
Abstract
Introduction
Laccases are green biocatalysts that possess attractive for the treatment of resistant environmental pollutants and dye effluents.
Purpose
To exploit the laccase of Anoxybacillus ayderensis SK3-4 that possesses dye decolorization ability at room and higher temperature, we characterized the enzyme in considerable detail and investigated its ability to decolorize different dyes.
Results
A bacterial laccase gene designed as LacAn from Anoxybacillus ayderensis SK3-4 of hot springs was cloned and expressed in Escherichia coli. LacAn is a monomeric protein with a molecular weight of 29.8 kDa. The optimum pH and temperature for syringaldazine oxidation were 7.0 and 75 °C, respectively. LacAn was stable at pH values ranging from 6.5 to 8.5 above 65 °C. The enzyme activity was significantly enhanced by Cu2+ and Mg2+ but inhibited by Zn2+ and Fe2+. Furthermore, LacAn showed high decolorization capability toward five dyes (direct blue 6, acid black 1, direct green 6, direct black 19, and acid blue 93) in the absence of redox mediators. It also demonstrated a wide temperature range, and it can retain its high decolorization ability even at high temperatures.
Conclusions
These properties including better enzymatic properties and efficiency to decolorize dyes demonstrate that the bacterial laccase LacAn has potentials for further industrial applications.
Similar content being viewed by others


Explore related subjects
Find the latest articles, discoveries, and news in related topics.Introduction
Laccases (benzenediol: oxygen oxidoreductases, EC 1.10.3.2) are a family of multicopper oxidases widely distributed in the bacteria, fungal, insect, plant kingdoms, and particularly in basidiomycetes polyphenols. They catalyze the one-electron oxidation of a wide range of aromatic compounds including phenols and amines by coupling to the reduction of oxygen to water (Hoegger et al., 2006; Baldrian, 2006). Laccases have been considered as green and environmental-friendly biological catalysts and have been the focus of an increasing contemporary investigation due to their diverse potential biotechnological applications, such as dye decolorization, biopulping, biobleaching, xenobiotic degradation, food processing, biopolymer modification, ethanol production, biosensor development, drug synthesis, and organic synthesis (Telke et al., 2011; Kudanga and Le Rose-Hill, 2014; Mate and Alcalde, 2016).
In the early stage of laccase application, the biological roles of fungal and plant laccases have been thoroughly studied and shown to be related to the degradation and synthesis of lignin (Leonowicz et al., 2001). However, recent studies have discovered that laccases are widespread in bacteria, such as Bacillus tequilensis (Sonica et al., 2014), Caldalkalibacillus thermarum (Sunil et al., 2018), Streptomyces griseorubens (Feng et al., 2015), and Agaricus bisporus (Othman et al., 2018), and their application is common in the industrial field. Industrial processes usually include harsh conditions, such as high temperature, acidic or alkaline pH, and high salt and detergents; thus, some laccases that are resistant to these harsh conditions are preferable (Hilden et al., 2009; Gunne and Urlacher, 2012). Of the reported bacterial laccases, several possess distinctive properties, including excellent activity and stability under alkaline conditions. For example, the Tth laccase from Thermus thermophilus exhibits extreme stability against heat with a half-life of more than 14 h at 80 °C (Miyazaki, 2005). The Ssl1 laccase from Streptomyces sviceus is highly alkali stable and resistant to detergents and organic solvents (Gunne and Urlacher, 2012). In addition, the SN4 laccase from Bacillus tequilensis is thermo-alkali stable and metal tolerant (Sonica et al., 2014). Bioinformatics analysis has demonstrated the high diversity of laccase or laccase-like enzymes in bacteria (Ausec et al., 2011), but bacterial laccase-like enzymes have yet to be exploited as promising laccase resources.
In this study, a putative laccase-like gene (designated lacAn) from Anoxybacillus ayderensis SK3-4 was cloned and heterologously expressed in Escherichia coli BL21 (DE 3). Anoxybacillus ayderensis SK3-4 was isolated from the Sungai Klah (SK) and Dusun Tua (DT) hot springs in Malaysia (Kahar et al., 2013). To exploit the laccase of Anoxybacillus ayderensis SK3-4 that possesses dye decolorization ability at room and higher temperature, we characterized the enzyme in considerable detail and investigated its ability to decolorize different dyes. LacAn possessed great enzymatic properties in terms of biochemical characteristics. The potential of the enzyme in the decolorization of industrial dyes in the absence of mediators was also evaluated.
Materials and methods
Chemicals
2,6-Dimethoxyphenol and 2,2′-azinobis (3-ethylben-zthiazoline-6-sulfonate) (ABTS), syringaldazine (SGZ), and 4-fluoro-2-methylphenol were from Sigma. Restriction enzymes and DNA ligase were from New England Biolabs (MA, USA). Agarose gel DNA extraction kit was from Qiagen, Hilden, Germany. Taq DNA polymerase and isopropyl-β-D-thiogalactoside (IPTG) were purchased from TaKaRa (Dalian, China). Direct blue 6 (DB6), acid black 1 (AB1), direct green 6 (DG6), direct black 19 (DB19), and acid blue 93 (AB93) were from Sigma-Aldrich (St. Louis, MO, USA). All other chemicals and reagents were of analytical grade.
Microorganisms and maintenance
E. coli BL21(DE3) was purchased from TransGen (Beijing, China) and was used as the host strain for plasmid propagation and protein expression. The E. coli strain was routinely grown in Luria-Bertani medium.
Cloning, expression, purification, and verification of identity
A series of potential laccase sequences were obtained on NCBI through bioinformatics technology (the access no. EPZ38526.1 from GeneBank). The target laccase-like gene (designated lacAn) was selected from Anoxybacillus sp. and optimized according to E. coli preference. Then the sequence was sent to General Biosystems (Anhui, China) for synthesis (Fig. S1). Finally, the synthesized sequence is inserted into the vector pUC18.
The open reading frame (a 804-bp fragment) of LacAn was amplified using the primer pair of GATATACATATGAACGATATATTTCGCCAAG (Nde I site italicized) and GTGGTGCTCGAGCCTCCAGCCAATAAGCGC (Xho I site italicized) based on the optimized gene sequence (pUC18-LacAn). Construction of plasmid pET22b-LacAn was performed according to the methods of Fang et al. (Fang et al., 2011). E. coli BL21(DE3) cells carrying pET22b-LacAn were grown at 37 °C in 200 mL of LB medium containing 100 μg/mL ampicillin. The cultivation temperature was reduced to 16 °C, and IPTG at a final concentration of 0.2 mM was added into the culture to induce enzyme expression when the OD600 of culture medium reached 0.6. After an additional incubation for 16 h, the cells were collected by centrifugation. The pellets were resuspended in cold 20 mM Tris-HCl buffer (pH 7.9) containing 500 mM NaCl and 5 mM imidazole, disrupted by sonication, and then centrifuged at 30,000×g for 30 min. The supernatant was applied to Ni-NTA (Novagen, Darmstadt, Germany) affinity chromatography to purify the recombinant LacAn. The purified protein was stored at 4 °C.
The molecular mass of the denatured protein was estimated by sodium dodecyl sulfate (SDS) polyacrylamide gel electrophoresis (PAGE). Proteins were stained with Coomassie brilliant blue R-250. Activity staining of LacAn was performed in 50 mM Na2HPO4-KH2PO4 buffer (pH 7.0) containing different substrates and 10 μM CuSO4. Protein concentration was determined using the BAC method (Bio-Rad, USA).
Sequence analysis of lacAn
The ORF of lacAn was determined using the ORF Finder provided by the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). Similar sequence searching was performed using BlastP at NCBI. The module structure of the enzyme was analyzed with simple modular architecture research tool SMART (http://smart.embl-heidelberg.de/). Multiple sequence alignment of lacAn with other related laccase sequences was performed using Clustal X 2.0.
Enzyme assay
The assay mixture consisted of 10 μL of appropriately diluted LacAn stock and 990 μL of 50 mM Na2HPO4-KH2PO4 buffer (pH 7.0) containing 100 μM SGZ (65,000 M−1 cm−1) and 10 μM CuSO4. The reaction was initiated by adding SGZ and enzyme into the solution. After incubation at 45 °C for 5 min, the mixture was transferred into ice-water bath for 30 s to stop the reaction and the absorbance was measured at 525 nm using a UV-visible spectrophotometer (Shimadzu uv-vis 1700, Japan). Alternative substrates for the measurement of laccase activity were 2 mM 2,6-DMP. Reactions with heat-treated LacAn were used as control. One activity unit (U) was defined as the amount of LacAn for oxidizing 1 μmol of substrate per minute. Protein concentration of LacAn was determined at 595 nm using the Modified Bradford Protein Assay Kit (Sangon, China) as bovine serum albumin as the standard.
Characterization of LacAn
The effect of pH on laccase activity was examined in the pH range of 4.5–9.0. Sodium citrate buffer (pH 4.5–5.5), Na2HPO4-KH2PO4 buffer (pH 5.5 to 8.0), and Tris-HCl buffer (pH 8.0–9.0) were used at a final concentration of 50 mM. The enzyme stability against pH was determined by measuring the residual activities of LacAn after incubation at 4 °C for 1 h in the aforementioned buffers. The effect of temperature on the activity was measured by incubating LacAn at pH 7.5 and a temperature range of 40–80 °C. Thermostability was determined by incubating LacAn at various temperatures (15–55 °C) at pH 7.5 for 15 min. The values and standard deviations are calculated based on three independent experiments.
The effects of 0.1 mM Mg2+, Co2+, Zn2+, Ca2+, Fe2+, EDTA 5% dimethyl sulfoxide (DMSO), 0.5 mM SDS, and 5% ethanol on LacAn activity were investigated by incubating LacAn with each effector for 15 min at 4 °C prior to substrate SGZ addition. The laccase assays were carried out under the aforementioned conditions. Control was carried out under the conditions with Na2SO4, KCl, or without NaCl in the normal manner.
Decolorization of textile azo dyes
The LacAn was used to oxidize 5 synthetic dyes of different classes which were shown in Table 1. The chemical structures of these dyes were shown in an additional figure (Fig. S2). Initial solutions (10 mM) of these dyes were prepared in DMSO and diluted to the required concentration and then used for the reaction. Initially, the reaction was mixture 1 mL contained 50 mM citrate-phosphate buffer (pH 7.5), crude enzyme (200 U/L), and dye solution (100 μM).
To evaluate the effect of the conditions on dye decolorization, the pH (5.5–8.5), temperature (55–80 °C), enzyme amount (50–250 U/L), and time (20–140 min) were optimized in turn. The reactions were incubated for 2 h, and the boiled LacAn as the control. The decolorization of these dyes by LacAn was monitored by the decrease in absorbance at the wavelength of each dye. The decolorization ratio was calculated according to the following equation:
Decolorization ratio (%) = (A − A0)/A0 × 100%
A0 and A represented the initial and final absorbance of the dye, respectively.
Results and discussion
Analysis of the LacAn sequence
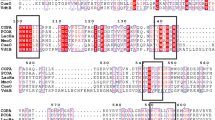
The putative ORF of lacAn consists of 804 nucleotides that encode a protein containing 267 amino acid residues. Putative conserved domains were detected according to the Pfam database (Pfam PF02578), demonstrating that LacAn belongs to the Cu-oxidase_4 superfamily (Beloqui et al., 2006). The relationship between lacAn and multiple selected laccase genes is shown in Fig. 1. Among the 12 identified copper sites in RL5, all sites were conserved in lacAn, which proved that LacAn is a bacterial laccase (Beloqui et al., 2006).

Multiple sequence alignment of LacAn with selected LacAn-related proteins. Amino acid sequences were retrieved from NCBI or the UniProt database. LacAn, a copper oxidase in this study (GeneBank accession no. EPZ38526.1); yfiH, a laccase from E. coli (UniProt accession no. P33644); 1t8h (UniProt accession no. P84138), Q5HQ05.1, Q5L0W5 were uncharacterized proteins from Bacillus stearothermophilus, Staphylococcus epidermidis RP62A, and Geobacillus kaustophilus; RL5, a laccase from the bovine rumen metagenome (NCBI accession no. AM269758). The amino acid residues binding to the three copper sites in RL5 are indicated with Δ
Purification of LacAn
After preliminary optimization of expression and purification, a protein with laccase activity and molecular mass of about 29.8 kDa (Fig. 2), named LacAn, was obtained. Under the corresponding optimum condition, the specificity of LacAn was 3.2 U mg−1 with substrate SGZ. At present, the molecular mass of most fungi and bacterial laccases range from 50 to 100 kDa (Brijwani et al., 2010; Sharma et al., 2007). However, the molecular mass of LacAn is only 29.8 kDa. Similar conditions were also found in the laccase protein SN4LAC of B. tequilensis (Sonica et al., 2014). Sondhi et al. suggested that this difference in molecular mass makes SN4LAC an interesting protein for studying the structure-function relationship of laccases (Sonica et al., 2014).

SDS-PAGE of overexpressed and Ni2+-NTA purified LacAn. Lane M, standard molecular weight marker; lane control, BL21(DE3) without pET22b-lacAn; lane sonicate, sonication product precipitate of BL21(DE3)-pET22b-lacAn; lane LacAn, purified LacAn
Characterization of LacAn
The optimum temperature of the purified LacAn was 75 °C (Fig. 3a). LacAn displayed more than 60% of the maximal activity with 180 min from 25 to 45 °C. In addition, the enzyme was stable at relatively high temperatures, with a half-life of 155 min at 65 °C when syringaldazine was used as a substrate (Fig. 3b). Obviously, LacAn, similar to many discovered bacterial laccases, showed great thermostability even at temperatures above 60 °C (Table S1). In addition, environmental factors also affected its thermostability. Anoxybacillus ayderensis SK3-4 originated from hot springs, which have a high temperature environment, explaining the stable thermostability of LacAn.

Effects of pH and temperature on the activity and stability of LacAn. a Optimum temperature of LacAn for enzyme activity was measured in 50 mM Na2HPO4-KH2PO4 at pH 7.0 with syringaldazine as a substrate. b Thermostability was investigated at 25 °C (triangle), 45 °C (circle), and 65 °C (square) by determining the enzyme activity under the same conditions as above. c Optimum pH of LacAn for enzyme activity was measured in 50 mM Na2HPO4-KH2PO4 at 75 °C with syringaldazine as a substrate. d pH stability was investigated at pH 8.5 (triangle), pH 7.5 (circle), and pH 6.5 (square) by determining the enzyme activity under the same conditions as above. Error bars represent standard deviations
Conversely, LacAn displayed the maximum activity at pH 7.0 (Fig. 3c). It was stable at pH values ranging from 6.5 to 8.5 at 75 °C and was the most stable at pH 7.5, retaining more than 80% of the original activity after incubation at 75 °C for 180 min while catalyzing syringaldazine (Fig. 3d). In general, bacterial laccases are functional in an alkaline environment and are active at pH 7.0–8.5, while fungal laccases are partially active in an acidic environment (Claus, 2003; Brander et al., 2014). LacAn is an extracellular thermo-alkali-stable laccase. The alkaline activity of LacAn is similar to those of other bacterial laccases in previous studies (Table S1). Thus, it may be further applied in industrial and biotechnological field. Additional, Km and Vmax of LacAn for the substrate SGZ were 14.2 μM and 10.6 μmol min−1 mg−1, respectively.
Metal ions bind to laccases and alter their stability. Cu2+ was important for LacAn activity because no activity was detected for the purified as isolated protein (Fig. 4a). The stimulation of laccase activity by Cu2+ observed in the study occurred probably due to the filling of type I or II copper binding sites with copper ions, highlighting the importance of Cu2+ ion in laccase function (Nagai et al., 2002; Kaushik and Thakur, 2013; Sonica et al., 2014). Zn2+ and Fe2+ decreased LacAn activity to 90% compared with that without metal ions. Inhibition of LacAn in the presence of Zn2+ was in accordance with the results from previously characterized fungal laccases (Murugesan et al., 2006; Sunil et al., 2018). The inhibition effect of Fe2+ may be due to its interaction with the electron transport system of laccase. The blockage of the access of the substrate or the transfer of electron at the T1 site results in inhibition in laccase activity (Murugesan et al., 2009). LacAn was almost stable in the presence of other metal ions. For example, Mg2+ increased the activity of LacAn to 133%, and Co2 + and Ca2+ increased the enzyme activity to 122% and 106%, respectively, similar to the results reported for B. tequilensis, may be due to their interaction with electron transport system of laccase (Sonica et al., 2014, Murugesan et al., 2006). The stability of LacAn in the presence of some metal ions makes LacAn suitable for applications where metal ions are present in high concentrations, such as in the pulp and paper industry and in wastewater containing heavy metals (Shraddha et al., 2011).

Effects of Cu2+, metal ions, inhibitors, and organic solvent on the enzyme activity of LacAn. a Optimum Cu2+ concentration of LacAn for enzyme activity was measured in 50 mM Na2HPO4-KH2PO4 at 75 °C and pH 7.0 with syringaldazine as a substrate. b Effects of metal ions, inhibitors, and organic solvent on the enzyme activity of LacAn. The enzyme activity was measured in 50 mM Na2HPO4-KH2PO4 at 75 °C and pH 7.0 with syringaldazine as a substrate, supplemented with 0.1 mM CuSO4. Error bars represent standard deviations
In addition, potential inhibitors exerted various effects on LacAn activity. Ionic surfactants reportedly inhibit laccase activity (Robles et al., 2002; Zhang et al., 2013). Under the above reaction conditions, 0.5 mM SDS significantly inhibited the enzyme activity, and only 13% of the enzyme activity was retained. This result may be due to binding of the ionic surfactant [below Critical Micelle Concentration (CMC)] to the enzyme which may cause the alterations in its enzymatic and physical characteristics (Robles et al., 2002; Sonica et al., 2014; Zhang et al., 2013). The effect of 10 mM EDTA was slightly weaker than SDS with 22% of the activity was retained, due to the deprivation of the Cu2+ ions present at type 1 copper center and inhibit the enzyme activity (Kaushik and Thakur, 2013). When DMSO and ethanol organic solvents were added, about 50% of the enzyme activity was retained (Fig. 5). However, as a comparison, ionic surfactants reportedly stimulate the activity of laccase from Azospirillum lipoferum and B. tequilensis (Diamantidis et al., 2000; Sonica et al., 2014).

Effect of pH (a), temperature (b), concentration of LacAn (c), and time (d) on the decolorization rate of LacAn oxidation dyes. The values and standard deviations were calculated based on three independent experiments
Decolorization of azo dyes by LacAn
The ability of LacAn to oxidize dyes was tested by single-factor optimization. When the five dyes were oxidized, the decolorization rates remained high when the pH was at 8.5 (Fig. 5a). The highest decolorization rates were achieved at pH 7.5, which was then chosen as the optimum pH and suitable for industrial applications. The temperatures were set at the range of 55–80 °C. Results showed that LacAn had good catalytic properties at high temperature and remained active within a wide temperature range. A high decolorization rate was achieved even at 80 °C (Fig. 5b). In addition, all five dyes were efficiently decolorized and the highest decolorization rates were observed even when 200 U/L LacAn was added (Fig. 5c). The decolorization rates of the dyes increased when the reaction time was extended. Finally, the highest decolorization rates of the five dyes showed that direct blue 6 and acid black 1 reached 33.15% ± 2.69% and 31.08% ± 1.31%, respectively, at 100 min of reaction. At 60 min of reaction, direct green 6 and direct black 19 reached 99.64% ± 1.13% and 51.34% ± 1.66%, respectively. Acid blue 93 reached 34.45% ± 1.52% at 80 min (Fig. 5d, Table 2).
Laccase can oxidize a variety of substrates for industrial applications, especially for the oxidation of dyes in industrial wastewater treatment, which has important environmental significance (Verma and Madamwar, 2003; Rai et al., 2005). LacAn is a bacterial laccase with high catalytic activity in alkaline pH. In this study, LacAn was able to oxidize five dyes under neutral to alkaline conditions and still had higher decolorization rates even at pH 8.5 (Fig. 5a). This result indicates that the bacterial laccase has better decolorization characteristics than fungal laccases under neutral alkaline conditions.
Most of the reported decolorization reaction temperatures of bacterial laccase are between 30 and 45 °C. With the increase in temperature, the activity of laccase decreased and the decolorization ability decreased, which limited the application of laccase decolorization (Kalmea et al., 2009; Molina-Guijarro et al., 2009; Pereira et al., 2009; Liu et al., 2011; Lu et al., 2012). LacAn can still decolorize dyes effectively at 80 °C, especially direct green 6 with a high decolorization rate of 99% (Fig. 5b). It enables LacAn to oxidize dyes at low and high temperatures and also shortens the reaction time at high temperatures, revealing its advantage of high temperature decolorization. LacAn has the advantage of high temperature application even in laccase derived from thermophilic bacteria. Compared with a laccase named TthLAC from Thermus thermophilus, when the azo dyes were oxidized by the TthLAC, the efficient decolorization was at temperature of 60–65 °C. It made the LacAn more widely used (Kumari et al., 2018).
The spatial structure of the dye macromolecule hinders its binding with the active center of the enzyme. Most laccases have low redox potential. Mediators such as ABTS and syringaldehyde must be added to oxidize dye substrates to complete the catalytic reaction and increase the decolorization efficiency (Guan et al., 2012; Yang et al., 2018). For example, the laccase LaclK from Kurthia huakuii was used to oxidize azo dyes (Guo et al., 2016). Only when ABTS was added as a mediator, LaclK showed obvious decolorization ability (Guo et al., 2016). A laccase derived from Bacillus subtilis cjp3 was used for dye oxidative decolorization, and results showed that reactive blue 19, reactive black 5, and indigo carmine can be effectively decolored by purified laccase with ABTS as a mediator. More than 86% of the dyes tested were removed at 4 h (Qiao et al., 2017). For the above laccases, without a mediator, there was no decolorization ability. But LacAn had an effective decolorization ability when the mediator was absent. Although the mediator could improve the decolorization efficiency, it would cause more pollution and increase the cost of decolorization. Due to the lack of mediators in decolorization, LacAn will have more important application value.
The LacAn used in this study still had a good decolorization effect on the selected dyes within a short reaction time even without any mediators (Fig. 5, Table 2). This result may be due to the better catalytic activity and stability of LacAn. Thus, LacAn has a wide application prospect in industrial wastewater treatment.
Conclusion
A bacterial laccase (LacAn) from Anoxybacillus ayderensis SK3-4 was cloned and successfully expressed in E. coli. The purified enzyme is smaller than other known laccases. In terms of biochemical characteristics, the optimum pH and temperature for SGZ oxidation were 7.0 and 75 °C, respectively. LacAn was stable at pH values ranging from 6.5 to 8.5 above 65 °C. Moreover, LacAn showed desirable stability at high temperatures and alkaline pH, and dye decolorization ability. These properties render LacAn a prospect for further industrial applications.
References
Ausec L, Zakrzewski M, Goesmann A, Schluter A, Mandic-Mulec I (2011) Bioinformatic analysis reveals high diversity of bacterial genes for laccase-like enzymes. PLoS ONE 6:e25724
Baldrian P (2006) Fungal laccases – occurrence and properties. FEMS Microbiol. Rev 30:215–242
Beloqui A, Pita M, Polaina J, Martinez-Arias A, Golyshina OV, Zumarraga M, Yakimov MM, Garcia-Arellano H, Alcalde M, Fernandez VM, Elborough K, Andreu JM, Ballesteros A, Plou FJ, Timmis KN, Ferrer M, Golyshin PN (2006) Novel polyphenol oxidase mined from a metagenome expression library of bovine rumen: biochemical properties, structural analysis, and phylogenetic relationships. J Biol Chem 281:22933–22942
Brander S, Mikkelsen J D, Kepp K P (2014) Characterization of an alkali and halide-resistant laccase expressed in E. coli: CotA from Bacillus clausii. PLoS One 9: e99402
Brijwani K, Rigdon A, Vadlani PV (2010) Fungal laccase: production, function, and applications in food processing. Enzyme Res 2010:149748
Claus H (2003) Laccases and their occurrence in prokaryotes. Arch Microbiol 179:145–150
Diamantidis G, Effosse A, Potier P, Bally R (2000) Purification and characterization of the first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum. Soil Biol Biochem 32:919–927
Fang ZM, Li TL, Wang Q, Zhang XC, Peng H, Fang W, Hong YZ, Ge HH, Xiao YZ (2011) A bacterial laccase from marine microbial metagenome exhibiting chloride tolerance and dye decolorization ability. Appl Microbiol Biotechnol 89:1103–1110
Feng HW, Zhang D, Sun YJ, Zhi Y, Mao L, Luo YQ, Xu L, Wang LM, Zhou P (2015) Expression and characterization of a recombinant laccase with alkalistable and thermostable properties from Streptomyces griseorubens. Appl Biochem Biotechnol 176:547–562
Guan ZB, Zhang N, Song CM, Zhou LX, Zhao H, Xu CW, Cai YJ, Liao XR (2012) Molecular cloning, characterization, and dye-decolorizing ability of a temperature- and pH-stable laccase from Bacillus subtilis X1. Appl Biochem Biotechnol. 172:1147–1157
Gunne M, Urlacher VB (2012) Characterization of the alkaline laccase Ssl1 from Streptomyces sviceus with unusual properties discovered by genome mining. PLoS One 7:e52360
Guo X, Zhou S, Wang YW, Song JL, Wang HM, Kong DL, Zhu J, Dong WW, He MX, Hu GQ, Ruan ZY (2016) Characterization of a highly thermostable and organic solvent-tolerant copper-containing polyphenol oxidase with dye-decolorizing ability from Kurthia huakuii LAM0618T. PLoS One 14:e0164810
Hilden K, Hakala TK, Lundell T (2009) Thermotolerant and thermostable laccases. Biotechnol Lett 31:1117–1128
Hoegger PJ, Kilaru S, James TY, Thacker JR, Kües U (2006) Phylogenetic comparison and classification of laccase and related multicopper oxidase protein sequences. FEBS J 273:2308–2326
Kahar UM, Chan KG, Salleh MM, Hii SM, Goh KM (2013) A high molecular-mass Anoxybacillus sp. SK3-4 amylopullulanase: characterization its relationship in carbohydrate utilization. Int J Mol Sci 28 14(6):11302–11318
Kalmea S, Jadhavb S, Jadhavb M, Govindwar S (2009) Textile dye degrading laccase from Pseudomonas desmolyticum NCIM 2112. Enzyme Microb Technol 44:65–71
Kaushik G, Thakur IS (2013) Purification, characterization and usage of thermotolerant laccase from Bacillus sp. for biodegradation of synthetic dyes. Appl Biochem Microbiol 49:352–359
Kudanga T, Le Roes-Hill M (2014) Laccase applications in biofuels production: current status and future prospects. Appl Microbiol Biotechnol 98:6525–6542
Kumari A, Kishor N, Guptasarma P (2018) Characterization of a mildly alkalophilic and thermostable recombinant Thermus thermophilus laccase with applications in decolorization of dyes. Biotechnol Lett 40:285–295
Leonowicz A, Cho NS, Luterek J, Wilkolazka A, Wasilewska MW, Matuszewska A, Hofrichter M, Wesenberg D, Rogalskai J (2001) Fungal laccase: properties and activity on lignin. Basic Microbiol 41:185–227
Liu YH, Ye M, Lu Y, Zhang X, Li G (2011) Improving the decolorization for textile dyes of a metagenome derived alkaline laccase by directed evolution. Appl Microbiol Biotechnol 91:667–675
Lu L, Zhao M, Wang TN, Zhao LY, Du MH, Li TL, Li DB (2012) Characterization and dye decolorization ability of an alkaline resistant and organic solvents tolerant laccase from Bacillus licheniformis LS04. Bioresour Technol 115:35–40
Mate DM, Alcalde M (2016) (2016) Laccase: a multi-purpose biocatalyst at the forefront of biotechnology. Microb Biotechnol. 10:1457–1467
Miyazaki K (2005) A hyper thermophilic laccase from Thermus thermophilus HB27. Extremophiles 9:415–425
Molina-Guijarro JM, Perez J, Munoz-Dorado J, Guillen F, Moya R, Hernandez M, Arias ME (2009) Detoxification of azo dyes by a novel pH-versatile, salt-resistant laccase from Streptomyces ipomoea. Int Microbial 12:13–21
Murugesan K, Arulmani M, Nam IH, Kim YM, Chang YS, Kalaichelvan PT (2006) Purification and characterization of laccase produced by a white rot fungus Pleurotus sajorcaju under submerged culture condition and its potential in decolorization of azo dyes. Appl Microbiol Biotechnol 72:939–946
Murugesan K, Kim Y, Jeon J, Chang Y (2009) Effect of metal ions on reactive dye decolorization by laccase from Ganoderma lucidum. J Hazard Mater 168:523–529
Nagai M, Sato T, Watanabe H, Saito K, Kawata M, Enei H (2002) Purification and characterization of an extracellular laccase from the edible mushroom Lentinula edodes, and decolorization of chemically different dyes. Appl Microbiol Biotechnol 60:327–335
Othman AM, Elsayed MA, Elshafei AM, Hassan MM (2018) Purification and biochemical characterization of two isolated laccase isoforms from Agaricus bisporus CU13 and their potency in dye decolorization. International Journal of Biological Macromolecules 113:1142–1148
Pereira L, Coelho AV, Viegas CA, Santos MM, Robalo MP, Martins LO (2009) Enzymatic biotransformation of the azo dye Sudan Orange G with bacterial CotA-laccase. J Biotechnol 139:68–77
Qiao W, Chu J, Ding S, Song X, Yu L (2017) Characterization of a thermo-alkali-stable laccase from Bacillus subtilis cjp3 and its application in dyes decolorization. J Environ Sci Health A Tox Hazard Subst Environ Eng 52:710–717
Rai HS, Bhattacharya MS, Singh J, Bansal TK, Vats P, Banerjee UC (2005) Removal of dyes from the effluent of textile and dyestuff manufacturing industry: a review of emerging techniques with reference to biological treatment. Crit Rev Environ Sci Technol 35:219–238
Robles A, Lucas R, Martinez-canamero M, Omar NB, Peres R, Galvez A (2002) Characterization of laccase activity produced by the hyphomycete Chalara (syn. Thielaviopsis) paradoxa CH32. Enzyme Microb Technol 31:516–522
Sharma P, Goel R, Capalash N (2007) Bacterial laccases. World J Microbiol Biotechnol 23:823–832
Shraddha SR, Sehgal S, Kamthania M, Kumar A (2011) Laccase: microbial sources, production, purification, and potential biotechnological applications. Enzyme Res. https://doi.org/10.4061/2011/217861
Sonica S, Prince S, Shilpa S, Neena P, Naveen G (2014) Purification and characterization of an extracellular, thermo-alkali-stable, metal tolerant laccase from Bacillus tequilensis SN4. PLOS ONE 9:e96951
Sunil G, Youri Y, Woo-Young S, Tae-Young K, Hor-Gil H (2018) A novel laccase from thermoalkaliphilic bacterium Caldalkalibacillus thermarum strain TA2.A1 able to catalyze dimerization of a lignin model compound. Appl Microbiol Biotechnol 102:4075–4086
Telke AA, Ghodake GS, Kalyani DC, Dhanve RS, Govindwar SP (2011) Biochemical characteristics of a textile dye degrading extracellular laccase from a Bacillus sp. ADR. Bioresour Technol 102:1752–1756
Verma P, Madamwar D (2003) Decolourization of synthetic dyes by a newly isolated strain of Serratia marcescens. World J Microbiol Biotechnol 19:615–618
Yang Q, Zhang M, Zhang M, Wang C, Liu Y, Fan X, Li H (2018) Characterization of a novel, cold-adapted, and thermostable laccase like enzyme with high tolerance for organic solvents and salt and potent dye decolorization ability, derived from a marine metagenomic library. Front Microbiol. https://doi.org/10.3389/fmicb.2018.02998
Zhang C, Zhang S, Diao H, Zhao H, Zhu X, Lu F, Lu Z (2013) Purification and characterization of a temperature and pH-stable laccase from the spores of Bacillus Vallismortis fmb-103 and its application in the degradation of malachite green. J Agric Food Chem 61:5468–5473
Research involving human participants and/or animals
N/A
Informed consent
N/A
Funding
This work was funded by the Natural Science Foundation of Anhui Province (1708085QC68), National Natural Science Foundation of China (31800049), National Natural Science Foundation of China (31800042), Foreign Visiting Program for Young Outstanding Talents in Anhui Province (gxgwfx2018067), and Talent Research Fund Project of Hefei University (16-17RC05)
Author information
Authors and Affiliations
Contributions
The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1.
Figure S1. DNA sequence of LacAn after codon optimizaion
Additional file 2.
Figure S2. The chemical structures of the 5 synthetic dyes
Additional file 3.
Table S1. Characteristics of LacAn compared with reported prokaryotic laccases
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, J., Chang, F., Tang, X. et al. Bacterial laccase of Anoxybacillus ayderensis SK3-4 from hot springs showing potential for industrial dye decolorization. Ann Microbiol 70, 51 (2020). https://doi.org/10.1186/s13213-020-01593-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13213-020-01593-6