
Abstract
Background
Myanmar is one of the six countries in the Greater Mekong Subregion (GMS) of Southeast Asia. Malaria vectors comprise many Anopheles species, which vary in abundance and importance in malaria transmission among different geographical locations in the GMS. Information about the species composition, abundance, and insecticide resistance status of vectorial systems in Myanmar is scarce, hindering our efforts to effectively control malaria vectors in this region.
Methods
During October and November 2019, larvae and adult females of Anopheles mosquitoes were collected in three sentinel villages of Banmauk township in northern Myanmar. Adult female mosquitoes collected by cow-baited tent collection (CBTC) and adults reared from field-collected larvae (RFCL) were used to determine mortality rates and knockdown resistance (kdr) against deltamethrin using the standard WHO susceptibility test. Molecular species identification was performed by multiplex PCR and ITS2 PCR, followed by DNA sequencing. The kdr mutation at position 1014 of the voltage-gated sodium channel gene was genotyped by DNA sequencing for all Anopheles species tested.
Results
A total of 1596 Anopheles mosquitoes from seven morphologically identified species groups were bioassayed. Confirmed resistance to deltamethrin was detected in the populations of An. barbirostris (s.l.), An. hyrcanus (s.l.), and An. vagus, while possible resistance was detected in An. annularis (s.l.), An. minimus, and An. tessellatus. Anopheles kochi was found susceptible to deltamethrin. Compared to adults collected by CBTC, female adults from RFCL had significantly lower mortality rates in the four species complexes. A total of 1638 individuals from 22 Anopheles species were molecularly identified, with the four most common species being An. dissidens (20.5%) of the Barbirostris group, An. peditaeniatus (19.4%) of the Hyrcanus group, An. aconitus (13.4%) of the Funestus group, and An. nivipes (11.5%) of the Annularis group. The kdr mutation L1014F was only detected in the homozygous state in two An. subpictus (s.l.) specimens and in a heterozygous state in one An. culicifacies (s.l.) specimen.
Conclusions
This study provides updated information about malaria vector species composition and insecticide resistance status in northern Myanmar. The confirmed deltamethrin resistance in multiple species groups constitutes a significant threat to malaria vector control. The lack or low frequency of target-site resistance mutations suggests that other mechanisms are involved in resistance. Continual monitoring of the insecticide resistance of malaria vectors is required for effective vector control and insecticide resistance management.
Graphical Abstract

Similar content being viewed by others

Background
Despite remarkable achievements in the battle against vector-borne diseases over the last decade, malaria remains a major public health problem in many developing countries, especially as progress has been stalled in recent years because of the COVID-19 pandemic. In 2020, there were an estimated 241 million malaria cases and 627,000 deaths from malaria worldwide [1]. Among the six Greater Mekong Subregion (GMS) countries of the WHO Southeast Asia Region, Myanmar had the highest malaria incidence, accounting for 71% of the indigenous cases of malaria in the GMS in 2020 [1].
Malaria control relies heavily on the use of indoor residual spraying (IRS) and long-lasting insecticide-treated nets (LLINs) to prevent mosquitoes from transmitting the malaria parasite to humans [2]. However, the extensive use of chemical insecticides to control disease vectors and other insect pests has resulted in insecticide resistance in many malaria vectors worldwide [3]. The widespread resistance to insecticides has become a significant threat to malaria vector control [4]. In Africa, insecticide resistance and its underlying mechanisms have been well documented in the primary malaria vector Anopheles gambiae complex. Resistance to pyrethroids was the most commonly reported, with two main types of known resistance mechanisms: changes at the target site of the insecticide to reduce sensitivity (e.g. knockdown resistance, kdr) and increases in the rate of insecticide metabolism due to increased activity or level of detoxification enzyme(s) [5]. In Southeast Asia, however, much less information is available about the status and mechanisms of insecticide resistance in the complex vector systems.
Before the 1980s, regular resistance monitoring of malaria vectors was rarely conducted in the GMS countries. In Vietnam, resistance monitoring indicated that DDT resistance had already existed before 1989 in the malaria vector An. epiroticus of the Sundaicus Complex. In the 1990s, resistance to pyrethroid insecticides was almost absent in all tested species except in some populations of An. vagus and the Minimus Complex [6]. There was no evidence of insecticide resistance in malaria vectors in Thailand before 1985. However, from 1986 to 2010, DDT resistance was found in the three primary malaria vectors, An. dirus (s.l.), An. minimus (s.l.), and An. maculatus (s.l.), as well as the three secondary vectors, An. aconitus, An. philippinensis, and An. nivipes. Permethrin resistance was also detected in a population of An. minimus (s.l.) from northern Thailand [6,7,8]. During 2003 and 2005, in the Mekong Delta region of southern Vietnam, An. epiroticus was found to be highly resistant to all pyrethroid insecticides tested. Anopheles vagus was found resistant to DDT and several pyrethroids in Vietnam and Cambodia. Anopheles minimus (s.l.) showed susceptibility to pyrethroid insecticides in Cambodia, Laos, and Thailand [6].
Recently, An. hyrcanus (s.l.) was found highly resistant to multiple insecticides tested (deltamethrin 0.05%, permethrin 0.75% and DDT 4%) in northeastern Thailand, with mosquito mortality ranging from 45 to 87% [9, 10], and at the Myanmar-Thailand border, with mortality ranging from 33 to 57% [11]. Resistance or possible resistance to multiple insecticides was also detected in An. barbirostris (s.l.), An. maculatus, and An. vagus in the Myanmar-Thailand border region, whereas the primary vectors An. dirus (s.l.) and An. minimus (s.l.) remained susceptible or had suspected resistance to pyrethroids and DDT [9, 11]. In China, high-level resistance to multiple insecticides has been found in An. sinensis [12,13,14,15].
The mechanisms of resistance to pyrethroid insecticides have been well documented in An. sinensis populations from China. Three non-synonymous kdr mutations (L1014F, L1014C, and L1014S) have been detected at codon L1014 of the para-type sodium channel gene in An. sinensis, and these kdr mutant alleles exhibited a patchy distribution in frequency from southern to central China. Near fixation of the kdr mutation was detected in populations from central China, but no kdr mutation was found in Yunnan Province of southwestern China, suggesting that kdr alone is insufficient to predict pyrethroid resistance [16]. Classification and regression tree statistical analysis found that metabolic detoxification was the most important resistance mechanism, whereas target site insensitivity of L1014 kdr mutation played a minor role [12]. Whole-genome sequencing (WGS) and RNA-seq analysis identified some transcripts and SNPs associated with phenotypic resistance to deltamethrin [17, 18]. In Thailand and Myanmar, no kdr mutation was detected in any Anopheles species tested except An. peditaeniatus of the Hyrcanus group, where the kdr mutation L1014S was detected at a low frequency [9, 11]. In Thailand, pre-exposure of An. hyrcanus (s.l.) to 4% piperonyl butoxide caused a significant increase in mortality or restored susceptibility, indicating a potential role of oxidases as a detoxifying enzyme resistance mechanism [9, 10].
Malaria vectors are complex in the GMS countries; many Anopheles species exist with varying abundance and importance in malaria transmission among different geographical regions. In south-central Vietnam, 24 Anopheles species were identified, with the predominant species being An. dirus in Binh Phuoc Province and An. maculatus in Dak Nong Province [19]. In Ubon Ratchathani Province of northeastern Thailand, 18 Anopheles species were identified, with An. peditaeniatus as the most abundant species (> 90%) [10]. Rattanarithikul et al. (2006) listed 73 species of Anopheles across Thailand [20]. In Myanmar, vector surveillance conducted during the 1970s identified 36 Anopheles species, and 10 species were infected with the malaria parasites [21]. However, little information is available about the current community structure and insecticide resistance of the vectorial systems in Myanmar. Such information is needed to guide the vector control practice in this region.
The objectives of this study are to determine Anopheles species composition and insecticide resistance status as well as the underlying resistance mechanisms in northern-central Myanmar, to determine the differences between sampling methods (field adult and larval collections) for resistance surveillance, to test the universal kdr primer set in different Anopheles species collected in Myanmar, and finally to determine the evolutionary relationships among species and species complexes.
Methods
Study sites and mosquito collection
During October–November 2019, entomological surveillance was carried out in three villages located in Banmauk Township, Katha District, Sagaing Region, northern-central Myanmar, namely Pin Hin Khar (24°25′33″N, 95°45′27″ E), Mankat (24°20′44″N, 9549′45″E), and Lay Thi (24°25′35″N, 95°51′58″E) (Fig. 1). In Myanmar, there are three weather seasons: cool dry season (November- February), hot dry season (March to May), and wet (monsoon) season (June to October) [22]. The late rainy season was selected for sampling because of higher mosquito density [23, 24], greater species richness, and relatively little variation among species in their abundance [25]. Anopheles adult female mosquitoes were collected every day for 10 days from each village using the cow-baited tent collection (CBTC) method [21, 26]. The collected female mosquitoes belonging to different species were separated by morphology [27] into different cages and reared for 2–3 days to conduct the insecticide susceptibility bioassays. Anopheles larvae were collected from different habitat types, including rice fields, rivers, streams, rainwater pools, and other water habitats. To minimize the sampling of siblings, no more than ten specimens (larvae and pupae) were collected from each habitat using a standard 350-ml capacity mosquito dipper. All the randomly selected larval habitats were at least 50 m away from each other. All the collected larvae and pupae were reared to adults (referred to as RFCL) under standard insectary conditions at 27 ± 1 °C, approximately 80% relative humidity, and a photoperiod of 12:12 (L:D) h. The emerged adult females were provided with cotton balls soaked in an 8% glucose solution. Non-blood-fed females 3–5 days post-emergence were separated by morphology into different cages and used for the bioassay as described below. Adult female Anopheles were individually transferred to a clean and transparent plastic tube using a HEPA Filter Mouth Aspirator (John W. Hock Company, Gainesville, FL, USA). The morphological identification of Anopheles individuals was performed while the mosquito was alive (a brief chill on ice if needed) following the taxonomic keys [27].

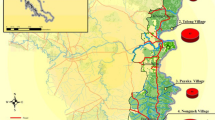
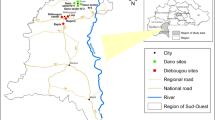
source at: www.esri.com
Map of study sites in Banmauk Township, Katha District, Sagaing Region of northern-central Myanmar. Red triangles indicate the locations of the three villages. The map was generated using ArcGIS Pro software based on map
Insecticide susceptibility bioassay
The adult female Anopheles mosquitoes obtained from both the CBTC and RFCL collections were used to conduct bioassays following the standard WHO insecticide susceptibility tube-test procedures for malaria vectors [5] to determine their susceptibility to the pyrethroid insecticide deltamethrin (0.05%). Test paper treated with silicone oil was used as the control. For each morphologically identified Anopheles species group, female mosquitoes with sample size > 50 were tested with 20–25 mosquitoes per tube and two control tubes without insecticide. The number of knocked-down mosquitoes was recorded at 0, 10, 15, 20, 30, 40, 50, and 60 min after mosquitoes were transferred into test tubes. After 1 h of exposure to the insecticide, the females were transferred into holding tubes, provided with cotton balls soaked in 8% glucose, and kept at 28 °C with a relative humidity of 80%. Mortality was recorded after a 24-h recovery period. Tests were replicated at least twice if enough specimens were collected. The bioassay results were interpreted per WHO guidelines updated in June 2018: confirmed resistance (mortality < 90%), possible resistance (mortality between 90 and 98%), and susceptible (mortality ≥ 98%) [5]. After bioassays, all specimens were individually transferred into a 1.5-ml Eppendorf tube containing silica gel and cotton wool and preserved at − 20 °C for further molecular species identification.
DNA extraction
All the bioassayed specimens (n = 1596) and untested samples (n = 42; all species with samples insufficient for the replicate test) were used for DNA extraction and molecular identification of Anopheles species. DNA extraction was performed by a rapid and simple method following the published protocol with modifications [28]. Briefly, a single leg of an ethanol-washed mosquito was transferred into a 1.5-ml Eppendorf tube containing 475 µl l × PBS solution and 25 µl 10% saponin solution. The sample was mixed well by gentle vortexing and centrifuged at 18,000 g for 10 min. After discarding the liquid with a 1-ml pipette, the sample was washed by adding 1 ml 1 × PBS solution and mixing well by vortexing. The sample was centrifuged at 18,000 g for 10 min, and then the supernatant was removed. After centrifuging at 18,000 g for 1 min, the additional liquid was removed using a 200-µl pipette, and the sample was dried at room temperature for 15 min. DNA from the mosquito leg was eluted by adding 80 µl of 20% Chelex solution and incubated for 10 min on a heating block shaker (Thomas Scientific, Swedesboro, NJ). After centrifuging at 18,000 g for 1 min, the supernatant (DNA) was transferred into a fresh collection tube and stored at − 20 °C until use.
Molecular identification of Anopheles species
Multiplex PCR was used for the accurate identification of Anopheles species in the Funestus group (An. minimus subgroup and An. funestus subgroup) [29], the Hyrcanus group [30], the Annularis group [31], and the Barbirostris group [32]. PCR and DNA sequencings of the internal transcribed spacer region (ITS2) of the nuclear ribosomal DNA were also carried out for a subset of samples to confirm the species identified by multiplex PCR and for all samples not determined by the multiplex PCR method. Amplification of the ITS2 region was performed using the primer pair ITS2A (5′-TGT GAA CTG CAG GAC ACA T-3′) and ITS2B (5′-TAT GCT TAA ATT CAG GGG GT-3′) [33]. The detailed PCR conditions and DNA sequencing procedures have been described previously [34].
Detection of kdr mutations
The kdr mutation of the voltage-gated sodium channel gene at codon L1014 residue was examined by PCR and DNA sequencing for all Anopheles species (n = 338) using the universal primer pair Ag-F (5'-GAC CAT GAT CTG CCA AGA TGG AAT-3') and An-kdr-R2 (5'-GAG GAT GAA CCG AAA TTG GAC-3') [35]. The PCR was conducted in a total reaction volume of 17 μl, including 2 μl of DNA template, 5 pmol of each primer, and 8.5 μl of DreamTaq Green PCR Master Mix (2X) (Thermo Fisher Scientific, Waltham, MA, USA). The thermocycling protocol consisted of an initial activation step of 3 min at 95 °C, followed by 35 amplification cycles of 30 s at 94 °C, 30 s at 55 °C, 45 s at 72 °C, and a final extension step of 6 min at 72 °C. Purification and DNA sequencing of PCR products were previously described [34].
Data analysis
Mortality rates of adult female mosquitoes were calculated after the bioassays for each species group. Abbott’s formula was used to correct mortality rates if mortality in the control tube was between 5 and 20% [36]. Tests with > 20% mortality in the control were excluded from the analysis. Time to 50% knockdown (KDT50) and its confidence interval for each Anopheles species were determined by the log-probit method described by Finney [37] using the Excel spreadsheet calculator [38]. Analysis of variance (ANOVA) was performed to determine differences in mortality rates among species and between the two sampling methods.
The CodonCode Aligner 9.0.1 (CodonCode Corp., Centerville, MA) was used to check the sequence quality and trim low-quality bases. Bio-Edit software was used to align the sequences and to calculate pairwise sequence identity and similarity from multiple sequence alignments. A threshold limit of 98% sequence similarity for ITS2 was used to classify sequences into species [34]. The consensus sequences were compared to the NCBI nr/nt database (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Phylogenetic analyses were performed using UPGMA with the Kimura 2-parameter model for ITS2 and kdr sequences. The tree nodes were evaluated by bootstrap analysis for 1000 replicates.
Results
Insecticide susceptibility bioassays
A total of 868 adult females were collected from cow-baited tents and separated based on morphology and abundance (minimum sample size > 50) into six main species groups (Barbirostris group, Hyrcanus group, Annularis group, Funestus group, An. tessellatus, and An. kochi) and others. Over 5000 Anopheles mosquito larvae and pupae were collected from the three study villages. A total of 770 adult females were reared from field-collected larvae (RFCL) and classified into six different species groups (Barbirostris group, Hyrcanus group, Annularis group, Funestus group, An. vagus, and An. kochi) and others. Deltamethrin susceptibility bioassays were performed on adult females from both CBTC and RFCL based on their species group. The mortality of mosquitoes without exposure to insecticide was < 5% in all control tubes. The laboratory strain of An. sinensis, when exposed to the insecticide, showed average mortality of > 99%. Deltamethrin resistance was found in mosquitoes of the Barbirostris group and the Hyrcanus group from both CBTC (mortality rate 77.9–81.1%) and RFCL (mortality rate 67.9–71.5%) as well as in An. vagus from RFCL (mortality rate 72.5 ± 2.9%) (Table 1, Fig. 2). Possible resistance to deltamethrin was found in mosquitoes of the Minimus group and the Annularis group from RFCL and in An. tessellatus from CBTC, while An. kochi mosquitoes from both CBTC and RFCL were found susceptible to deltamethrin. A significant difference in mortality rate was found among species (ANOVA, F(6,85) = 12.7, P < 0.001). Significantly lower mortality rates were found in adult females reared from field-collected larvae compared to those from CBTC in all species combined (ANOVA, F(1,90) = 6.7, P < 0.01) and in four out of the seven species groups (ANOVA, F(1,56) = 3.8, P < 0.05) (Table 1). The knockdown time ranged from 11.6 to 24.9 min in mosquitoes from CBTC and from 15.8 to 26.6 min in mosquitoes from RFCL (Table 1).

Mortality rate of Anopheles mosquitoes exposed to deltamethrin following the WHO susceptibility test procedure for insecticide monitoring in malaria vectors. The mortality rate was recorded after a 24-h recovery period. NA, not available. Red dashed line indicates cutoff for confirmed resistance (< 90%) (WHO 2018 update). Anopheles sinensis susceptible laboratory strain showed 100% mortality
Molecular identification of Anopheles species
The bioassayed mosquito samples from four species groups were genotyped by multiplex PCR and classified into seven species: An. dissidens (formerly Anopheles barbirostris species A1) (n = 336), An. peditaeniatus (n = 318), An. aconitus (n = 220), An. nivipes (n = 189), An. sinensis (n = 108), An. annularis (n = 70), and An. minimus (n = 28) (Table 2). PCR and DNA sequencing of the ITS2 region confirmed the identities of these species (n > 20 per species) and identified four additional species: An. nigerrimus (n = 2) from the Hyrcanus group and An. culicifacies (s.l.) (n = 10), An. varuna (n = 3), and An. jeyporiensis (n = 1) from the Funestus group. The ITS2 sequences of An. culicifacies (s.l.) showed > 99% similarity to multiple subspecies: An. culicifacies B, C, and E (GenBank AJ534247, AJ534643, and AJ534645). Thus, the accurate subspecies of this complex could not be determined by the ITS2 sequences. All specimens from each of the three species groups An. kochi, An. tessellatus, and An. vagus were confirmed by ITS2 sequencing. In addition, 42 specimens of the minor species not used for bioassays were also sequenced and classified into eight Anopheles species from five groups (Jamesii, Maculatus, Dirus, Subpictus, and Stephensi) compared to the ITS2 sequences retrieved from GenBank (Table 2). The ITS2 sequences of An. subpictus (s.l.) showed > 99% similarity to An. subpictus A cluster (GenBank KC191825), which includes multiple biological forms: An. subpictus A, C, and D. Thus, the accurate biological forms of this complex could not be determined by the ITS2 sequences. Phylogenetic analysis indicated that species from the same species group were generally clustered together with large bootstrap values, such as for the Hyrcanus, Maculatus, and Funestus groups (Fig. 3a). The Barbirostris group included only one species, An. dissidens, with the longest ITS2 sequence length (1.8 k), which fell into a different cluster in the phylogenetic tree.

Phylogenetic tree inferred from UPGMA method based on ITS2 sequences (a) and kdr sequences (b). Bootstrap support values ≥ 50% are displayed by colored dots next to the branches. Tips of the tree are labeled as GenBank accession followed by Anopheles species identified in this study
Kdr mutations in the voltage-gated sodium channel gene
In total, 338 specimens from 22 Anopheles species were successfully sequenced for the kdr genes covering codon L1014. The L1014F kdr mutation was detected in the homozygous state in two An. subpictus (s.l.) specimens and in the heterozygous state in one An. culicifacies (s.l.) specimen. All other species were wild type at the kdr L1014 locus (Additional file 1: Fig. S1). Phylogenetic analysis of the 22 Anopheles species indicated that the kdr sequences generally clustered together within the same species group with large bootstrap values and showed a similar pattern with that inferred from the UPGMA method based on the ITS2 sequences (Fig. 3b).
Discussion
Understanding vector community structure, species distribution, vector abundance, and insecticide resistance is essential for developing cost-effective strategies to control malaria vectors. This study provides updated information about Anopheles species composition and abundance surveyed from adult and larva collections of field mosquito populations and, for the first time to our knowledge, reports the insecticide resistance of seven species or species complexes in northern-central Myanmar. The kdr primer set was confirmed to work for all the Anopheles species identified in this study.
Out of 22 Anopheles species identified molecularly, four (An. dissidens, An. peditaeniatus, An. aconitus, and An. nivipes) were the most common (accounting for ~ 65%), while the rest were below 10% each. Previous surveillance of malaria vectors conducted in the nearby township (Indaw, Saging Division) identified 19 Anopheles species, with An. philippinensis, An. fluviatilis (s.l.), An. jamesii, and An. vagus as the four most common species based on morphological identification [21]. However, this study did not find four previously reported species (An. philippinensis, An. fluviatilis, An. karwari, and An. majidi), possibly because of the short duration for sample collection or different sampling methods used. In contrast, this study identified seven previously unreported species (An. dirus, An. peditaeniatus, An. nigerrimus, An. nivipes, An. sawadwongporni, An. dravidicus, and An. jeyporiensis), which were further confirmed by molecular identification. The changes in species composition and abundance may reflect local environmental changes causing species to shift their habitat range, biting and resting behaviors, and host choice as they track their ecological niches [39].
The widespread use of insecticide-treated nets (ITNs) and IRS has been shown to alter the species composition, abundance, and behavior of vector populations [40, 41]. In Thailand, IRS resulted in a higher proportional decrease of An. dirus (s.l.) compared to An. minimus (s.l.) [42], whereas ITN use in China caused a more substantial decrease of the endophilic and anthropophilic An. lesteri (synonym: An. anthropophagus) [43] and An. minimus (s.l.) [44] than of the exophagic and zoophilic An. sinensis. In Nepal, residual spraying has proved to be effective against indoor resting species, e.g. An. annularis, An. culicifacies (s.l.), An. splendidus and An. vagus, and some outdoor resting species, e.g. An. fluviatilis (s.l.) and An. maculatus (s.l.) [45]. Shifts of malaria vectors in biting behavior such as shifts to outdoor or early biting and shifts to zoophily or exophily have been observed in An. dirus (s.l.) and An. minimus (s.l.) of Thailand [42, 46, 47]. The level of behavioral heterogeneity of Anopheles species and populations is much higher in Southeast Asia than on the other continents according to ecological situations [48, 49]. These heterogeneities of species and behavioral shifting may result in differences in epidemiological importance and will have a great potential to increase malaria transmission in regions with a high coverage of ITNs and IRS [40, 41, 50]. Thus, there is an urgent need for additional control measures tackling malaria transmission by these vector populations.
Previous parasitological studies identified by morphology only one primary vector (An. minimus) and six secondary vectors (An. aconitus, An. annularis, An. culicifacies (s.l.), An. sinensis, An. maculatus, and An. philippinensis) in Indaw township [21]. In this study, An. dirus (a primary vector in Southeast Asia) was also found at low density. Whereas the secondary vector An. philippinensis was not detected, the closely related, morphologically similar species An. nivipes of the Annularis group was collected in Banmauk township. These results may reflect the effects of seasonality, sampling methods, and sampling duration in species composition and richness. Among the four common species identified in this study, An. dissidens (formerly An. barbirostris species A1), currently known from Thailand, China, and Myanmar [51], has been reported as a potential vector of P. vivax in Thailand [52, 53]. Anopheles peditaeniatus of the Hyrcanus group has been reported as a secondary vector of Japanese encephalitis virus in China and India [54,55,56] and a potential vector of Brugia malayi filarial worms [57]. Anopheles peditaeniatus has also been reported to be positive for P. falciparum infection by ELISA in Sri Lanka [58], Thailand [57, 59], and Indonesia [60]. The common species An. nivipes in Banmauk is a recognized malaria vector in Bangladesh, Thailand, and India [61]. The rare species An. nigerrimus of the Hyrcanus group in Banmauk was also recognized as a suspected vector of malaria in Bangladesh [62].
Out of the seven Anopheles groups tested in this study, three (Barbirostris, Hyrcanus, and Vagus) showed resistance to the pyrethroid insecticide deltamethrin, suggesting the presence of insecticide resistance in malaria vectors from northern Myanmar. Anopheles dissidens is the only common species detected in the Barbirostris group from Banmauk, whereas three species in the Hyrcanus group were found, with An. peditaeniatus being the predominant (~ 3/4), followed by An. sinensis (~ 1/4) and An. nigerrimus (< 1%). Resistance to deltamethrin has been previously reported in the Barbirostris group, the Hyrcanus group, and An. vagus from the Thai-Myanmar border on the Myanmar side [11] and in the Hyrcanus group from China [12,13,14,15, 50] and northeast Thailand [9] as well as in An. vagus from Bangladesh [63]. In the Hyrcanus group, all the resistant specimens were An. peditaeniatus, consistent with those findings from northeast Thailand [10]. However, the Annularis group, the Funestus group, An. tessellatus, and An. kochi were found to be susceptible (in the adult collection) or possibly resistant (in the larvae collection), similar to those detected in the Thai-Myanmar border area [11]. In the Sagaing Division of Myanmar, deltamethrin resistance in the Hyrcanus group has also been confirmed by the WHO, while other Anopheles species are susceptible or remain to be tested [64].
Previous findings showed that mosquito sampling and preparation methods significantly affected the mortality rates in the standard WHO tube resistance bioassay [65,66,67]. In Africa, Oliver, et al. (2014) reported that pyrethroid resistance levels in laboratory-reared non-blood-fed An. arabiensis females steadily decreased with age, whereas blood-fed females could significantly increase or maintain resistance compared to non-blood-fed females at the same age. Similar results were also found in the adult females reared from field-collected larvae of An. gambiae in western Kenya [67]. This study found that field-collected mixed-age blood-fed adult females of all four Anopheles species groups had significantly increased mortality rates against deltamethrin compared to non-blood-fed adult females reared from the larvae collection, consistent with a previous report of An. sinensis populations from central China [65]. Therefore, to minimize the effect of confounding factors such as mosquito age and blood-feeding status, the use of non-blood-fed adult 3–5 day-old female Anopheles is recommended to assess the susceptibility of field malaria vector populations to insecticides [5].
The insecticide resistance mechanism is still largely unknown in most Anopheles vector species from the GMS. Previous findings showed that both target site mutations (kdr) and metabolic detoxification play important roles in the deltamethrin resistance of An. sinensis field populations from China [12,13,14,15]. In An. peditaeniatus, a high allele frequency of L1014F kdr mutation (97.6%) was reported in Vietnam [68], whereas the L1014S kdr mutation was found in Vietnam, Thailand, and the Thai-Myanmar border at low frequencies [10, 11, 68]. The L1014S kdr mutation was also found in An. vagus, An. sinensis, and An. paraliae at low frequencies in Vietnam, but not in Thailand or Myanmar [10, 11, 68]. Among the 22 Anopheles species tested in this study, the kdr mutation (L1014F) was only detected in An. subpictus (s.l.) and at a low allele frequency in An. culicifacies (s.l.). The lack or scarcity of target-site resistance mutations suggested the involvement of other resistance mechanisms. The other resistane mechanisms include metabolic resistance by increased activities of detoxification enzymes (e.g. cytochrome P450 monooxygenases, glutathione S-transferase, and non-specific esterases), penetration resistance through thickening or remodeling of the cuticle to prevent the absorption or penetration of insecticide, and behavioral resistance by a change from their normal activity to avoid contact with or ingestion of a lethal dose of an insecticide [69]. Of the six families of P450s (CYP4, 6, 9, 12, 18, and 28), the CYP6 family has been well documented as being involved in insecticide resistance in the malaria vector An. gambiae [70,71,72]. Overexpression of cytochrome P450 isoforms (CYP6AA2, CYP6AA3, and CYP6P7) was also observed in laboratory-selected pyrethroid-resistant An. minimus mosquitoes in Thailand [73, 74].
This study has some limitations. First, we only conducted Anopheles resistance monitoring in a short period of time in three villages, considering the difficulty of accessing the study sites in northern central Myanmar. Second, identifying malaria vectors at the species level based on morphological traits remains a challenge due to the labor-intensiveness and difficulty separating species from groups or species complexes that are morphologically indistinguishable. Third, we could not collect enough mosquitoes of less abundant species (or species complex) of Anopheles for bioassay. Thus, only seven predominant species were assessed for susceptibility to insecticide in the study. Future studies should consider including testing different insecticides, collecting mosquitoes using multiple methods across different seasons, and covering a larger region to obtain a clear picture of the insecticide resistance situation in Myanmar. Molecular identification of Anopheles species should also be strengthened so that more species can be identified by the regular PCR method.
Bioassay and molecular monitoring of insecticide resistance status in local malaria vectors are crucial for resistance management and vector control. This study reported the presence of deltamethrin resistance in three Anopheles species complexes [An. barbirostris (s.l.), An. hyrcanus (s.l.), and An. vagus] tested in northern Myanmar, suggesting that these species complexes are probably subjected to the high selective pressure of insecticides used in the local ecological environment [75]. In Myanmar, pyrethroid insecticides have been intensively used for malaria control in the interior and along the periphery of human habitation areas since 1992 [76]. The use of pyrethroid insecticides in vector control and agricultural practices has potentially exerted selection pressure for resistance in mosquito populations, which may explain the deltamethrin resistance detected in the Anopheles complexes in this study [75]. Several strategies have been recommended to manage the resistance evolution in vector populations against chemical insecticides, such as minimizing insecticide use, avoiding persistent chemicals, using insecticide mixture or insecticide mosaic, and rotating insecticides with different chemical classes or different control methods [3].
Conclusions
This study updated Anopheles species composition, vector abundance, and insecticide resistance status in northern Myanmar. The presence of confirmed resistance to pyrethroid insecticide and the absence of knockdown mutation (kdr) in An. dissidens, An. peditaeniatus, and An. vagus have important implications in pyrethroid-based vector control and resistance management. To identify alternatives to pyrethroids for vector control, further studies are needed to document the non-target-site resistance mechanisms (e.g. rapid metabolic detoxification) and the susceptibility status of malaria vectors to other classes of insecticides (e.g. carbamates, organophosphates, and insect growth regulators).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request. The ITS2 sequences obtained in the study are available in GenBank with accession numbers OM060212–OM060233. The kdr sequences obtained in the study are available in GenBank with accession numbers OL979443–OL979465.
Abbreviations
- GMS:
-
Greater Mekong Subregion
- WHO:
-
World Health Organization
- COVID:
-
Coronavirus Disease
- CBTC:
-
Female adults from cow-baited tent collection
- RFCL:
-
Female adults reared from field-collected larvae
- kdr :
-
Knockdown resistance
- KT50:
-
Time to knockdown 50% mosquitoes
- MR:
-
Mortality rate
- CI:
-
Confidence interval
- S:
-
Susceptible
- PR:
-
Probably resistant
- R:
-
Resistant
- ITS2:
-
Ribosomal internal transcribed spacer 2
- PCR:
-
Polymerase chain reaction
- DDT:
-
Dichlorodiphenyltrichloroethane
- ANOVA:
-
Analysis of variance
- UPGMA:
-
Unweighted pair group method with arithmetic mean
References
World Health Organization. World malaria report 2021. Geneva, Switzerland: WHO; 2021.
World Health Organization. World malaria report 2013. Geneva, Switzerland: WHO; 2013.
Corbel V, N’Guessan R. Distribution, mechanisms, impact and management of insecticide resistance in malaria vectors: a pragmatic review. In: Manguin S, editor. Anopheles mosquitoes - New insights into malaria vectors: Rijeka: Intech; 2013.
Hemingway J. The role of vector control in stopping the transmission of malaria: threats and opportunities. Philos Trans R Soc Lond B Biol Sci. 2014;369:20130431.
World Health Organization. Test procedures for insecticide resistance monitoring in malaria vector mosquitoes (updated in June 2018). Geneva, Switzerland: WHO; 2018.
Van Bortel W, Trung HD, Thuan LK, Sochantha T, Socheat D, Sumrandee C, et al. The insecticide resistance status of malaria vectors in the Mekong region. Malaria J. 2008;7:102.
Chareonviriyahpap T, Aum-aung B, Ratanatham S. Current insecticide resistance patterns in mosquito vectors in Thailand. Southeast Asian J Trop Med Public Health. 1999;30:184–94.
Chareonviriyaphap T, Bangs MJ, Suwonkerd W, Kongmee M, Corbel V, Ngoen-Klan R. Review of insecticide resistance and behavioral avoidance of vectors of human diseases in Thailand. Parasit Vectors. 2013;6:280.
Sumarnrote A, Overgaard HJ, Marasri N, Fustec B, Thanispong K, Chareonviriyaphap T, et al. Status of insecticide resistance in Anopheles mosquitoes in Ubon Ratchathani province Northeastern Thailand. Malar J. 2017;16:299.
Sumarnrote A, Overgaard HJ, Corbel V, Thanispong K, Chareonviriyaphap T, Manguin S. Species diversity and insecticide resistance within the Anopheles hyrcanus group in Ubon Ratchathani Province Thailand. Parasit Vectors. 2020;13:525.
Chaumeau V, Cerqueira D, Zadrozny J, Kittiphanakun P, Andolina C, Chareonviriyaphap T, et al. Insecticide resistance in malaria vectors along the Thailand-Myanmar border. Parasit Vectors. 2017;10:165.
Zhong D, Chang X, Zhou G, He Z, Fu F, Yan Z, et al. Relationship between knockdown resistance, metabolic detoxification and organismal resistance to pyrethroids in Anopheles sinensis. PLoS ONE. 2013;8:e55475.
Chang X, Zhong D, Fang Q, Hartsel J, Zhou G, Shi L, et al. Multiple resistances and complex mechanisms of Anopheles sinensis mosquito: a major obstacle to mosquito-borne diseases control and elimination in China. PLoS Negl Trop Dis. 2014;8:e2889.
Qin Q, Li Y, Zhong D, Zhou N, Chang X, Li C, et al. Insecticide resistance of Anopheles sinensis and An. vagus in Hainan Island, a malaria-endemic area of China. Parasit Vectors. 2014;7:92.
Chen S, Qin Q, Zhong D, Fang X, He H, Wang L, et al. Insecticide resistance status and mechanisms of Anopheles sinensis (Diptera: Culicidae) in Wenzhou, an important coastal port city in China. J Med Entomol. 2019;56:803–10.
Chang X, Zhong D, Lo E, Fang Q, Bonizzoni M, Wang X, et al. Landscape genetic structure and evolutionary genetics of insecticide resistance gene mutations in Anopheles sinensis. Parasit Vectors. 2016;9:228.
Chang X, Zhong D, Wang X, Bonizzoni M, Li Y, Zhou G, et al. Genomic variant analyses in pyrethroid resistant and susceptible malaria vector, Anopheles sinensis. G3 (Bethesda). 2020;10:2185–93.
Zhu G, Zhong D, Cao J, Zhou H, Li J, Liu Y, et al. Transcriptome profiling of pyrethroid resistant and susceptible mosquitoes in the malaria vector Anopheles sinensis. BMC Genomics. 2014;15:448.
Ngo C, Dubois G, Sinou V, Parzy D, Le H, Harbach R, et al. Diversity of Anopheles mosquitoes in Binh Phuoc and Dak Nong Provinces of Vietnam and their relation to disease. Parasit Vectors. 2014;7:316.
Rattanarithikul R, Harrison BA, Harbach RE, Panthusiri P, Coleman RE, Panthusiri P. Illustrated keys to the mosquitoes of Thailand. IV. Anopheles. Southeast Asian J Trop Med Public Health. 2006; 37(Suppl 2):1–128.
Oo TT, Storch V, Becker N. Review of the anopheline mosquitoes of Myanmar. J Vector Ecol. 2004;29:21–40.
Perez-Guzman PN, Carlos Junior Alcantara L, Obolski U, de Lima MM, Ashley EA, Smithuis F, et al. Measuring mosquito-borne viral suitability in Myanmar and implications for local zika virus transmission. PLoS Curr. 2018; 10:ecurrents.outbreaks.7a6c64436a3085ebba37e5329ba169e6.
Kibret S, Wilson GG, Ryder D, Tekie H, Petros B. Malaria impact of large dams at different eco-epidemiological settings in Ethiopia. Trop Med Health. 2017;45:4.
Adugna T, Getu E, Yewhalaw D. Species diversity and distribution of Anopheles mosquitoes in Bure district Northwestern Ethiopia. Heliyon. 2020;6:e05063.
Franklin DC, Whelan PI. Tropical mosquito assemblages demonstrate ‘textbook’ annual cycles. PLoS ONE. 2009;4:e8296.
St Laurent B, Oy K, Miller B, Gasteiger EB, Lee E, Sovannaroth S, et al. Cow-baited tents are highly effective in sampling diverse Anopheles malaria vectors in Cambodia. Malar J. 2016;15:440.
Oo TT, Kaiser A, Becker N. Illustrated keys to the anopheline mosquitoes of Myanmar. J Vector Ecol. 2006;31:9–16.
Wooden J, Kyes S, Sibley CH. PCR and strain identification in Plasmodium falciparum. Trends Parasitol. 1993;9:303–5.
Garros C, Koekemoer LL, Coetzee M, Coosemans M, Manguin S. A single multiplex assay to identify major malaria vectors within the African Anopheles funestus and the Oriental An. minimus groups. Am J Trop Med Hyg. 2004;70:583–90.
Hempolchom C, Otsuka Y, Baimai V, Thongsahuan S, Saeung A, Taai K, et al. Development of a multiplex PCR assay for the identification of eight species members of the Thai Hyrcanus group (Diptera: Culicidae). Appl Entomol Zool. 2013;48:469–76.
Walton C, Somboon P, Harbach RE, Zhang S, Weerasinghe I, O’loughlin SM, et al. Molecular identification of mosquito species in the Anopheles annularis group in southern Asia. Med Vet Entomol. 2007;21:30–5.
Brosseau L, Udom C, Sukkanon C, Chareonviriyaphap T, Bangs MJ, Saeung A, et al. A multiplex PCR assay for the identification of five species of the Anopheles barbirostris complex in Thailand. Parasit Vectors. 2019;12:223.
Beebe NW, Saul A. Discrimination of all members of the Anopheles punctulatus complex by polymerase chain reaction–restriction fragment length polymorphism analysis. Am J Trop Med Hyg. 1995;53:478–81.
Zhong D, Hemming-Schroeder E, Wang X, Kibret S, Zhou G, Atieli H, et al. Extensive new Anopheles cryptic species involved in human malaria transmission in western Kenya. Sci Rep. 2020;10:16139.
Syafruddin D, Hidayati AP, Asih PB, Hawley WA, Sukowati S, Lobo NF. Detection of 1014F kdr mutation in four major Anopheline malaria vectors in Indonesia. Malar J. 2010;9:315.
Abbott WS. A method of computing the effectiveness of an insecticide. 1925. J Am Mosq Control Assoc. 1987; 3:302–3.
Finney DJ. Probit analysis. J Am Pharm Assoc Am Pharm Assoc (Baltim). 1952;41:627.
Mekapogu AR. Finney's probit analysis spreadsheet calculator. 2021. https://probitanalysis.wordpress.com/. Accessed 26 Mar 2022.
Editorial. Shifting habitats. Nat Clim Chang. 2020; 10:377.
Durnez L, Coosemans M. Residual transmission of malaria: An old issue for new approaches. In: Manguin S, editor. Anopheles mosquitoes - New insights into malaria vectors. Rijeka: InTech; 2013.
Carnevale P, Manguin S. Review of issues on residual malaria transmission. J Infect Dis. 2021;223:S61-80.
Ismail IA, Notananda V, Schepens J. Studies on malaria and responses of Anopheles balabacensis balabacensis and Anopheles minimus to DDT residual spraying in Thailand. Acta Trop. 1975;32:206–31.
Li ZZ, Zhang MC, Wus YG, Zhong BL, Lin GY, Huang H. Trial of deltamethrin impregnated bed nets for the control of malaria transmitted by Anopheles sinensis and Anopheles anthropophagus. Am J Trop Med Hyg. 1989;40:356–9.
Zhang Z, Yang C. Application of deltamethrin-impregnated bednets for mosquito and malaria control in Yunnan, China. Southeast Asian J Trop Med Public Health. 1996;27:367–71.
Reisen WK, Pradhan SP, Shrestha JP, Shrestha SL, Vaidya RG, Shrestha JD. Anopheline mosquito (Diptera: Culicidae) ecology in relation to malaria transmission in the inner and outer terai of Nepal, 1987–1989. J Med Entomol. 1993;30:664–82.
Ismail IA, Phinichpongse S, Boonrasri P. Responses of Anopheles minimus to DDT residual spraying in a cleared forested foothill area in central Thailand. Acta Trop. 1978;35:69–82.
Tananchai C, Tisgratog R, Juntarajumnong W, Grieco JP, Manguin S, Prabaripai A, et al. Species diversity and biting activity of Anopheles dirus and Anopheles baimaii (Diptera: Culicidae) in a malaria prone area of western Thailand. Parasit Vectors. 2012;5:211.
Trung HD, Bortel WV, Sochantha T, Keokenchanh K, Briët OJ, Coosemans M. Behavioural heterogeneity of Anopheles species in ecologically different localities in Southeast Asia: a challenge for vector control. Trop Med Int Health. 2005;10:251–62.
Morgan K, Somboon P, Walton C. Understanding Anopheles diversity in Southeast Asia and its applications for malaria control. In: Manguin S, editor. Anopheles mosquitoes - New insights into malaria vectors. Rijeka: Intech; 2013.
Zhang S, Guo S, Feng X, Afelt A, Frutos R, Zhou S, et al. Anopheles vectors in mainland China while approaching malaria elimination. Trends Parasitol. 2017;33:889–900.
Wilai P, Namgay R, Made Ali RS, Saingamsook J, Saeung A, Junkum A, et al. A multiplex PCR based on mitochondrial COI sequences for identification of members of the Anopheles barbirostris Complex (Diptera: Culicidae) in Thailand and other countries in the region. Insects. 2020;11:409.
Phattanawiboon B, Jariyapan N, Mano C, Roytrakul S, Paemanee A, Sor-Suwan S, et al. Salivary gland proteome during adult development and after blood feeding of female Anopheles dissidens mosquitoes (Diptera: Culicidae). PLoS ONE. 2016;11:e0163810.
Thongsahuan S, Baimai V, Junkum A, Saeung A, Min GS, Joshi D, et al. Susceptibility of Anopheles campestris-like and Anopheles barbirostris species complexes to Plasmodium falciparum and Plasmodium vivax in Thailand. Mem Inst Oswaldo Cruz. 2011;106:105–12.
Kanojia PC, Shetty PS, Geevarghese G. A long-term study on vector abundance & seasonal prevalence in relation to the occurrence of Japanese encephalitis in Gorakhpur district Uttar Pradesh. Indian J Med Res. 2003;117:104–10.
Mourya DT, Ilkal MA, Mishra AC, Jacob PG, Pant U, Ramanujam S, et al. Isolation of Japanese encephalitis virus from mosquitoes collected in Karnataka state, India from 1985 to 1987. Trans R Soc Trop Med Hyg. 1989;83:550–2.
Hallin Z, Dengyun Z, Huafang S, Zhuqing M, Zhengda G, Jiayu Z, et al. The natural infection rate of mosquitoes by Japanese Encephalitis B Virus in Yunnan Province, China. In: Dengue Bulletin. WHO Regional Office for South-East Asia.1992. https://apps.who.int/iris/handle/10665/146039. Accessed 26 Mar 2022.
Saeung A, Hempolchom C, Baimai V, Thongsahuan S, Taai K, Jariyapan N, et al. Susceptibility of eight species members in the Anopheles hyrcanus group to nocturnally subperiodic Brugia malayi. Parasit Vectors. 2013;6:5.
Amerasinghe PH, Amerasinghe FP, Konradsen F, Fonseka KT, Wirtz RA. Malaria vectors in a traditional dry zone village in Sri Lanka. Am J Trop Med Hyg. 1999;60:421–9.
Gingrich JB, Weatherhead A, Sattabongkot J, Pilakasiri C, Wirtz RA. Hyperendemic malaria in a Thai village: dependence of year-round transmission on focal and seasonally circumscribed mosquito (Diptera: Culicidae) habitats. J Med Entomol. 1990;27:1016–26.
Sugiarto, Kesumawati Hadi U, Soviana S, Hakim L. Confirmation of Anopheles peditaeniatus and Anopheles sundaicus as malaria vectors (Diptera: Culicidae) in Sungai Nyamuk village, Sebatik Island North Kalimantan, Indonesia using an enzyme-linked immunosorbent assay. J Med Entomol. 2016; 53:1422–4.
Prakash A, Walton C, Bhattacharyya DR, Mohapatra PK, Mahanta J. Characterization of ITS2 rDNA of Anopheles philippinensis and Anopheles nivipes (Diptera: Culicidae) from north-east India. Southeast Asian J Trop Med Public Health. 2006;37:1134–8.
Alam MS, Khan MG, Chaudhury N, Deloer S, Nazib F, Bangali AM, et al. Prevalence of anopheline species and their Plasmodium infection status in epidemic-prone border areas of Bangladesh. Malar J. 2010;9:15.
Alam MS, Al-Amin HM, Khan WA, Haque R, Nahlen BL, Lobo NF. Preliminary report of pyrethroid resistance in Anopheles vagus, an important malaria vector in Bangladesh. Am J Trop Med Hyg. 2020;103:810–1.
World Health Organization. WHO malaria threats map. Geneva. Switzerland: WHO; 2021.
Xu T, Zhong D, Tang L, Chang X, Fu F, Yan G, et al. Anopheles sinensis mosquito insecticide resistance: comparison of three mosquito sample collection and preparation methods and mosquito age in resistance measurements. Parasit Vectors. 2014;7:54.
Oliver SV, Brooke BD. The effect of multiple blood-feeding on the longevity and insecticide resistant phenotype in the major malaria vector Anopheles arabiensis (Diptera: Culicidae). Parasit Vectors. 2014;7:390.
Machani MG, Ochomo E, Sang D, Bonizzoni M, Zhou G, Githeko AK, et al. Influence of blood meal and age of mosquitoes on susceptibility to pyrethroids in Anopheles gambiae from Western Kenya. Malaria J. 2019;18:112.
Verhaeghen K, Van Bortel W, Trung HD, Sochantha T, Keokenchanh K, Coosemans M. Knockdown resistance in Anopheles vagus, An. sinensis, An. paraliae and An. peditaeniatus populations of the Mekong region. Parasit Vectors. 2010;3:59.
Riveron JM, Tchouakui M, Mugenzi L, Menze B, Chiang M, Wondji CS. Insecticide resistance in malaria vectors: An update at a global scale. In: Manguin S, editor. Towards malaria elimination - A leap forward. Intech; 2018.
Nikou D, Ranson H, Hemingway J. An adult-specific CYP6 P450 gene is overexpressed in a pyrethroid-resistant strain of the malaria vector Anopheles gambiae. Gene. 2003;318:91–102.
Stevenson BJ, Bibby J, Pignatelli P, Muangnoicharoen S, O’Neill PM, Lian LY, et al. Cytochrome P450 6M2 from the malaria vector Anopheles gambiae metabolizes pyrethroids: Sequential metabolism of deltamethrin revealed. Insect Biochem Mol Biol. 2011;41:492–502.
Mitchell SN, Stevenson BJ, Müller P, Wilding CS, Egyir-Yawson A, Field SG, et al. Identification and validation of a gene causing cross-resistance between insecticide classes in Anopheles gambiae from Ghana. Proc Natl Acad Sci USA. 2012;109:6147–52.
Rongnoparut P, Boonsuepsakul S, Chareonviriyaphap T, Thanomsing N. Cloning of cytochrome P450, CYP6P5, and CYP6AA2 from Anopheles minimus resistant to deltamethrin. J Vector Ecol. 2003;28:150–8.
Rodpradit P, Boonsuepsakul S, Chareonviriyaphap T, Bangs MJ, Rongnoparut P. Cytochrome P450 genes: molecular cloning and overexpression in a pyrethroid-resistant strain of Anopheles minimus mosquito. J Am Mosq Control Assoc. 2005;21:71–9.
Overgaard HJ, Sandve SR, Suwonkerd W. Evidence of anopheline mosquito resistance to agrochemicals in northern Thailand. Southeast Asian J Trop Med Public Health. 2005;36:152–7.
Kawada H, Oo SZM, Thaung S, Kawashima E, Maung YNM, Thu HM, et al. Co-occurrence of point mutations in the voltage-gated sodium channel of pyrethroid-resistant Aedes aegypti populations in Myanmar. PLoS Negl Trop Dis. 2014;8:e3032.
Acknowledgements
We thank our field team of the Southeast Asia ICEMR project in Myanmar for collecting mosquito samples. We also thank the editor and the anonymous reviewers for their constructive comments that improved this manuscript. This work is supported by the National Institutes of Health (NIH) Grant U19AI089672.
Author information
Authors and Affiliations
Contributions
DZ, MPK, and PLA contributed to overall study design. MPK, PLA, MMM, and MTS identified study locations, helped to collect mosquito samples, and identified vector species. DZ, MMM, PLA, XW, and QQ conducted mosquito bioassay or genotyping. DZ, XW, GZ, and GY analyzed the data. DZ, LC, JS, GZ, GY, XW, MTS, MPK, and PLA drafted and revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethical approval for the study was obtained from the institutional scientific ethical review board of University of California, Irvine, CA, USA, and the Ethical Review Committee, Department of Medical Research, Ministry of Health and Sports, Yangon, Myanmar. Permission was sought from the chief of each study site. All methods used in this study were performed in accordance with the relevant guidelines and regulations.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Partial sequences showing the kdr L1014 allele (highlighted) in the 22 Anopheles species from northern-central Myanmar. Numbers in parentheses indicate the sample size.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhong, D., Aung, P.L., Mya, M.M. et al. Community structure and insecticide resistance of malaria vectors in northern-central Myanmar. Parasites Vectors 15, 155 (2022). https://doi.org/10.1186/s13071-022-05262-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05262-2