
Abstract
Saffron (Crocus sativus) is a medicinally important plant. The Kashmir valley (J&K, India) emblematizes one of the major and quality saffron producing areas in the world. Nonetheless, the area has been experiencing a declining trend in the production of saffron during the last decade. Poor disease management is one of the major reasons for declining saffron production in the area. Endophytes are known to offer control against many diseases of host plant. During the present study, culturable bacterial endophytes were isolated from saffron plant, identified and assessed for plant growth promoting activities. Molecular and phylogenetic analysis grouped the fifty-four bacterial isolates into eleven different taxa, viz. Bacillus licheniformis, B. subtilis, B. cereus, B. humi, B. pumilus, Paenibacillus elgii, B. safensis, Brevibacillus sp., Pseudomonas putida, Staphylococcus hominis and Enterobacter cloacae. The results were also supported with the identification based on BIOLOG system. B. licheniformis was the dominant endophyte in both leaves and corms of saffron. 81 % isolates showed lipase activity, 57 % cellulase, 48 % protease, 38 % amylase, 33 % chitinase and 29 % showed pectinase activity. 24 % of the isolates were phosphate solublizers, 86 % showed siderophore production and 80 % phytohormone production potential. The present repository of well characterized bacterial endophytes of saffron, have plant growth promoting potential which can be explored further for their respective roles in the biology of the saffron plant.
Similar content being viewed by others


Background
Saffron, commonly called as Zafran, Kesar, Kong, Kong Posh etc. and scientifically known as Crocus sativus belongs to the family Iridaceae of the order asparagales (Katariya et al. 2011). It is a small perennial plant with purple colored flowers. The flower stalk and leaves rise from corm, which is an underground part of the plant (Bhargava 2011). Apart from its uses as flavouring and colouring agent, the spice also possesses several medicinal properties (Katariya et al. 2011). Traditionally, stigmas of the plant are used for the treatment of different human disorders. Some of the reported pharmacological properties of the various solvent extracts of Crocus sativus include: anticancer (Singla and Bhat 2011; Bakshi et al. 2010); antinociceptive and anti-inflammatory (Hosseinzadeh and Younesi 2002); anticonvulsant and antidepressant (Akhondzadeh et al. 2005; Dharmananda 2005); antioxidant (Papandreou et al. 2006); hypolipidemic (Asdaq and Inamdar 2010); antityrosinase (Sariri et al. 2011); immunomodulatory (Kianbakht and Ghazavi 2011) and many other bioactivities (Bhargava 2011; Katariya et al. 2011).
Jammu and Kashmir is the only state in India where saffron is grown. In Kashmir, Pampore town is famous worldwide for its high grade saffron (Kozgar and Jabeen 2012). Unfortunately, total area under saffron cultivation in Kashmir has declined from 5707 ha in 1997 to 2667 ha in 2007 whereas the yield has observed a dip from 15.95 to 5.61 tonnes in the decade (Husaini et al. 2013). Biological stress caused by various pathogens is one of the main reasons for the declining trend in saffron yield. Some of the common fungal pathogens which infect the saffron and cause corm rot are Rhizoctonia crocorum, Phoma crocoplila, Fusarium moniliforme, Macrophomina phaseolina, Fusarium oxysporum, f. sp. solani, F. pallidoroseum, F. equiseti, Mucor sp., Penicillium sp. and Sclerotium rolfsii (Husaini et al. 2010). Among these pathogens, Fusarium oxysporum f. solani and Fusarium oxysporum f. sp. gladioli are the most devastating in Kashmir raising the corm rot incidence rate to 70–85 % leading to huge loss in terms of crop yield (Kalha et al. 2007). Fusarium corm rot of saffron caused by F.oxysporum f. sp. gladioli is also reported to cause huge yield losses in Italy as well (Primo et al. 2002).
Plants host useful bacteria as endophytes, epiphytes and rhizobacteria. Bacterial endophytes dwell host plant tissues in a symptomless manner and therefore, function as plant growth promoters (PGPs). Plants are associated with plethora of such significant microbes having biocontrol, stress resistance, phytoremediation and growth promoting potential (Ryan et al. 2008). Bacterial genera viz. Bacillus and Pseudomonas have been reported to be extensively used as biocontrol agents against root rot diseases, as well as plant growth promoters of different crop plants (Preston 2004; Dasgupta et al. 2006; Gheorghe et al. 2008).
Diversity studies are important to understand the ecological role of endophytic microbes in the host plant system. In addition to genetic and taxonomic diversity, metabolic diversity is also important to determine the exact functional role of such microbes. Carbon source utilization is widely used to determine the functional diversity of bacterial communities of a specific environment. Knowledge about the endophytic fungal diversity of saffron is limited to few reports (Raj et al. 2013) whereas the endophytic bacterial diversity of saffron is still not known. In order to fill this lacunae, the present study was undertaken to explore the bacterial inhabitants of saffron plant endosphere and determine their putative role in the biosystem of the plant.
Results and discussion
During the present study, a total of fifty-four bacterial endophytes were isolated from 252 surface sterilized leaf and corm segments of healthy saffron plants. Screening based on morphological and biochemical features broadly classified the isolates into 21 different morphospecies (Table 1; Additional file 1: Tables S1 and S2) which include fifteen phenotypically different endophytic bacterial isolates from leaves of saffron and six isolates from the corm. Isolation rate (IR) values represent percent number of endophytes isolated with respect to total number of tissue segments incubated. Isolation rate of bacterial endophytes in leaf tissue was found to be higher as compared to those in corm tissue (Table 2), which might be attributed to the variation of endophytic population with the growth stages of the plant (Zinniel et al. 2002).
16S rDNA sequences of all the isolates showed 99–100 % similarity with already available sequences in NCBI. On this basis twenty-one different morphotypes selected from fifty-four isolates were classified into eleven different bacterial species viz. Bacillus pumilus, B. licheniformis, B. subtilis, B. cereus, B. humi, Brevibacillus sp., Pseudomonas putida, Paenibacillus elgii, Bacillus. safensis, Staphylococcus. hominis and Enterobacter. cloacae (Table 1).
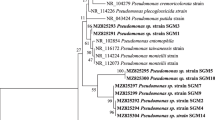
Phylogenetic analysis divided the isolates into two main groups namely Firmicutes and Proteobacteria. The optimal tree with the sum of branch length 1.17970862 is shown in Fig. 1. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) were shown above the branches (Fig. 1) (Felsenstein 1985).

Evolutionary relationship among taxa of the endophytic bacterial isolates and with the reference taxa
TS14 and TS27 isolates formed a common clade with P. putida strain Dn1.2 and E.cloacae strain SB3013, respectively, with 100 % similarity (Fig. 1). Isolates TS9, TS26, TS3 and TS15 shared the clades with B. humi, S. hominis, Brevibacillus and P. elgii, with 100 % similarity (Fig. 1). TS2, TS8, TS20, TS22 were grouped with B. pumilus and isolates TS5, TS13 with B. subtilis strains. TS4, TS7, TS11, TS16, TS17, TS18 were grouped with B. licheniformis strains (Fig. 1). Bacillus species were obtained as prominent bacterial endophytes in the present study (Tables 1, 3; Fig. 2) which is supported by earlier reports that Bacillus strains are frequently obtained as endophytes following isolation by culture dependent approach.

Isolation frequency (% values shown on bars) of bacterial endophytes from corm and leaf tissue of saffron. on X axis B.lic, Bacillus licheniformis; B.pum, B. pumilus, B.saf, B. safensis; B.bei, B. beijingensis; B.cer, B. cereus; B.sub, B. subtilis; B.hum, B. humi; Brev, Brevibacillus; P.elg, Paenibacillus elgii; P.put, Pseudomonas putida; S.hom, Staphylococcus hominis; E.clo, Enterobacter cloacae
Bacillus licheniformis was found to be the dominant bacterial endophyte of saffron with dominance value of 27.78 % followed by B. pumilus with the value 18.53 % (Table 3). Colonisation frequency of dominant endophytes was more in leaf segments than the corm segments of saffron (Table 3). However, Naik et al. (2009) have reported dominant endophytes with more colonisation frequency in roots than in leaf segments of rice. The difference in colonisation frequency of bacterial endophytes among different tissue segments in the present study can be explained on the basis of the fact that colonization frequency of endophytes is tissue dependent as influenced by structure and substrate differences in different tissues of same host plant (Fisher and Petrini 1988; Okane et al. 1998). Isolation frequency of bacterial endophytes from the corm and leaf tissues of saffron indicated presence of Bacillis licheniformis, B. pumilus and Paenibacillus sp. in both the tissues, while B. safensis, Staphylococcus hominis and Enterobacter cloacae, were restricted only to corm tissue. Enterobacter sp. and Staphylococcus species have been reported as endophytes of corn, cotton and apple (McInroy and Kloepper 1995; Phukon et al. 2013). On the other hand B. cereus, B. subtilis, B. humi, Brevibacillus sp. and Pseudomonas putida were restricted to leaf tissue of saffron. Bourgue et al. (2013) also reported Bacillus subtilis and Bacillus pumilus as endophytes from leaves of switchgrass. These differences reflect tissue specificity of individual endophytic bacterial species which may be the consequence of metabolic ability of endophytic microbes to utilize specific substrate (Carroll and Petrini 1983). Shannon-Wiener index (Table 4) indicated that the diversity of bacterial endophytes was more in leaf tissue (1.94) than in corm tissue (1.54) of saffron. The evenness index was higher in corm tissue and lower in leaf tissue of saffron due to frequent isolations of the same species and rare or no appearance of other isolates in the leaf tissue of saffron (Table 4; Fig. 2). Variation in evenness index accounts for unequal distribution of individuals (Maheswari and Rajagopal 2013).
Percent substrate richness values of different isolates based on carbon source utilization results (BIOLOG) are shown in Fig. 3. These values represent the metabolic potential of the isolates and the variation in the substrate richness values depicts the functional diversity of the isolates. Bacillus licheniformis strain TS4 was found to be metabolically most active among all the isolates with substrate richness value of 77 % (Figs. 3, 4). Amylase, cellulase, protease and lipase enzyme production potential of microbes is related to their host plant colonization ability and nutrition, whereas chitinase and phosphate solubilization, siderophore and phytohormone production potential is related to host plant growth promotion. Seventeen isolates were found to be positive for lipase activity, twelve isolates for cellulase activity, ten for protease activity, eight for amylase, seven for chitinase and six isolates showed pectinase activity (Table 1; Fig. 4). Eighteen isolates were found to be positive for siderophore production and IAA production each (Table 1; Fig. 4). As already reported by other workers, saffron plants contain pectins in high amount (Katariya et al. 2011). The pectin solublizing endophytes might be metabolizing host pectins for their nutrition and inturn may be producing some useful metabolites for the host plant. Another significant report implies that endophytes capable of degrading pectic substances are more likely to be latent pathogens of the host plant (Choi et al. 2005). So, pectin solublising bacterial isolates seem to be the latent pathogens of saffron. Five isolates were found to solublize tricalcium phosphate in Pikovskayas agar (Table 1; Fig. 4). The results are in accordance with the report of in vitro solubilisation of inorganic phosphate by endophytic bacteria from rice varieties (Duangpaeng et al. 2013). Chitinase producing isolates might be helpful in plant defense against fungal pathogens.

Percent substrate richness of the culturable endophytic bacterial isolates of saffron

Percentage of isolates showing different metabolic activities; P+ phosphate solublisation, SD+ siderophore production, Iaa+ indole acetic acid production
Isolate TS13, identified as Bacillus subtilis was found to be positive for plant growth promoting activities like siderophore and indole acetic acid production. B. subtilis has already been reported to possess saffron plant growth promotion potential. According to Eldin et al. (2008) saffron corms treated with B. subtilis suspension result in speeding of corm growth, increased stigma biomass as well as increased carotenoid pigment production. Liu et al. (2009) reported that the ginseng seedlings exhibit increased biocontrol, growth promotion and stress resistance properties when sown in soil inoculated with B. subtilis. B. licheniformis strains TS4, 7, 11,16 and 17 have shown phytohormone production potential. Isolate TS-2 a B. pumilus strain was screened as an agent with maximum enzyme production potential (Table 1). B. pumilus and B. licheniformis isolated from the rhizosphere of Alnus glutinosa have been reported to produce physiologically active gibberllins having role in plant growth promotion (Manero et al. 2001). B. licheniformis has also been reported to possess antagonistic potential against tomato and strawberry gray mould, Botrytis cinerea (Lee et al. 2006; Kim et al. 2007). Paenibacillus elgii strain TS15 was found to be positive for all the plant growth promotion activities tested in the present study. Paenibacillus isolated from the rhizosphere of Calendula sp. has been reported to possess biocontrol and plant growth promotion potential (Ryu et al. 2005; Kaki et al. 2013). B. safensis strain TS22 and B. humi strain TS9 were found to be postive for cellulase, siderophore as well as IAA production. To the best of our knowledge, Bacillus humi and B. safensis have not been reported as endophytes from any plant so far.
Identification results for most of the endophytic bacterial isolates (80 %) obtained from BIOLOG coincide with those based on 16S rDNA sequences, further confirming the identification of the isolates (Table 1). Hung and Annapurna (2004) also used BIOLOG for the identification of endophytic bacterial isolates from nodules of soyabean. Isolates identified as same species on the basis of molecular characterisation (Fig. 1) differed in their carbon source utilization profile (Fig. 3), enzyme production profile and antifungal potential (Table 1; Fig. 4). Therefore, these isolates can be considered as different strains of the, however, further studies need to be undertaken to confirm. Different Bacillus spp. reported in the present study as endophytes in saffron plant include Bacillus licheniformis, B. subtilis, B. pumilus and B. cereus, Brevibacillus sp., B. humi, Paenibacillus sp. and B. safensis (Tables 1, 3) (Figs. 1, 2). Bacillus species have been reported as potential plant growth promoters as well as biocontrol agents against different plant pathogens (Dasgupta et al. 2006; Gheorghe et al. 2008). Ryu et al. (2005) used Bacillus species viz. Bacillus pumilus, Bacillus subtilis and Paenibacillus for growth promotion of Arabidopsis sp. Predominance of Bacillus species can be attributed to their multienzyme as well as antibiotic production potential. Bacillus species have been reported to be potential antagonists of different plant pathogens due to their ability to produce a broad variety of antibiotics (Dikin et al. 2006). Ahmed et al. (2007) have also reported Bacillus sp. to suppress root rot disease caused by F. oxysporum, causal agent of saffron corm rot. Chitinolytic potential of the isolates might be responsible for their antifungal activity (Devkota et al. 2011). Results indicate that most of the isolates possess plant growth promotion traits and may have important role in biology of saffron by promoting plant growth directly, indirectly or synergistically.
Conclusion
The present study reports presence of morphologically and metabolically diverse endophytic bacterial assemblage in Crocus sativus (saffron). Endophytic bacteria have been isolated from corm (belowground) and leaf (aboveground) tissue of saffron. It has been observed that multiple plant tissue sampling, polyphasic characterisation and mathematical analysis of data are helpful in determining the endophytic biodiversity in a given plant species. However, more tissue sampling is required to uncover the complete culturable bacterial endophytic microbiome of saffron. Moreover, minimal media can also be used for the isolation of slow growing bacterial isolates, if any. The metabolic analysis of the isolates demonstrated that they produce number of hydrolytic enzymes which might play important role in nutrient cycling. The plant growth promoting properties indicate their potential as biotechnologically important cultures. Further studies are required to exactly determine the role of the endophytic isolates in the biology of saffron plant. Potential isolates from the present repository of saffron endophytes can be used for field trials to confirm the feasibility of using the same in plant growth promotion and disease management programmes. Such studies would facilitate the selection of promising cultures possessing desirable host plant growth promotion properties and can be exploited as biofertilizers in saffron fields. The use of microbial agents are becoming indispensable alternative to chemical fertilizers. So this would surely be a step forward towards sustainable and developed agriculture by minimising the use of chemicals in the environment.
Methods
Collection of the plant material
Symptomless corms were collected from saffron fields of Pampore, Kashmir (34.02°N; 74.93°E; 5164 feet high), India, in August 2013 (dormant stage of the saffron plant). Collected samples were brought to laboratory under low temperature conditions and processed for isolation of endophytes. Healthy corms were used for the direct isolation of bacterial endophytes whereas some of them were sown in the garden soil. Leaves were collected in November 2013 (during vegetative phase) and used for the isolation of bacterial endophytes.
Isolation of endophytes
The corms and leaves were surface sterilized by following the protocol of Santos et al. (2003) with slight modifications. Sterilization involved washing of selected plant parts in running water for 10 min, washing twice in distilled water for 1 min followed by treatment with 70 % (v/v) ethanol for 1 min. Subsequently, the explants were treated with 0.5 % (v/v) sodium hypochlorite for 30 s and again treated with 70 % ethanol for 1 min. Each step was followed by washing with sterile water for 2 min. Surface sterilized plant parts were later dried on sterile filter paper. After proper sterilization, leaves and corms of saffron were cut into small segments (0.5–1 cm). The segments were placed on three different media plates, viz. Nutrient agar, Potato dextrose agar and 1 % Water agar, supplemented with 50 µg/ml cycloheximide (Verma et al. 2009), for the isolation of bacterial endophytes. Effectiveness of surface sterilization was validated by following the protocol of Verma et al. (2009) with slight modifications. Surface sterilized tissue segments were stirred in sterile water and 500 µl of the suspension was inoculated on nutrient agar plates. As another control, surface sterilized explants were touched on the media plates and observed for the growth of surface bacteria, if any. All the inoculated plates were incubated at 37 °C and monitored at regular intervals for the growth. Endophytes thus observed growing on explants were isolated, purified and stored as slants at 4 °C till further use.
Screening of bacterial endophytes
Morphologically and biochemically different isolates were selected for further study. Morphological features studied included colony texture, margins, colour, elevation, growth, form, shape of bacterial cells (rod, cocci or coccobacillus) and Gram’s staining reaction (Steinbach and Shetty 2001). Biochemical characterisation was done using Bacterial identification kit (HiMedia KB003) as per user manual instructions (Results available as supplementary data).
Phylogenetic diversity analysis
Genomic DNA was isolated from the selected bacterial cultures using Himedia bacterial DNA purification kit as per manufacturer’s instructions. Universal bacterial primers were used for the amplification of 16S rRNA gene viz 16Bacf- 5′AGAGTTTGATCCTGGCTCAG 3′ (forward primer) and U1492R- 5′ GGTTACCTTGTTACGACTT 3′(reverse primer) (Doty et al. 2005). PCR program included initial denaturation at 94 °C for 4 min followed by 30 cycles of denaturation at 94 °C for 1 min; annealing at 54 °C for 50 s, extension at 72 °C for 2 min and final extension at 72 °C for 10 min.
The resulting PCR products were purified by gel extraction using DNA purification kit (Axygen) as per manufacturer’s protocol. The purified 16S rDNA amplicons were sequenced by Sanger’s method at SciGenom labs Chennai, India. The obtained sequences were then compared with public databases at the NCBI site by using the BLASTn algorithm (Altschul et al. 1997). Sequences were submitted to GenBank and accession numbers were obtained. Phylogenetic and molecular evolutionary analysis were conducted using MEGA version 6 (Tamura et al. 2013). The evolutionary history was inferred using the UPGMA method (Sneath and Sokal 1973). The evolutionary distances were computed using the Kimura 2-parameter method (Kimura 1980) and are in the units of the number of base substitutions per site.
Screening of metabolic diversity
The selected isolates were analysed for different metabolic activities including carbon source utilization ability, enzyme production potential viz lipase, amylase chitinase, cellulase, protease and pectinase as well as plant growth promoting properties like phosphate solubilisation, siderophore production, indole acetic acid production. Determination of functional diversity in terms of carbon source utilization ability of the isolates and their subsequent species level identification was carried out by using BIOLOG identification system with OMNILOG version 2.3.01; reader version Rev1.07 and GEN III database (Garland and Mills 1991; Miller and Rhoden 1991; Amaresan et al. 2012). A similarity index value of more than or equal to 0.5 was considered to indicate good species match (Wu et al. 2013). Percent substrate richness value was calculated for each bacterial isolate by determining the number of carbon compounds utilized per microplate (Miglia et al. 2007). Plate assays were done to assess the enzyme production potential of the isolates for amylase (starch agar); lipase (tributyrin agar); pectinase (MP-5 agar); protease (0.5 % gelatine agar); cellulose (1 % carboxymethylcellulose agar); chitinase (0.7 % chitin agar) (Kumar et al. 2012). Different enzyme media were inoculated with the bacterial isolates and observed after 48 h incubation at 37 °C. Phosphate solubilization potential of the isolates was assessed on Pikovskayas agar (Kumar et al. 2012). Saffron endophytes were analysed for phytohormone IAA production by following the method of Gordon and Weber (1951). Siderophore production was detected by the formation of orange halos on chrome azurol S agar (CAS) agar plates after 48 h incubation at 37 °C following the protocol of Alexander and Zuberer (1991).
On the basis of data on morphological, molecular and biochemical characterization, repetitive isolation of same bacterial endophytes was determined and isolation rate, colonization frequency, isolation frequency and dominance values were calculated.
Percent Isolation rate (IR) of bacterial endophytes was calculated as (Maheswari and Rajagopal 2013):
\(\frac{NI}{Nt} \times 100\) where NI is number of endophytic isolates obtained from plant segments and Nt is total number of segments incubated (Nt).
Percent colonization frequency (CF) of endophytes was calculated as (Maheswari and Rajagopal 2013):
\(\frac{Ncol}{Nt} \times 100\) where Ncol is number of plant segments colonised by single endophyte.
The dominance of endophytes was calculated (Verma et al. 2007) as:
\(\frac{CF}{{\sum {CF} }}\) where CF is the percentage colonization frequency of a given endophyte and ∑CF is the sum of the percentage colony frequencies of all the endophytes.
Percent isolation frequency (IF) of the endophytes was calculated as (Maheswari and Rajagopal 2013):
\(\frac{N1}{N2} \times 100\) where N1 is number of times single isolate obtained and N2 is total number of isolates obtained.
For analysis of endophytic bacterial diversity in saffron plant tissues Shanon diversity index (H) and Species eveness (E) values were determined (Verma et al. 2007).
References
Ahmed IH, Lubuschagne N, Korsten L (2007) Screening rhizobacteria for biological control of Fusarium rot and crown rot of Sorghum in Ethiopia. Biol Cont 40:97–106
Akhondzadeh S, Tahmacebi-Pour N, Noorbala AA, Amini H, Fallah-Pour H, Jamshidi AH, Khani M (2005) Crocus sativus L. in the treatment of mild to moderate depression: a double-blind, randomized and placebo-controlled trial. Phytother Res 19:148–151
Alexander DB, Zuberer DA (1991) Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol Fertile Soil 12:39–45
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Amaresan N, Jayakumar V, Kumar K, Thajuddin N (2012) Endophytic bacteria from tomato and chilli, their diversity and antagonistic potential against Ralstonia solanacearum. Arch Phytopathology Plant Protect 45:344–355
Asdaq SM, Inamdar MN (2010) Potential of Crocus sativus (saffron) and its constituent, crocin, as hypolipidemic and antioxidant in rats. Appl Biochem Biotechnol 162:358–372
Bakshi H, Sam S, Rozati R, Sultan P, Islam T, Rathore B, Lone Z, Sharma M, Triphati J, Saxena RC (2010) DNA fragmentation and cell cycle arrest: a hallmark of apoptosis induced by crocin from Kashmiri saffron in a human pancreatic cancer cell line. Asian Pac J Cancer Prev 11:675–679
Bhargava VK (2011) Medicinal uses and pharmacological properties of Crocus sativus L. (saffron). Int J Pharm Pharm Sci 3:22–26
Bourgue FG, Aliferis KA, Seguin P, Rani M, Samson R, Jabaji S (2013) Isolation and characterization of indigenous endophytic bacteria associated with leaves of switchgrass (Panicum virgatum L.) cultivars. J Appl Microbiol 114:836–853
Carroll G, Petrini O (1983) Patterns of substrate utilization by some fungal endophytes from coniferous foliage. Mycol 75:53–63
Choi YW, Hodgkiss IJ, Hyde KD (2005) Enzyme production by endophytes of Brucea javanica. J Agri Technol 1:55–66
Dasgupta SM, Khan N, Nautiyal CS (2006) Biologic control ability of plant growth–promoting Paenibacillus lentimorbus NRRL B-30488 isolated from milk. Curr Microbiol 53:502–505
Devkota HK, Maharjan BL, Baral B, Singh A, Yami KD (2011) Invitro screening of antifungal activity of rhizospheric bacteria and possible role of chitinase in antifungal activity. Nepal J Sci Tech 12:304–311
Dharmananda S (2005) ITM articles: Saffron: an anti-depressant herb. Institute for Traditional Medicine, Portland, Oregon. http://www.itmonline.org/articles/saffron/saffron.html
Dikin A, Sijam K, Kadir J, Seman IA (2006) Antagonistic bacteria against Schizophyllum commune FR in Penninsular Malaysia. Biotrop 13:111–121
Doty SL, Dosher MR, Singleton GL (2005) Identification of an endophytic Rhizobium in stems of Populus. Symbiosis 39:27–35
Duangpaeng A, Phetcharat P, Chanthapho S, Okuda N (2013) Screening of endophyte bacteria for phosphate solublisation from organic rice. In: Proceeding of the science and engineering. I-SEEC 2012, pp 61–66
Eldin MS, Elkholy S, Fernandez JA, Junge H, Cheetham R, Guardiola J, Weathers P (2008) Bacillus subtilis FZB24 affects flower quantity and quality of saffron (Crocus sativus). Planta Med 74:1316–1320
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783–791
Fisher PJ, Petrini O (1988) Tissue specificity by fungi endophytic in Ulex europaeus. Sydowia 40:46–50
Garland JL, Mills AL (1991) Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl Environ Microbiol 57:2351–2359
Gheorghe A, Jecu L, Voicu A, Popea F, Rosu A, Roseanu A (2008) Biological control of phytopathogen microorganisms with antagonist bacteria. Chem Eng Trans 14:509–516
Gordon SA, Weber RP (1951) Colorimetric estimation of indole acetic acid. Plant Physiol 26:192–195
Hosseinzadeh H, Younesi HM (2002) Antinociceptive and anti-inflammatory effects of Crocus sativus L. stigma and petal extracts in mice. BMC Pharmacol 15:2–7
Hung PQ, Annapurna K (2004) Isolation and characterization of endophytic bacteria in soybean (Glycine sp.). Omonrice 12:92–101
Husaini AM, Hassan B, Ghani MY (2010) Saffron (Crocus sativus L. Kashmirianus) cultivation in Kashmir:practices and problems. Funct Plant Sci Biotechnol 4:108–115
Husaini AM, Bhat MA, Kamili AN, Mir MA (2013) Kashmir saffron in crisis. Curr Sci 104:686–687
Jones JB, Chase AR, Harris GK (1993) Evaluation of the Biolog GN Microplate system for identification of some plant-pathogenic bacteria. Plant Dis 77:553–558
Kaki AA, Chaouche NK, Dehimat L, Milet A, Ali MY, Ongena M, Thonart P (2013) Biocontrol and plant growth promotion characterization of Bacillus species isolated from Calendula officinalis rhizosphere. Indian J Microbiol 53:447–452
Kalha CS, Gupta V, Gupta D (2007) First report of sclerotial rot of saffron caused by Sclerotium rolfsii in India. Plant Dis 91(9):1203
Katariya DC, Nilakshi N, Vijay GR, Abhyankar MM (2011) Detailed profile of Crocus sativus. Int J Pharm Bio Sci 2:530–540
Kianbakht S, Ghazavi A (2011) Immunomodulatory effects of saffron: a randomized double-blind placebo-controlled clinical trial. Phytother Res 25:1801–1805
Kim JH, Lee SH, Kim CS, Lim EK, Choi KH, Kong HG, Kim DW, Lee SW, Moon BJ (2007) Biological control of strawberry gray mold caused by Botrytis cinerea using Bacillus licheniformis N1 formulation. J Microbiol Biotechnol 17:438–444
Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16:111–120
Kozgar MI, Jabeen N (2012) Extracts of Kashmiri saffron in service to human race and present ground realities. Curr world Environ 7(2):275–280
Kumar G, Kanaujia N, Bafana A (2012) Functional and phylogenetic diversity of root-associated bacteria of Ajuga bracteosa in Kangra valley. Microbiol Res 167:220–225
Lee JP, Lee SW, Kim CS, Son JH, Song JH, Lee KY, Kim HJ, Jung SJ, Moon BJ (2006) Evaluation of formulations of Bacillus licheniformis for the biological control of tomato gray mold caused by Botrytis cinerea. Biol Control 37:329–337
Liu X, Pang J, Yang Z (2009) The biocontrol effect of Trichoderma and Bacillus subtilis SY1. J Agric Sci 1:132–136
Maheswari S, Rajagopal K (2013) Biodiversity of endophytic fungi in Kigelia pinnata during two different seasons. Curr Sci 104:515–518
Manero FJG, Solano BR, Probanza A, Mehouachi J, Tadeo FR, Talon M (2001) The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol Plantarum 111:206–211
McInroy JA, Kloepper JW (1995) Survey of indigenous bacterial endophytes from cotton and sweet corn. Plant Soil 173:337–342
Miglia KJ, McArthur ED, Redman RS, Rodriguez RJ, Zak JC, Freeman DC (2007) Genotype, soil type, and locale effects on reciprocal transplant vigor, endophyte growth, and microbial functional diversity of a narrow sagebrush hybrid zone in salt creek canyon, Utah. Am J Bot 94:425–436
Miller JM, Rhoden DL (1991) Preliminary evaluation of Biolog, a carbon source utilization method for bacterial identification. J Clin Microbiol 26:1143–1147
Naik BS, Shashikala J, Krishnamurthy YL (2009) Study on the diversity of endophytic communities from rice (Oryza sativa L.) and their antagonistic activities in vitro. Microbiol Res 164:290–296
Okane I, Nakagiri A, Ito T (1998) Endophytic fungi in leaves of Ericaceous plants. Can J Bot 76:657–663
Papandreou MA, Kanakis CD, Polissiou MG, Efthimiopoulos S, Cordopatis P, Margarity M, Lamari FN (2006) Inhibitory activity on amyloid-beta aggregation and antioxidant properties of Crocus sativus stigmas extract and its crocin constituents. J Agric Food Chem 54:8762–8768
Phukon M, Sahu P, Srinath R, Nithya A, Babu S (2013) Unusual occurrence of Staphylococcus warneri as endophyte in fresh fruits along with usual Bacillus spp. J food safety 33:102–106
Preston GH (2004) Plant perceptions of plant growth-promoting Pseudomonas. Philos Trans R Soc Lond B Biol Sci 359:907–918
Primo PD, Cappelli C, Katan T (2002) Vegetative compatibility grouping of Fusarium oxysporum f. sp. gladioli from saffron. Eur J Plant Pathol 108:869–875
Raj P, Khan SS, Modak M, Lone ZA, Rather SA, Yaqoob M (2013) Biodiversity of endophytic fungi in saffron (Crocus sativus) and antimicrobial activity of their crude extract. Indo Am J Pharm Res 3:3702–3713
Ryan RP, Germaine K, Franks A, Ryan DJ, Dowling DN (2008) Bacterial endophytes: recent developments and applications. FEMS Microbiol Lett 278:1–9
Ryu CM, Hu CH, Locy RD, Kloepper JW (2005) Study of mechanisms for plant growth promotion elicited by Rhizobacteria in Arabidopsis thaliana. Plant Soil 268:285–292
Santos RMG, Rodrigues-Fo E, Rocha WC, Teixeira MFS (2003) Endophytic fungi from Melia azedarach. World J Microbiol Biotechnol 19:767–770
Sariri R, Sabbaghzadeh R, Poumohamad F (2011) In-vitro antioxidant and antityrosinase activity of methanol extracts from Crocus sativus flowers. Pharmacol 3:1–11
Singla RK, Bhat GV (2011) Crocin: an overview. Indo Global J Pharma Sci 1:281–286
Sneath PHA, Sokal RR (1973) Numerical Taxonomy. Freeman, San Francisco
Steinbach W, Shetty A (2001) Use of the diagnostic bacteriology laboratory: a practical review for the clinician. Postgrad Med J 77:148–156
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729
Verma VC, Gond SK, Kumar A, Kharwar RN, Strobel G (2007) The endophytic mycoflora of bark, leaf, and stem tissues of Azadirachta indica A. Juss (neem) from Varanasi (India). Microb Ecol 54:119–125
Verma VC, Gond SK, Kumar A, Mishra A, Kharwar RN, Gange AC (2009) Endophytic Actinomycetes from Azadirachta indica A. Juss.: isolation, diversity and anti-microbial activity. Microb Ecol 57:749–756
Wu XQ, Yuan WM, Tian XJ, Fan B, Fang X, Ye JR, Ding XL (2013) Specific and functional diversity of endophytic bacteria from pine wood nematode Bursaphelenchus xylophilus with different virulence. Int J Biol Sci 9:34–44
Zinniel DK, Lambrecht P, Harris NB, Feng Z, Kuczmarski D, Higley P, Ishimaru CA, Arunakumari A, Barletta RG, Vidaver AK (2002) Isolation and characterization of endophytic colonizing bacteria from agronomic crops and prairie plants. Appl Environ Microbiol 68:2198–2208
Authors’ contributions
The work presented here was carried out in collaboration between all authors. SK defined the research theme and planned the experiments. TS performed most of the experimental work. TS, SK and MKD evaluated the results and wrote the manuscript. All authors read and approved the final version of the manuscript.
Acknowledgements
The authors are thankful to Department of Biotechnology, Govt of India for financial assistance and facilities. The DBT, Bioinformatics Facility of the School is also acknowledged.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Additional file
40064_2015_1435_MOESM1_ESM.pdf
Additional file 1. Morphological and biochemical characterisation of bacterial endophytes isolated from leaf and corm of saffron.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sharma, T., Kaul, S. & Dhar, M.K. Diversity of culturable bacterial endophytes of saffron in Kashmir, India. SpringerPlus 4, 661 (2015). https://doi.org/10.1186/s40064-015-1435-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40064-015-1435-3