
Abstract
Background
Conventional agriculture relies on chemical pesticides and fertilizers, which can degrade ecosystems. A reduction of these harmful practices is required, replacing (or integrating) them with more eco-friendly approaches, such as microbial inoculation. Tomato is an important agricultural product, with a high content of bioactive compounds (folate, ascorbate, polyphenols, and carotenoids). The focus of this research was to investigate the plant growth-promoting (PGP) abilities of bacterial strains isolated from different tomato cultivars, with the aim to develop systems to improve plant health and crop productivity based on microbial inoculation.
Methods
A pool of different tomato cultivars already available on the market and new tomato hybrids were selected based on their nutritional quality (high content of biologically active compounds). A total of 23 strains were isolated from tomato roots (11 rhizospheric strains and 12 root endophytes). The cultivable isolates were analyzed for a number of different PGP traits: organic acids (OA), indole acetic acid (IAA), ACC deaminase, and siderophore production. The effects of microbial inoculation on root growth of Arabidopsis thaliana were also evaluated using a Vertical Agar Plate assay.
Results
A high percentage of the isolated strains tested positive for the following PGP traits: 73 % were able to produce OA, 89 % IAA, 83 % ACC deaminase, and 87 % siderophores. The most striking result were remarkable increases in the formation of root hairs for most of the inoculated plants. This effect was obvious for all A. thaliana seedlings inoculated with the isolated endophytes, and for the 50 % of the seedlings inoculated with the rhizospheric strains.
Conclusions
A better knowledge of the plant growth-promotion activity of these strains can provide an important contribution to increase environmental sustainability in agriculture.

Screening of plant growth-promoting (PGP) effects of bacteria associated with different tomato cultivars.
Similar content being viewed by others


Background
Feeding an increasing number of people is one of the major challenges of the twenty-first century [1, 2]. Established strategies to enhance crop productivity are the use of chemical fertilizers, manures, and pesticides. However, these approaches often have a negative impact on the environment: leaching of nitrate into groundwater, surface run-off of phosphorus and nitrogen, and eutrophication of aquatic ecosystems [3]. The growing interest in environmental sustainability has led to considerable efforts to minimize the use of chemical fertilizers and pesticides, replacing (or integrating) these conventional approaches with more eco-friendly methods, such as the application of beneficial microorganisms [4–6]. Plants form mutually beneficial associations with microbes [7]. These associations play essential roles in agricultural and food safety, and contribute to the environmental equilibrium [8]. A clear distinction should be drawn between bacteria residing in the rhizosphere or phyllosphere (the aerial habitat influenced by plants) and bacteria living inside the plant, the so-called endophytes. Endophytic bacteria reside in specific tissues of the plant (such as root cortex or xylem) and develop a close association with the plant, with exchange of nutrients, enzymes (lipase, catalase, oxidase, etc.), functional agents (siderophores, biosurfactants, etc.), and also “signals” [9, 10]. Endophytes colonize their plant host tissues in which they persist without exerting the negative effects of a pathogen (disruption of respiration, photosynthesis, translocation of nutrients, transpiration, etc.). On the contrary, the presence of these endophytic bacteria in the host plant leads to beneficial effects on its health and/or growth. Plant growth-promoting bacteria promote plant health and growth via three mechanisms: phytostimulation, biofertilization, and biocontrol [11].
Phytostimulators enhance plant growth in a direct way, usually by the production of phytohormones (auxins, cytokinins, gibberellins) [12]. The production of plant hormones such as indole-3-acetic acid (IAA) is classified as “direct” promotion. The synthesis of 1-aminocyclopropane-1-carboxylate (ACC) deaminase can be included in the same group: ACC deaminase cleaves ACC, the immediate precursor of ethylene, and thereby reduces its biosynthesis; ethylene inhibits growth of roots and shoots; therefore lower levels of this plant hormone lead to plant growth promotion.
Biofertilizing strains can fix nitrogen or increase the availability of phosphorus and iron.
Biocontrol agents protect plants from infections by phytopathogens (through competition for nutrients, induced systemic resistance, production of antimicrobial secondary metabolites). Siderophore production is considered an indirect biocontrol trait; iron chelation limits the amount of trace metals available to potential plant pathogens [13].
Tomato represents one of the most important agricultural products. It can be consumed not only as raw fruit, but also transformed by industrial processes (pulped, canned, incorporated in sauce). It is known to be rich in several bioactive compounds (folate, ascorbate, polyphenols, carotenoids) [14–16].
Recent studies underline the importance of the microbial metabolism and host interaction for growth, quality, and health of edible and medicinal plants [17–19]. The bacterial community composition of tomato plants and the influence of inoculation of rhizobacteria on plant health is also a topic of increasing scientific interest [20–23]. A recent study showed that Trichoderma-enriched biofertilizer enhances production and nutritional quality of tomato and minimizes NPK fertilizer use [24]. However, the unraveling of the correlation between the in vitro detection of Plant Growth-Promoting (PGP) traits in bacterial isolates and a real contribution to the host plant health in vivo is still lacking. Moreover, in this study we not only investigated the PGP characteristics of bacteria isolated from roots of tomato cultivars already available on the market, but we also focused on new tomato hybrids of industrial interest. These new tomato hybrids, both fresh fruits and transformed products, could have a high potential for the agro-food industry because of their nutritional quality (high content of biologically active compounds) [14–16]. A screening of PGP traits of the isolates was performed to detect ACC deaminase, organic acids (OA), indole acetic acid (IAA), and siderophore (SID) production. The effects of microbial inoculation on root development were evaluated using a standardized test system with Arabidopsis thaliana grown on Vertical Agar Plates (VAPs).
Methods
Isolation of bacteria from different tomato cultivars and new tomato hybrids
Isolation of bacteria from the rhizosphere
In 2012, eleven bacterial strains, colonizing the rhizosphere of tomato plants, were collected from an experimental field situated in the Campania Region (southern Italy). The soil had originated from limestone and tuff which have been covered with a thick layer of volcanic material. High levels of oxygen and organic matter and a high availability of potassium contribute to the high fertility of the soil (see Additional file 1). Tomato roots were sampled at the late stage of maturation, placed in 50 mL tube with sterile phosphate-buffered saline (PBS; NaH2PO4·H2O, 6.33 g L−1; Na2HPO4·7H2O, 16.5 g L−1; pH 7.4), and stored at 4 °C until isolation. Samples were rinsed several times in PBS in order to remove adhering soil particles. Root cuttings were rubbed against a sterile 200-µm nylon mesh to retain roots particles and large cell debris. A sucrose solution (9 ml; 4 % w/v) was added to the filtrates obtained from 1 g of roots. Serial 10-fold dilutions of the suspensions in the same sucrose solution were prepared, and 100 µl aliquots were spread on modified Luria–Bertani (LB) plates containing 4 g L−1 of NaCl instead of 10 g L−1. After 7 days incubation at 30 °C, single colonies were selected and further purified on the basis of morphological differences. Subsequently, all strains were grown aerobically in modified Luria–Bertani (LB) medium (NaCl 0.4 % w/v).
Isolation of endophytic bacteria from roots
In 2013, 12 strains were isolated from tomato plant roots from the above-mentioned experimental field, but this time a specific protocol was applied to isolate endophytic bacteria. Fresh roots were collected in sterile 50 ml Falcon tubes with 25 ml P-buffer (per L: 6.33 g of NaH2PO4·H2O; 16.5 g of Na2HPO4·7H2O; 200 µl Tween 40). Samples were washed several times with P-buffer until the roots were free of all attached soil particles; then distilled H2O was used to remove foam (Tween 40). Sterilization was performed using a 5 % sodium hypochlorite solution for 5 min, and then the roots were rinsed 5 times in sufficient volumes of new sterile water. After rinsing, the samples were cut in smaller fragments with a sterile razor blade and crushed with sterile mortar and pestle in sterile 10 mM MgSO4-solution (1 g roots in 10 ml). Serial 10-fold dilutions of the suspensions were prepared, and 100 µl aliquots were streaked on Luria–Bertani (LB) plates. To verify the surface sterility of the roots, 100 µl of the last rinsing water was plated on LB medium.
Bacterial strain and growth media
The eleven rhizospheric strains and the twelve root endophytic strains were grown aerobically at 30 °C while shaking (120 rpm) in modified (NaCl 0.4 % w/v) and standard (NaCl 1.0 % w/v) Luria–Bertani (LB) medium, respectively. The pH of medium was 7.0–7.2. Inocula for the assays to detect plant growth-promoting (PGP) traits were prepared as described below. 10 µl of the glycerol-stocked bacterial isolates stored at −80 °C were grown in 96-well plates containing 1 mL of liquid LB medium for 48 h at 30 °C with shaking. The cultures were centrifuged at 4000 rpm for 20 min at room temperature and then the supernatant was discharged. Cell pellets were washed twice with 1 mL of 10 mM MgSO4 and re-suspended in 650 µL of MgSO4.
Genotypical analysis of the isolates
Genomic DNA of isolated bacteria was extracted using the DNeasy® 96 Blood and Tissue Kit (Qiagen). The qualitative and quantitative analysis of the extracted DNA was performed using the Nanodrop ND-1000 Spectrophotometer (Isogen Life Sciences). An aliquot (1 µl) of the extracted DNA of each bacterial strain was used in a PCR reaction without further purification to amplify a fragment of the 16S ribosomal DNA coding region. Two primers, the universal 1392R (5′-ACGGGCGGTGTGTGTRC-3′) and the bacteria-specific 27F (5′-AGAGTTTGATCMTGGCTCAG-3′), were used for the amplification. PCR products were sequenced by Macrogen Europe (Amsterdam, The Netherlands). Sequencing data were processed using the DNATools application developed by Sam Achten and Lenny Jorissen (Hasselt University). The Smith–Waterman algorithm was chosen for local sequence alignment and the consensus was obtained by comparing the quality of bases. The obtained sequences were compared with reference strains in the National Center for Biotechnology Information (NCBI) database.
Plant growth-promoting (PGP) traits of selected bacterial isolates
ACC-deaminase activity
Bacterial isolates with the ability to produce ACC deaminase were identified according to the method developed by Belimov et al. (2005) that was slightly modified [25, 26]. 250 µl of each last bacterial suspension in 10 mM MgSO4 (see Bacterial strain and growth media) was added to 1.2 mL of salts minimal medium (SMN) containing 5 mM ACC as a sole source of N. The cultures were incubated at 30 °C for 72 h with shaking (150 rpm), then harvested by centrifugation at 4000 rpm for 20 min at room temperature. The supernatant was discharged and the pellets were re-suspended in 100 µl of 0.1 M Tris–HCl buffer (pH = 8.5); cells were disrupted by the addition of 3 µl toluene followed by vigorous vortexing for 10 min. Subsequently, 10 µL of 0.5 M ACC and 100 µL of 0.1 M Tris–HCl buffer (pH = 8.5) were added and followed by shaking for 10 min. The microplates were incubated for 30 min at 30 °C with shaking (150 rpm). Then, 690 µl of 0.56 N HCl and 150 µl of 0.2 % 2, 4-dinitrophenylhydrazine reagent (in 2 N HCl) were added to the cell suspensions. The plates were kept at 30 °C for 30 min, supplemented with 1 ml of 2 N NaOH. Well plates without ACC were used as negative controls. The change of color from yellow to brown was considered as positive.
IAA production
The ability of the strains to produce indole acetic acid (IAA) was tested using the method of Gordon and Weber (1951) slightly modified [27, 28]. Briefly, 50 µl of each last bacterial suspension in 10 mM MgSO4 (see Bacterial strain and growth media) was inoculated in 96-well plates containing 1 mL 1/10 diluted 869-rich medium complemented with 50 mg mL−1 tryptophan and incubated at 30 °C for 4 days at 150 rpm in the dark. Afterward, bacterial cultures were centrifuged at 4000 rpm for 20 min, and 0.5 mL of the supernatant was added to 1 mL of Salkowski reagent (98 mL 35 % HClO4, 2 mL 0.5 M FeCl3) and gently vortexed. After 20 min, the development of a pink color was considered as positive (bacteria producing IAA).
Organic acids production
For the identification of the strains able to produce various organic acids, a protocol developed by Cunningham and Kuiack [29] was used. 50 µl of each last bacterial suspension in 10 mM MgSO4 (see Bacterial strain and growth media) was inoculated in 800 µl of Sucrose Tryptone medium (ST) containing 20 g L−1 sucrose and 5 g L−1 tryptone. ST medium was supplemented with 10 mL of trace elements solution of the following composition: NaMoO4 20 mg L−1, H3BO3 20 mg L−1, CuSO4·5H2O 20 mg L−1; FeCl3 100 mg L−1; MnCl2·4H2O 20 mg L−1; ZnCl2 280 mg L−1. After 15 min, the color change of the alizarine red pH indicator from red (pH > 6) to yellow (pH ~ 5 or below) was considered as positive.
Siderophore production
The ability of the isolates to produce siderophores was evaluated through a qualitative test in liquid 284 medium (that stimulates siderophore production) with chrome azurol S (CAS) shuttle solution [30]. Briefly, 50 µl of each last bacterial suspension in 10 mM MgSO4 (see Bacterial strain and growth media) was inoculated in 96-well plates containing 800 µl of 284 medium prepared at three different iron concentrations: without iron, with 0.25 µM, and 3 µM Fe(III) citrate. The microplates were incubated at 30 °C at 150 rpm for 5 days. After incubation, 100 µl of the blue Chromium-Azurol S (CAS) reagent was added. Plates were kept at room temperature for 4 h, and the change of color from blue to orange/yellow was considered as positive.
Evaluation of the effects of microbial inoculation on root growth
A Vertical Agar Plate (VAP) assay was performed to evaluate the effects of microbial inoculation on root development of A. thaliana seedlings [31, 32].
Growth medium and growth conditions
The basic medium was based on a 50-fold dilution of Gamborg’s B5 medium and contained KNO3 0.5 mM, MgSO4·7H2O 0.02 mM, CaCl2·6H2O 0.02 mM, NaH2PO4 0.022 mM, MnSO4·H2O 0.94 µM, (NH4)2SO4 0.02 mM as macronutrients, KI 90 nM, H3BO3 0.97 nM, ZnSO4·7H2O 0.14 nM, CuSO4·5H2O 2 nM, Na2MoO4·2H2O 20.6 nM, CoSO4·H2O 2.6 nM as micronutrients, FeCl3 3.6 µM, 2-[N-Morpholino]ethanesulfonic acid 2.56 mM MES (Sigma), pH = 5.7. Only for the plates used for germination, this basic medium was complemented with 5 g L−1 sucrose. A. thaliana seeds were surface sterilized for 1 min in 0.1 % Cl-solution, followed by five washes with sterile deionized water. The seeds were rinsed 4 times for 5 min in a larger volume of sterile deionized water and then dried in a laminar air flow. Sterilized seeds were stored for 3 days at 4 °C in the dark, then transferred with a sterile toothpick onto 12 × 12 cm transparent plates containing 40 mL of solid medium (1 % agar nr. 4 for plant tissue culture) complemented with sucrose. Plates were incubated vertically in a growth chamber at 22 °C, with a 16-h light/8-h dark regime and a light intensity of 150 µmol m−2 s−1. After 6 days, plantlets growing on the surface of the agar were transferred to fresh growth media.
Vertical Agar Plate (VAP) assay
In order to estimate the effects of bacterial inoculation on root growth, a Vertical Agar Plate (VAP) assay was performed. 5 ml of 48 h culture of each strain was centrifuged (4000 rpm, 15 min) and brought to OD660 = 0.5 using 10 mM MgSO4·7H2O, then 400 µl of this inoculum was transferred to fresh growth medium and equally distributed over the surface of the plate. For the “control,” 400 µl 10 mM MgSO4·7H2O was used.
Arabidopsis thaliana seedlings were prepared as described previously and transferred to the VAPs (five plants per plate, in triplicate). Plates were incubated vertically in a growth chamber at 22 °C, with a 16-h light/8-h dark regime and a light intensity of 150 µmol m−2 s−1.
Analysis of root growth
Root growth in the vertical agar plates was monitored daily for 8 days, and the plates were then scanned at 300 dpi (Canon CanoScan 4400F). Primary root length and total lateral root length were calculated after analysis of scanned images using the RootNav image analysis tool (University of Nottingham) [33]. 2D pictures of the roots were also created with the same software in order to graphically compare the root system architectures. Root hair visibility of scanned images was improved using the GNU Image Manipulation Program (GIMP).
Statistical analysis
All statistical analyses were performed using the 3.1.1 version of R (The R Foundation for Statistical Computing, Vienna, Austria). Since for all the data groups considered, normality and homoscedasticity assumptions were met, parametric tests were used in all analyses. Student’s t test was used for comparing two averages, while ANOVA with post hoc Tukey’s Honest Significant Difference correction for multiple correction was applied when considering more than two averages.
Results
Genotypic characterization
After the isolation, all morphologically different bacteria were purified, but only 2/3 of the strains showed to be cultivable in the subsequent steps; hence, 16S rDNA-based identification was performed on 15 strains (6 rhizospheric and 9 endophytic strains).
Among the 15 strains that were genotypically characterized, five belong to the genus Agrobacterium, and others to the genera Microbacterium (2), Bacillus (2), Rhizobium (2), Ensifer (1), Chryseobacterium (1), Pseudomonas (1), and Rhodococcus (1) (Table 1).
Screening of the plant growth-promoting (PGP) traits
All isolates were tested for their in vitro putative PGP traits. Specific tests were performed in order to detect ACC deaminase, organic acids (OA), indole acetic acid (IAA), and siderophore (SID) production. Results indicate a high percentage of strains showing PGP traits; in particular, 73 % were able to produce organic acids (OA), 89 % IAA, 83 % ACC deaminase, and 87 % siderophores (Table 2).
It is remarkable that for most root endophytes siderophore production was detected. Also among the rhizosphere strains a high percentage of siderophore producers was found; in fact, only the two strains belonging to genus Microbacterium tested negative for this feature.
For the other traits that were investigated (ACC deaminase, organic acids (OA), and indole acetic acid (IAA) production) also high percentages of positives were detected. All the isolates showed capability to exhibit at least one of the three PGP traits, and most of the strains tested positive for at least two of the three traits. Agrobacterium strains tested positive for all features.
Vertical Agar Plate (VAP) assay
After inoculation of most of the strains, the length of the main root was comparable to the control (not inoculated seedlings) or even shorter (Table 3, see Additional file 2). More specifically, concerning the rhizosphere bacteria, inoculation of the two Microbacterium sp. (numbers 2 and 8) and the two Bacillus sp. (numbers 4 and 7) strains resulted in a root length similar (±5 %) to the control; inoculation of strains 5, 6 and Chryseobacterium sp. (number 3) led to significant inhibitions (p value <0.05) of the primary root length, while only for Ensifer sp. (number 1) a trend of primary root length increase was observed (no significant difference). Concerning the root endophytes, only seedlings inoculated with strains Rhizobium sp. (number 11) and Agrobacterium sp. (number 13) showed root lengths comparable to the control (±5 %), while for the other endophytic strains shorter primary roots were found. More specifically, for strain number 10, Pseudomonas sp. (number 9), Agrobacterium sp. (number 15), and Rhizobium sp. (number 16) we registered significant differences (p value <0.05) compared to the control; for Agrobacterium sp. (number 18) a relevant but not significant difference (p = 0.053) was observed, and for the other strains (Rhodococcus sp. n. 12 and Agrobacterium sp. n. 14 and n. 17) there was a trend of lower values without being significantly different from the control.
Similar results were observed for the lateral roots; in general, the total lateral root length of inoculated plants was lower than that of the control plants (not inoculated seedlings), in some cases comparable to it (Table 4). More specifically, the total lateral root lengths of seedlings inoculated with the rhizosphere strains Ensifer sp. (number 1) and Chryseobacterium sp. (number 3) were comparable (±5 %) to the control; for strains 5 and 6 and Bacillus sp. (number 4) a decreasing trend was observed but only for strain 8 this decrease was relevant (p = 0.054), while for Bacillus sp. (number 7) and the two Microbacterium sp. (numbers 2 and 8) trends to increased total lateral root length were registered. All plants inoculated with the root endophytes showed inhibitions of the total lateral root length. Only for strain number 10, statistical analysis showed a significant difference, but relevant differences were observed for Agrobacterium sp. (numbers 15 and 18) and Rhizobium sp. (number 16); p values were, respectively, 0.069, 0.052, and 0.072.
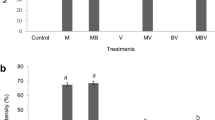
The most striking effects of bacterial inoculation on root development were strong increase in root hairs formation for most of the inoculated seedlings. Increases of root hairs formation were observed for plants inoculated with the rhizospheric strains 5, 6, Ensifer sp. (number 1) and Chryseobacterium sp. (number 3), and for all seedlings inoculated with endophytes (Fig. 1).

VAP test—increase in root hairs. Comparison of the increase in root hairs due to the inoculation with the selected strains. Ctrl control
Discussion
In this study, we report the isolation of a total of 23 strains from the roots of tomato plants. In particular, 11 strains were isolated from the rhizosphere of 4 tomato cultivars (commercial names: Plus Licopene, San Marzano giallo, Indigo Perù, and Super San Marzano), while 12 endophytes were isolated from 3 different cultivars (San Marzano, Black Tomato, and a new hybrid created by cross-pollination between these two species).
The highest efficiencies (in terms of root hair development and OA, IAA, ACC deaminase, and siderophore production) were registered for endophytic strains. In comparison to the rhizospheric ones, endophytes interact more closely with their host because of their localization in the plant tissues [34]. Such a close link leads to highly evolved mutualistic interactions, from which both plants and bacteria benefit. Endophytes represent a rich source of bioactive compounds that can positively influence plant growth through a number of different mechanisms [35]. Moreover, bacterial endophytes are competing with phytopathogens due to the fact that they colonize the same ecological niches; due to this, they often develop abilities that protect plants from infections (biocontrol agents) [36]. For these reasons, we expected to find higher efficiency in terms of PGP traits for endophytes compared to the strains isolated from the tomato rhizosphere.
This difference between the rhizosphere strains and the endophytes was also highlighted by the statistical analysis of the outcomes of the VAP test. A comparison, separately for endophytes and rhizosphere strains, between inoculated and non-inoculated plants showed statistically significant effects of inoculation on primary root length (p value = 0.03) and lateral root length (p value = 0.005) for the endophytes, but not for the rhizosphere strains (p values = 0.28 and 0.82, respectively).
This dissimilarity between rhizosphere strains and endophytes in general was once more retrieved in the form of a statistically significant difference between rhizosphere strains and endophytes in general for their effects on lateral root length (p value = 0.0001), although this was not observed for primary root length (p value = 0.8). This discrepancy between lateral and primary root length suggests a significant interaction between parameter (primary or lateral root length) and bacterial origin (endosphere or rhizosphere), which was confirmed by an ANOVA.
The obtained results also suggest a causal relationship between indole acetic acid (IAA) production capacity and the stimulation of root hair development in A. thaliana. IAA is the most studied auxin; in fact, auxin and IAA are often used as interchangeable terms [37]. It was suggested that the quantity of IAA produced by Sphingomonas sp. (11.23 ± 0. 93 μM/ml), an endophyte isolated from the leaves of Tephrosia apollinea, should be linked to the increases of surface area and root length, the loss of cell wall and the release of exudates in tomato (Solanum lycopersicum L.) plants [38]. It is known that root hair initiation and elongation are among the best characterized auxin-associated phenotypes [39–41]. High levels of IAA promote the formation of lateral roots and induce a decrease in primary root length and increases in root hair development [42]. In our work, all isolates that increased root hair development in A. thaliana in the VAP assay, also showed positive results in the IAA production test. Based on this, we may speculate that, for the investigated strains, IAA synthesis plays an essential role in the regulation of this PGP trait.
Conclusions
The improved capacity to acquire nutrients and water due to the increase of the surface area of the plant’s root system induced by inoculation of most of the isolates, makes them potentially exploitable to enhance crop productivity. However, the in vitro detection of PGP traits in natural endophytic bacteria does not implicate a general and predictable improvement of growth and health of all types of host plants in vivo [43], and therefore further investigations are needed to assess if the strains that we isolated from the different tomato cultivars effectively induce beneficial effects in vivo in tomato and also in other plant species.
Moreover, important processes of many beneficial rhizosphere-colonizing microorganisms, including some PGP traits (i.e., siderophore production), are regulated by Quorum Sensing (QS) [44]. QS is the mechanism that describes the coordinated gene expression that microbial communities show at high cell density [45]. Further analyses are in progress to investigate QS activity in the isolated strains.
References
Berg G. Plant-microbe interactions promoting plant growth and health: perspectives for controlled use of microorganisms in agriculture. Appl Microbiol Biotechnol. 2009;84(1):11–8.
Adesemoye AO, Kloepper JW. Plant-microbes interactions in enhanced fertilizer-use efficiency. Appl Microbiol Biotechnol. 2009;85(1):1–12.
Camargo JA, Alonso A. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: a global assessment. Environ Int. 2006;32(6):831–49.
Farrar K, Bryant D, Cope-Selby N. Understanding and engineering beneficial plant-microbe interactions: plant growth promotion in energy crops. Plant Biotechnol J. 2014;12(9):1193–206.
Hardoim PR, van Overbeek LS, van Elsas JD. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008;16(10):463–71.
Enya J, Shinohara H, Yoshida S, Negishi TTH, Suyama K, Tsushima S. Culturable leaf-associated bacteria on tomato plants and their potential as biological control agents. Microb Ecol. 2007;53(4):524–36.
Lugtenberg B, Kamilova F. Plant-growth-promoting rhizobacteria. Annu Rev Microbiol. 2009;63:541–56.
Mendes R, Garbeva P, Raaijmakers JM. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol Rev. 2013;37(5):634–63.
Parsek MR, Greenberg EP. Acyl-homoserine lactone quorum sensing in Gram-negative bacteria: a signaling mechanism involved in associations with higher organisms. Proc Natl Acad Sci USA. 2000;97(16):8789–93.
Hardoim PR, van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A, Döring M, Sessitsch A. The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol Rev. 2015;79(3):293–320.
Bloemberg GV, Lugtenberg BJJ. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr Opin Plant Biol. 2001;4(4):343–50.
Weyens N, van der Lelie D, Taghavi S, Vangronsveld J. Phytoremediation: plant-endophyte partnerships take the challenge Phytoremediation: plant-endophyte partnerships take the challenge. Curr Opin Biotechnol. 2009;20(2):248–54.
Jin CW, Ye YQ, Zheng SJ. An underground tale: contribution of microbial activity to plant iron acquisition via ecological processes. Ann Bot. 2014;113(1):7–18.
Tommonaro G, De Prisco R, Abbamondi GR, Nicolaus B. Bioactivity of tomato hybrid powder: antioxidant compounds and their biological activities. J Med Food. 2013;16(4):351–6.
Tommonaro G, de Prisco R, Abbamondi GR, Marzocco S, Saturnino C, Poli A, Nicolaus B. Evaluation of antioxidant properties, total phenolic content, and biological activities of new tomato hybrids of industrial interest. J Med Food. 2012;15(5):483–9.
Tommonaro G, Caporale A, De Martino L, Popolo A, De Prisco R, Nicolaus B, Abbamondi GR, Saturnino C. Antioxidant and cytotoxic activities investigation of tomato seed extracts. Nat Prod Res. 2014;28(10):764–8.
Koberl M, Schmidt R, Ramadan EM, Bauer R, Berg G. The microbiome of medicinal plants: diversity and importance for plant growth, quality and health. Front Microbiol. 2013;4:400.
Palaniyandi SA, Yang SH, Zhang L, Suh J-W. Effects of actinobacteria on plant disease suppression and growth promotion. Appl Microbiol Biotechnol. 2013;97(22):9621–36.
Garcia-Fraile P, Carro L, Robledo M, Ramirez-Bahena M-H, Flores-Felix J-D, Teresa Fernandez M, Mateos PF, Rivas R, Mariano Igual J, Martinez-Molina E, et al. Rhizobium promotes non-legumes growth and quality in several production steps: towards a biofertilization of edible raw vegetables healthy for humans. Plos One. 2012;7(5):e38122.
Botta AL, Santacecilia A, Ercole C, Cacchio P, Del Gallo M. In vitro and in vivo inoculation of four endophytic bacteria on Lycopersicon esculentum. New Biotechnol. 2013;30(6):666–74.
Yim W, Seshadri S, Kim K, Lee G, Sa T. Ethylene emission and PR protein synthesis in ACC deaminase producing Methylobacterium spp. inoculated tomato plants (Lycopersicon esculentum Mill.) challenged with Ralstonia solanacearum under greenhouse conditions. Plant Physiol Biochem. 2013;67:95–104.
Palaniyandi SA, Damodharan K, Yang SH, Suh JW. Streptomyces sp. strain PGPA39 alleviates salt stress and promotes growth of ‘Micro Tom’ tomato plants. J Appl Microbiol. 2014;117(3):766–73.
Goudjal Y, Toumatia O, Yekkour A, Sabaou N, Mathieu F, Zitouni A. Biocontrol of Rhizoctonia solani damping-off and promotion of tomato plant growth by endophytic actinomycetes isolated from native plants of Algerian Sahara. Microbiol Res. 2014;169(1):59–65.
Molla AH, Md. Haque M, Md. Haque A, Ilias GNM. Trichoderma-enriched biofertilizer enhances production and nutritional quality of tomato (Lycopersicon esculentum Mill.) and minimizes NPK fertilizer use. Agric Res 2012, 1(3):265–72.
Belimov AA, Hontzeas N, Safronova VI, Demchinskaya SV, Piluzza G, Bullitta S, Glick BR. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol Biochem. 2005;37(2):241–50.
Truyens S, Jambon I, Croes S, Janssen J, Weyens N, Mench M, Carleer R, Cuypers A, Vangronsveld J. The effect of long-term cd and ni exposure on seed endophytes of Agrostis capillaris and their potential application in phytoremediation of metal-contaminated soils. Int J Phytorem. 2014;16(7–8):643–59.
Gordon SA, Weber RP. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951;26(1):192–5.
Patten CL, Glick BR. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl Environ Microbiol. 2002;68(8):3795–801.
Cunningham JE, Kuiack C. Production of citric and oxalic acids and solubilization of calcium-phosphate by Penicillium bilaii. Appl Environ Microbiol. 1992;58(5):1451–8.
Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem. 1987;160(1):47–56.
Zhang HM, Forde BG. An Arabidopsis MADS box gene that controls nutrient-induced changes in root architecture. Science. 1998;279(5349):407–9.
Remans T, Nacry P, Pervent M, Girin T, Tillard P, Lepetit M, Gojon A. A central role for the nitrate transporter NRT2.1 in the integrated morphological and physiological responses of the root system to nitrogen limitation in Arabidopsis. Plant Physiol. 2006;140(3):909–21.
Pound MP, French AP, Atkinson JA, Wells DM, Bennett MJ, Pridmore T. RootNav: navigating images of complex root architectures. Plant Physiol. 2013;162(4):1802–14.
Weyens N, Beckers B, Schellingen K, Ceulemans R, Croes S, Janssen J, Haenen S, Witters N, Vangronsveld J. Plant-associated bacteria and their role in the success or failure of metal phytoextraction projects: first observations of a field-related experiment. Microb Biotechnol. 2013;6(3):288–99.
Ryan RP, Germaine K, Franks A, Ryan DJ, Dowling DN. Bacterial endophytes: recent developments and applications. FEMS Microbiol Lett. 2008;278(1):1–9.
Gupta P, Puniya B, Barun S, Asthana M, Kumar A. Isolation and characterization of endophytes from different plants: effects on growth of Pennisetum typhoides. Biosci, Biotechnol Res Asia. 2014;11(1):223–34.
Gamalero E, Glick BR. Mechanisms used by plant growth-promoting bacteria. In: Maheshwari DK, editor. Bacteria in agrobiology: plant nutrient management. Berlin, Heidelberg: Springer; 2011. p. 17–46.
Khan AL, Waqas M, Kang S-M, Al-Harrasi A, Hussain J, Al-Rawahi A, Al-Khiziri S, Ullah I, Ali L, Jung H-Y, et al. Bacterial endophyte Sphingomonas sp LK11 produces gibberellins and IAA and promotes tomato plant growth. J Microbiol. 2014;52(8):689–95.
Overvoorde P, Fukaki H, Beeckman T. Auxin control of root development. Cold Spring Harb Perspect Biol 2010;2(6):a001537.
Taghavi S, Garafola C, Monchy S, Newman L, Hoffman A, Weyens N, Barac T, Vangronsveld J, van der Lelie D. Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Appl Environ Microbiol. 2009;75(3):748–57.
Cho HT, Cosgrove DJ. Regulation of root hair initiation and expansin gene expression in Arabidopsis. Plant Cell. 2002;14(12):3237–53.
Vacheron J, Desbrosses G, Bouffaud M-L, Touraine B, Moenne-Loccoz Y, Muller D, Legendre L, Wisniewski-Dye F, Prigent-Combaret C. Plant growth-promoting rhizobacteria and root system functioning. Front Plant Sci 2013;4:356.
Long HH, Schmidt DD, Baldwin IT. Native bacterial endophytes promote host growth in a species-specific manner; phytohormone manipulations do not result in common growth responses. Plos One. 2008;3(7):e2702.
Ortiz-Castro R, Contreras-Cornejo HA, Macias-Rodriguez L, Lopez-Bucio J. The role of microbial signals in plant growth and development. Plant Signal Behav. 2009;4(8):701–12.
Abbamondi GR, De Rosa S, Iodice C, Tommonaro G. Cyclic dipeptides produced by marine sponge-associated bacteria as quorum sensing signals. Nat Prod Commun. 2014;9(2):229–32.
Authors’ contributions
GRA, JV, GT, and BN designed the study, participated in its coordination, and helped to draft the manuscript. GRA performed the laboratory studies, wrote the manuscript, and made the corrections. WdMR contributed to carry out the PGP bioassays. WS participated in data acquisition and statistical analysis. ST, NW, and CI contributed in experimental designing. All authors read and approved the final manuscript.
Acknowledgements
This work was supported by a dedicated grant from the Italian Ministry of Economy and Finance to the National Research Council for the project “Innovazione e Sviluppo del Mezzogiorno, Conoscenze Integrate per Sostenibilita’ ed Innovazione del Made in Italy Agroalimentare, Legge n. 191/2009” and the UHasselt Methusalem project 08M03VGRJ. Funding was also received from the COST Action (FA1103, COST-STSM-ECOST-STSM-FA1103-090914-049031) through a Short-Term Scientific Mission for GRA to Hasselt University.
The authors would like to thank Bram Beckers and Marijke Gielen (Hasselt University) for assistance with the Genotypical analysis and the Vertical Agar Plate (VAP) assay, and for comments that greatly improved the manuscript.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Additional files
40538_2015_51_MOESM1_ESM.pdf
Additional file 1. Soil characteristics analyzed in 2012 and 2013. a organic matter, b electrical conductivity of a saturated soil paste extract (ECe), c Total soil N by Kjeldahl method.
40538_2015_51_MOESM2_ESM.pdf
Additional file 2. VAP test - Comparison of the root system architectures. Vertical Agar Plate test. Comparison of the root system architectures of non-inoculated and inoculated Arabidopsis seedlings through the 2D pictures created with the RootNav image analysis tool (University of Nottingham).
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Abbamondi, G.R., Tommonaro, G., Weyens, N. et al. Plant growth-promoting effects of rhizospheric and endophytic bacteria associated with different tomato cultivars and new tomato hybrids. Chem. Biol. Technol. Agric. 3, 1 (2016). https://doi.org/10.1186/s40538-015-0051-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-015-0051-3