
Abstract
Background
The excretory/secretory (ES) antigen of Trichinella spiralis muscle larvae (ML) is currently the most widely used diagnostic antigen to detect T. spiralis infection. However, this antigen has certain drawbacks, such as a complicated ES antigen preparation process and lower sensitivity during the early phase of infection. The aim of this study was to investigate the features of a novel T. spiralis trypsin (TsTryp) and evaluate its potential diagnostic value for trichinellosis.
Methods
The TsTryp gene was cloned and recombinant TsTryp (rTsTryp) expressed. Western blotting and an enzyme-linked immunosorbent assay (ELISA) were performed to confirm the antigenicity of rTsTryp. The expression pattern and distribution signature of TsTryp at various life-cycle stages of T. spiralis were analyzed by quantitative PCR, western blotting and the immunofluorescence test. An ELISA with rTsTryp and ML ES antigens was used to detect immunoglobulins G and M (IgG, IgM) in serum samples of infected mice, swine and humans. The seropositive results were further confirmed by western blot with rTsTryp and ML ES antigens.
Results
TsTryp expression was observed in diverse T. spiralis life-cycle phases, with particularly high expression in the early developmental phase (intestinal infectious larvae and adults), with distribution observed mainly at the nematode outer cuticle and stichosome. rTsTryp was identified by T. spiralis-infected mouse sera and anti-rTsTryp sera. Natural TsTryp protease was detected in somatic soluble and ES antigens of the nematode. In mice infected with 200 T. spiralis ML, serum-specific IgG was first detected by rTsTryp-ELISA at 8 days post-infection (dpi), reaching 100% positivity at 12 dpi, and first detected by ES-ELISA at 10 dpi, reaching 100% positivity at 14 dpi. Specific IgG was detected by rTsTryp 2 days earlier than by ES antigens. When specific IgG was determined in serum samples from trichinellosis patients, the sensitivity of rTsTryp-ELISA and ES antigens-ELISA was 98.1% (51/52 samples) and 94.2% (49/52 samples), respectively (P = 0.308), but the specificity of rTsTryp was significantly higher than that of ES antigens (98.7% vs. 95.4%; P = 0.030). Additionally, rTsTryp conferred a lower cross-reaction, with only three serum samples in total testing positive from 11 clonorchiasis, 20 cysticercosis and 24 echinococcosis patients (1 sample from each patient group).
Conclusions
TsTryp was shown to be an early and highly expressed antigen at intestinal T. spiralis stages, indicating that rTsTryp represents a valuable diagnostic antigen for the serodiagnosis of early Trichinella infection.
Graphical Abstract

Similar content being viewed by others

Background
Trichinellosis is an important zoonotic parasitosis caused by nematodes of the genus Trichinella. In humans, Trichinella infection occurs through the ingestion of raw or semi-cooked meat of animals infected with Trichinella larvae, with Trichinella spiralis the prime causative agent [1, 2]. This parasitic disease can occur worldwide, with 50 trichinellosis outbreaks reported in the European Union between 2015 and 2019 [3] and 13 trichinellosis outbreaks involving 1604 cases and 12 deaths were recorded in Southeast Asian countries from 2001 to 2021 [4]. In China, specifically, eight trichinellosis outbreaks were documented during the 2009–2020 period, with 479 patients and two deaths reported [5]. The consumption of undercooked pork and pork products are the predominant source of T. spiralis infection. Trichinellosis therefore remains a thorny challenge and threat to public health and food safety [6].
Following the ingestion of Trichinella-infected meat, muscle larvae (ML) are liberated and develop into intestinal infectious larvae (IIL) in the small intestine, where they undergo molting 4 times to ultimately develop into adult worms (AW). Following copulation, the mature females produce newborn larvae (NBL), which penetrate the skeletal muscles through the circulatory and lymphatic systems and develop into the encapsulated ML [7]. The clinical manifestations of trichinellosis at these different development stages differ. The initial intestinal stage of the infection is often accompanied by nonspecific gastrointestinal symptoms, including nausea, emesis, abdominal pain and diarrhea. The major symptoms and signs at the muscular stage (acute stage) are fever, periorbital edema, myalgia or muscle weakness, skin eruption, among others [8, 9]. Clinical diagnosis of trichinellosis is rather difficult due to its non-pathognomonic symptomatology in the initial stage of infection, especially in sporadic cases and upon presentation of early cases of a new outbreak. Moreover, medical practitioners from non-endemic areas are usually unfamiliar with the manifestations of trichinellosis, and patients are often not taken seriously and/or misdiagnosed.
The definitive diagnosis of trichinellosis can be established on the basis of epidemiological investigation (focusing on a history of raw or undercooked meat consumption), muscle biopsy and serological testing. However, muscle biopsy is not suitable for patients with a mild infection and/or during early infection [10]. At the present time, T. spiralis ML excretory/secretory (ES) antigen-based serological assays are extensively applied for the detection of the infection, as recommended by the International Commission on Trichinellosis (ICT) and the Food and Agriculture Organization of the UN (FAO)/WHO/World Organization for Animal Health (OIE) [11, 12]. Nevertheless, the main drawbacks of this ML ES antigen are a low sensitivity during the early infection period due to a 3- to 5-week-long window between Trichinella infection and seroconversion for positivity. Seroconversion in patients with trichinellosis often occurs between the third and fifth weeks post-infection [13, 14]. In addition to exposure level (e.g. the infectious dose), stage-specific antigens used in the immunodiagnosis are also an important factor affecting the detection of Trichinella-specific immunoglobulin G (IgG). Recently, the presence of circulating antigen and DNA in serum samples as alternative markers for detecting T. spiralis infection have received increasing attention. However, some studies showed that the levels of free antigens circulating in the blood are generally low due to the existence of immune complexes. The authors of one study reported that the overall positive rate of circulating antigen in patients with clinical trichinellosis was only 30–50% and, in addition, the persistent period of T. spiralis DNA was transitory in infected host’s blood [15]. Therefore, it is necessary to explore novel detection/diagnostic antigens to achieve a simple, accurate, reliable and early diagnosis of trichinellosis. An enzyme-linked immunosorbent assay (ELISA) with ML ES antigens has demonstrated superior performance in terms of sensitivity as well as an acceptable specificity and reproducibility; it has been widely applied to the serological diagnosis of trichinellosis [12, 16]. However, the ES antigens prepared from the ML were found to be stage specific and may not contain epitopes identified by the antibodies produced at the intestinal stage [17]. At the enteral phase of T. spiralis infection, surface and ES proteins from IIL and AW are expressed early during infection to simulate the host’s immune system and trigger production of enteral Trichinella-specific antibodies. Therefore, the surface and ES proteins from the IIL and AW might contain the early intestinal stage-specific expressed protein molecule markers for early diagnosis of trichinellosis [18, 19].
In one study, T. spiralis adult crude antigens were probed by serum from infected swine or mice 7 days post-infection (dpi) [20]. Recombinant proteins from IIL or pre-adults have been identified by serum from infected swine as early as 15–20 dpi [21, 22]. Trichinella spiralis-specific IgG in infected mouse sera was detected by indirect ELISA with AW and IIL ES antigens at 8–10 dpi, but not detected by ELISA with the ML ES antigens prior to 12 dpi. Moreover, the ELISA with these worm ES antigens also had low false positivity with infection serum of other parasites [23, 24]. Therefore, novel diagnostic antigens for trichinellosis need to be further characterized and developed, with a focus on worms of the intestinal stage (IIL and AW) of T. spiralis [25, 26].
Additionally, the preparation of the ML ES antigens is cumbersome and time-consuming, requiring first the collection of ML from T. spiralis-infected laboratory animals and then their culture to prepare ES antigens in vitro [27, 28]. Although ES antigens are less complex than worm somatic crude antigens, they still have the potential to cross-react with other parasitic helminths [29]. Thus, the ICT recommended that positive results of ELISA with ML ES antigens need to be further confirmed by western blot [12]. Recently, the preparation of recombinant antigens using DNA recombinant technology has proved to be time-saving and relatively cost-effective and to have a good reproducibility. Therefore, recombinant Trichinella antigens have the potential to be serodiagnostic antigens for trichinellosis [30,31,32].
In preliminary studies, we identified a novel highly expressed trypsin of T. spiralis (TsTryp, GenBank: XM_003381619.1) in IIL ES antigens using sera from infected mice at 8–10 dpi and sera from patients with early trichinellosis [26, 33, 34]. TsTryp is a likely to be a potential early diagnostic antigen. In the present study we aimed to further determine the biological properties of TsTryp and evaluate its prospective serodiagnostic values for early trichinellosis.
Methods
Worm and animals
The T. spiralis isolate used in this study (ISS534) was obtained from an infected domestic pig in Henan province of China [35]. In our laboratory, this nematode was passaged in mice at 6-month intervals. For the present study, we bought 20 female BALB/c mice (age: 8 weeks; weight: 20–25 g each) from Henan Provincial Medical Laboratory Animal Center (Zhengzhou, China). All of the mice were raised in individual ventilated cage under the specific-pathogen-free (SPF) condition, and each mouse was orally infected with 200 T. spiralis ML.
Serum samples
Blood samples (100 μl) were collected from the tail vein of each of the 20 mice infected with 200 T. spiralis ML on alternate days between 0 and 30 dpi [31]. Serum samples were collected from mice infected with 200 ML of T. spiralis (T1), Trichinella nativa (T2), Trichinella britovi (T3), Trichinella pseudospiralis (T4) and Trichinella nelsoni (T7) at 35 dpi. Sera from mice infected with Angiostrongylus cantonensis, Capillaria hepatica, Schistosoma japonicum and Spirometra mansoni plerocercoids and from Toxoplasma gondii were collected in our laboratory or were a gift from our Chinese colleagues. Fifty individual serum samples from uninfected healthy mice were used as the negative serum control. Eighteen large white pigs were inoculated orally with 5000 ML; infected pigs were sacrificed at 70 dpi, and infected serum (swine infection serum) was obtained. Serum samples from swine naturally infected with Ascaris suum were a present from Prof. Ming Xin Song (Northeast Agricultural University, China).
Fifty-two serum samples were obtained from 52 patients treated for trichinellosis during two trichinellosis outbreaks that occurred in Yunnan province, southwestern China in 2003 (20 patients [age range: 12–65 years; 12 males]; serum collected at 19 dpi) and 2013 (32 patients [age range: 15–70 years; 25 males]; serum collected at 35 dpi). All patients had a history of consuming raw or undercooked meat and showed typical clinical trichinellosis manifestations, including fever, myalgia, periorbital or facial edema and eosinophilia. All patients tested positive for Trichinella-specific IgG antibodies by the ML ES antigen ELISA; muscle biopsy was performed in two trichinellosis patients at 35 dpi and encapsulated ML were found [8, 24]. All serum samples of these trichinellosis patients were tested again in our laboratory by ELISA with ML or AW ES antigens; the anti-Trichinella IgG was positive [24]. The etiological Trichinella species of two trichinellosis outbreaks was not identified. Since only one species T. spiralis of the Trichinella genus was found in southwestern China, the causative agent of the two trichinellosis outbreaks is likely to be T. spiralis [36]. Serum samples from patients with other parasite infections were conserved in our laboratory as follows: five cases with hepatic capillariasis, three cases with ancylostomiasis (Ancylostoma duodenale), 12 cases with lymphatic filarialsis (Wuchereria bancrofti), 25 cases with paragonimiasis (Paragonium skrjabini), 11 cases with clonorchiasis (Clonorchis sinensis), 30 cases with schistosomiasis (Schistosoma japonicum), eight cases with sparganosis (Spirometra mansoni sparganum), 20 cases with cysticercosis (Taenia solium cysticercus) and 24 cases with cystic echinococcosis (Echinococcus granulosus). The confirmatory diagnosis of the patients with parasitic diseases was made on the basis of serological tests and fecal parasitological examination [24, 37]. For comparison, 100 serum samples form healthy individuals residing in non-trichinellosis endemic areas were collected and tested as a negative result for the parasite infection tests mentioned above.
Collection of worms and its antigens
Murine muscles infected with T. spiralis at 42 dpi were artificially digested and the ML were obtained as previously reported [38]. The IIL and AW were isolated from the intestines of infected mice at 6 h post-infection (hpi) and 3 and 6 dpi, respectively [39, 40]. Female AW at 6 dpi were first washed in phosphate-buffered saline (PBS), then cultivated at 37 °C for 18 h in RPMI 1640 medium (Sangon, Shanghai, China) supplemented with 100 μg/ml streptomycin and 200 U/ml penicillin, following which NBL were recovered [27, 41]. Somatic soluble proteins (crude antigens) from various T. spiralis developmental phases, including ML, IIL, AW and NBL, were prepared as described previously [42, 43]. The ML ES antigens were prepared according to a previously reported method [23]. Briefly, the ML were washed throughly in sterile normal saline solution and serum-free RPMI-1640 medium containing 200 U penicillin/ml and 100 μg streptomycin/ml, and then cultivated at 5000 ML/ml in the same culture medium under 5% CO2 at 37 °C for 18 h. After cultivation, the medium containing the ES proteins were filtered through a 0.2-μm membrane into a 50-ml conical tube and then centrifuged at 5000 g, 4 °C for 2 h. The supernatant was filtered once again through an Amicon Ultra 3K centrifugal filter. The protein concentration of ML ES antigens determined by the Bradford assay was 1.29 mg/ml. The filtered supernatant was preserved at − 80 °C until use.
Analysis of TsTryp sequence and phylogeny
The NCBI GenBank was searched for the peptide sequence of TsTryp (GenBank: XM_003381619.1). The physicochemical characteristics, functional domain, transmembrane domain and signal peptide of TsTryp were analyzed and predicted by the bioinformatics online websites ExPasy, NCBI, SignalP 5.0 and TMHMM [44]. The tertiary structure and enzyme active sites of TsTryp were predicted using CN3D and PyMOL software [45]. Phylogenetic analysis of amino acid sequences of TsTryp and of trypsin of other Trichinella species was performed using MEGA7.0 software, and their phylogeny was constructed on the basis of neighbor-joining (NJ) method [46, 47]. The GenBank accession numbers of trypsin’s peptide sequence from different Trichinella species and other organisms are as follows: T. nativa (KRZ48719.1), T. britovi (KRY59262.1), T. psendospiralis (KRY77859.1), T. murrelli (KRX44632.1), Trichinella T6 (KRX79013.1), T. nelsoni (KRX20911.1), Trichinella T8 (KRZ96745.1), Trichinella T9 (KRX62817.1), T. papuae (KRZ66153.1), T. patagoniensis (KRY20428.1), T. zimbabwensis (KRZ13763.1), Trichuris suis (KFD57430.1), Trichuris trichiura (CDW57755.1), Caenorhabditis elegans (NP_501379.2), Necator americanus (XP_013293798.1), Ancylostoma caninum (RCN34081.1), Anisakis simplex (VDK43792.1), Brugia malayi (XP_042934028.1), Steinernema carpocapsae (TKR57687.1), Homo sapiens (XP_011526282.1) and Mus musculus (P69525.1).
Preparation of recombinant TsTryp and anti-recombinant TsTryp serum
The RNAs of IIL were isolated using TRIzol (Sangon). The TsTryp coding sequence of 1956 bp was amplified by PCR with specific primers: (i) 5′-CCGAATTCCTACCAGATGACAA ATGTGGATT-3′ containing the restriction enzymatic digestion site of EcoRI (underlined); and (ii) 5′-TCGTCGACTCAAAGTGTCTG AAAACGGGAAC-3′ containing the restriction enzymatic digestion site of SalI (underlined). The pET-32a plasmid was used to link PCR products and to construct the recombinant expression plasmid pET-32a/TsTryp, which were transferred into Escherichia coli BL21 (DE3). Subsequently, the pET-32a/TsTryp plasmid was induced for 6 h at 25 °C with 0.8 mM IPTG as inducer [48]. Recombinant TsTryp (rTsTryp) and thioredoxin (TRX) protein were purified using the Ni–NTA Sefinose Resin Kit (BBI Life Sciences Corp., Shanghai, China) [45]. rTsTryp and its antigenicity were analyzed by western blot as reported before [49].
Thirty female mice were immunized by subcutaneous injection with rTsTryp (20 mice, 20 μg/mouse) and TRX tag protein (10 mice, 20 μg/mouse) emulsified with complete Freund’s adjuvant. Boost immunization with rTsTryp and TRX protein emulsified in incomplete Freund’s adjuvant at a 1:1 ratio was administered twice with a 2-week interval between boosters [44, 50]. At 2 weeks after the last immunization, blood samples were obtained via tail-vein bleeding, and anti-rTsTryp and anti-TRX immune sera were isolated; the antibody level of anti-rTsTryp and anti-TRX IgG was detected by conventional indirect ELISA using rTsTryp and TRX protein (2.0 μg/ml) [51]. The specific IgG titer of both anti-rTsTryp and anti-TRX IgG was 1:105, and the optical density (OD) value of anti-rTsTryp and anti-TRX IgG was 1.33 ± 0.53 (n = 20) and 1.20 ± 0.32 (n = 10), respectively. Serum samples were also collected from uninfected normal mice prior to rTsTryp immunization; these samples were considered to be the pre-immune serum.
Western blot
rTsTryp, ML crude antigens and ES antigens were analyzed using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and the protein products were electrophoretically transferred to a nitrocellulose (NC) membrane (MerckMillipore, Burlington, MA, USA). For blocking, the membranes were placed in non-fat milk (5%) dissolved in Tris-buffer saline containing 0.05% Tween (TBST) at 37 °C for 2 h. The strip was then probed with the different sera mentioned above (infected serum, anti-rTsTryp serum, anti-TRX serum and pre-immune serum) at 1:200 overnight at 4 °C [42]. After rinsing in TBST, the strip was incubated with horse radish peroxidase (HRP)-conjugated anti-mouse/swine/human IgG as the secondary antibodies, respectively (1:10,000 dilutions; SouthernBiotech., Birmingham, AL, USA) at room temperature for 1 h. Finally, staining was performed with the substrate 3, 3′-diaminobenzidine (DAB; Sigma-Aldrich, St Louis, MO, USA) [52, 53]. To determine the expression levels of TsTryp at diverse T. spiralis stages, the strips were visualized by enhanced chemiluminescence (ECL) using the ultrasensitive ECL chemiluminescence reagent kit (ECL; Meilunbio, China) and the bands then analyzed using ImageJ software [54].
Quantitative PCR
Trichinella spiralis at different developmental stages were collected and their total RNAs were extracted using TRIzol reagent (Sangon). The PrimeScript RT Reagent Kit (TaKaRa, Kyoto, Japan) was used to reverse transcribe the RNAs into complementary DNA (cDNA) [55, 56]. The SYBR Green PCR Master Mix (Takara) was used for quantitative PCR (qPCR), using the TsTryp primer sequences for qPCR (5′-GGAGCTGAATCATCTTATGG-3′ and 5′-CGTCTATCTGCTTTACCACC-3′). The expression level of TsTryp messenger RNA (mRNA) was normalized to that of T. spiralis glyceraldehyde 3-phosphate dehydrogenase (GAPDH; GenBank: AF452239) as the internal reference. Each assay was carried out in triplicate. Relative quantification of TsTryp was analyzed by the 2−ΔΔCt method [57].
Immunofluorescence test
Natural TsTryp expression on the surface of cuticles during the different T. spiralis worm phases and tissue localization were investigated by the immunofluorescence test (IFT) on entire worms as well as worm cross-sections [58, 59]. Briefly, fresh worms at diverse stages were fixed in 4% paraformaldehyde followed by embedding. Serial 3-µm-thick worm sections were cut and fixed on glass slides. Sections and whole worms were sealed at room temperature for 2 h using PBS supplemented with 5% normal goat serum, followed by washing in PBS before being probed for 10 h at 4 °C with infection serum, anti-rTsTryp serum, pre-immune serum and anti-TRX serum, respectively, diluted at 1:20. On the second day, the treated parasites and cross-sections were washed in PBS, then stained by FITC-goat anti-mouse IgG conjugate (Santa Cruz Biotechnology, Dallas, TX, USA) at 1:100 dilutions, at 37 °C in dark for 1 h, and examined by fluorescent microscopy (Olympus, Tokyo, Japan) [60, 61].
Assay of anti-Trichinella antibodies by ELISA
The optimal coating concentration of rTsTryp and ML ES antigens, dilution of serum samples and conjugates were first assessed by the checkerboard titration method, as reported previously [62]. The titration results revealed that 2.0 μg/ml of rTsTryp and ES antigens, respectively, was the optimal coating concentration for detecting specific IgG and immunoglobulin M (IgM); the optimal dilutions of mouse, swine and human serum were 1:100 to detect IgG and 1:50 to detect mouse IgM; the optimal conjugate dilution was 1:10,000 dilutions for HRP-conjugated anti-human/mouse/swine IgG; and 1:5000 dilutions were optimal for HRP-anti-mouse IgM conjugate. The ELISA was used to assay anti-Trichinella antibodies as mentioned previously [8, 37]. In brief, the microtiter plate (Corning Inc., Corning, NY, USA) was coated using rTsTryp and ML ES antigens, which were diluted in carbonate bicarbonate buffer (0.05 M, pH 9.6) and incubated overnight at 4 °C. The plate was blocked using 5% skimmed milk diluted in PBS containing 0.05% Tween 20 (PBST) and probed at 37 °C for 2 h with various serum samples (mouse, swine and human sera). After three rinses in PBST, the plate was incubated at 37 °C for 1 h with HRP-conjugated anti-human/mouse/swine IgG or anti-mouse IgM (Southern Biotech.) diluted in PBST. Following further washes, coloration was performed with the substrate O-phenylenediamine dihydrochloride (OPD; Sigma-Aldrich), which was dissolved in phosphate citric acid buffer containing 0.2 M Na2HPO4 and 0.1 M citric acid and supplemented with H2O2, for 30 min; the reaction was stopped with 2 M H2SO4 (50 μl/well). The OD values (absorbance at 492 nm) were measured using a microplate reader (Tecan Austria GmbH, Grödig, Austria). The cutoff value was defined as 2.1-fold the mean OD value of negative control sera of 50 uninfected mice, 55 uninfected pigs or 100 healthy persons [63]. The cutoff values for rTsTryp-ELISA mouse serum IgG and IgM were 0.241 and 0.266, respectively, and those of the ES antigens-ELISA were 0.256 and 0.270, respectively. The cutoff values of the rTsTryp-ELISA and ES antigens-ELISA were 0.271 and 0.281, respectively, for testing swine serum IgG and 0.357 and 0.426, respectively, for testing human serum IgG.
Statistical analysis
The data were analyzed using SPSS version 22.0 software (SPSS IBM Corp., Armonk, NY, USE). Normality and homogeneity of data variances was checked prior to performing analysis of variance (ANOVA). Data were first assessed for normal distribution by the Shapiro–Wilk test. Under the test standard α = 0.05, P > 0.05 indicated that the data were subject to normal distribution. The homogeneity of data variances was also checked by homogeneity of variance test in one-way ANOVA; P > 0.05 was considered to be consistent with homogeneity of variance. The results are presented as the mean ± standard deviation (SD). Expression levels of rTsTryp at diverse T. spiralis worm stages were analyzed by one-way ANOVA. The sensitivity and specificity of rTsTryp and ML ES antigens were compared by the Chi-square test (χ2). P-values < 0.05 were accepted as indicating statistical significance.
Results
Biological features of TsTryp
The TsTryp cDNA encoding 667 amino acid residues comprises 2004 bp. This protein has a molecular weight of 71.6 kDa and isoelectric point (pI) of 8.83. The N terminal of TsTryp has a typical hydrophobic structure, and all 67 amino acids are outside of the membrane. Alignment analysis of the amino acid sequence of TsTryp with those of the trypsin of eight encapsulated Trichinella species (T. patagoniensis, T. nelsoni, T. murrelli, Trichinella T6, T. nativa, T. britovi, Trichinella T8 and Trichinella T9) revealed 94.5%, 94.2%, 94.0%, 94.0%, 93.9%, 93.7%, 93.5% and 93.4% similarity, respectively. Also, a similarity of 75.6%, 75.4% and 71.4% between TsTryp and the trypsin from three non-encapsulated Trichinella species (T. papuae, T. zimbabwensis and T. pseudospiralis), respectively, was also found (Additional file 1: Figure S1). Prediction of the subcellular localization of TsTryp revealed that TsTryp had an extracellular localization and might be part of secretory protein. Moreover, prediction of TsTryp structure suggested that TsTryp had two independent and similar trypsin-like domains and that there were three active sites (Asp, Ser and His) in each domain (Figs. 1a, b).

Tertiary structure of TsTryp and phylogeny of trypsin in 22 organisms by NJ method. a Online analysis of the NCBI and SMART databases showed that TsTryp had two identical and independent trypsin-like serine protease domains (Tryp_SPc). b The NCBI-Cn3D online website was used to conduct homology modeling for TsTryp and predict its tertiary structure. The secondary structure of TsTryp comprises alpha-helices (green cylinders), beta-strands (brown sheets) and random coils (gray strands). TsTryp had two similar trypsin-like domains; the three enzyme active sites of Asp, Ser and His are shown as yellow spheres. c Phylogenetic tree of the trypsin of Trichinella spiralis and other nematodes were constructed by NJ method in MEGA7.0 software. NJ, Neighbor-joining; TsTryp, T. spiralis trypsin

Identification of rTsTryp. A SDS-PAGE of rTsTryp. Lanes: M, Protein marker; 1, lysates of non-induced BL21 containing the pET-32a/TsTryp plasmid; 2, lysates of induced BL21 with pET-32a/TsTryp; 3, purified rTsTryp. b Western blot of rTsTryp. Lanes: M, Protein marker; 1, lysates of non-induced pET-32a/TsTryp was not identified by serum of infected mice (murine infection serum); 2, lysates of pET-32a/TsTryp following induction were identified using murine infection serum; 3, 4, purified rTsTryp probed using murine infection serum (lane 3) and monoclonal antibody to His tag (lane 4), respectively. Arrow indicates the rTsTryp protein band of 91.6 kDa. BL21, Escherichia coli strain BL21; rTsTryp, recombinant T. spiralis trypsin; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis
The phylogeny of TsTryp with trypsins from other nematodes is shown in Fig. 1c. Specifically, the phylogenetic tree demonstrates that the genus Trichinella is monophyletic, with TsTryp having a relatively closer genetic relationship with the enteral parasitic nematodes of the genus Trichuris (T. suis and T. trichiura). Two clear clades can be seen in the genus Trichinella: an encapsulated clade (T. patagoniensis, T. nelsoni, T. murrelli, Trichinella T6, T. nativa, , T. britovi, Trichinella T8, Trichinella T9) and a non-encapsulated clade (T. papuae, T. zimbabwensis, T. pseudospiralis).
Expression and identification of rTsTryp
The TsTryp gene was cloned and the rTsTryp protein expressed. After induction with IPTG, the TRX-His + TsTryp fusion protein was expressed in E. coli BL21 carrying the pET-32a/TsTryp plasmid. SDS‒PAGE of the purified rTsTryp showed that rTsTryp had an apparent single protein band of 91.6 kDa, which corresponded to the predicted size, including the 71.6-kDa TsTryp and the approximately 20-kDa TRX + His-tag (Fig. 2a). The results were further validated by rTsTryp being recognized by T. spiralis infection serum on the western blot (Fig. 2b).
Following separation of the soluble antigens and ES antigens of ML (Fig. 3a), western blot analysis showed that native TsTryp of 31.8–71.6 kDa in the crude soluble antigens of ML were detected by anti-rTsTryp serum while the infected serum detected the native proteins of 31.0–90.5 kDa. In ES antigens of the ML, natural TsTryp of 32.7–71.6 kDa was detected by anti-rTsTryp serum, while infection serum identified natural proteins of 42.2–71.6 kDa. However, the crude and ES antigens were not identified by anti-TRX serum and uninfected normal serum (Fig. 3b). Native TsTryp of 71.6 kDa in crude antigens of ML, IIL, AW at 3 and 6 dpi (3-/6-day AW) and NBL was also observed by using anti-rTsTryp serum (Fig. 3c). Several forms of natural TsTryp of 31–71.6 kDa from ML, IIL and 3-day AW ES antigens were also identified by anti-rTsTryp serum (Fig. 3d). The results demonstrated that native TsTryp was expressed during diverse T. spiralis life-cycle phases and that TsTryp and its different isoforms belonged to the group of secretory proteins.

Antigenicity analysis of rTsTryp. a SDS-PAGE of rTsTryp. Lanes: M, Protein marker; 1, muscle larvae crude antigens; 2, muscle larvae ES antigens; 3, purified rTsTryp. b Western blot of rTsTryp. Lanes: M, Protein marker; 1, 2, 3, natural TsTryp in muscle larva crude antigens (lane 1), ES antigens (lane 2) and rTsTryp (lane 3), identified by anti-rTsTryp serum; 4–9, natural TsTryp in muscle larva crude antigens (lane 4) and ES antigens (lane 5) and rTsTryp (lane 6) probed using infection serum, but not probed using uninfected murine serum (lanes 7, 8, 9, respectively); 10, 11, 12, natural TsTryp in muscle larva crude antigens (lane 10) and ES antigens (lane 11) not recognized by anti-TRX serum, but rTsTryp was recognized by anti-TRX serum (lane 12). c Western blot of natural TsTryp in crude antigens of muscle larvae, IIL, 3 dpi adult worms, 6 dpi adult worms and NBL (lanes 1–5, respectively), identified using immune serum against rTsTryp. d Natural TsTryp in ES antigens of muscle larvae, IIL and 3 dpi adult worms (lanes 1, 2, 3, respectively) were identified using immune serum against rTsTryp by western blot. ES, Excretory/secretory; IIL, intestinal infective larvae; NBL, newborn larvae; rTsTryp, recombinant T. spiralis trypsin; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; TRX, thioredoxin
Expression level of TsTryp mRNA and protein at various T. spiralis developmental stages
The qPCR data demonstrated that the TsTryp gene was expressed during all life-cycle stages of T. spiralis, including ML, IIL, 3-/6-day AW and NBL (Fig. 4a). The expression level of TsTryp mRNA clearly differed at the different phases (F = 2365.300, P < 0.0001). Notably, the mRNA level of TsTryp at the IIL stage was 3.01-fold higher than that at the ML stage (F = 4412.472, P < 0.0001), and the TsTryp transcriptional level in 3-day-/6-day AW was also remarkably higher than that of the ML (2.22- and 2.87-fold higher, respectively; F3d AW = 724.522, F6d AW = 9166.032, P < 0.0001). The western blot results showed that the expression level of TsTryp in the IIL and 3-/6-day-old AW was significantly higher than that at the ML stage (1.50- 1.35- and 1.33-fold higher, respectively; FIIL = 45.751, P = 0.002; F3d AW = 8.910, P = 0.041; F6d AW = 10.415, P = 0.032) (Fig. 4b). These results demonstrated that TsTryp was expressed during all life-cycle stages of the nematode and that TsTryp was a highly expressed protein early during the IIL and AW stages; consequently, it is likely an early diagnostic molecular marker.

Expression of TsTryp at different developmental stages of T. spiralis. a qPCR test for TsTryp transcription level at different T. spiralis stages, calculated based on the 2−ΔΔCt method. The fold changes in TsTryp gene level were normalized to the respective GAPDH control. b Western blot of TsTryp protein expression level during various worm stages. TsTryp expression in crude antigens of ML larvae, IIL, 3-/6 dpi AW and NBL was assayed by western blot using immune serum against rTsTryp. The data were derived from three separate tests. Asterisk indicates a statistical significant difference at *P < 0.05 compared to the ML stage. AW, Adult worms; d, day post-infection; ES, excretory/secretory; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; h, hour post-infection; IIL, intestinal infective larvae; ML, muscle larvae; mRNA, messenger RNA; NBL, newborn larvae; qPCR, quantitative PCR; rTsTryp, recombinant T. spiralis trypsin
Expression and localization of TsTryp in various T. spiralis life-cycle phases
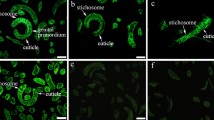
The expression and distribution pattern of TsTryp in the surface of the whole worm and within the worm body was determined by the IFT with anti-rTsTryp serum. A clear green immune fluorescence signal was detected in the epicuticle of T. spiralis at different developmental phases (6- and 12-h IIL, 3- and 6-day AW, NBL), with the exception of the ML (Fig. 5). The results of the IFT with anti-rTsTryp serum on the prepared tissue sections suggested that immune fluorescence was present in the cuticle, stichosome and uterus of female embryos (Fig. 6). No immune fluorescence staining was detected on whole worms or cross-sections using normal serum and anti-TRX serum. Taken together, these results demonstrated that TsTryp is a surface and secretory protein of this parasite and that it is mainly expressed on the surface of intestinal worms (IIL and AW) and their stichosome.

Expression of TsTryp in the epicuticle of diverse developmental stages of T. spiralis by IFT. Entire intact worms were incubated with immune serum against rTsTryp, and a green immunofluorescence signal was observed on the outer cuticle of T. spiralis at diverse life-cycle phases, including 6- and 12-hpi IIL, 3- and 6-dpi AW and NBL, but not ML. Pre-immune serum and anti-TRX serum did not detect any immunofluorescence signal in the worms. Trichinella-infected murine serum was applied as a positive control of reproducibility. Scale bars: 50 μm (ML, 6- and 12-hpi IIL); 100 μm (3- and 6-dpi AW); 25 μm (NBL). AW, Adult worms; IFT, immunofluorescence test; IIL, intestinal infective larvae; ML, muscle larvae; NBL, newborn larvae; rTsTryp, recombinant T. spiralis trypsin; TRX, thioredoxin

Localization of TsTryp in T. spiralis worm tissue by IFT. Green immunofluorescence signal was detected in the epidermis and stichosome and around the uterus of female embryos. No immunofluorescence was probed by pre-immune serum and anti-TRX serum in worm cross-sections. Scale bars: 50 μm. AW, Adult worm; IFT, immunofluorescence test; IIL, intestinal infective larvae; ML, muscle larvae; rTsTryp, recombinant T. spiralis trypsin
Quality assessment of rTsTryp and ES antigens by Western blot
Prior to performing the ELISA, we assessed the quality of rTsTryp and ML ES antigens by western blot using T. spiralis-infected murine sera collected at 14 and 35 dpi. The rTsTryp and ML ES antigens were first separated by SDS-PAGE, and visualization of the products showed a clear individual band of 91.6 kDa for rTsTryp and nine clear bands of 60.0, 49.1, 44.5, 40.6, 33.6, 32.4, 26.9, 24.7 and 21.3 kDa, respectively, for the ML ES antigens (Fig. 7a). These distinct bands of rTsTryp and ES antigens were authenticated by murine infection sera collected at 14 and 35 dpi. Using 20 infection serum samples collected at 14 dpi, three main bands (49.1, 44.5 and 40.6 kDa) of ES antigens were identified, with an identical band pattern in all serum samples (Fig. 7b); also, one distinct band of rTsTryp at 91.6 kDa was identified by the 20 infection serum samples collected at 14 dpi (Fig. 7c). Using 30 murine infection serum samples collected at 35 dpi, the same three main bands (49.1, 44.5, 40.6 kDa) of ES antigens were recognized, but more clearly, as well as an additional band (60 kDa) (Fig. 7D), suggesting that the 60-kDa band may be expressed in the ML stage and that it might be regarded as a diagnostic marker for the muscular phase of T. spiralis infection in mice. The distinct band of rTsTryp at 91.6 kDa was also detected by the 30 infection serum samples collected at 35 dpi (Fig. 7e). In contrast, rTsTryp and ML ES antigens were not recognized by 30 uninfected normal murine serum samples (Fig. 7f, g). The results indicated that the rTsTryp and ML ES antigens prepared in this study were of good quality and that the band pattern identified by the infection serum had good reproducibility.

Quality assessment of rTsTryp and ES antigens by western blot. ML ES antigens and rTsTryp were identified by western blot using T. spiralis-infected murine serum collected at 14 and 35 dpi, respectively. a SDS-PAGE assay of rTsTryp and ML ES antigens. Lanes: M, Protein marker; 1, ML ES antigens; 2, rTsTryp. b Western blot of ES antigens using 20 murine infection serum samples collected at 14 dpi (lanes 1–20), showing recognition of three main bands (49.1, 44.5, 40.6 kDa) of ES antigens. c Western blot of rTsTryp using 20 murine infection serum samples collected at 14 dpi (lanes 1–20), showing recognition of one distinct band of rTsTryp at 91.6 kDa. d Western blot of ES antigens using 30 murine infection serum samples collected at 35 dpi (lanes 1–30), showing recognition of four distinct bands (60, 49.1, 44.5, 40.6 kDa) of ES antigens. e Western blot of rTsTryp using 30 murine infection serum samples collected at 35 dpi (lanes 1–30), showing one distinct rTsTryp band at 91.6 kDa that was more evident than shown in c. f, g rTsTryp (f) and ML ES antigens (g) were not recognized by 30 uninfected murine serum samples. dpi, Days post-infection; ES, excretory/secretory; ML, muscle larvae; rTsTryp, recombinant T. spiralis trypsin; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis
Detection of Trichinella-specific IgG in infected murine serum
Trichinella-specific IgG levels in serum samples from mice infected with 200 T. spiralis ML and other parasites were measured using the rTsTryp-ELISA and ES antigens-ELISA (Table 1). In serum collected from mice infected with 200 T. spiralis ML at 30 dpi, the positive rate of specific IgG was 100% by the two antigens. The two antigens had no cross-reaction with serum collected from mice infected with 30 A. cantonensis third-stage larvae, 200 C. hepatica infective eggs, 60 C. sinensis metacercaria, 100 S. japonicum cercariae, and 4 × 104 T. gondii (RH strain) tachyzoites, although the ES antigens had a cross-reaction with one serum sample (3.33%) collected from S. mansoni plerocercoid-infected mice. However, the subsequent confirmatory test proved that this one positive serum in the ES-ELISA test was negative on western blot analysis (Additional file 2: Figure S2), indicating that the ES-ELISA had one false positive with murine sparganum infection serum.
Seroconversion for positivity of mice experimentally infected with T. britovi and T. nelsoni was 100% at 35 dpi (Table 2), and seroconversion for positivity of specific IgG in mice infected with T. nativa was not significantly different by rTsTryp-ELISA and ES antigens-ELISA (χ2 = 0.015, P = 0.903). However, seropositivity in mice infected with T. pseudospiralis was only 42.86% by rTsTryp-ELISA and 34.29% by ES antigens-ELISA (χ2 = 0.241, P = 0.624). Serum samples from T. pseudospiralis-infected mice collected at 35 dpi were also tested by the ELISA with T. pseudospiralis ML crude antigens. The results showed that the seropositive rate was 100% (35/35) (OD values: 1.99 ± 0.90), while the anti-Trichinella IgG level of 50 normal mouse serum samples was negative (OD values: 0.13 ± 0.03), suggesting that the seroconversion had occurred in all T. pseudospiralis-infected mice at 35 dpi. These findings suggested that rTsTryp and T. spiralis ES antigens were applicable to the serodiagnosis of infection with encapsulated Trichinella species, but not to that of infection with non-encapsulated T. pseudospiralis.
Dynamic changes of serum specific IgG and IgM during 2–30 dpi
We used 20 mice experimentally infected with 200 T. spiralis ML to measure Trichinella-specific IgG and IgM levels in serum samples collected at 2–30 dpi by rTsTryp-ELISA and ES antigens-ELISA. Specific IgG was first detected in 35% (7/20) of the infected mice at 8 dpi by rTsTryp-ELISA and in 40% (8/20) of infected mice at 10 dpi by ES antigens-ELISA (χ2 = 0.107, P = 0.744). Anti-Trichinella IgG seroconversion reached 100% (20/20) at 12 dpi by rTsTryp-ELISA and 14 dpi by ES-ELISA (Fig. 8a, b), and persisted to 30 dpi. At 6 dpi, serum specific IgM was first detected in 75% (15/20) of infected mice by rTsTryp-ELISA and in 35% (7/20) of infected mice by ES-ELISA (χ2 = 6.465, P = 0.011). Detection of specific IgM reached 100% (20/20) at 8 dpi by rTsTryp-ELISA and at 10 dpi by ES antigens-ELISA (Fig. 8c, d). Anti-Trichinella IgM reached peak levels in serum collected at 18–20 dpi, but began to decrease significantly in serum collected on 22 dpi and onwards, although it was still positive by two antigens in serum collected at 30 dpi. These results indicated that the specific antibodies IgG and IgM were detected by rTsTryp 2 days earlier than the ES antigen.

Dynamic changes in serum specific IgG and IgM in serum of mice infected with 20 T. spiralis larvae collected at 2–30 dpi, detected by ELISA. a, b Specific IgG levels in serum of infected mice assayed by rTsTryp-ELISA (a) and ES antigens-ELISA (b). c, d Serum specific IgM levels of infected mice assayed by rTsTryp-ELISA (c) and ES antigens-ELISA (d). The dashed lines in the figures are the cutoff values of IgG and IgM. dpi, Days post-infection; ELISA, enzyme-linked immunosorbent assay; ES, excretory/secretory; IgG/IgM, immunoglobulin G/M; OD, optical density; rTsTryp, recombinant T. spiralis trypsin
In order to further verify the accuracy of the ELISA test, we used western blot with two antigens to confirm the anti-Trichinella IgG positive serum according to the ELISA test. The results are shown in Fig. 9a. Seven serum samples collected at 8 dpi that were positive in the rTsTryp-ELISA test (negative in the ES-ELISA test) were proved to be positive by western blot with rTsTryp; 13 negative serum samples at 8 dpi were still negative according to western blot with rTsTryp. Western blot with rTsTryp showed that at 10 dpi, 14 positive serum samples in the rTsTryp-ELISA test were verified to be positive, whereas six negative serum samples in the rTsTryp-ELISA test were still negative (Fig. 9b). Positive serum at 10 dpi according to the ES-ELISA test was also evaluated by western blot with ES antigens; the results revealed that eight positive serum samples at 10 dpi in the ES-ELISA test were validated to be weakly positive, while the 12 negative serum samples were still negative (Fig. 9c). These results demonstrated that the detection of Trichinella-specific IgG in the serum of infected mice was the same by ELISA and western blot. However, the positive bands of rTsTryp (91.6 kDa) and ES antigens (49.1 kDa) identified by infection sera at 8–10 dpi were indistinct and weak on western blot.

Western blot confirmation of positive serum in the ELISA test. a Western blot confirmation of positive serum at 8 dpi according to the rTsTryp-ELISA test. Seven positive serum samples (lanes 1–7) and 13 negative serum samples (lanes 8–20) in the rTsTryp-ELISA test were verified to be positive and negative, respectively, by rTsTryp-Western blot. b Western blot confirmation of positive serum at 10 dpi according to the rTsTryp-ELISA test. The 14 positive serum samples (lanes 1–14) and 6 negative serum samples (lanes 15–20) in the rTsTryp-ELISA test proved to be positive and negative, respectively, by rTsTryp-western blot. c Western blot confirmation of positive serum at 10 dpi in the ES-ELISA test. The 8 positive serum samples (lanes 1–8) and 12 negative serum samples (lanes 9–20) according to the ES-ELISA test were validated to be weakly positive and negative, respectively, by ES-western blot. All of the positive bands of rTsTryp and ES antigens recognized by western blot with early infection serum were indistinct and weak. The black arrow indicates the rTsTryp band of 91.6 kDa recognized by infection sera; the red arrow indicates the ES antigen band of 49.1 kDa identified by murine infection serum at 10 dpi. dpi, Days post-infection; ELISA, enzyme-linked immunosorbent assay; ES, excretory/secretory; M, protein marker; rTsTryp, recombinant T. spiralis trypsin
Trichinella-specific IgG in serum of T. spiralis-infected pigs
Specific IgG levels in pigs experimentally infected with T. spiralis were measured using rTsTryp-ELISA and ES antigens-ELISA. Seropositivity rate in serum of T. spiralis-infected swine collected at 70 dpi was 100% by rTsTryp-ELISA and ES-ELISA (Table 3). There was no cross-reaction with serum from pigs infected with A. suum and normal porcine sera by two antigens.
Serum anti-Trichinella IgG in trichinellosis patients
The sensitivity of the rTsTryp-ELISA and ES antigens-ELISA to detect serum Trichinella-specific IgG was 98.1% (51/52 samples) and 94.2% (49/52), respectively (χ2 = 1.040, P = 0.308) in trichinellosis patients. The sensitivity of the two antigens was 100% (52/52) for late trichinellosis patients at 35 dpi. The sensitivity of rTsTryp and ES antigens was 95.00% (19/20 samples) and 85.00% (17/20), respectively (χ2 = 1.111, P = 0.292) for serum samples collected from early trichinellosis patients at 19 dpi (Table 4). The specificity of rTsTryp was 98.7% (235/238), which was significantly higher than the 95.4% (227/238) of ES antigens (χ2 = 4.710, P = 0.030). rTsTryp did not cross-react with serum of patients with other parasitosis (hepatic capillariasis, filariasis, ancylostomiasis, paragonimiasis, schistosomiasis and sparganosis) or with serum from healthy individuals; but it did cross-react with one serum sample from one patient with echinococcosis, clonorchiasis or cysticercosis, respectively (Additional file 3: Figure S3). These findings demonstrated that the sensitivity of rTsTryp and ES antigens was not significant different, but the specificity of rTsTryp was remarkably higher than that of the ES antigens.
Discussion
Trypsin is an important serine protease of the chymotrypsin subfamily that plays a critical role in catalyzing protein hydrolysis [64]. Results from previous studies indicated that serine proteases actively participate in the invasion and pathogenic process of some parasites [65]. Serine proteases have been found to be the most plentiful proteinases in ES proteins at different stages of T. spiralis development [39, 66, 67]. These stage-specific serine proteases elicit distinct immune responses during T. spiralis infection [68, 69]. In previous studies, we found that the expressions of serine proteases clearly increased in the IIL phase compared to the ML phase [70], suggesting that serine proteases were involved in the IIL intrusion of the gut mucosa. The serine proteases highly expressed in the enteral IIL phase are exposed early to the host’s local intestinal mucosal and systematic immune system, thereby eliciting the production of intestinal IIL stage-specific antibodies; consequently, serine proteases may represent potential early diagnostic antigens of trichinellosis [66].
In the present study, rTsTryp was expressed and purified. As expected, rTsTryp protein exhibited good immunogenicity, and the immunization of mice with rTsTryp elicited an rTsTryp-specific antibody response with high IgG titer. Western blot results revealed that rTsTryp was recognized by both anti-rTsTryp serum and infection serum. While some native TsTryp at 31.0–71.6 kDa in ES antigens of T. spiralis ML, IIL and 3-day AW were identified by anti-rTsTryp serum, this is possibly explained by: (i) TsTryp belonging to the superfamily of serine proteases, and diverse serine proteases have the similar functional domains and common epitopes; (ii) TsTryp having two trypsin-like domains [71]; and (iii) TsTryp possessing various isoforms, post-translational modifications and protein processing [8, 31, 72, 73].
The results of qPCR and western blot showed that TsTryp protein was expressed in all T. spiralis worm phases (ML, IIL, AW and NBL), although its expression level was distinctly higher in the intestinal stages (IIL, AW). The IFT results demonstrated that TsTryp was distributed on the surface of IIL- and AW-stage worms, and principally localized in the cuticle and stichosome of the nematode; western blot also revealed that several native TsTryp in ES antigens of the nematode was recognized by anti-rTsTryp serum and infection serum. These findings indicated that TsTryp is a surface and secretory protein that is highly expressed in worms of this parasite at the enteral stage. Previous studies showed that other serine proteases of T. spiralis (TsSP, TsSP1.1 and TsSP-1.2) expressed in different worm stages also participated in larval invasion [37, 67, 74]. The results of the current study suggested that TsTryp is a serine protease expressed early during infection and that it may play a significant role in invading gut mucosa and inducing the antibody response at the intestinal stage of T. spiralis infection.
Serological methods are currently in wide use for the detection of the specific antibody IgG in Trichinella infection, with ELISA and western blot with ML ES antigens being the most commonly used serological tests [13]. The ES antigens-ELISA is usually used for the preliminary screening of Trichinella infection, whereas western blot is used as a confirmatory test to verify or exclude the non-negative results in the ES antigens-ELISA test [75,76,77]. In the present study, the quality of rTsTryp was first assessed by western blotting using murine infection sera. The results showed that one clear rTsTryp band (91.6 kDa) and three ES antigen bands (49.1, 44.5 and 40.6 kDa) were distinctly recognized by murine infection serum collected at 14 dpi. An additional band (60 kDa) of ES antigens was also ultimately recognized by murine infection serum collected at 35 dpi, suggesting that this 60-kDa band might be expressed only at the ML stage, and that it is possibly a diagnostic marker for the muscular phase of T. spiralis infection. The ES antigen band pattern recognized by murine infection serum was similar to those recognized by swine infection sera [77]. Although the ML ES antigens had higher specificity than the ML somatic crude antigens, different bands (40–70 kDa) are usually recognized by infection serum on western blot [11]. Small variations in the molecular weight of the bands did not affect the identification of infection sera, but careful observation of the protein migration distances is required [78]. rTsTryp always had a distinct and single band at 91.6 kDa on western blot with infection sera collected at different times post infection, demonstrating that the visual observation of the western blot band pattern of rTsTryp was more simple and obvious compared to that of the ML crude and ES antigens [22, 63].
In T. spiralis-infected mice, anti-Trichinella IgM reached peak levels at 18–20 dpi, followed by a significant decrease beginning at 22 dpi; this trend suggests that Trichinella-specific IgM is only useful for the diagnosis of early trichinellosis [15, 79]. The specific IgG seroconversion reached 100% at 12 dpi by the rTsTryp-ELISA and at 14 dpi by the ES-ELISA. Moreover, rTsTryp had good specificity and did not cross-react with sera from uninfected normal mice and mice infected with other parasites (A. cantonensis, Capillaria hepatica, Clonorchis sinensis, Schistosoma japonicum, Spirometra mansoni and T. gondii). The reason for the low sensitivity of rTsTryp-ELISA and ES-ELISA to detect T. pseudospiralis infection sera may be due to the distant phylogeny between encapsulated and non-encapsulated Trichinella species. The similarity of the amino acid sequences of trypsin of T. pseudospiralis and T. spiralis was only 71.4%. The encapsulated and non-encapsulated Trichinella species belong to two different clades in the phylogenetic relationship. It may be that TsTryp has more common antigenic epitopes among encapsulated species (T. nativa, T. britovi and T. nelsoni) and fewer common antigenic epitopes in non-encapsulated species (T. pseudospiralis). As such, TsTryp might be specific antigen of the encapsulated Trichinella species. The findings of this study indicated that the detection time of specific IgG and IgM in infected mice could be advanced by 2 days using rTsTryp; namely, rTsTryp detected specific antibodies 2 days earlier than ES antigens, thereby demonstrating that rTsTryp is superior to the ES antigens for the early diagnosis of Trichinella infection. Additionally, only one 49.1-kDa band of ES antigens was recognized by infection murine serum at 10 dpi, which differs from the three bands of ML ES antigens (40–70 kDa) usually recognized by infection serum [80]. It is interesting that on the rTsTryp-western blot, anti-Trichinella IgG was also detected 2 days earlier than the ES antigens.
Although the sensitivity of rTsTryp-ELISA for detecting Trichinella-specific IgG antibody in serum from patients with early trichinellosis at 19 dpi was higher than that of ES antigens, there was no statistical significance between two antigens, likely due to an insufficient number of samples from these patients. Importantly, the specificity of rTsTryp was significantly higher than of ES antigens. rTsTryp cross-reacted with only one serum sample collected from each of a patient with clonorchiasis, cysticercosis or echinococcosis. The sensitivity and specificity of rTsTryp for detecting Trichinella-specific IgG antibody in human serum was similar to that of other recombinant antigens of serine protease family from T. spiralis (rTs31, rTsSP and rTsEla) [8, 31, 37] and cystatin-like protein (rCLP) [81]. It was not possible to validate false-positive human serum in the rTsTryp-ELISA test by western blot due to an insufficient number of human serum samples available in our laboratory. Moreover, anti-Trichinella IgM levels in serum from trichinellosis patients was not tested due to long-term sample storage logistics. Our results demonstrated that the sensitivity of rTsTryp and ES antigens for detecting Trichinella-specific IgG in human serum was not statistically different, but that the specificity of rTsTryp was significantly higher that of the ES antigens. Therefore, rTsTryp could be regarded as an alternative diagnostic antigen for immunodiagnosis of early T. spiralis infection. However, the sensitivity, specificity and reproducibility of rTsTryp needs to be further evaluated on a larger sample size of early trichinellosis patients and patients with other helminthic infections and diseases.
The ES antigens-ELISA is the serological test of choice for detecting anti-Trichinella IgG in domestic swine due the simplicity of the text as well as low costs and fast detection. Previous studies have shown that ES-ELISA has a higher sensitivity than the digestion of 1 g of muscle sample in pigs with low larval burdens (i.e., < 3 larvae per gram, l pg) [82]. The infection level (inoculation dose) of T. spiralis ML in pigs is closely related to the detectable time of serum specific antibodies. In pigs experimentally infected with 25–10,000 larvae, anti-Trichinella IgG antibodies were detected at 25–60 dpi, respectively, indicating that the humoral immune response was inoculation dose dependent [83]. However, no direct correlation between antibody level and the ultimate larval burdens in muscle tissues in serologically positive pigs by ELISA has been found [12]. In a previous study, serum Trichinella-specific IgG in pigs inoculated with 300 larvae was first detected at 30 dpi by ES antigens-ELISA, which was also verified using western blot [77]. In another study, recombinant T. spiralis Serpin was prepared and used to test T. spiralis-infected pig serum; the results showed that at 6 weeks after infection, 45 serum samples obtained from 34 pigs infected with various doses (1000, 5000, 10,000 and 30,400 larvae) exhibited 100% positivity in both ES-ELISA and Serpin ELISA [28]. Our results demonstrated that anti-Trichinella IgG positivity was 100% at 70 dpi in pigs infected with 5000 larvae using rTsTryp-ELISA and ES-ELISA. It was not possible to assay early swine infection sera by the two antigens in the present study. Therefore, from a veterinary practical point of view, early swine infection serum collected at various time points post-infection should be tested using rTsTryp-ELISA in future studies.
Trichinella spiralis contains phenotypically distinct developmental stages throughout its life-cycle (ML, IIL, AW and NBL). The IIL molt 4 times and grow into the AW within 10–30 hpi. Cuticle replacement occurs in each molting process and in larvae emerging from the cuticle of the previous stage [40]. The outer cuticle of each T. spiralis stage expresses different protein molecules during larval growth and the molting process, so each worm stage has its stage-specific antigens [25, 84, 85]. Therefore, at the same time as ensuring specificity, the preparation and mixed use of two or three stage-specific recombinant antigens, namely recombinant stage-specific antigens from early intestinal stages (IIL1-4 or AW), will greatly increase the sensitivity of detecting specific antibodies and enable the development of a more accurate early serodiagnosis of Trichinella infection [13].
In conclusion, TsTryp was present at various T. spiralis developmental stages, with significantly higher levels expressed at the IIL and adult stages, and was primarily distributed in the cuticle and stichosome of this nematode. rTsTryp is an ES antigen with good immunogenicity. Compared with the widely used ML ES antigens, rTsTryp has a similar sensitivity but higher specificity for detecting anti-Trichinella IgG. Therefore, rTsTryp is a valuable tool to detect serum specific antibodies against Trichinella, and it may be a promising alternative antigen for the serodiagnosis of Trichinella infection.
Availability of data and materials
The data supporting the conclusion of this research have been included within the article.
Abbreviations
- AW:
-
Adult worms
- dpi:
-
Days post-infection
- ECL:
-
Enhanced chemiluminescence
- ELISA:
-
Enzyme-linked immunosorbent assay
- ES:
-
Excretory/secretory
- hpi:
-
Hours post-infection
- ICT:
-
International Commission on Trichinellosis
- IFT:
-
Immunofluorescence test
- IIL:
-
Intestinal infectious larvae
- ML:
-
Muscle larvae
- NBL:
-
Newborn larvae
- NJ:
-
Neighbor-joining
- PBST:
-
Phosphate-buffered saline containing 0.05% Tween 20
- pI:
-
Isoelectric point
- (r)TsTryp:
-
(Recombinant) T. spiralis trypsin
- TBST:
-
Tris-buffer saline containing 0.05% Tween 20
References
Murrell KD, Pozio E. Worldwide occurrence and impact of human trichinellosis, 1986–2009. Emerg Infect Dis. 2011;17:2194–202.
Rostami A, Gamble HR, Dupouy-Camet J, Khazan H, Bruschi F. Meat sources of infection for outbreaks of human trichinellosis. Food Microbiol. 2017;64:65–71.
European Food Safety Authority, European Centre for Disease Prevention and Control. The European Union One Health 2019 Zoonoses report. EFSA J. 2021;19:e06406.
Yera H, Bory S, Khieu V, Caron Y. Human trichinellosis in Southeast Asia, 2001–2021. Food Waterborne Parasitol. 2022;28:e00171.
Zhang XZ, Wang ZQ, Cui J. Epidemiology of trichinellosis in the People’s Republic of China during 2009–2020. Acta Trop. 2022;229:106388.
Vasilev S, Mitic I, Mirilovic M, Plavsa D, Milakara E, Plavsic B, et al. Trichinella infection in Serbia from 2011 to 2020: a success story in the field of One Health. Epidemiol Infect. 2023;151:e20.
Despommier DD. How does Trichinella spiralis make itself at home? Parasitol Today. 1998;14:318–23.
Cui J, Wang L, Sun GG, Liu LN, Zhang SB, Liu RD, et al. Characterization of a Trichinella spiralis 31 kDa protein and its potential application for the serodiagnosis of trichinellosis. Acta Trop. 2015;142:57–63.
Dupouy-Camet J, Kociecka W, Bruschi F, Bolas-Fernandez F, Pozio E. Opinion on the diagnosis and treatment of human trichinellosis. Expert Opin Pharmacother. 2002;3:1117–30.
Gottstein B, Pozio E, Nockler K. Epidemiology, diagnosis, treatment, and control of trichinellosis. Clin Microbiol Rev. 2009;22:127–45.
Gamble HR, Pozio E, Bruschi F, Nockler K, Kapel CM, Gajadhar AA. International Commission on Trichinellosis: recommendations on the use of serological tests for the detection of Trichinella infection in animals and man. Parasite. 2004;11:3–13.
Bruschi F, Gomez-Morales MA, Hill DE. International Commission on Trichinellosis: Recommendations on the use of serological tests for the detection of Trichinella infection in animals and humans. Food Waterborne Parasitol. 2019;14:e00032.
Gomez-Morales MA, Cherchi S, Ludovisi A. Serological testing for Trichinella infection in animals and man: current status and opportunities for advancements. Food Waterborne Parasitol. 2022;27:e00165.
Dupouy-Camet J, Raffetin A, Rosca EC, Yera H. Clinical picture and diagnosis of human trichinellosis. In: Bruschi F, editor. Trichinella and trichinellosis. Amsterdam: Academic Press; 2021. p. 333–52.
Liu LN, Jing FJ, Cui J, Fu GY, Wang ZQ. Detection of circulating antigen in serum of mice infected with Trichinella spiralis by an IgY-IgM mAb sandwich ELISA. Exp Parasitol. 2013;133:150–5.
Gomez-Morales MA, Ludovisi A, Amati M, Cherchi S, Pezzotti P, Pozio E. Validation of an ezyme-linked immunosorbent assay for diagnosis of human trichinellosis. Clin Vaccine Immunol. 2008;15:1723–9.
Liu JY, Zhang NZ, Li WH, Li L, Yan HB, Qu ZG, et al. Proteomic analysis of differentially expressed proteins in the three developmental stages of Trichinella spiralis. Vet Parasitol. 2016;231:32–8.
Wang ZQ, Shi YL, Liu RD, Jiang P, Guan YY, Chen YD, et al. New insights on serodiagnosis of trichinellosis during window period: early diagnostic antigens from Trichinella spiralis intestinal worms. Infect Dis Poverty. 2017;6:41.
Xu DM, Wen H, Wang LA, Hu CX, Qi X, Sun GG, et al. Identification of early diagnostic antigens in soluble proteins of Trichinella spiralis adult worms by Western blot. Trop Biomed. 2017;34:191–8.
Yang J, Pan W, Sun X, Zhao X, Yuan G, Sun Q, et al. Immunoproteomic profile of Trichinella spiralis adult worm proteins recognized by early infection sera. Parasit Vectors. 2015;8:20.
Tang B, Liu M, Wang L, Yu S, Shi H, Boireau P, et al. Characterisation of a high-frequency gene encoding a strongly antigenic cystatin-like protein from Trichinella spiralis at its early invasion stage. Parasit Vectors. 2015;8:78.
Zocevic A, Lacour SA, Mace P, Giovani B, Grasset-Chevillot A, Vallee I, et al. Primary characterization and assessment of a T. spiralis antigen for the detection of Trichinella infection in pigs. Vet Parasitol. 2014;205:558–67.
Sun GG, Liu RD, Wang ZQ, Jiang P, Wang L, Liu XL, et al. New diagnostic antigens for early trichinellosis: the excretory-secretory antigens of Trichinella spiralis intestinal infective larvae. Parasitol Res. 2015;114:4637–44.
Sun GG, Wang ZQ, Liu CY, Jiang P, Liu RD, Wen H, et al. Early serodiagnosis of trichinellosis by ELISA using excretory-secretory antigens of Trichinella spiralis adult worms. Parasit Vectors. 2015;8:484.
Liu RD, Cui J, Liu XL, Jiang P, Sun GG, Zhang X, et al. Comparative proteomic analysis of surface proteins of Trichinella spiralis muscle larvae and intestinal infective larvae. Acta Trop. 2015;150:79–86.
Wang ZQ, Liu RD, Sun GG, Song YY, Jiang P, Zhang X, et al. Proteomic Analysis of Trichinella spiralis adult worm excretory-secretory proteins recognized by sera of patients with early trichinellosis. Front Microbiol. 2017;8:986.
Wu Z, Nagano I, Takahashi Y, Maekawa Y. Practical methods for collecting Trichinella parasites and their excretory-secretory products. Parasitol Int. 2016;65:591–5.
Lobanov VA, Konecsni KA, Purves RW, Scandrett WB. Performance of indirect enzyme-linked immunosorbent assay using Trichinella spiralis-derived Serpin as antigen for the detection of exposure to Trichinella spp. in swine. Vet Parasitol. 2022;309:109744.
Mahannop P, Setasuban P, Morakote N, Tapchaisri P, Chaicumpa W. Immunodiagnosis of human trichinellosis and identification of specific antigen for Trichinella spiralis. Int J Parasitol. 1995;25:87–94.
Jung D, Teifke JP, Karger A, Michael K, Venz S, Wittmann W, et al. Evaluation of baculovirus-derived recombinant 53-kDa protein of Trichinella spiralis for detection of Trichinella-specific antibodies in domestic pigs by ELISA. Parasitol Res. 2007;100:429–37.
Hu CX, Jiang P, Yue X, Zeng J, Zhang XZ, Song YY, et al. Molecular characterization of a Trichinella spiralis elastase-1 and its potential as a diagnostic antigen for trichinellosis. Parasit Vectors. 2020;13:97.
Grzelak S, Bien-Kalinowska J, Stachyra A. Trichinella britovi recombinant proteins produced in Pichia pastoris expression system for specific IgG antibody detection in the sera of mice and pigs infected with Trichinella spp. Exp Parasitol. 2022;242:108386.
Liu RD, Jiang P, Wen H, Duan JY, Wang LA, Li JF, et al. Screening and characterization of early diagnostic antigens in excretory-secretory proteins from Trichinella spiralis intestinal infective larvae by immunoproteomics. Parasitol Res. 2016;115:615–22.
Mitreva M, Jasmer DP, Zarlenga DS, Wang ZY, Abubucker S, Martin J, et al. The draft genome of the parasitic nematode Trichinella spiralis. Nat Genet. 2011;43:228–74.
Bai SJ, Han LL, Liu RD, Long SR, Zhang X, Cui J, et al. Oral vaccination of mice with attenuated Salmonella encoding Trichinella spiralis calreticulin and serine protease 1.1 confers protective immunity in BALB/c mice. PLoS Negl Trop Dis. 2022;16:e0010929.
Wang ZQ, Cui J, Shen LJ. The epidemiology of animal trichinellosis in China. Vet J. 2007;173:391–8.
Sun GG, Song YY, Jiang P, Ren HN, Yan SW, Han Y, et al. Characterization of a Trichinella spiralis putative serine protease Study of its potential as sero-diagnostic tool. Plos Neglect Trop Dis. 2018;12:e0006485.
Gajadhar AA, Noeckler K, Boireau P, Rossi P, Scandrett B, Gamble HR. International Commission on Trichinellosis: recommendations for quality assurance in digestion testing programs for Trichinella. Food Waterborne Parasitol. 2019;16:e00059.
Sun GG, Ren HN, Liu RD, Song YY, Qi X, Hu CX, et al. Molecular characterization of a putative serine protease from Trichinella spiralis and its elicited immune protection. Vet Res. 2018;49:59.
Ren HN, Liu RD, Song YY, Zhuo TX, Guo KX, Zhang Y, et al. Label-free quantitative proteomic analysis of molting-related proteins of Trichinella spiralis intestinal infective larvae. Vet Res. 2019;50:1–14.
Yue WW, Yan SW, Zhang R, Cheng YK, Liu RD, Long SR, et al. Characterization of a novel pyruvate kinase from Trichinella spiralis and its participation in sugar metabolism, larval molting and development. Plos Neglect Trop Dis. 2022;16:e0010881.
Ren HN, Guo KX, Zhang Y, Sun GG, Liu RD, Jiang P, et al. Molecular characterization of a 31 kDa protein from Trichinella spiralis and its induced immune protection in BALB/c mice. Parasit Vectors. 2018;11:1–12.
Hu CX, Zeng J, Hao HN, Xu YXY, Liu F, Liu RD, et al. Biological properties and roles of a Trichinella spiralis inorganic pyrophosphatase in molting and developmental process of intestinal larval stages. Vet Res. 2021;52:1–16.
Hu YY, Zhang R, Yan SW, Yue WW, Zhang JH, Liu RD, et al. Characterization of a novel cysteine protease in Trichinella spiralis and its role in larval intrusion, development and fecundity. Vet Res. 2021;52:113.
Yue X, Sun XY, Liu F, Hu CX, Bai Y, Da Yang Q, et al. Molecular characterization of a Trichinella spiralis serine proteinase. Vet Res. 2020;51:125.
Ren HN, Bai SJ, Wang Z, Han LL, Yan SW, Jiang P, et al. A metalloproteinase Tsdpy31 from Trichinella spiralis participates in larval molting and development. Int J Biol Macromol. 2021;192:883–94.
Hao HN, Song YY, Ma KN, Wang BN, Long SR, Liu RD, et al. A novel C-type lectin from Trichinella spiralis mediates larval invasion of host intestinal epithelial cells. Vet Res. 2022;53:85.
Xu J, Liu RD, Bai SJ, Hao HN, Yue WW, Xu YXY, et al. Molecular characterization of a Trichinella spiralis aspartic protease and its facilitation role in larval invasion of host intestinal epithelial cells. Plos Neglect Trop Dis. 2020;14:e0008269.
Hu CX, Xu YXY, Hao HN, Liu RD, Jiang P, Long SR, et al. Oral vaccination with recombinant Lactobacillus plantarum encoding Trichinella spiralis inorganic pyrophosphatase elicited a protective immunity in BALB/c mice. Plos Neglect Trop Dis. 2021;15:e0009865.
Bai Y, Ma KN, Sun XY, Liu RD, Long SR, Jiang P, et al. Molecular characterization of a novel cathepsin L from Trichinella spiralis and its participation in invasion, development and reproduction. Acta Trop. 2021;224:106112.
Lei JJ, Hu YY, Liu F, Yan SW, Liu RD, Long SR, et al. Molecular cloning and characterization of a novel peptidase from Trichinella spiralis and protective immunity elicited by the peptidase in BALB/c mice. Vet Res. 2020;51:111.
Guo KX, Bai Y, Ren HN, Sun XY, Song YY, Liu RD, et al. Characterization of a Trichinella spiralis aminopeptidase and its participation in invasion, development and fecundity. Vet Res. 2020;51:78.
Zeng J, Zhang R, Ma KN, Han LL, Yan SW, Liu RD, et al. Characterization of a novel aminopeptidase P from Trichinella spiralis and its participation in the intrusion of intestinal epithelial cells. Exp Parasitol. 2022;242:108376.
Song YY, Lu QQ, Han LL, Yan SW, Zhang XZ, Liu RD, et al. Proteases secreted by Trichinella spiralis intestinal infective larvae damage the junctions of the intestinal epithelial cell monolayer and mediate larval invasion. Vet Res. 2022;53:19.
Liu CY, Ren HN, Song YY, Sun GG, Liu RD, Jiang P, et al. Characterization of a putative glutathione S-transferase of the parasitic nematode Trichinella spiralis. Exp Parasitol. 2018;187:59–66.
Yan SW, Hu YY, Song YY, Ren HN, Shen JM, Liu RD, et al. Characterization of a Trichinella spiralis cathepsin X and its promotion for the larval invasion of mouse intestinal epithelial cells. Vet Parasitol. 2021;297:109160.
Yang F, Yang DQ, Song YY, Guo KX, Li YL, Long SR, et al. In vitro silencing of a serine protease inhibitor suppresses Trichinella spiralis invasion, development, and fecundity. Parasitol Res. 2019;118:2247–55.
Li JF, Guo KX, Qi X, Lei JJ, Han Y, Yan SW, et al. Protective immunity against Trichinella spiralis in mice elicited by oral vaccination with attenuated Salmonella-delivered TsSP1.2 DNA. Vet Res. 2018;49:1–12.
Qi X, Han Y, Jiang P, Yue X, Ren HN, Sun GG, et al. Oral vaccination with Trichinella spiralis DNase II DNA vaccine delivered by attenuated Salmonella induces a protective immunity in BALB/c mice. Vet Res. 2018;49:1–12.
Liu CY, Song YY, Ren HN, Sun GG, Liu RD, Jiang P, et al. Cloning and expression of a Trichinella spiralis putative glutathione S-transferase and its elicited protective immunity against challenge infections. Parasit Vectors. 2017;10:1–9.
Zhang XZ, Yue WW, Bai SJ, Hao HN, Song YY, Long SR, et al. Oral immunization with attenuated Salmonella encoding an elastase elicits protective immunity against Trichinella spiralis infection. Acta Trop. 2022;226:106263.
Liu LN, Wang ZQ, Zhang X, Jiang P, Qi X, Liu RD, et al. Characterization of Spirometra erinaceieuropaei plerocercoid cysteine protease and potential application for serodiagnosis of sparganosis. Plos Neglect Trop Dis. 2015;9:e0003807.
Sun GG, Lei JJ, Guo KX, Liu RD, Long SR, Zhang X, et al. Primary assessment of a T. spiralis putative serine protease for early serological detection of experimental trichinellosis. Trop Biomed. 2019;36:792–802.
Yang Y, Wen YJ, Cai YN, Vallee I, Boireau P, Liu MY, et al. Serine proteases of parasitic helminths. Korean J Parasitol. 2015;53:1–11.
Nagano I, Wu ZL, Nakada T, Boonmars T, Takahashi Y. Molecular cloning and characterization of a serine proteinase gene of Trichinella spiralis. J Parasitol. 2003;89:92–8.
Grzelak S, Stachyra A, Stefaniak J, Mrowka K, Moskwa B, Bien-Kalinowska J. Immunoproteomic analysis of Trichinella spiralis and Trichinella britovi excretory-secretory muscle larvae proteins recognized by sera from humans infected with Trichinella. PLoS ONE. 2020;15:e0241918.
Liu F, Song YY, Zhang R, Liu RD, Jiang P, Cui J, et al. Cloning and expression of a new Trichinella spiralis serine protease and its rRole in invading host intestinal epithelium. Iran J Parasitol. 2022;17:375–84.
Bermudez-Cruz RM, Fonseca-Linan R, Grijalva-Contreras LE, Mendoza-Hernandez G, Ortega-Pierres MG. Proteomic analysis and immunodetection of antigens from early developmental stages of Trichinella spiralis. Vet Parasitol. 2016;231:22–31.
Gao H, Tang B, Bai X, Wang LB, Wu XP, Shi HN, et al. Characterization of an antigenic serine protease in the Trichinella spiralis adult. Exp Parasitol. 2018;195:8–18.
Ren HJ, Cui J, Yang W, Liu RD, Wang ZQ. Identification of differentially expressed genes of Trichinella spiralis larvae after exposure to host intestine milieu. PLoS ONE. 2013;8:e67570.
Trap C, Fu BQ, Le Guerhier F, Liu MY, Le Rhun D, Romand T, et al. Cloning and analysis of a cDNA encoding a putative serine protease comprising two trypsin-like domains of Trichinella spiralis. Parasitol Res. 2006;98:288–94.
Bien J, Cabaj W, Moskwa B. Proteomic analysis of potential immunoreactive proteins from muscle larvae and adult worms of Trichinella spiralis in experimentally infected pigs. Folia Parasitol. 2015;62:022.
Robinson MW, Connolly B. Proteomic analysis of the excretory-secretory proteins of the Trichinella spiralis L1 larva, a nematode parasite of skeletal muscle. Proteomics. 2005;5:4525–32.
Wang B, Wang ZQ, Jin J, Ren HJ, Liu LN, Cui J. Cloning, expression and characterization of a Trichinella spiralis serine protease gene encoding a 35.5 kDa protein. Exp Parasitol. 2013;134:148–54.
Frey CF, Schuppers ME, Nockler K, Marinculic A, Pozio E, Kihm U, et al. Validation of a western blot for the detection of anti-Trichinella spp. antibodies in domestic pigs. Parasitol Res. 2009;104:1269–77.
Cuttell L, Gomez-Morales MA, Cookson B, Adams PJ, Reid SA, Vanderlinde PB, et al. Evaluation of ELISA coupled with Western blot as a surveillance tool for Trichinella infection in wild boar (Sus scrofa). Vet Parasitol. 2014;199:179–90.
Gondek M, Bien J, Nowakowski Z. Use of ELISA and Western blot for serological detection of antibodies to E-S antigens of Trichinella spiralis muscle larvae in sera of swine experimentally infected with Trichinella spiralis. Vet Immunol Immunopathol. 2018;203:13–20.
Gomez-Morales MA, Ludovisi A, Amati M, Blaga R, Zivojinovic M, Ribicich M, et al. A distinctive western blot pattern to recognize Trichinella infections in humans and pigs. Int J Parasitol. 2012;42:1017–23.
Serrano FJ, Perez-Martin JE, Carron A, Navarrete I. Comparison of IgM, IgG1 and IgG2 responses to Trichinella spiralis and Trichinella britovi in swine. Parasite. 2001;8:S133–5.
Supcharoengoon U, Reamtong O, Dekumyoy P, Watthanakulpanich D, Limpanont Y, Zhiyue L, et al. Evaluation of indirect-ELISA using eluted antigens from Trichinella spiralis muscle larvae for diagnosis of swine trichinellosis. Acta Trop. 2022;235:106644.
Liu Y, Xu N, Li Y, Tang B, Yang H, Gao W, et al. Recombinant cystatin-like protein-based competition ELISA for Trichinella spiralis antibody test in multihost sera. PLoS Negl Trop Dis. 2021;15:e0009723.
Kapel CM, Gamble HR. Infectivity, persistence, and antibody response to domestic and sylvatic Trichinella spp. in experimentally infected pigs. Int J Parasitol. 2000;30:215–21. https://doi.org/10.1016/s0020-7519(99)00202-7.
Wang N, Bai X, Ding J, Lin J, Zhu F, Luo X, et al. Trichinella infectivity and antibody response in experimentally infected pigs. Vet Parasitol. 2021;297:109111.
Jungery M, Clark NW, Parkhouse RM. A major change in surface antigens during the maturation of newborn larvae of Trichinella spiralis. Mol Biochem Parasitol. 1983;7:101–9.
Ortega-Pierres G, Chayen A, Clark NW, Parkhouse RM. The occurrence of antibodies to hidden and exposed determinants of surface antigens of Trichinella spiralis. Parasitology. 1984;88:359–69.
Acknowledgements
We would like to thank YY Song and SW Yan for their help in animal experiments.
Funding
This research was funded by grants of the National Natural Science Foundation of China (No. No. 81971952, 82172300 and 82372276).
Author information
Authors and Affiliations
Contributions
JC and ZQW designed the research. LLH, QQL, YLL, WWZ, PR, RDL, JC and ZQW conducted the tests. LLH, JC and ZQW wrote the manuscript. LLH collected and analyzed experimental data and prepared the figures. All authors reviewed the manuscript and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study was performed according to National Guidelines for Experimental Animal Welfare (Minister of Science and Technology, People’s Republic of China, 2006). All experimental protocols were authorized by the Life Science Ethics Committee of Zhengzhou University (No. ZZUIRB GZR 2021-0044). Blood samples from patients were only acquired from adults who provided written informed consent.
Consent for publication
Not applicable.
Competing interests
No relevant conflicts of interest to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Multiple alignment of peptide sequences of TsTryp with trypsin of other species/genotypes of Trichinella. The sequence of TsTryp (XM_003381619.1) was aligned with trypsin of other Trichinella species/genotypes. The Clustal W analysis in BioEdit software was used to compare peptide sequences of trypsin within the genus Trichinella. The black background sequence represents the same part of TsTryp sequence with other species, the gray background sequence represents conservative substitutive residues and numbers behind the sequence represent percentage similarity with the TsTryp sequence.
Additional file 2: Figure S2.
Western blot confirmation of one false positive serum of sparganum-infected mice in the ES-ELISA test. A Analysis of ML ES antigens by SDS-PAGE. Lanes: M, Protein marker; 1, muscle larvae ES antigens. B Western blot confirmation of one false ES-ELISA positive serum. Lanes: M, Protein marker; 1, muscle larvae ES antigens recognized using T. spiralis-infected murine serum; 2, muscle larvae ES antigens not recognized using uninfected mouse serum; 3, muscle larvae ES antigens were not identified using sparganum-infected mouse serum which was false positive in ES-ELISA test.
Additional file 3: Figure S3.
Scatter plot of absorbance values at 492 nm of rTsTryp-ELISA (A) and ES antigens-ELISA (B) for testing specific IgG in sera from patients infected with Trichinella and other parasites. Horizontal dashed lines are cutoff values from rTsTryp-ELISA and ES antigens ELISA.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Han, L.L., Lu, Q.Q., Li, Y.L. et al. Application of a recombinant novel trypsin from Trichinella spiralis for serodiagnosis of trichinellosis. Parasites Vectors 17, 9 (2024). https://doi.org/10.1186/s13071-023-06067-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-06067-7