest in MF, then NB + MF, and AB) and it indicated indeed high C utilization, as documented also by synergy between GLU, other enzymes and aboveground, root biomass (PCA, Additional file 1: Figure S1) and by high positive Pearson’s correlation (p ≤ 0.001) with Phos, ARS as well as moderate correlation with AGB_fresh, AGB_dry (Additional file 2: Figure S2). Similarly, phosphatase (Phos) activity was the highest in MF, followed by NB + MF and AB treatment (Fig. 2c). Such alteration in Phos activity of richly supplied soil (via mineral fertilization) could be also attributed to changes in soil pH as depicted by [25], that biochar-derived changes in soil pH are the key mediators of Phos activity in soils. In addition to GLU and Phos activity, the enhancement of nitrogen mining enzyme N-acetyl-β-D-glucosaminidase (NAG) through MF and NB + MF amendments could be related to the increased nitrogen availability to microbes which was in accord with the findings of [63]. PCA synergy (Additional file 1: Figure S1) and Pearson’s high correlation (Additional file 2: Figure S2) of NAG with TN, DOC, Phos, and ARS corroborated the previously reported, that the application of fertilizers can significantly induce the activity of these enzymes [64]. Arylsulfatase-mediated organosulfur mineralization was only significantly increased in MF treatment too (Fig. 2f). Our results agree with [13], which study referred to increased activity of C and N degrading enzymes under the application of biochar and mineral fertilizer combinations. However, urease activity, an enzyme responsible for the release of ammonium from organic nitrogenous compounds, demonstrated no significant changes in all treatments, and even decreased in the MF treatment (Fig. 2e). It could be assumed lower demand for ammonium nitrogen in the biochar-treated treatments due to the high adsorption potential of pyrolyzed matter [50]. The results and assumptions lead to the conclusion that, compared to the control, both MF and NB + MF showed a significantly lower N acquisition ratio (Table 2).
Effect of amendments on basal and substrate-induced microbial respirations
The exogenous addition of biochar and different (including mineral) fertilizers has been shown to stimulate soil microbial respiration [34, 65, 66]. In the present study, the soil basal as well as some substrate-induced respiration (Tre-, Ala-IR) was significantly enhanced by the co-applied amendments, biochar and MF (Fig. 3). AB showed the strongest enhancement in respiration types, which were the parameters most affected by activated biochar, as hypothesized. This could be due to the fact that biochar might act as C source for microbes and that the AB was produced by value addition of nutrients (mainly of N, P). Hence the availability of readily soluble nutrients stimulated microbial communities resulting in enhanced microbial respiration [67, 68]. Moreover, this enhancement of microbial respirations under the AB and NB + MF treatments might be due to the enhanced dissolution of soil carbonates through biochar-derived organic acids and offering a suitable niche for microbes to degrade readily available C compounds (relevant for SIR) and finally due to the ability of biochar to adsorb organic compounds on its surface resulting in more microbial growth and hence enhanced respiration [69,70,71]. Despite enhanced carbon utilization, as evidenced by increased C acquisition ratio values of NB + MF and AB (Table 2), carbon limitation was observed in these treatments (vector parameter in Table 2), likely a consequence of accelerated consumption of labile carbon (increasing from NB + MF to AB) (Fig. 1c). Similar enhancing effect of biochar amendment on the activity of specific microorganisms in soil, leading to degradation of labile soil organic matter, was referred [72]. The highest C limitation was calculated for MF treatment (Table 2), which finding may explain the most decreased values of all determined respiration types (Fig. 3a–f).
Effect of amendments on plant biomass
It has been described that biochar addition and fertilization impact positively on crop productivity. These positive effects are associated with improved physico-chemical properties of the soil including, but not limited to, soil structure modification, enhanced nutrient retention capacity, increased water holding capacity of the soils and improved soil aeration leading to better root development [22, 73,74,75]. In addition, it has been reported previously that the application of biochar alone and or in combination with other organic and inorganic fertilizers improved gas exchange and physiological parameters (e.g. photosynthetic rate, transpiration, water use efficiency) of plants [76,77,78]. The application of biochar (NB) alone and combined with mineral fertilizer (NB + MF) significantly enhanced lettuce biomass in the present study (Fig. 4). The possible explanation behind such increases might be the increased pH, EC, and associated plant nutrients in the soil, which changes may consequently result in enhanced biomass accumulation [8, 21, 51]. Our results were in accordance with the findings of [79], where enhanced growth of tomato plants due to applied biochar was reported. The authors clarified the role of biochar to retain NH4+ ions on its surface which ultimately enhanced N availability to plant resulting in increased biomass accumulation. This shows the direct involvement of biochar in plant growth promotion together with other readily available nutrient sources (mineral fertilizer) in the present study. However, the improved accumulation of nutrients from soil in plant biomass was coupled (for most determined elements, mainly N, Mg, and K) strictly with mineral fertilization, i.e. presumably biochar was capable to absorb nutrient during activation phase as well as stabilize them in soil when co-applied with MF (exerted in the treatments NB + MF and AB, Fig. 5). Similar positive effect of biochar on N and K uptake into plant biomass if combined with mineral fertilizer was reported [80, 81]. The enhanced activity of nutrient solubilizing enzymes (GLU, Phos NAG, ARS, Ure) in the present study (Fig. 2b–f) also supports the hypothesis that biochar addition enhances soil nutrients and improve their availability to plants which ultimately accumulate more biomass. This was further corroborated by the results of Pearson’s correlation, whereby Phos, NAG, GLU activity was significantly (p ≤ 0.05 or less) positively linked to plant biomass features AGB_fresh and AGB_dry (moderate-to-high correlation), Additional file 2: Figure S2. Furthermore, AGB_fresh and AGB_dry highly correlated with N-biomass and Mg-biomass (p ≤ 0.001, Additional file 2: Figure S2). In contrast to expectations, biochar had no effect on plant calcium content (Fig. 5d), and did not enhance plant phosphorus uptake (Figs. 2c, 5c). Mineral fertilization likely increased soluble phosphorus, however this was not taken up by plants or microbes at a rate sufficient to prevent phosphorus limitation (indicated by the angle parameter in Table 2). Calcium was reported to be strongly adsorbed by biochar and also decreases availability of phosphorus when complexed with P [82], thus, only MF treatment provided increased Ca content in the lettuce biomass, in the line with the previous reports of similar research [83].
Conclusion
In this study, the impacts on several soil health indicators and geochemical parameters of combinations of biochar with and without mineral fertilizer were tested. Soil health indicators (soil enzymes, basal and substrate-induced respiration) responded favourably to all soil amendments, resulting in enhanced nutrient acquisition enzymes (C, N and P) which resulted in improved soil fertility and consequent higher crop (lettuce) biomass. Nutrient content in the plant biomass was the most dependent on the mineral fertilization, albeit the increased element accumulation was provided also by activated biochar. The activation of biochar by pre-mixing rather than merely co-applying mineral fertilizer was shown to reduce the leaching of N, P from added fertilizing amendments, demonstrating the longevity benefits of this pre-application stage. Research is now required to determine whether the initial trends seen in this study are reproducible at a field scale and thus be recommended for soil amendment in some agronomic circumstances.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AB:
-
Activated biochar
- AGB:
-
Aboveground biomass
- Ala:
-
L-Alanine
- ANOVA:
-
One-way analysis of variance
- ARS:
-
Arylsulfatase
- BET:
-
Brunauer–Emmett–Teller surface
- BR:
-
Basal soil respiration
- DHA:
-
Dehydrogenase activity
- DM:
-
Dry matter
- DOC:
-
Dissolved organic carbon
- EC:
-
Electrical conductivity
- Glc:
-
D-Glucose
- GLU:
-
β-Glucosidase
- Man:
-
D-Mannose
- MF:
-
Mineral fertilizer
- NAG:
-
N-Acetyl-β-D-glucosaminidase
- NAG-IR:
-
Respiration induced by N-acetyl-β-D-glucosamine
- NB:
-
(Non-activated) biochar
- p:
-
P-value
- PCA:
-
Principal component analysis
- Phos:
-
Phosphatase
- r:
-
Correlation coefficient
- SIR:
-
Substrate-induced soil respiration
- SOM:
-
Soil organic matter
- TN:
-
Total nitrogen
- TOC:
-
Total organic carbon
- Tre:
-
D-Trehalose
- Ure:
-
Urease
References
Mustafa A, Hu X, Abrar MM, Shah SAA, Nan S, Saeed Q, et al. Long-term fertilization enhanced carbon mineralization and maize biomass through physical protection of organic carbon in fractions under continuous maize cropping. Appl Soil Ecol. 2021;165:103971. https://doi.org/10.1016/j.apsoil.2021.103971.
West PC, Gerber JS, Engstrom PM, Mueller ND, Brauman KA, Carlson KM, et al. Leverage points for improving global food security and the environment. Science. 2014;345(6194):325–8. https://doi.org/10.1126/science.1246067.
Hairani A, Osaki M, Watanabe T. Effect of biochar application on mineral and microbial properties of soils growing different plant species. Soil Sci Plant Nutr. 2016;62(5–6):519–25. https://doi.org/10.1080/00380768.2016.1212648.
Holatko J, Bielska L, Hammerschmiedt T, Kucerik J, Mustafa A, Radziemska M, et al. Cattle manure fermented with biochar and humic substances improve the crop biomass, microbiological properties and nutrient status of soil. Agronomy. 2022;12(2):368. https://doi.org/10.3390/agronomy12020368.
Saletnik B, Zagula G, Bajcar M, Czernicka M, Puchalski C. Biochar and biomass ash as a soil ameliorant: the effect on selected soil properties and yield of giant Miscanthus (Miscanthus x giganteus). Energies. 2018;11(10):2535. https://doi.org/10.3390/en11102535.
Mustafa A, Minggang X, Ali Shah SA, Abrar MM, Nan S, Baoren W, et al. Soil aggregation and soil aggregate stability regulate organic carbon and nitrogen storage in a red soil of southern China. J Environ Manage. 2020;270:110894. https://doi.org/10.1016/j.jenvman.2020.110894.
Mukherjee A, Lal R. Biochar impacts on soil physical properties and greenhouse gas emissions. Agronomy. 2013;3(2):313–39. https://doi.org/10.3390/agronomy3020313.
Sohi SP, Krull E, Lopez-Capel E, Bol R. A review of biochar and its use and function in soil. Advances in agronomy. Amsterdam: Elsevier; 2010. p. 47–82.
Hammerschmiedt T, Holatko J, Pecina V, Huska D, Latal O, Kintl A, et al. Assessing the potential of biochar aged by humic substances to enhance plant growth and soil biological activity. Chem Biol Technol Agric. 2021. https://doi.org/10.1186/s40538-021-00242-7.
Masulili A, Utomo WH, Ms S. Rice husk biochar for rice based cropping system in acid soil 1. The characteristics of rice husk biochar and its influence on the properties of acid sulfate soils and rice growth in West Kalimantan, Indonesia. J Agric Sci. 2010. https://doi.org/10.5539/jas.v2n1p39.
Yuan J-H, Xu R-K, Qian W, Wang R-H. Comparison of the ameliorating effects on an acidic ultisol between four crop straws and their biochars. J Soils Sediments. 2011;11(5):741–50. https://doi.org/10.1007/s11368-011-0365-0.
Hammer EC, Forstreuter M, Rillig MC, Kohler J. Biochar increases arbuscular mycorrhizal plant growth enhancement and ameliorates salinity stress. Appl Soil Ecol. 2015;96:114–21. https://doi.org/10.1016/j.apsoil.2015.07.014.
Song D, Chen L, Zhang S, Zheng Q, Ullah S, Zhou W, et al. Combined biochar and nitrogen fertilizer change soil enzyme and microbial activities in a 2-year field trial. Eur J Soil Biol. 2020;99:103212. https://doi.org/10.1016/j.ejsobi.2020.103212.
Glaser B, Lehmann J, Zech W. Ameliorating physical and chemical properties of highly weathered soils in the tropics with charcoal - a review. Biol Fertil Soils. 2002;35(4):219–30. https://doi.org/10.1007/s00374-002-0466-4.
Chan KY, Van Zwieten L, Meszaros I, Downie A, Joseph S. Using poultry litter biochars as soil amendments. Soil Research. 2008;46(5):437. https://doi.org/10.1071/sr08036.
Lehmann J, da Silva Jr JP, Steiner C, Nehls T, Zech W, Glaser B. Nutrient availability and leaching in an archaeological anthrosol and a ferralsol of the central amazon basin: fertilizer, manure and charcoal amendments. Plant Soil. 2003;249(2):343–57. https://doi.org/10.1023/a:1022833116184.
Hussain M, Farooq M, Nawaz A, Al-Sadi AM, Solaiman ZM, Alghamdi SS, et al. Biochar for crop production: potential benefits and risks. J Soils Sediments. 2016;17(3):685–716. https://doi.org/10.1007/s11368-016-1360-2.
Saha A, Basak BB, Gajbhiye NA, Kalariya KA, Manivel P. Sustainable fertilization through co-application of biochar and chemical fertilizers improves yield, quality of Andrographis paniculata and soil health. Ind Crops Prod. 2019;140:111607. https://doi.org/10.1016/j.indcrop.2019.111607.
Tagoe SO, Horiuchi T, Matsui T. Effects of carbonized and dried chicken manures on the growth, yield, and N content of soybean. Plant Soil. 2008;306:211–20.
Alburquerque JA, Salazar P, Barrón V, Torrent J, del Campillo MdC, Gallardo A, et al. Enhanced wheat yield by biochar addition under different mineral fertilization levels. Agron Sustain Dev. 2013;33(3):475–84. https://doi.org/10.1007/s13593-012-0128-3.
Scotti R, Bonanomi G, Scelza R, Zoina A, Rao MA. Organic amendments as sustainable tool to recovery fertility in intensive agricultural systems. J Soil Sci Plant Nutr. 2015. https://doi.org/10.4067/s0718-95162015005000031.
Schulz H, Glaser B. Effects of biochar compared to organic and inorganic fertilizers on soil quality and plant growth in a greenhouse experiment. J Plant Nutr Soil Sci. 2012;175(3):410–22. https://doi.org/10.1002/jpln.201100143.
Atkinson CJ, Fitzgerald JD, Hipps NA. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: a review. Plant Soil. 2010;337(1–2):1–18. https://doi.org/10.1007/s11104-010-0464-5.
Tian J, Wang J, Dippold M, Gao Y, Blagodatskaya E, Kuzyakov Y. Biochar affects soil organic matter cycling and microbial functions but does not alter microbial community structure in a paddy soil. Sci Total Environ. 2016;556:89–97. https://doi.org/10.1016/j.scitotenv.2016.03.010.
Lehmann J, Rillig MC, Thies J, Masiello CA, Hockaday WC, Crowley D. Biochar effects on soil biota—a review. Soil Biol Biochem. 2011;43(9):1812–36. https://doi.org/10.1016/j.soilbio.2011.04.022.
Bastida F, Luis Moreno J, Teresa H, García C. Microbiological degradation index of soils in a semiarid climate. Soil Biol Biochem. 2006;38(12):3463–73. https://doi.org/10.1016/j.soilbio.2006.06.001.
Gul S, Whalen JK, Thomas BW, Sachdeva V, Deng H. Physico-chemical properties and microbial responses in biochar-amended soils: Mechanisms and future directions. Agr Ecosyst Environ. 2015;206:46–59. https://doi.org/10.1016/j.agee.2015.03.015.
Bamminger C, Marschner B, Jüschke E. An incubation study on the stability and biological effects of pyrogenic and hydrothermal biochar in two soils. Eur J Soil Sci. 2014;65(1):72–82. https://doi.org/10.1111/ejss.12074.
Khadem A, Raiesi F. Responses of microbial performance and community to corn biochar in calcareous sandy and clayey soils. Appl Soil Ecol. 2017;114:16–27. https://doi.org/10.1016/j.apsoil.2017.02.018.
Lehmann J, Joseph S. Biochar for environmental management: science, technology and implementation. London: Routledge; 2015.
Warnock DD, Mummey DL, McBride B, Major J, Lehmann J, Rillig MC. Influences of non-herbaceous biochar on arbuscular mycorrhizal fungal abundances in roots and soils: results from growth-chamber and field experiments. Appl Soil Ecol. 2010;46(3):450–6. https://doi.org/10.1016/j.apsoil.2010.09.002.
Hale L, Luth M, Crowley D. Biochar characteristics relate to its utility as an alternative soil inoculum carrier to peat and vermiculite. Soil Biol Biochem. 2015;81:228–35. https://doi.org/10.1016/j.soilbio.2014.11.023.
Anderson CR, Condron LM, Clough TJ, Fiers M, Stewart A, Hill RA, et al. Biochar induced soil microbial community change: Implications for biogeochemical cycling of carbon, nitrogen and phosphorus. Pedobiologia. 2011;54(5–6):309–20. https://doi.org/10.1016/j.pedobi.2011.07.005.
Brtnicky M, Datta R, Holatko J, Bielska L, Gusiatin ZM, Kucerik J, Hammerschmiedt T, Danish S, Radziemska M, Mravcova L, Fahad S, Kintl A, Sudoma M, Ahmed N, Pecina. A critical review of the possible adverse effects of biochar in the soil environment. Sci Total Environ. 2021;796:18. https://doi.org/10.1016/j.scitotenv.2021.148756.
Beheshti M, Etesami H, Alikhani HA. Effect of different biochars amendment on soil biological indicators in a calcareous soil. Environ Sci Pollut Res Int. 2018;25(15):14752–61. https://doi.org/10.1007/s11356-018-1682-2.
Trakal L, Raya-Moreno I, Mitchell K, Beesley L. Stabilization of metal(loid)s in two contaminated agricultural soils: comparing biochar to its non-pyrolysed source material. Chemosphere. 2017;181:150–9. https://doi.org/10.1016/j.chemosphere.2017.04.064.
Doi R, Ranamukhaarachchi SL. Soil dehydrogenase in a land degradation-rehabilitation gradient: observations from a savanna site with a wet/dry seasonal cycle. Rev Biol Trop. 2009;57:223–34. https://doi.org/10.15517/rbt.v57i1-2.11317.
Campbell CD, Chapman SJ, Cameron CM, Davidson MS, Potts JM. A rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil. Appl Environ Microbiol. 2003;69(6):3593–9. https://doi.org/10.1128/AEM.69.6.3593-3599.2003.
ISO_20130. Soil quality — Measurement of enzyme activity patterns in soil samples using colorimetric substrates in micro-well plates. Geneva, Switzerland: International Organization for Standardization; 2018.
Cui J, Zhang S, Wang X, Xu X, Ai C, Liang G, et al. Enzymatic stoichiometry reveals phosphorus limitation-induced changes in the soil bacterial communities and element cycling: evidence from a long-term field experiment. Geoderma. 2022;426:116124. https://doi.org/10.1016/j.geoderma.2022.116124.
Moorhead DL, Sinsabaugh RL, Hill BH, Weintraub MN. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol Biochem. 2016;93:1–7. https://doi.org/10.1016/j.soilbio.2015.10.019.
Lu RK. Analytical methods of soil agrochemistry. Beijing, China: Chinese Agriculture Science and Technology Press; 1999.
Olsen SR, Sommers LE. Phosphorus. In: Page AL, editor. Methods of soil analysis: part 2 chemical and microbiological properties. 2nd ed. Madison, USA: ASA and SSSA; 1982. p. 403–30.
Nowosielski O. Methods for determination of fertilisation requirements. 2nd ed.; PWRiL: Warsaw, Poland. 1974.
Ryan J, Estefan G, Rashid A. Soil and plant analysis lab manual. Aleppo, Syria: ICARDA; 2001.
Jones Jr. Laboratory guide for conducting soil tests and plant analysis. Boca Raton: CRC Press; 2001. https://doi.org/10.1201/9781420025293.
R_Core_Team. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2020.
Zar JH. Biostatistical analysis. 2nd ed. Englewood Cliffs: Prentice-Hall, Inc.; 1984.
Hinkle DE, Wiersma W, Jurs SG. Applied statistics for the behavioral sciences. 5th ed. Boston, Mass: Houghton Mifflin; 2003.
Hollister CC, Bisogni JJ, Lehmann J. Ammonium, nitrate, and phosphate sorption to and solute leaching from biochars prepared from corn stover (L.) and oak wood (spp.). J Environ Qual. 2013;42(1):137–44. https://doi.org/10.2134/jeq2012.0033.
Jones DL, Rousk J, Edwards-Jones G, DeLuca TH, Murphy DV. Biochar-mediated changes in soil quality and plant growth in a three year field trial. Soil Biol Biochem. 2012;45:113–24. https://doi.org/10.1016/j.soilbio.2011.10.012.
Bird JA, Herman DJ, Firestone MK. Rhizosphere priming of soil organic matter by bacterial groups in a grassland soil. Soil Biol Biochem. 2011;43:718–25.
Feng Z, Fan Z, Song H, Li K, Lu H, Liu Y, et al. Biochar induced changes of soil dissolved organic matter: the release and adsorption of dissolved organic matter by biochar and soil. Sci Total Environ. 2021;783:147091. https://doi.org/10.1016/j.scitotenv.2021.147091.
Mitchell K, Beesley L, Šípek V, Trakal L. Biochar and its potential to increase water, trace element, and nutrient retention in soils. Biochar in agriculture for achieving sustainable development goals. Amsterdam: Elsevier; 2022. p. 25–33.
Samarakoon U, Weerasinghe P, Weerakkody W. Effect of electrical conductivity (EC) of the nutrient solution on nutrient uptake, growth and yield of leaf lettuce (Lactuca sativa L.) in stationary culture. Tropical Agricultural Research 2006;18:13–21.
Dong Y, Chen RR, Petropoulos E, Yao TY, Yu BQ, Lin XG, et al. Microbial carbon use efficiency in coastal soils along a salinity gradient revealed by ecoenzymatic stoichiometry. J Geophys Res Biogeosci. 2022;127(8):11. https://doi.org/10.1029/2022jg006800.
Nawaz H. Soil electrical conductivity as affected by biochar under summer crops. Int J Environ Sci Nat Resour. 2018. https://doi.org/10.19080/ijesnr.2018.14.555887.
Rehrah D, Reddy MR, Novak JM, Bansode RR, Schimmel KA, Yu J, et al. Production and characterization of biochars from agricultural by-products for use in soil quality enhancement. J Anal Appl Pyrol. 2014;108:301–9. https://doi.org/10.1016/j.jaap.2014.03.008.
Zeka M, Njira KOW, Bailey L, Chimungu JG, Ligowe IS, Nalivata PC. 2022. https://doi.org/10.21203/rs.3.rs-2173583/v1 (preprint)
Zhao Y, Li X, Li Y, Bao H, Xing J, Zhu Y, et al. Biochar acts as an emerging soil amendment and its potential ecological risks: a review. Energies. 2022;16(1):410. https://doi.org/10.3390/en16010410.
Luo D, Wang L, Nan H, Cao Y, Wang H, Kumar TV, et al. Phosphorus adsorption by functionalized biochar: a review. Environ Chem Lett. 2022. https://doi.org/10.1007/s10311-022-01519-5.
Sial TA, Lan Z, Wang L, Zhao Y, Zhang J, Kumbhar F, et al. Effects of different biochars on wheat growth parameters, yield and soil fertility status in a silty clay loam soil. Molecules. 2019;24(9):1798. https://doi.org/10.3390/molecules24091798.
Zhang L, Sun X-Y, Tian Y, Gong X-Q. Biochar and humic acid amendments improve the quality of composted green waste as a growth medium for the ornamental plant Calathea insignis. Sci Hortic. 2014;176:70–8. https://doi.org/10.1016/j.scienta.2014.06.021.
Andert J, Mumme J. Impact of pyrolysis and hydrothermal biochar on gas-emitting activity of soil microorganisms and bacterial and archaeal community composition. Appl Soil Ecol. 2015;96:225–39. https://doi.org/10.1016/j.apsoil.2015.08.019.
Brtnicky M, Hammerschmiedt T, Elbl J, Kintl A, Skulcova L, Radziemska M, et al. The potential of biochar made from agricultural residues to increase soil fertility and microbial activity: impacts on soils with varying sand content. Agronomy. 2021;11(6):1174. https://doi.org/10.3390/agronomy11061174.
Zavalloni C, Alberti G, Biasiol S, Vedove GD, Fornasier F, Liu J, et al. Microbial mineralization of biochar and wheat straw mixture in soil: a short-term study. Appl Soil Ecol. 2011;50:45–51. https://doi.org/10.1016/j.apsoil.2011.07.012.
Asirifi I, Werner S, Heinze S, Saba CKS, Lawson IYD, Marschner B. Short-term effect of biochar on microbial biomass, respiration and enzymatic activities in wastewater irrigated soils in Urban Agroecosystems of the West African Savannah. Agronomy. 2021;11(2):271. https://doi.org/10.3390/agronomy11020271.
Haring V, Manka’abusi D, Akoto-Danso EK, Werner S, Atiah K, Steiner C, et al. Effects of biochar, waste water irrigation and fertilization on soil properties in West African urban agriculture. Sci Rep. 2017;7(1):10738. https://doi.org/10.1038/s41598-017-10718-y.
Jin H, Lehmann J, Thies JE. Soil microbial community response to amending corn soils with corn stover charcoal. Conference of International Biochar Initiative. Newcastle, UK 2008.
Steinbeiss S, Gleixner G, Antonietti M. Effect of biochar amendment on soil carbon balance and soil microbial activity. Soil Biol Biochem. 2009;41(6):1301–10. https://doi.org/10.1016/j.soilbio.2009.03.016.
Tamir G, Shenker M, Heller H, Bloom PR, Fine P, Bar-Tal A. Can soil carbonate dissolution lead to overestimation of soil respiration? Soil Sci Soc Am J. 2011;75(4):1414–22. https://doi.org/10.2136/sssaj2010.0396.
Mitchell PJ, Simpson AJ, Soong R, Simpson MJ. Shifts in microbial community and water-extractable organic matter composition with biochar amendment in a temperate forest soil. Soil Biol Biochem. 2015;81:244–54. https://doi.org/10.1016/j.soilbio.2014.11.017.
Chan KY, Van Zwieten L, Meszaros I, Downie A, Joseph S. Agronomic values of greenwaste biochar as a soil amendment. Soil Res. 2007;45(8):629. https://doi.org/10.1071/sr07109.
Lair GJ, Gerzabek MH, Haberhauer G, Jakusch M, Kirchmann H. Response of the sorption behavior of Cu, Cd, and Zn to different soil management. J Plant Nutr Soil Sci. 2006;169(1):60–8. https://doi.org/10.1002/jpln.200521752.
Lima JRdS, de Moraes SW, de Medeiros EV, Duda GP, Corrêa MM, Martins Filho AP, et al. Effect of biochar on physicochemical properties of a sandy soil and maize growth in a greenhouse experiment. Geoderma. 2018;319:14–23. https://doi.org/10.1016/j.geoderma.2017.12.033.
Trupiano D, Cocozza C, Baronti S, Amendola C, Vaccari FP, Lustrato G, et al. The effects of biochar and its combination with compost on lettuce (Lactuca sativa L.) growth, soil properties, and soil microbial activity and abundance. Int J Agron. 2017;2017:1–12. https://doi.org/10.1155/2017/3158207.
Uzoma KC, Inoue M, Andry H, Fujimaki H, Zahoor A, Nishihara E. Effect of cow manure biochar on maize productivity under sandy soil condition. Soil Use Manag. 2011;27(2):205–12. https://doi.org/10.1111/j.1475-2743.2011.00340.x.
Viger M, Hancock RD, Miglietta F, Taylor G. More plant growth but less plant defence? First global gene expression data for plants grown in soil amended with biochar. GCB Bioenergy. 2015;7(4):658–72. https://doi.org/10.1111/gcbb.12182.
Vaccari FP, Maienza A, Miglietta F, Baronti S, Di Lonardo S, Giagnoni L, et al. Biochar stimulates plant growth but not fruit yield of processing tomato in a fertile soil. Agr Ecosyst Environ. 2015;207:163–70. https://doi.org/10.1016/j.agee.2015.04.015.
Borchers A, Pieler T. Programming pluripotent precursor cells derived from Xenopus embryos to generate specific tissues and organs. Genes (Basel). 2010;1(3):413–26. https://doi.org/10.3390/genes1030413.
Huang C, Sun X, Wang L, Storer P, Siddique KHM, Solaiman ZM. Nutrients leaching from tillage soil amended with wheat straw biochar influenced by fertiliser type. Agriculture. 2021;11(11):1132.
Luo D, Wang L, Nan H, Cao Y, Wang H, Kumar TV, et al. Phosphorus adsorption by functionalized biochar: a review. Environ Chem Lett. 2023;21(1):497–524. https://doi.org/10.1007/s10311-022-01519-5.
Ogunyemi AM, Otegbayo BO, Fagbenro JA. Effects of NPK and biochar fertilized soil on the proximate composition and mineral evaluation of maize flour. Food Sci Nutr. 2018;6(8):2308–13. https://doi.org/10.1002/fsn3.808.
Acknowledgements
Not applicable.
Funding
This research was supported by the project FCH-S-23-8297 of Ministry of Education, Youth and Sports of the Czech Republic, by the Ministry of Agriculture of the Czech Republic, institutional support MZE-RO1223 and with the Institutional support of the Ministry of Agriculture of the Czech Republic in the frame of the Long-Term Conception of the Development of the Research Organization Agricultural Research, Ltd. Troubsko.
Author information
Authors and Affiliations
Contributions
Conceptualization, TH, AM, LB and MB; methodology, MB, JH, PR, AK, and AM; resources AK, TB, JH, LT, COO, and AM; writing—original draft preparation, MB, AM, JH; visualization, TH, COO, and TB; supervision, PR, LT, and JH; project administration, MB, AK, LT and JH. All authors contributed to the data interpretation, writing—review and editing and final approval for publication.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
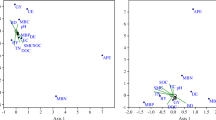
Additional file 1: Figure S1.
Rolf plot for principal component analysis (PCA).
Additional file 2: Figure S2.
Pearson correlation matrix. The asterisks indicate a level of significance in statistical difference between the variables: ˙ for p ≤ 0.1, * for p ≤ 0.05, ** for p ≤ 0.01, *** for p ≤ 0.001.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

{kind=link}
{kind=link}
Cite this article
Brtnicky, M., Mustafa, A., Hammerschmiedt, T. et al. Pre-activated biochar by fertilizers mitigates nutrient leaching and stimulates soil microbial activity. Chem. Biol. Technol. Agric. 10, 57 (2023). https://doi.org/10.1186/s40538-023-00430-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-023-00430-7