
Abstract
Background
Wolbachia pipientis is an endosymbiont bacterium that induces cytoplasmic incompatibility and inhibits arboviral replication in mosquitoes. This study aimed to assess Wolbachia prevalence and genetic diversity in different mosquito species from Cape Verde.
Methods
Mosquitoes were collected on six islands of Cape Verde and identified to species using morphological keys and PCR-based assays. Wolbachia was detected by amplifying a fragment of the surface protein gene (wsp). Multilocus sequence typing (MLST) was performed with five housekeeping genes (coxA, gatB, ftsZ, hcpA, and fbpA) and the wsp hypervariable region (HVR) for strain identification. Identification of wPip groups (wPip-I to wPip-V) was performed using PCR–restriction fragment length polymorphism (RFLP) assay on the ankyrin domain gene pk1.
Results
Nine mosquito species were collected, including the major vectors Aedes aegypti, Anopheles arabiensis, Culex pipiens sensu stricto, and Culex quinquefasciatus. Wolbachia was only detected in Cx. pipiens s.s. (100% prevalence), Cx. quinquefasciatus (98.3%), Cx. pipiens/quinquefasciatus hybrids (100%), and Culex tigripes (100%). Based on the results of MLST and wsp hypervariable region typing, Wolbachia from the Cx. pipiens complex was assigned to sequence type 9, wPip clade, and supergroup B. PCR/RFLP analysis revealed three wPip groups in Cape Verde, namely wPip-II, wPip-III, and wPip-IV. wPip-IV was the most prevalent, while wPip-II and wPip-III were found only on Maio and Fogo islands. Wolbachia detected in Cx. tigripes belongs to supergroup B, with no attributed MLST profile, indicating a new strain of Wolbachia in this mosquito species.
Conclusions
A high prevalence and diversity of Wolbachia was found in species from the Cx. pipiens complex. This diversity may be related to the mosquito's colonization history on the Cape Verde islands. To the best of our knowledge, this is the first study to detect Wolbachia in Cx. tigripes, which may provide an additional opportunity for biocontrol initiatives.
Graphical Abstract

Similar content being viewed by others

Background
Wolbachia pipientis (Alphaproteobacteria, Rickettsiales) is an obligate intracellular gram-negative bacterium and proteobacterial symbiont found in a variety of invertebrates, including insects, crustaceans, arachnids, and filarial nematodes [1]. Currently, the Wolbachia genus is subdivided into 17 supergroups (A–F; H–Q, and S), and most species known belong to supergroups A and B [2].
Wolbachia is transmitted vertically through host eggs and can influence longevity and reproduction, including feminization, parthenogenesis, and incompatibility between the female and male sex cells [3]. The best-known phenotype induced by Wolbachia in arthropods is cytoplasmic incompatibility (CI). It occurs when males harboring Wolbachia are crossed with uninfected females or between individuals infected with incompatible strains [4, 5]. The generally accepted model stipulates that cytoplasmic incompatibility results from a Wolbachia “modification” factor (mod; toxin) in the sperm that blocks early embryogenesis, and a Wolbachia “rescue” factor (resc; antitoxin) produced in the oocyte that allows the diploid zygote to develop if the cross is compatible [6, 7].
Besides cytoplasmic incompatibility, Wolbachia can inhibit viral replication in mosquitoes, including Zika, dengue, West Nile, and chikungunya arboviruses in Aedes aegypti [8, 9]. Other studies also suggest inhibition of pathogens such as Plasmodium falciparum in Anopheles stephensi and Anopheles gambiae and West Nile virus in Culex quinquefasciatus [1, 10, 11]. These abilities make Wolbachia a promising tool against mosquito-borne diseases and possibly an alternative to conventional vector control programs using insecticides. In fact, the release of males harboring incompatible Wolbachia into target populations has successfully decreased reproduction by sterilization [12, 13]. The release of Ae. aegypti transfected with the Wolbachia wMel strain (derived from Drosophila melanogaster) led to the establishment of Ae. aegypti populations infected with Wolbachia and a proven decrease in dengue incidence in Australia [14] and Malaysia [15].
Cape Verde is threatened by several species of vector mosquitoes, including Ae. aegypti, Anopheles arabiensis, Cx. quinquefasciatus, and Culex pipiens sensu stricto (s.s.) [16]. Integrated vector control strategies are mainly directed against An. arabiensis and Ae. aegypti, using chemical insecticides, diesel, and biological control with Gambusia sp. fish [17]. However, despite control efforts, the country had its first dengue epidemic in 2009, followed by an outbreak of Zika in 2015–2016 [18] and a malaria outbreak in 2017 [19].
There is no data on the genetic diversity of Wolbachia infecting mosquitoes (Diptera: Culicidae) from the Cape Verde islands. This knowledge would be a first step for the design and implementation of programs to suppress mosquito populations through cytoplasmic incompatibility. In this context, the present study aims to detect and genetically characterize Wolbachia in populations of Culicidae from Cape Verde.
Methods
Study area and sample collection
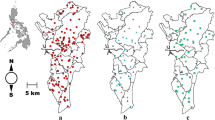
An entomological survey was carried out in Cape Verde between February and June 2021. Larval and adult mosquito samples were collected on six islands (Santiago, Brava, Fogo, Maio, Santo Antão, and Boavista; Fig. 1) using BG-Sentinel and Centers for Disease Control and Prevention (CDC) light traps, dorsal aspirators, dippers, and pipettes. All collection sites were geo-referenced with a portable global positioning system (GPS) device (Garmin eTrex 10).

Map of the North Atlantic region showing the geographic location of the Cape Verde islands. Mosquito samples were collected on the islands of Santo Antão, Boavista, Maio, Santiago, Fogo, and Brava (highlighted in black)
Mosquitoes were identified to species/complex using the Ribeiro et al. [20] identification key and stored individually in microtubes containing silica gel (for adults) or 80% ethanol (for larvae). For genetic analysis, DNA was extracted from single specimens using cetrimonium bromide (CTAB) 2% and proteinase K, according to Weeks et al. [21].
Species of the An. gambiae complex were identified by polymerase chain reaction (PCR) according to Scott et al. [22] using primer sequences described in Table S1 (Additional file 1: Table S1). PCR was performed using 12.5 µl of Xpert TaqPLUS Mastermix (GriSP), 0.1 µM of ME and UN primers, 0.05 µM of GA primer, and 0.15 µM of AR primer, plus 1 µl of DNA template and water to a final volume of 25 µl. Cycling conditions were as follows: one cycle at 95 °C for 5 min, 30 cycles at 94 °C for 30 s, 50 °C for 30 s, and 72 °C for 30 s; and a final cycle of 72 °C for 5 min.
For the Cx. pipiens complex, specimens were identified to species by PCR amplification of acetylcholinesterase-2 (ace-2) gene sequences using primers described by Smith & Fonseca [23] (Additional file 1: Table S1). PCR was performed using 12.5 µl of Xpert TaqPLUS Mastermix (GriSP), 0.4 µM of ACEquin and B1246 primer, 0.2 µM of ACEpip primer, 1 µl of DNA template and water to a final volume of 25 µl. Cycling conditions were performed as follows: one cycle at 94 °C for 5 min, 35 cycles at 94 °C for 30 s, 55 °C for 30 s, 72 °C for 1 min, and one cycle at 72 °C for 5 min.
Whenever necessary, morphological identification of species other than the above was supported with the sequencing of a 710-base-pair (bp) fragment of cytochrome c oxidase subunit 1 mitochondrial gene (COI) with primers LCOI1490_F1 and HCOI2198_R1 (Additional file 1: Table S1) according to Folmer et al. [24]. PCR was performed using 1X PCR buffer, 2 mM MgCl2, 0.2 mM dNTPs, 1 U Taq polymerase (Robust HotStart PCR Kit, Roche/Kapa Biosystems), 0.5 µM of each primer, 2 µl of DNA template, and water to a final volume of 20 µl. Cycling conditions were as follows: initial denaturation at 94 °C for 3 min; 40 cycles at 94 °C for 50 s; annealing at 45 °C during 30 s and 72 °C for 1 min; and final elongation at 72 °C for 5 min.
Screening of Wolbachia
Wolbachia detection in mosquito samples was performed by amplifying a 610-bp region of the Wolbachia surface protein gene (wsp) using primers 81F and 691R (Additional file 1: Table S2) described by Zhou et al. [25]. The amplification reaction comprised 12.5 µl of Xpert TaqPLUS Mastermix (GriSP), 0.4 µM of each primer, 1 µl of DNA template, and water to a final volume of 25 µl. Cycling conditions were as follows: one cycle at 95 °C for 3 min, 35 cycles at 95 °C for 1 min, 55 °C for 1 min, 72 °C for 1 min, and one cycle at 72 °C for 10 min.
All PCR products from the assays described above were analyzed by electrophoresis on a 1.5% agarose gel stained with GreenSafe Premium (NZYTech).
Wolbachia multilocus sequence typing (MLST) and wsp typing
Wolbachia genotyping was performed through amplification and sequencing of five MLST loci (gatB, coxA, hcpA, ftsZ, fbpA) and the wsp hypervariable region [26, 27]. The primer pairs for each locus and the size of amplified products are shown in supplemental materials (Additional file 1: Table S3).
PCR for each locus was performed using 1X PCR buffer, 0.2 mM dNTPs, 1.5 mM MgCl2, 0.5U Taq polymerase (Robust HotStart PCR Kit, Roche/Kapa Biosystems), 1 µM of each primer, 2 µl of DNA template, and water to a final volume of 40 µl. Cycling conditions were as follows: initial denaturation at 94 °C for 2 min; 37 cycles at 94 °C for 30 s, annealing at 54 °C (for hcpA, gatB, ftsZ, and coxA), and 59 °C (fbpA and wsp) for 45 s, and 72 °C for 90 s; and final elongation at 72 °C for 10 min.
Five microliters of PCR product from each locus was used in electrophoresis to confirm amplification. The remaining 35 µl was purified using an Exo/SAP Go PCR purification kit (GriSP) and sent for direct DNA sequencing at STAB Vida (Oeiras, Portugal) using forward and reverse primers.
Wolbachia MLST and hypervariable wsp sequences were edited and aligned using BioEdit (version 7.0.9.0). Consensus and concatenated sequences (gatB, coxA, fbpA, ftsZ, hcpA, and wsp hypervariable region [HVR]) were queried in the Wolbachia MLST database (https://pubmlst.org/bigsdb?db=pubmlst_wolbachia_seqdef) for strain characterization. Sequences were also subjected to the nucleotide Basic Local Alignment Search Tool (BLAST) to verify the similarity with deposited sequences in GenBank (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Phylogenetic analysis was conducted using the gamma-distributed Tamura 3-parameter nucleotide substitution model, and a neighbor-joining tree was generated employing 1000 bootstraps in Molecular Evolutionary Genetics Analysis version 11 (MEGA11) [28].
Identification of wPip groups by PCR–RFLP
Identification of wPip groups (wPip-I to wPip-V) was performed using a PCR–restriction fragment length polymorphism (RFLP) assay based on the ankyrin (ANK) Wolbachia marker pk1 [29,30,31]. A PCR that amplifies a 1300-bp fragment of the ANK domain gene (pk1) was performed with primers pk1_For and pk1_Rev (Additional file 1: Table S2) [32]. The reaction components included 10 µl of Xpert TaqPLUS Mastermix (GriSP), 0.4 µM of each primer, 2 µl of DNA template, and water to a final volume of 20 µl. Cycling conditions were as follows: one cycle at 94ºC for 5 min; 35 cycles at 94 °C for 30 s, 52ºC for 30 s, and 72 °C for 90 s; and a final cycle of 72 °C for 5 min. PCR product was analyzed by electrophoresis on a 2% agarose gel stained with GreenSafe Premium (NZYTech).
The pk1 PCR product was digested with restriction enzymes TaqαI and PstI to identify different wPip groups [29]. Digestion with TaqαI was performed with the following reaction mixture: 2 µl of Buffer C (NZYTech), 10 µl of the PCR product, 18 µl of water, and 2 µl TaqαI enzyme (NZYTech) at 10U/µl. The mixture was placed in a thermal cycler at 65ºC for 90 min. The reaction was stopped by adding 0.02 mM of ethylenediaminetetraacetic acid (EDTA) (pH = 8) to each tube, and the digestion product was visualized by electrophoresis on a 2% agarose gel. Each allele (wPip group) was detected according to the size of the resulting fragments: allele “a” or “e” (wPip-I or wPip-V; 991, 251, 107 bp); “b” (wPip-III; 669, 665 bp); “c” (wPip-II; 851, 498 bp); “d” (wPip-IV; 497, 251, 107 bp) [29].
If alleles "a" or "e" (wPip-I or wPip-V) were present, the two were differentiated by digesting the pk1 PCR product with the PstI restriction enzyme. For this purpose, a reaction mixture was prepared with 2 µl of Buffer A (NZYTech), 12 µl pk1 PCR product, 1 µl PstI enzyme (NZYTech) at 10U/µl, and 5 µl of water. The mixture was incubated at 37 °C for 1 h, and the reaction stopped by incubating at 80 °C for 20 min. Digested DNA fragments were separated by electrophoresis on a 2% agarose gel. wPip alleles resulting from pstI digestion included “a” (wPip- I; 903, 303, 141 bp) and “e” (wPip-V; 903, 430 bp) [29, 30].
Sequencing of pk1 PCR products was performed to confirm the RFLP profile. For this purpose, the pk1 PCR product was purified as described above for the MLST and sent for direct sequencing using reverse and forward primers. Sequences were subjected to the nucleotide BLAST, and phylogenetic analysis was performed using the gamma-distributed Tamura 3-parameter nucleotide substitution model, and a neighbor-joining tree was generated using 1000 bootstraps in MEGA software version 11.0.11.
Results
Mosquito species identification
A total of 1648 mosquitoes (303 larvae and 1345 adults) were collected (Additional file 2: Table S4 for details). Species identification by morphological characters revealed the presence of Ae. aegypti (n = 663, 40.2%), Aedes caspius (n = 39, 2.4%), An. gambiae sensu lato (s.l.) (n = 49, 3.0%), Anopheles pretoriensis (n = 275, 16.7%), Cx. pipiens s.l. (n = 584, 35.4%), Culex thalassius (n = 7, 0.4%), Culex tigripes (n = 3, 0.2%), and Culiseta longiareolata (n = 28, 1.7%).
Ribosomal DNA PCR for identifying species of the An. gambiae complex revealed that all collected specimens from this complex belonged to An. arabiensis. For the Cx. pipiens complex, specimens were identified by ace-2 PCR as Cx. pipiens s.s. (n = 10, 1.7%), Cx. quinquefasciatus (n = 545, 93.3%), and Cx. pipiens/Cx. quinquefasciatus hybrids (n = 29, 5.0%).
Screening of Wolbachia
The wsp fragment was amplified only in Cx. pipiens s.s. (10/10 = 100% prevalence), Cx. quinquefasciatus (536/545 = 98.3%), Cx. pipiens/Cx. quinquefasciatus hybrids (29/29 = 100%), and Cx. tigripes (3/3 = 100%). The remaining species were negative for Wolbachia.
Wolbachia MLST and wsp typing
We analyzed 80 mosquitoes that were positive for wsp for Wolbachia MLST and wsp typing. Allelic profiles resulting from MLST loci and the wsp hypervariable region sequencing revealed that Wolbachia from Cx. pipiens s.s., Cx. quinquefasciatus, and Cx. pipiens/Cx. quinquefasciatus hybrids belong to sequence type 9, wPip clade, and supergroup B Wolbachia (Table 1). The same result was obtained from phylogenetic analysis using concatenated sequences of MLST loci (coxA, gatB, ftsZ, fbpA, hcpA) and the wsp hypervariable region (Fig. 2).

Phylogenetic tree generated from concatenated sequences of MLST loci (coxA, gatB, ftsZ, fbpA, hcpA) and the wsp hypervariable region. Numbers on branches indicate percentage bootstrap support (1000 replicates). Reference sequences were obtained from the Wolbachia MLST database and are marked by full circles. Each Wolbachia supergroup is marked with a different color: yellow, supergroup B; black, supergroup A; red, supergroup D; and green, supergroup F. The scale bar indicates the number of substitutions
For Cx. tigripes, the allelic profile obtained was unavailable in the MLST database, thus not allowing the determination of a sequence type. However, the phylogenetic analysis indicates that Wolbachia from Cx. tigripes also belongs to supergroup B but integrates a distinct clade from wPip (Fig. 2).
wPip groups and their distribution in the archipelago
Results from pk1 PCR–RFLP showed the occurrence of three different wPip groups in Cape Verde, namely wPip-IV (88.9%), wPip-II (7.4%), and wPip-III (3.7%) (Table 2). The wPip-IV group was detected in Cx. quinquefasciatus from five islands (Santiago, Brava, Santo Antão, Maio and Boavista) and in Cx. pipiens s.s. from Santo Antão. The wPip-II group was detected only in Cx. pipiens s.s. from Maio, while wPip-III was found exclusively in Cx. quinquefasciatus and Cx. pipiens/quinquefasciatus hybrids from the island of Fogo (Table 2).
Sequencing of pk1 PCR products confirmed the observed RFLP profiles and similarity with pk1 sequences deposited in GenBank (Fig. 3).

Phylogenetic tree generated from pk1 sequences by Bayesian analysis. Known wPip group pk1 sequences are marked by full circles. Numbers on branches indicate percentage bootstrap support (1000 replicates). The scale bar indicates the number of substitutions
Discussion
Wolbachia has garnered substantial attention for its ability to control diseases transmitted by mosquitoes. This study represents the first assessment of Wolbachia's prevalence and genetic diversity in mosquitoes from Cape Verde. Our objective is to expand knowledge of this bacterium through our findings and illustrate its potential for controlling mosquito-borne diseases in the archipelago.
The MLST and wsp typing results revealed that Wolbachia from Cx. pipiens and Cx. quinquefasciatus and their hybrids belong to the wPip clade and share a monophyletic origin within Wolbachia group B. The same results were obtained by Atyame et al. [33] and Dumas et al. [30] when studying Wolbachia genetic diversity from Cx. pipiens s.l. populations originating from different regions of the world. According to the authors, these findings suggest that wPip strains comprise a recent clade of the Wolbachia supergroup B [30, 33].
The analysis of the fast-evolving pk1 gene revealed further variation within the wPip strain, indicating the presence of wPip-II, wPip-III, and wPip-IV groups in Cape Verde. The occurrence of different wPip groups suggests multiple introduction events into the archipelago. In the past, Cape Verde was a maritime hub between Europe and mainland Africa, and the intense movement of ships may explain the diversity of wPip found on the islands. This result contrasts with that of the southwestern Indian Ocean islands, in which Wolbachia infecting Cx. quinquefasciatus all belonged to the wPip-I group [13]. It is noteworthy that wPip-I was the only group found in mainland Sub-Saharan Africa, South America, and Southeast Asia, whereas only wPip-III was detected in North America [30, 33]. Europe shows the highest diversity, with all five groups of the wPip clade being found in this continent [30]. The presence of wPip-II, wPip-III, and wPip-IV groups in Cape Verde islands suggests at least three introduction events of Wolbachia possibly originating from Europe. However, a North American origin for the wPip-III group in Fogo Island cannot be excluded. Interestingly, the differences found in the genetic composition of the wPip clade among islands agree with the genetic structure of the Cx. pipiens complex in Cape Verde. Previous microsatellite-based analysis suggested that Cx. quinquefasciatus from Fogo Island may comprise a genetic ancestry cluster distinct from the other islands [34]. What was previously considered an admixed Cx. quinquefasciatus population in Fogo Island [34] may, in fact, represent a genetically differentiated population originating from a wPip-III group source population.
The absence of the African wPip-I group from Cape Verde Cx. quinquefasciatus is not easily explained. Mainland Africa would be the natural candidate for a source population of wPip-I Cx. quinquefasciatus that would have colonized the Cape Verdean islands, as suggested for the southwestern Indian Ocean islands [30]. However, Cx. quinquefasciatus was predominantly infected by the wPip-IV group. This result may suggest that Cx. quinquefasciatus from Cape Verde may have derived from a yet to be sampled wPip-IV population of mainland Africa. Another explanation would involve the cytoplasmic transfer of wPip-IV from European Cx. pipiens s.s. to wPip-I Cx. quinquefasciatus via hybridization, followed by the latter's replacement through cytoplasmic incompatibility (CI). High levels of CI have been reported in crosses between wPip-II and wPip-IV, as well as between wPip-III- and wPip-IV-infected mosquitoes [7, 29]. Studies involving experimental crosses would be required to assess CI between wPip-I and wPip-IV and whether this CI would confer an adaptive advantage to wPip-IV-infected mosquitoes.
Wolbachia was not detected in Ae. aegypti from Cape Verde islands, which is consistent with most surveys on this species where no evidence of Wolbachia natural infection was found [35,36,37,38]. The presence of Wolbachia in Ae. aegypti has been reported on only a few occasions, including those from New Mexico, the USA [39], and Kuala Lumpur, Malaysia [40]. However, the possibility of Wolbachia detection in Ae. aegypti being the result of an infection with a Wolbachia-carrying nematode or of environmental contamination during field collections could not be excluded [36]. Wolbachia was also not detected in An. arabiensis and An. pretoriensis from Cape Verde. While this result is in line with most studies that screened for Wolbachia in Anopheles species [41, 42], there have been a few reports on the presence of the endosymbiont in An. gambiae and An. coluzzii from Mali [43], An. gambiae from the Democratic Republic of Congo, and An. coluzzii in Ghana [44]. Shaw et al. [45] concluded that Wolbachia natural Anopheles infections do not induce cytoplasmic incompatibility or sex ratio distortion but show a negative correlation with Plasmodium infection, suggesting that Wolbachia may interfere with malaria transmission.
This study reports for the first time the presence of Wolbachia in Cx. tigripes. Phylogenetic analyses indicate that Wolbachia isolated from this mosquito belongs to supergroup B, with no attributed MLST profile. This result suggests the presence of a new strain of Wolbachia infecting Cx. tigripes in Santiago Island. Culex tigripes is the only predatory mosquito in Cape Verde [46], and on the island of Santiago, its larvae are often found in breeding sites associated with Cx. pipiens s.l. species [47]. Our results exclude environmental contamination by Cx. pipiens s.l. Wolbachia since we detected Wolbachia in both larvae and an adult male of Cx. tigripes (Additional file 2: Table S4). More importantly, the concatenated sequences of the MLST loci and the wsp HVR region clearly showed that the strain detected in Cx. tigripes forms a monophyletic group separate from the wPip clade. Our phylogenetic analyses also exclude contamination with Wolbachia from supergroups D and F, which are generally found in nematodes [2, 48].
The use of Wolbachia-based methods in vector management holds significant promise. The newly detected Wolbachia strain in Cx. tigripes from Cape Verde encourages further research to assess their ability to be firmly established in major vector trans-infected lines, induce cytoplasmic incompatibility, or reduce the ability to transmit pathogens. Proof of these abilities may offer an additional opportunity for biocontrol initiatives.
The incompatible insect technique (IIT), a variation of the sterile insect technique (SIT), can be performed by taking advantage of the wPip-induced cytoplasmic incompatibility. Studies have indicated that Cx. pipiens s.l. mosquitoes infected with identical Wolbachia wPip groups tend to exhibit cytoplasmic compatibility, while crossing between mosquitoes carrying different wPip groups is often incompatible [13, 31]. As a result, our findings regarding the natural occurrence of wPip groups in Cape Verde can provide valuable insights for implementing control programs for Cx. pipiens s.l. in the archipelago.
Experiments conducted in semi-field conditions on La Réunion showed that the mating between local Cx. quinquefasciatus wPip-I females and non-native males carrying the wPip-IV (Istanbul strain) resulted in 100% embryonic mortality [13]. Altinli et al. [29] demonstrated naturally occurring CI patterns between wPip-IV-harboring males and wPip-I- or wPip-II-harboring females in Cx. pipiens s.l. populations from Turkey. These observations reveal that IIT based on wPip-inducing IC could be employed to control Cx. pipiens populations. The same methodology can be implemented in Cape Verde considering the data we gathered on the prevalence and distribution of wPip groups in the archipelago. It would be worthwhile to analyze the pattern of cytoplasmic incompatibility among the different wPip groups in Cape Verde and determine whether an island-specific wPip group could be used to regulate Cx. pipiens s.l. populations on another island. As an alternative, male Cx. pipiens from other regions of the world carrying wPip groups not present in Cape Verde, could be introduced into the archipelago to sterilize local females. It is noteworthy that IIT based on wPip-inducing IC could be a favorable alternative to the costly radiation and genetic manipulation methods, and its implementation would provide a more advantageous solution for low-income nations.
Conclusion
Our study revealed that Wolbachia is widespread in Cx. pipiens s.l. from the Cape Verde islands but absent from other mosquito species except for Cx. tigripes,, where a novel Wolbachia strain was unveiled. The three distinct wPip groups circulating in Cx. pipiens s.l. suggest multiple introduction events in the archipelago, possibly of non-African origin. The finding of a novel Wolbachia strain in Cx. tigripes may provide an additional candidate to be used in biocontrol approaches. Further studies would be required to isolate this new Wolbachia strain to be used in transfection studies with major mosquito vectors in order to assess its potential impact on mosquito fitness and vector competence.
Availability of data and materials
Sequences generated in this study are available in the GenBank database: pk1 sequences (OQ223307-OQ223325); ftsZ (OQ223326-OQ223348); hcpA (OQ223349-OQ223371); fbpA (OQ223372-OQ223394); coxA (OQ225016-OQ225038); gatB (OQ225039-OQ225061); and wsp (OQ236526-OQ236548). All reference sequence accession numbers (GenBank) and MLST database IDs are included in the article.
Abbreviations
- CI:
-
Cytoplasmic incompatibility
- MLST:
-
Multilocus sequence typing
- RFLP:
-
Restriction fragment length polymorphism
- Wsp:
-
Wolbachia surface protein
References
Glaser RL, Meola MA. The native Wolbachia endosymbionts of Drosophila melanogaster and Culex quinquefasciatus increase host resistance to West Nile virus infection. PLoS ONE. 2010;5:e11977.
Kaur R, Shropshire JD, Cross KL, Leigh B, Mansueto AJ, Stewart V, et al. Living in the endosymbiotic world of Wolbachia: a centennial review. Cell Host Microbe. 2021;29:879–93.
Werren JH, Baldo L, Clark ME. Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol. 2008;6:741–51.
Flores HA, O’Neill SL. Controlling vector-borne diseases by releasing modified mosquitoes. Nat Rev Microbiol. 2018;16:508–18.
Duron O, Bernard C, Unal S, Berthomieu A, Berticat C, Weill M. Tracking factors modulating cytoplasmic incompatibilities in the mosquito Culex pipiens. Mol Ecol. 2006;15:3061–71.
Bonneau M, Atyame C, Beji M, Justy F, Cohen-Gonsaud M, Sicard M, et al. Culex pipiens crossing type diversity is governed by an amplified and polymorphic operon of Wolbachia. Nat Commun. 2018;9:1491.
Atyame C, Labbé P, Dumas E, Milesi P, Charlat S, Fort P, et al. Wolbachia divergence and the evolution of cytoplasmic incompatibility in Culex pipiens. PLoS ONE. 2014;9:e87336.
Kittayapong P, Chansang C, Chansang U, Mongkalangoon P, Ninphanomchai S, Limohpasmanee W. Combined sterile insect technique and incompatible insect technique: The first proof-of-concept to suppress Aedes aegypti vector populations in semi-rural settings in Thailand. PLoS Negl Trop Dis. 2019;13:e0007771.
van den Hurk AF, Hall-Mendelin S, Pyke AT, Frentiu FD, McElroy K, Day A, et al. Impact of Wolbachia on Infection with Chikungunya and Yellow Fever Viruses in the Mosquito Vector Aedes aegypti. PLoS Negl Trop Dis. 2012;6:e1892.
Hughes GL, Koga R, Xue P, Fukatsu T, Rasgon JL. Wolbachia infections are virulent and inhibit the human malaria parasite Plasmodium falciparum in Anopheles gambiae. PLoS Pathog. 2011;7:e1002043.
Bian G, Joshi D, Dong Y, Lu P, Zhou G, Pan X, et al. Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science. 2013;340:748–51.
Atyame CM, Pasteur N, Dumas E, Tortosa P, Tantely ML, Pocquet N, et al. Cytoplasmic incompatibility as a means of controlling Culex pipiens quinquefasciatus mosquito in the islands of the south-western indian ocean. PLoS Negl Trop Dis. 2011;5:e1440.
Atyame CM, Cattel J, Lebon C, Flores O, Dehecq JS, Weill M, et al. Wolbachia-based population control strategy targeting Culex quinquefasciatus mosquitoes proves efficient under semi-field conditions. PLoS ONE. 2015;10:e0119288.
Ryan PA, Turley AP, Wilson G, Hurst TP, Retzki K, Brown-Kenyon J, et al. Establishment of wMel Wolbachia in Aedes aegypti mosquitoes and reduction of local dengue transmission in Cairns and surrounding locations in northern Queensland. Australia Gates Open Res. 2020;3:1547.
Nazni WA, Hoffmann AA, NoorAfizah A, Cheong YL, Mancini MV, Golding N, et al. Establishment of Wolbachia Strain wAlbB in Malaysian Populations of Aedes aegypti for Dengue Control. Curr Biol. 2019;29:4241-4248.e5.
Alves J, Gomes B, Rodrigues R, Silva J, Arez AP, Pinto J, et al. Mosquito fauna on the Cape Verde islands (West Africa): an update on species distribution and a new finding. J Vector Ecol. 2010;35:307–12.
Salgueiro P, Serrano C, Gomes B, Alves J, Sousa CA, Abecasis A, et al. Phylogeography and invasion history of Aedes aegypti, the Dengue and Zika mosquito vector in Cape Verde islands (West Africa). Evol Appl. 2019;12:1797–811.
Faye O, de Lourdes MM, Vrancken B, Prot M, Lequime S, Diarra M, et al. Genomic epidemiology of 2015–2016 Zika virus outbreak in Cape Verde. Emerg Infect Dis. 2020;26:1084–90.
DePina AdJ, Dia AK, de Ascenção A, Martins S, Ferreira MC, Moreira AL, et al. Knowledge, attitudes and practices about malaria in Cabo Verde: a country in the pre-elimination context. BMC Public Health. 2019;19:850.
Ribeiro H, Ramos HC. Guia ilustrado para a identificação dos mosquitos de Angola. 4th ed. Lisboa: Soc. Portuguesa Entomologia; 1995.
Weeks AR, van Opijnen T, Breeuwer JAJ. AFLP fingerprinting for assessing intraspecific variation and genome mapping in mites. Exp Appl Acarol. 2000;24:775–93.
Scott JA, Brogdon WG, Collins FH. Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am J Trop Med Hyg. 1993;49:520–9.
Smith JL, Fonseca DM. Rapid assays for identification of members of the Culex (Culex) pipiens complex, their hybrids, and other sibling species (Diptera: Culicidae). Am J Trop Med Hyg. 2004;70:339–45.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994;3:294–9.
Zhou W, Rousset F, O’Neill S. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc R Soc B Biol Sci. 1998;265:509–15.
Baldo L, Hotopp JCD, Jolley KA, Bordenstein SR, Biber SA, Choudhury RR, et al. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl Environ Microbiol. 2006;72:7098–110.
Jolley KA, Bray JE, Maiden MCJ. Open-access bacterial population genomics: BIGSdb software, the PubMLST. org website and their applications. Wellcome Open Res. 2018;3:1–20.
Tamura K, Stecher G, Kumar S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38:3022–7.
Altinli M, Gunay F, Alten B, Weill M, Sicard M. Wolbachia diversity and cytoplasmic incompatibility patterns in Culex pipiens populations in Turkey. Parasit Vectors. 2018;11:1–9.
Dumas E, Atyame CM, Milesi P, Fonseca DM, Shaikevich EV, Unal S, et al. Population structure of Wolbachia and cytoplasmic introgression in a complex of mosquito species. BMC Evol Biol. 2013;13:181.
Sicard M, Namias A, Perriat-Sanguinet M, Carron E, Unal S, Altinli M, et al. Cytoplasmic incompatibility variations in relation with Wolbachia cid genes divergence in Culex pipiens. mBio. 2021;12:e02797-20.
Duron O, Boureux A, Echaubard P, Berthomieu A, Berticat C, Fort P, et al. Variability and expression of ankyrin domain genes in Wolbachia variants infecting the mosquito Culex pipiens. J Bacteriol. 2007;189:4442–8.
Atyame C, Delsuc F, Pasteur N, Weill M, Duron O. Diversification of Wolbachia endosymbiont in the Culex pipiens mosquito. Mol Biol Evol. 2011;28:2761–72.
Gomes B, Alves J, Sousa CA, Santa-Ana M, Vieira I, Silva TL, et al. Hybridization and population structure of the Culex pipiens complex in the islands of Macaronesia. Ecol Evol. 2012;2:1889–902.
Goindin D, Cannet A, Delannay C, Ramdini C, Gustave J, Atyame C, et al. Screening of natural Wolbachia infection in Aedes aegypti, Aedes taeniorhynchus and Culex quinquefasciatus from Guadeloupe (French West Indies). Acta Trop. 2018;185:314–7.
Ross PA, Callahan AG, Yang Q, Jasper M, Arif MAK, Afizah AN, et al. An elusive endosymbiont: does Wolbachia occur naturally in Aedes aegypti? Ecol Evol. 2020;10:1581–91.
Gloria-Soria A, Chiodo TG, Powell JR. Lack of evidence for natural Wolbachia infections in Aedes aegypti (Diptera: Culicidae). J Med Entomol. 2018;55:1354–6.
Torres R, Hernandez E, Flores V, Ramirez JL, Joyce AL. Wolbachia in mosquitoes from the Central Valley of California, USA. Parasit Vectors. 2020;13:558.
Kulkarni A, Yu W, Jiang J, Sanchez C, Karna AK, Martinez KJL, et al. Wolbachia pipientis occurs in Aedes aegypti populations in New Mexico and Florida, USA. Ecol Evol. 2019;9:6148–56.
Teo CHJ, Lim PKC, Voon K, Mak JW. Detection of dengue viruses and Wolbachia in Aedes aegypti and Aedes albopictus larvae from four urban localities in Kuala Lumpur. Malaysia Trop Biomed. 2017;34:583–97.
Chrostek E, Gerth M. Is Anopheles gambiae a natural host of wolbachia? MBio. 2019;10:1–10.
Sawadogo SP, Kabore DA, Tibiri EB, Hughes A, Gnankine O, Quek S, et al. Lack of robust evidence for a Wolbachia infection in Anopheles gambiae from Burkina Faso. Med Vet Entomol. 2022;36:301–8.
Gomes FM, Hixson BL, Tyner MDW, Ramirez JL, Canepa GE, Alves Silva TL, et al. Effect of naturally occurring Wolbachia in Anopheles gambiae s. l mosquitoes from Mali on Plasmodium falciparum malaria transmission. Proc Natl Acad Sci USA. 2017;114:12566–71.
Jeffries CL, Lawrence GG, Golovko G, Kristan M, Orsborne J, Spence K, et al. Novel Wolbachia strains in Anopheles malaria vectors from Sub-Saharan Africa. Wellcome Open Res. 2018;3:113.
Shaw WR, Marcenac P, Childs LM, Buckee CO, Baldini F, Sawadogo SP, et al. Wolbachia infections in natural Anopheles populations affect egg laying and negatively correlate with Plasmodium development. Nat Commun. 2016;7:11772.
Duarte EH. The haematophagous arthropods (Animalia: Arthropoda) of the Cape Verde Islands: a review. Zool Caboverdiana. 2014;4:31–42.
Correia W, Varela I, Spencer H, Alves J, Heleno DE. Characterization of mosquito breeding sites in the Cape Verde islands with emphasis on major vectors. Int J Mosq Res. 2015;2:192–9.
Bandi C, Anderson TJC, Genchi C, Blaxter ML. Phylogeny of Wolbachia in filarial nematodes. Proc R Soc B Biol Sci. 1998;265:2407–13.
Acknowledgements
We are grateful to the National Institute of Public Health for the laboratory support in Cape Verde, and to technicians from the Ministry of Health for their assistance in field work. We would like to thank Prof. Paulo Almeida for providing DNA controls of Cx. pipiens and Cx. quinquefasciatus used in the species identification PCR assay.
Funding
This work was funded by national funds through FCT—Fundação para a Ciência e Tecnologia, I.P., within the framework of the project ARBOMONITOR (PTDC/BIA-OUT/29477/2017. Aires da Moura was funded by the Ph.D. fellowship program of Camões I.P.
Author information
Authors and Affiliations
Contributions
AJFM, CAS, and JP designed the study. AJFM, EMS, and SLV performed field work and specimen identification. AJFM and VV performed the molecular laboratory work. AJFM, SLV, CAS, and JP drafted the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Primer sequences used for molecular identification of mosquito species collected in Cape Verde islands. Table S2. Primers used for PCR detection of Wolbachia and genotyping of wPip I–V groups by PCR-RFLP. Table S3. Primers used for Wolbachia MLST loci and wsp hypervariable region amplification and sequence analysis.
Additional file 2: Table S4
. Mosquito species collected on each island and tested for Wolbachia using wsp.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
da Moura, A.J.F., Valadas, V., Da Veiga Leal, S. et al. Screening of natural Wolbachia infection in mosquitoes (Diptera: Culicidae) from the Cape Verde islands. Parasites Vectors 16, 142 (2023). https://doi.org/10.1186/s13071-023-05745-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-05745-w