
Abstract
In this paper, we propose a single species logistic model with feedback control and additive Allee effect in the growth of species. The basic aim of the paper is to discuss how the additive Allee effect and feedback control influence the above model’s dynamical behaviors. Firstly, the existence and stability of equilibria are discussed under three different cases, i.e., weak Allee effect, strong Allee effect, and the critical case. Secondly, we prove the occurrence of saddle-node bifurcation and transcritical bifurcation with the help of Sotomayor’s theorem. The above dynamical behaviors are richer and more complex than those in the traditional logistic model with feedback control. We find that both Allee effect and feedback control can increase the species’ extinction property. We also reveal some new bifurcation phenomena which do not exist in the single-species model with feedback control (Fan and Wang in Nonlinear Anal., Real World Appl. 11(4):2686–2697, 2010 and Lin in Adv. Differ. Equ. 2018:190, 2018).
Similar content being viewed by others

1 Introduction
For single species model, especially the logistic model, the dynamical behaviors, such as the stability, persistence, permanence, extinction, and existence of positive periodic solutions, have been extensively studied during the last decades. Also, feedback control of species in ecosystem has become a focus of intense research for the ecological balance. It is well known that when a certain component of an ecosystem changes, it inevitably causes a series of corresponding changes in other components, which in turn affects the component that originally changed. This process is called feedback control [3]. Its function is to enable the ecosystem to reach and maintain the steady state. By applying the feedback control method to the population ecosystem, a large number of scholars have extensively investigated the logistic model, and many interesting results have been obtained. For example, Gopalsamy and Weng [4] firstly introduced a feedback control variable into the delay logistic model and proposed the following ecosystem:
By constructing some suitable Lyapunov functional, the authors [4] obtained the global stability under certain condition. Subsequently, Fan and Wang [1] considered the global asymptotical stability of the following logistic equation with feedback control:
By use of a new method combined with the upper and lower solution technique, which is different from the Lyapunov function method, the authors in [1] proved that system (1.2) has a unique positive equilibrium \(E^{*}(x^{*},u^{*})\) which is globally asymptotically stable. In addition, the single species with pure delay or distributed delay, the mutualism system, the epidemic model, the discrete Lotka–Volterra competitive system, and the cooperative system with feedback control were investigated in [5–10], and [11], respectively. Very recently, in [12] the author considered a logistic model with feedback of fractional order.
On the other hand, in this vast world, the relationship of species is complex and diverse. Allee [13] pointed out that if the population is too sparse, mating between the populations will be more difficult, and then the ability to cope with predators will be weakened, which in turn will lead to lower birth rates and higher death rates. This phenomenon is called the Allee effect [14]. In recent years, the ecological model with Allee effect has been extensively studied. For example, Conway and Smoller [15], Bazykin [16] proposed the following single species model with Allee effect through incorporating a continuous growth function:
Here r denotes the intrinsic per capita growth rate of the population and K is the carrying capacity. If \(m\leq 0\), it shows the weak Allee effect, while if \(0< m< K\), it shows the strong Allee effect. As was pointed out in [17] and [18], the strong Allee effect introduces a population threshold, below which the species may be extinct, and above which it survives. In contrast, a population with weak Allee effect does not have such threshold. For more details on the Allee effect, please see [19–21].
Furthermore, Dennis [22] firstly deduced the additive Allee effect in the following equation:
Here, \(\frac{m}{x+a}\) is the term of additive Allee effect. m and a are constants which indicate the severity of the Allee effect. Moreover, the criteria of the additive Allee effect are as follows:
- (a)
If \(0< m< a\), then the Allee effect in (1.4) is weak;
- (b)
If \(m>a\), then the Allee effect in (1.4) is strong.
Consequently, an increasing number of researchers have taken into account the additive Allee effect when analyzing the rich phenomenon in ecology and biology. For example, in [23], the authors presented the Hopf bifurcation and stability analysis for a delayed logistic equation with additive Allee effect. Moreover, the predator-prey model, the diffusive or the impulsive ecological model, the fractional order Leslie–Gower model, and the hybrid bioeconomic system with the additive Allee effect were investigated in [24–31], and [32], respectively.
Recently, Lin [2] proposed a single species logistic model with Allee effect and feedback control
The author in [2] shows that the species may be extinct when the Allee effect is large. As we know, the traditional logistic system with feedback control always admits a unique positive equilibrium which is globally attractive. In other words, the Allee effect makes system (1.5) become unstable, i.e., the system could collapse under large perturbation. Also, the latest papers on population models have inspired us a lot, see for instance [33–36]. Hence, one can naturally propose the following interesting and meaningful question: How can the additive Allee effect influence the logistic system with feedback control? Does the logistic system with additive Allee effect and feedback control still admit a unique positive equilibrium? Motivated by the above, in this paper, we shall investigate the following single species logistic model with additive Allee effect and feedback control:
where x and u represent the density of the population and feedback control variable, respectively. Here a, b, c, and m are all positive constants. The biological meaning of m and a in (1.6) is the same as that in system (1.4). For system (1.6), we shall present the stability and bifurcation analysis. Moreover, the impact of the additive Allee effect and feedback control on the dynamical behaviors will be discussed in this paper.
Thus far, to the best of the authors’ knowledge, this paper is the first work which proposes and investigates the logistic model with additive Allee effect and feedback control. The detailed contribution is in two aspects. One is to discuss the existence and stability for the above system under three different Allee effect cases, i.e., \(m< a\), \(m>a\), and \(m=a\). The other is to propose the bifurcation phenomenon under the strong Allee effect case (\(m>a\)). Compared with [1] and [2], the differences are listed as follows. Firstly, the system in [1] and [2] admits at most one positive equilibrium. However, system (1.6) can admit one or two positive equilibria when the additive Allee effect constants m and a satisfy certain condition. The above shows that the additive Allee effect can have a direct impact on the species’ permanence and extinction. Secondly, the system in [1] and [2] has a trivial equilibrium which is always unstable. However, for system (1.6), in order to decide whether the trivial equilibrium is stable or not, one should compare the size of m with a. In other words, comparing with [1] and [2], system (1.6) is experiencing more complex dynamics such as the transcritical bifurcation and saddle-node bifurcation when incorporating the additive Allee effect and feedback control.
The rest of the paper is organized as follows. In Sect. 2, we discuss the existence and stability of equilibrium for system (1.6). In Sect. 3, we present certain condition under which the saddle-node bifurcation and transcritical bifurcation occur. We also carry out some numerical simulations to verify the main results. In Sect. 4, we discuss and give a conclusion to end this paper.
2 Existence and stability of equilibria
In this section, we discuss the existence and stability of the equilibria for system (1.6). By simple computation, we obtain that system (1.6) always has a trivial equilibrium \(E_{0}(0,0)\), and other interior equilibria shall be investigated as follows.
2.1 The existence of positive equilibria
The equilibria of system (1.6) are determined by the following equation:
Easily one can find that the system always admits the trivial equilibrium \(E_{0}(0,0)\). From the view of biology, we only consider the positive equilibria, and so we pay attention to the positive solution of the following equation:
Let the discriminant of equation (2.2) denoted by \(\triangle (m)\) be as follows:
and let \(m^{\ast }\) be the unique root of \(\triangle (m)=0\). Through simple calculation, we have
System always has a trivial equilibrium \(E_{0}(x_{0},u_{0})\). If \(m< m^{*}\), then \(\bigtriangleup (m)>0\). So (1.6) has two different equilibria, i.e., \(E_{1}(x_{1},u_{1})\), \(E_{2}(x_{1},u_{1})\). Here \(x_{1}=\frac{1-a-abc-\sqrt{\bigtriangleup (m)}}{2(1+bc)}\), \(u_{1}=cx_{1}\), \(x_{2}=\frac{1-a-abc+\sqrt{\bigtriangleup (m)}}{2(1+bc)}\), and \(u_{2}=cx_{2}\). However, if \(m>m^{*}\), then \(\bigtriangleup (m)<0\). So (1.6) has no equilibria. Also, if \(m=m^{*}\), \(\bigtriangleup (m)=0\), then (1.6) has a unique equilibrium \(E_{3}(x_{3},u_{3})\) where \(x_{3}=\frac{1-a-abc}{2(1+bc)}\) and \(u_{3}=cx_{3}\). Through the above analysis, we can obtain the existence of the positive equilibria in the case of weak Allee effect, strong Allee effect, and \(m=a\) as follows.
Theorem 2.1
(The case of weak Allee effect, i.e., \(m< a\))
System (1.6) has a unique positive equilibrium\(E_{2}(x_{2},u_{2})\).
Theorem 2.2
(The case of strong Allee effect, i.e., \(m>a\))
-
(i)
If \(a(1+bc)<1\) and
-
(ii)
If\(a(1+bc)\geq 1\), then system (1.6) has no positive equilibria.
Theorem 2.3
(The case of \(m=a\))
-
(i)
If\(a(1+bc)<1\), then system (1.6) has a unique positive equilibrium\(E_{2}(x_{2},u_{2})\).
-
(ii)
If\(a(1+bc)\geq 1\), then system (1.6) has no positive equilibria.
2.2 Stability of the equilibria \(E_{0}\), \(E_{1}\), \(E_{2}\), and \(E_{3}\)
Firstly, we investigate the stability of the trivial equilibrium \(E_{0}\). The Jacobian matrix of system (1.6) is calculated as follows:
So the Jacobin matrix at \(E_{0}(0,0)\) is
By direct calculation, we can obtain two eigenvalues of \(J_{E_{0}}\), i.e., \(\lambda _{1}=1-\frac{m}{a}\) and \(\lambda _{2}=-1<0\). So, if \(0< m< a\), we have \(\lambda _{1}>0\), then \(E_{0}(0,0)\) is a hyperbolic saddle. If \(m>a\), we have \(\lambda _{1}<0\), then \(E_{0}(0,0)\) is a hyperbolic stable node.
Secondly, as was pointed out in Theorem 2.2, when \(a(1+bc)<1\) and \(a< m< m^{*}\), there exist two positive equilibria \(E_{1}(x_{1},u_{1})\) and \(E_{2}(x_{2},u_{2})\). The Jacobian matrix about the equilibria \(E_{i}(x_{i},u_{i})\), \(i=1,2\), is given by
Combining with (2.1), the determinant and the trace of the Jacobian matrix are given by
After straightforward computation, we have
Therefore, \(E_{1}\) is always a saddle and \(E_{2}\) is a hyperbolic stable node.
In all, in the following, the stability of the equilibria in the case of weak Allee effect, strong Allee effect, and \(m=a\) is discussed in Theorems 2.4, 2.5, and 2.6, respectively.
Theorem 2.4
(The case of weak Allee effect, i.e., \(m< a\))
-
(1)
\(E_{0}(0,0)\)is a hyperbolic saddle.
-
(2)
\(E_{2}(x_{2},u_{2})\)is a hyperbolic stable node.
Corollary 2.1
When\(m< a\), for system (1.6), the unique positive equilibrium\(E_{2}(x_{2},u_{2})\)is globally asymptotically stable.
Proof
As was mentioned before, \(E_{2}\) is locally asymptotically stable. So we only need to prove that \(E_{2}\) is attractive. Define the Lyapunov function \(V(x,u)=x-x_{2}-x_{2}\ln \frac{x}{x_{2}}+\frac{b}{2c}(u-u_{2})^{2}\). Obviously, \(V(x,u)\) is positive definite. By use of \(1-x_{2}-\frac{m}{x_{2}+a}-bu_{2}=0\) and \(u_{2}=cx_{2}\), we can obtain the derivative of \(V(x,u)\) along the positive solution of (1.6) as follows:
We can conclude that \(\dot{V}(x,u)\leq 0\) since \(m< a\) and \(\limsup_{t\rightarrow \infty } x(t)\leq 1\). Also \(\dot{V}(x,u)=0\) if and only if \(x=x_{2}\), \(u=u_{2}\). Thus one can deduce that \(E_{2}(x_{2},u_{2})\) is globally asymptotically stable. The proof of Corollary 2.1 is finished. □
Theorem 2.5
(The case of strong Allee effect, i.e., \(m>a\))
\(E_{0}(0,0)\)is always a hyperbolic stable node. Suppose that\(a(1+bc)<1\),
- (1)
If\(a< m< m^{*}\), then\(E_{1}\)is a saddle and\(E_{2}\)is a hyperbolic stable node.
- (2)
If\(m=m^{*}\), then\(E_{3}\)is an attracting saddle-node.
Proof
In what follows, we only prove that \(E_{3}\) is a saddle-node when \(m=m^{*}\). Let \(X=x-x_{3}\), \(U=u-u_{3}\), we transform system (1.6) to the following system:
Applying the Taylor expansion of \(\frac{m}{X+x_{3}+a}\), system (2.8) can be rewritten as
where \(P(X)\) denotes the power series with term \(X^{i}\) satisfying \(i\geq 4\). The Jacobian matrix of system (2.9) evaluated at the origin, i.e., the Jacobian matrix of system (1.6) at \(E_{3}\), can be calculated as follows:
then the eigenvalues of \(J_{E_{3}}\) are \(\lambda _{1}=0\), \(\lambda _{2}=bcx_{3}-1\). Under the transformation
system (2.9) can be rewritten as
where \(P_{1}(x,u)\), \(P_{2}(x,u)\) denote the power series with term \(x^{i}u^{j}\) satisfying \(i+j\geq 4\) and
Introducing a new time variable \(\tau =(bcx_{3}-1)t\), for notational simplicity, we still retain t to denote τ and obtain
where \(e_{ij}=\frac{c_{ij}}{bcx_{3}-1}\), \(f_{ij}=\frac{d_{ij}}{bcx_{3}-1}\), \(c_{ij}\), \(d_{ij}\) (\(i+j= 2,3\)) are the same as those in (2.12). When \(m=m^{\ast }\), the coefficient of \(x^{2}\) can be simplified as \(e_{20}= \frac{(1-a-abc)(1+a+abc)^{2}}{8(1+bc)^{2}(1-bcx_{3})(x_{3}+a)^{3}}>0\). Considering the new time variable τ and using Theorem 7.1 in [37], we can conclude that the equilibrium \(E_{3}\) is an attracting saddle-node. This completes the proof. □
Remark 2.1
When \(a(1+bc)<1\), \(m>m^{\ast }\) or \(a(1+bc)\geq 1\), we can conclude that \(E_{0}(0,0)\) is globally asymptotically stable. The detailed reason is as follows. If \(a(1+bc)<1\), \(m>m^{\ast }\), or \(a(1+bc)\geq 1\), system (1.6) has a unique stable equilibrium \(E_{0}(0,0)\). And the solution of system (1.6) is bounded, so the solution cannot extend to the infinity. Also, there exists no limit cycle since system (1.6) has no interior equilibria. Thus \(E_{0}(0,0)\) is global asymptotically stable.
Theorem 2.6
(The case of \(m=a\))
-
(1)
\(E_{0}(0,0)\)is an attracting saddle-node.
-
(2)
\(E_{2}\)is locally asymptotically stable.
Proof
Here we only need to prove that \(E_{0}(0,0)\) is a saddle-node. If \(m=a\), (2.6) can be rewritten as
then the eigenvalues of \(J_{E_{0}}\) are \(\lambda _{1}=0\), \(\lambda _{2}=-1\). So \({E_{0}}\) is a non-hyperbolic equilibrium, we shall analyze the stability of \(E_{0}\) by use of Theorem 7.1 in [37]. Let
System (1.6) can be changed as follows:
Introducing a new time variable \(\tau =-t\), for notational simplicity, we still retain to denote τ and have
Applying the Taylor expansion of \(\frac{1}{x_{1}+a}\), we can obtain
Here \(R_{1}(x_{1},u_{1})\) and \(R_{2}(x_{1},u_{1})\) denote the power series with term \(x_{1}^{i}u_{1}^{j}\) satisfying \(i+j\geq 4\). Let \(u_{1}+\bar{Q}(x_{1},u_{1})=0\), we can get the implicit function \(u_{1}=\phi (x_{1})=(c-\frac{c}{a}+bc^{2})x_{1}^{2}+M(x_{1})\), and then \(\bar{P}(x_{1},u_{1})=\bar{P}(x_{1},\phi (x_{1}))=\frac{a+abc-1}{a}x_{1}^{2}+T(x_{1})\). Here \(M(x_{1})\) and \(T(x_{1})\) denote the power series with term \(x_{1}^{i}\) satisfying \(i\geq 3\) and \(i\geq 4\), respectively. Considering the new time variable τ and using Theorem 7.1 in [37], we can conclude that the equilibrium \(E_{0}\) is an attracting saddle-node. The proof is complete. □
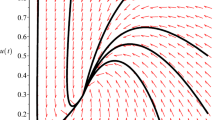
The corresponding illustrative phase portraits in Theorems 2.4–2.6 are shown in Figs. 1–3, respectively.

The phase portraits of system (1.6) when \(m< a\). Here \(E_{0}(0,0)\) is a hyperbolic saddle and \(E_{2}(x_{2},u_{2})\) is a stable node

The phase portraits of system (1.6) when \(m>a\): (a) Two distinct positive equilibria \(E_{1}\) and \(E_{2}\). \(E_{1}\) is a saddle and \(E_{2}\) is locally asymptotically stable. (b) Unique positive equilibrium \(E_{3}\) which is a saddle-node. (c), (d), (e) No positive equilibria

The phase portraits of system (1.6) when \(m=a\): (a) Two equilibria \(E_{0}\) and \(E_{2}\). \(E_{0}\) is a saddle-node and \(E_{2}\) is a stable node. (b), (c) A trivial equilibrium \(E_{0}\) which is a saddle-node
3 Bifurcation
For the case of weak Allee effect, the unique positive equilibrium is globally asymptotically stable. Thus there exist no bifurcation phenomena. In the following, we discuss the occurrence of the saddle-node bifurcation and transcritical bifurcation under the cases of strong Allee effect and the critical case when \(m=a\).
3.1 Saddle-node bifurcation
From Sect. 2, we have analyzed the existence of the equilibria \(E_{1}\), \(E_{2}\), \(E_{3}\). In other words, if \(0< m< m^{\ast }\), then system (1.6) has two positive equilibria, i.e., \(E_{1}(x_{1},u_{1})\) and \(E_{2}(x_{2},u_{2})\). If \(m=m^{\ast }\), the system has a unique equilibrium \(E_{3}(x_{3},u_{3})\). If \(m>m^{\ast }\), the system has no positive equilibria. The appearance or annihilation of equilibria may lead to the occurrence of saddle-node bifurcation which exists when \(m=m_{\mathrm{SN}}\triangleq \frac{(1+a+abc)^{2}}{4(1+bc)}\). We will prove the existence of saddle-node bifurcation as follows.
Theorem 3.1
System (1.6) undergoes the saddle-node bifurcation when\(m=m_{\mathrm{SN}}\).
Proof
In order to prove the occurrence of saddle-node bifurcation at \(m=m_{\mathrm{SN}}\), we use Sotomayor’s theorem in [38] to verify the transversality condition when \(m=m_{\mathrm{SN}}\). The Jacobian matrix at \(E_{3}\) is given by
and combining with (2.1), the determinant and the trace of the Jacobian matrix are given by
Obviously \(\operatorname{Det}[J_{E_{3}}]=0\), then \(J_{E_{3}}\) has a unique zero eigenvalue, named \(\lambda _{1}\).
Let V and W be two eigenvectors respectively corresponding to the eigenvalue \(\lambda _{1}\) for the matrices \(J(E_{3})\) and \(J(E_{3})^{T}\). After simple calculation, we have
Moreover,
Here \(F_{1}=x(1-x-\frac{m}{x+a})-bxu\) and \(F_{2}=-u+cx\). One could easily see that V and W satisfy the transversality condition
which means that the saddle-node bifurcation occurs at \(E_{3}\) when \(m=m_{\mathrm{SN}}\). Thus we complete the proof. □
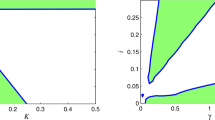
As was mentioned before, the number of interior equilibria of system (1.6) changes from zero to two when m passes from one side of \(m=m_{\mathrm{SN}}\) to the other side. For \(a= 0.5\), \(b = 1\), \(c = 0.25\), we get \(m _{\mathrm{SN}} = 0.528125\). For \(m< m_{\mathrm{SN}}\), system (1.6) has two distinct interior equilibria \(E_{1}(x_{1}, u_{1} )\) and \(E_{2}(x_{2}, u_{2})\), which collided with each other if \(m=m_{\mathrm{SN}}\), and no interior equilibria if \(m>m_{\mathrm{SN}}\). The above result is shown in Fig. 4.

The saddle-node bifurcation of system (1.6)
3.2 Transcritical bifurcation
As was mentioned before, system (1.6) has a boundary equilibrium \(E_{0}\). Moreover, we find that \(E_{0}\) coincides with \(E_{1}\) when \(m=a\). And the stability of \(E_{0}\) changes when m varies from \(m< a\) to \(m>a\). Hence, the appearance or annihilation of equilibria may lead to the occurrence of transcritical bifurcation which exists when
We obtain the occurrence of transcritical bifurcation as follows.
Theorem 3.2
System (1.6) undergoes the transcritical bifurcation when\(m=m_{\mathrm{TC}}\).
Proof
We still use Sotomayor’s theorem in [38] to verify the occurrence of the transcritical bifurcation when \(m=a\). The Jacobian matrix at \(E_{0}\) is given by
Obviously \(J_{E_{0}}\) has a unique zero eigenvalue, named \(\lambda _{1}\).
Let V and W be two eigenvectors respectively corresponding to the eigenvalue \(\lambda _{1}\) for the matrices \(J(E_{0})\) and \(J(E_{0})^{T}\), after simple calculation, we have
Moreover,
Therefore, V and W satisfy the transversality condition
which means that when \(m=a\), the transcritical bifurcation occurs at \(E_{0}\). This completes the proof of Theorem 3.2. □
The above result is shown in Fig. 5.

The transcritical bifurcation of system (1.6)
4 Conclusion
In this paper, different from [1] and [2], we have paid attention to the impact of the additive Allee effect and feedback control on the logistic model. We have obtained the dynamic behaviors under three different Allee effect cases, i.e., \(m< a\), \(m>a\), and \(m=a\).
For the case \(m< a\), besides a trivial equilibrium which is a saddle, system (1.6) also has a unique positive equilibrium which is globally asymptotically stable. The above is stated in Theorem 2.1 and Theorem 2.4. In other words, the additive Allee effect has no impact on the species if the Allee effect is weak. The obtained results coincide with those in [1] and [2].
For the case \(m>a\), the detailed results are divided into two cases. When \(a\geq \frac{1}{1+bc}\) or \(a< \frac{1}{1+bc}\), \(m>m^{*}\), system (1.6) has no positive equilibrium. If \(a<\frac{1}{1+bc}\) and \(a< m< m^{*}\), system (1.6) has a trivial equilibrium \(E_{0}\) and two positive equilibria \(E_{1}\) and \(E_{2}\). Here \(E_{0}\), \(E_{1}\) are stable nodes and \(E_{2}\) is a saddle. When \(a<\frac{1}{1+bc}\) and \(m=m^{*}\), system (1.6) has a unique positive equilibrium \(E_{3}\) which is an attracting saddle-node. All the above shows that whether the species can survive or not depends on the additive Allee effect constants m and a. The smaller these two parameters, the better the survival of the population. Furthermore, in Theorem 3.1 and Theorem 3.2, we have revealed that the system undergoes the saddle-node bifurcation around the equilibrium \(E_{3}\) and the transcritical bifurcation around \(E_{0}\).
For the critical case \(m=a\), if \(a<\frac{1}{1+bc}\), system (1.6) has a trivial equilibrium \(E_{0}\) which is a saddle-node and a unique positive equilibrium \(E_{2}\) which is globally asymptotically stable. However, if \(a\geq \frac{1}{1+bc}\), system (1.6) has no positive equilibrium, which is presented in Theorem 2.3 and Theorem 2.6. All the above shows that if \(m=a\), whether the species can survive or not depends on the feedback control constants b and c. The feedback control increases the species’ extinction probability.
To sum up, the obtained results have revealed that both the additive Allee effect and feedback control have a vital influence on the species’ permanence, extinction, and stability. In detail, the species can survive when the additive Allee effect is weak. However, when the additive Allee effect and feedback control is strong, the risk of extinction of the species will be greater. Especially, from Theorems 2.2–2.3 and Theorems 2.5–2.6, one can conclude that the feedback control strength also has a great effect on the dynamic behaviors. The above is completely different from the corresponding results in [1] and [2]. In fact, both [1] and [2] conclude that the feedback control variable cannot alter the species’ permanence or extinction. Interestingly, Theorems 2.2–2.3 and Theorems 2.5–2.6 tell us that not only the Allee effect but also feedback control can make system (1.6) become “unstable”. The main results in this paper are a good extension and supplement to those in [1] and [2].
References
Fan, Y.H., Wang, L.L.: Global asymptotical stability of a logistic model with feedback control. Nonlinear Anal., Real World Appl. 11(4), 2686–2697 (2010)
Lin, Q.F.: Stability analysis of a single species logistic model with Allee effect and feedback control. Adv. Differ. Equ. 2018(1), 190 (2018)
Wiener, N.: Cybernetics or Control and Communication in the Animal and the Machine. MIT Press, Cambridge (1948)
Gopalsamy, K., Weng, P.X.: Feedback regulation of logistic growth. Int. J. Appl. Math. Comput. Sci. 16(1), 177–192 (1993)
Fang, S.L., Jiang, M.H.: Stability and Hopf bifurcation for a regulated logistic growth model with discrete and distributed delays. Commun. Nonlinear Sci. Numer. Simul. 14, 4292–4303 (2009)
Li, Z., Han, M.A., Chen, F.D.: Almost periodic solutions of a discrete almost periodic logistic equation with delay. Appl. Math. Comput. 232(1), 743–751 (2014)
Chen, F.D., Yang, J.H., Chen, L.J.: Note on the persistent property of a feedback control system with delays. Nonlinear Anal., Real World Appl. 11(2), 1061–1066 (2010)
Chen, F.D., Yang, J.H., Chen, L.J., Xie, X.D.: On a mutualism model with feedback controls. Appl. Math. Comput. 214(2), 581–587 (2009)
Chen, L.J., Sun, J.T.: Global stability of an SI epidemic model with feedback controls. Appl. Math. Lett. 28, 53–55 (2014)
Chen, L.J., Chen, F.D.: Extinction in a discrete Lotka–Volterra competitive system with the effect of toxic substances and feedback controls. Int. J. Biomath. 8(1), 1550012 (2015)
Yang, K., Miao, Z.S., Chen, F.D., Xie, X.D.: Influence of single feedback control variable on an autonomous Holling-II type cooperative system. J. Math. Anal. Appl. 435(1), 874–888 (2016)
Hoang, M.T., Nagy, A.M.: Uniform asymptotic stability of a logistic model with feedback control of fractional order and nonstandard finite difference schemes. Chaos Solitons Fractals 123, 24–34 (2019)
Allee, W.C.: Animal Aggregations: A Study in General Sociology. University of Chicago Press, Chicago (1931)
Stephens, P.A., Sutherland, W.J., Freckleton, R.P.: What is the Allee effect? Oikos 87, 185–190 (1999)
Conway, E.D., Smoller, J.A.: Global analysis of a system of predator–prey equations. SIAM J. Appl. Math. 46(4), 630–642 (1986)
Bazykin, A.D.: Nonlinear Dynamics of Interacting Populations. World Scientific, Singapore (1998)
Wang, J.F., Shi, J.P., Wei, J.J.: Predator–prey system with strong Allee effect in prey. J. Math. Biol. 62, 291–331 (2011)
Wang, M.H., Kot, M.: Speeds of invasion in a model with strong or weak Allee effects. Math. Biosci. 171, 83–97 (2001)
Min, N., Wang, M.X.: Hopf bifurcation and steady-state bifurcation for a Leslie–Gower prey–predator model with strong Allee effect in prey. Discrete Contin. Dyn. Syst., Ser. A 39(2), 1071–1099 (2019)
Yu, T.T., Tian, Y., Guo, H.J., Song, X.Y.: Dynamical analysis of an integrated pest management predator–prey model with weak Allee effect. J. Biol. Dyn. 13(1), 218–244 (2019)
Zhang, J.M., Zhang, L.J., Bai, Y.Z.: Stability and bifurcation analysis on a predator–prey system with the weak Allee effect. Mathematics 7(5), 432 (2019)
Dennis, B.: Allee effects: population growth, critical density, and the chance of extinction. Nat. Resour. Model. 3, 481–538 (1989)
Elabbasy, E.M., Elmorsi Waleed, A.I.: Hopf bifurcation and stability analysis for a delayed logistic equation with additive Allee effect. Comput. Ecol. Softw. 5(2), 175–186 (2015)
Aguirre, P., González-Olivares, E., Sáez, E.: Two limit cycles in a Leslie–Gower predator–prey model with additive Allee effect. Nonlinear Anal., Real World Appl. 10, 1401–1416 (2009)
Pal, P.J., Tapan, S.H.: Dynamical complexity of a ratio-dependent predator–prey model with strong additive Allee effect. Appl. Math. 146, 287–298 (2015)
Jiang, J., Song, Y.L., Yu, P.: Delay-induced triple-zero bifurcation in a delayed Leslie-type predator–prey model with additive Allee effect. Int. J. Bifurc. Chaos 26(7), 1650117 (2016)
Liu, Y.W., Liu, Z.E., Wang, R.Q.: Bogdanov–Takens bifurcation with codimension three of a predator–prey system suffering the additive Allee effect. Int. J. Biomath. 10(3), 1750044 (2017)
Cai, Y.L., Zhao, C.D., Wang, W.M., Wang, J.F.: Dynamics of a Leslie–Gower predator–prey model with additive Allee effect. Appl. Math. Model. 39(7), 2092–2106 (2015)
Yang, L., Zhong, S.M.: Dynamics of an impulsive diffusive ecological model with distributed delay and additive Allee effect. J. Appl. Math. Comput. 48(1–2), 1–23 (2015)
Yang, L., Zhong, S.M.: Dynamics of a diffusive predator–prey model with modified Leslie–Gower schemes and additive Allee effect. Comput. Appl. Math. 34(2), 671–690 (2015)
Suryanto, A., Darti, I., Anam, S.: Stability analysis of a fractional order modified Leslie–Gower model with additive Allee effect. Int. J. Math. Math. Sci. 2017, Article ID 8273430 (2017)
Liu, C., Wang, L.P., Lu, N., Yu, L.F.: Modelling and bifurcation analysis in a hybrid bioeconomic system with gestation delay and additive Allee effect. Adv. Differ. Equ. 2018(1), 278 (2018)
Cai, Y.L., Gui, Z.J., Zhang, X.B., Shi, H.B., Wang, W.M.: Bifurcations and pattern formation in a predator–prey model. Int. J. Bifurc. Chaos 28(11), 1850140 (2018)
Zhang, H.S., Cai, Y.L., Fu, S.M., Wang, W.M.: Impact of the fear effect in a prey–predator model incorporating a prey refuge. Appl. Math. Comput. 356, 328–337 (2019)
Wang, J., Cai, Y.L., Fu, S.M., Wang, W.M.: The effect of the fear factor on the dynamics of a predator–prey model incorporating the prey refuge. Chaos, Interdiscip. J. Nonlinear Sci. 29(8), 083109 (2019)
Yang, B., Cai, Y.L., Wang, K., Wang, W.M.: Optimal harvesting policy of logistic population model in a randomly fluctuating environment. Phys. A, Stat. Mech. Appl. 526, 120817 (2019)
Zhang, Z.F., Ding, T.R., Huang, W.Z., Dong, Z.X.: Qualitative Theory of Differential Equation. Science Press, Beijing (1992) (in Chinese): English edition: Transl. Math. Monogr. Vol. 101 (Am. Math. Soc., Providence)
Perko, L.: Differential Equations and Dynamical Systems, 3rd edn. Texts in Applied Mathematics, vol. 7. Springer, New York (2001)
Acknowledgements
Not applicable.
Availability of data and materials
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
Funding
This work was supported by the National Natural Science Foundation of China under Grant 11601085, the Natural Science Foundation of Fujian Province under Grant 2017J01400 and 2018J01664, the Scientific Research Foundation of Fuzhou University under Grant GXRC-17026 and GXRC-18047.
Author information
Authors and Affiliations
Contributions
All authors contributed equally to the writing of this paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that there is no conflict of interests.
Consent for publication
Not applicable.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lv, Y., Chen, L. & Chen, F. Stability and bifurcation in a single species logistic model with additive Allee effect and feedback control. Adv Differ Equ 2020, 129 (2020). https://doi.org/10.1186/s13662-020-02586-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-020-02586-0