
Abstract
Background
Predators play a critical role in regulating larval mosquito prey populations in aquatic habitats. Understanding predator-prey responses to climate change-induced environmental perturbations may foster optimal efficacy in vector reduction. However, organisms may differentially respond to heterogeneous thermal environments, potentially destabilizing predator-prey trophic systems.
Methods
Here, we explored the critical thermal limits of activity (CTLs; critical thermal-maxima [CTmax] and minima [CTmin]) of key predator-prey species. We concurrently examined CTL asynchrony of two notonectid predators (Anisops sardea and Enithares chinai) and one copepod predator (Lovenula falcifera) as well as larvae of three vector mosquito species, Aedes aegypti, Anopheles quadriannulatus and Culex pipiens, across instar stages (early, 1st; intermediate, 2nd/3rd; late, 4th).
Results
Overall, predators and prey differed significantly in CTmax and CTmin. Predators generally had lower CTLs than mosquito prey, dependent on prey instar stage and species, with first instars having the lowest CTmax (lowest warm tolerance), but also the lowest CTmin (highest cold tolerance). For predators, L. falcifera exhibited the narrowest CTLs overall, with E. chinai having the widest and A. sardea intermediate CTLs, respectively. Among prey species, the global invader Ae. aegypti consistently exhibited the highest CTmax, whilst differences among CTmin were inconsistent among prey species according to instar stage.
Conclusion
These results point to significant predator-prey mismatches under environmental change, potentially adversely affecting natural mosquito biocontrol given projected shifts in temperature fluctuations in the study region. The overall narrower thermal breadth of native predators relative to larval mosquito prey may reduce natural biotic resistance to pests and harmful mosquito species, with implications for population success and potentially vector capacity under global change.
Graphical Abstract

Similar content being viewed by others


Background
Population size in aquatic ecosystems is known to be largely dependent on ecological interactions such as competition and predation pressure [1, 2]. Predation plays a pivotal role in regulating problematic species (e.g. proliferating mosquitoes) through density-mediated effects, whereby population numbers are directly controlled through predatory removal or through indirect, trait-mediated effects such as compromised fecundity, growth rate and longevity of prey [3, 4]. Key mosquito genera (e.g. Aedes, Anopheles, Culex) are of public concern globally, transmitting pathogens that cause common debilitating diseases to humans (e.g. chikungunya, dengue, multiple kinds of encephalitis, elephantiasis, malaria, yellow fever, Zika), livestock (Rift Valley fever) and wildlife (avian malaria, West Nile) [5, 6]. Moreover, mosquitoes are semi-aquatic insects that colonize and develop (egg, larva, pupae) in aquatic habitats across natural, urban and peri-urban environments [7].
Naturally, mosquitoes coexist in aquatic microhabitats with a large faunal community [8], including aquatic predators that curb mosquito populations [9,10,11,12,13,14]. In these environments, both the predator and mosquito prey experience stressful thermal extremes, with variable effects on performance given differential thermal physiological responses [15, 16]. Mosquitoes breed in diverse, often cryptic, aquatic habitats such as rock crevices, phytotelmata (e.g. tree holes), animal hoof prints, artificial containers as well as larger-scale temporary and permanent water bodies [17, 18]. These environments are prone to extreme environmental fluctuations, which are expected to become the new norm in a warming world, typically becoming more intense, prolonged and frequent [19,20,21]. Mosquitoes have also adapted to colonize clean to highly compromised water quality sources, sunlit or shaded and of varying nutrient levels [22]. Predators of mosquitoes can persist in these environments as wholly aquatic organisms, access through aerial dispersal as semi-aquatic predators or be purposefully introduced as agents for desired ecosystem services [23]. The variable utilization of different water bodies by mosquito larvae, and their aquatic predators, has implications for performance of these organisms and ultimately predator-prey interaction outcomes. Understanding these dynamics could thus prove useful in determining the sustainability of natural and augmentative mosquito biological control in aquatic habitats, particularly within the context of changing environments [24].
Predator-prey interaction strengths can be mediated by abiotic environmental factors [25]. Particularly temperature is important in this regard, affecting organismal physiology, ecology, metabolism and overall fitness [26, 27]. Temperature in water bodies is essential as a regulatory mechanism that drives biochemical and physiological processes [28, 29], with implications for behaviour, performance and predator-prey interaction outcomes [30,31,32]. Indeed, empirical studies have shown environmental variability likely affects higher trophic levels, e.g. predators, more significantly than prey [33,34,35]. Furthermore, thermal performance is highly enigmatic and varies among species, ontogeny, age [36] and size [29]. Moreover, natural enemy efficacy also depends on the fate of bottom-up and top-down effects, which have been reported to favour pest and vector species [37]. As such, even slight alterations to temperature can compromise or heighten species fitness, community interactions and structure [38, 39]. This makes predicting the fate of natural enemy effects in the face of climate change highly complex. Additionally, at the autecological level, understanding how organismal critical thermal limits (CTLs) of varied species are affected by oscillating temperatures can be useful for broader ecological inferences, such as interaction dynamics between species [40, 41]. Definitively, CTLs represent temperatures at which an organism stops activity. Ecologically, activity here represents key fitness traits, e.g. mating, swimming and foraging ability [27, 42].
The effects of temperature are critical in determining the fate of trophic interactions under changing environments [43]. However, whilst studies have focused on terrestrial environments [34, 35, 44], to our knowledge, few have concerned aquatic habitats in the context of thermal tolerance, particularly for vector mosquitoes and their predators. Here, we aimed to assess thermal tolerance (lower and upper) of three regionally abundant aquatic mosquito predators (Enithares chinai and Anisops sardea [Insecta: Hemiptera], Lovenula falcifera [Copepoda: Calanoida]) and their vectorially important larval mosquito prey (Aedes aegypti, Anopheles quadriannulatus, Culex pipiens [Diptera: Culicidae]) in a semi-arid subtropical southern African landscape. All three mosquito species are commonly encountered in peri- and urban landscapes of the study region [45,46,47], with Ae. aegypti and Cx. pipiens well-known vectors of various pathogens that cause disease [48,49,50]. Anopheles quadriannulatus is not currently a known vector to human pathogens [51], however, is susceptible to Plasmodium infection [52, 53]. Physiological limits of this species may nevertheless serve as reasonable proxies of congeneric malaria vector species. While increases in extreme temperature events are predicted to be the future norm [19,20,21, 54,55,56], semi-arid southern Africa is projected to be particularly impacted by shifting climatic conditions [57]. We thus hypothesized that: (1) larval thermal tolerance would vary across mosquito species, with container-breeding specialists (principally Ae. aegypti) having the widest thermal window; (2) for predators, the wholly aquatic copepod would have the narrowest thermal window; (3) all predators would have narrower and therefore asynchronized thermal windows compared to the mosquito species.
Materials and methods
Animal collection and maintenance
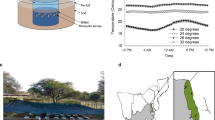
Adult Lovenula falcifera and Anisops sardea were collected from a clay-lined temporary pond, Central District, Botswana (Fig. 2a; 022° 52′ 16.0 S; 027° 47′ 42.7 E), while Enithares chinai were collected from a concrete-lined water pool on the Botswana International University of Science and Technology (BIUST) campus (022° 35′ 46.8 S; 027° 07′ 30.5 E). The predators were housed in separate aerated 3-L plastic containers (covered with a net to prevent winged predator escape), comprising ~2 L of a 50:50 ratio of habitat water and matured tap water (kept for 48 h for dechlorination). These were placed in climate chambers (HPP 260, Memmert GmbH + Co.KG, Germany) set at 28 °C ± 2 and 65 % ± 10 relative humidity under a 12:12 light:dark photocycle. All predator species were fed Cx. pipiens larvae ad libitum. Culex pipiens larvae (accession number: MT741514) originated from egg rafts sampled from a concrete-lined water body situated in BIUST campus (022° 35′ 05.7 S; 027° 06′ 58.7 E). Aedes aegypti larvae (accession number: MK571449) were collected using a 1000 µm mesh net from a 20-L container holding ~10 L rain water in a homestead in Palapye village (022′ 32′ 97.6 S; 027′ 11′ 50.4 E) while An. quadriannulatus larvae (accession number: MT741513) were sourced from stagnant river water near Hogs Creek (022′ 34′ 79.3 S; 028′ 19′ 96.1 E). The larvae were reared, separately according to species, to different instar stages in 3-L plastic containers holding ~2 L matured tap water housed in climate chambers (as above) and fed with crushed rabbit food pellets ad libitum (Westerman’s Premium, Durban, South Africa). Both predators and their prey were kept at similar densities (10 individuals/L in a 3-L container holding ~2 L of a 50:50 ratio of habitat water and matured tap water) to avoid overcrowding effects on thermal fitness [58]. Both the predators and prey were collected between January and February 2020 and experienced similar thermal environments. Prior to all experiments, predators were kept for at least 7 days in laboratory rearing conditions while prey developmental stages were monitored until appropriate instars sizes were reached.
Experimental design
We assessed CTLs in an experimental design with respect to (1) predators (3 species types: L. falcifera, A. sardea, E. chinai) and (2) mosquito larvae (3 species: Cx. pipiens, Ae. aegypti, An. quadriannulatus) across their instar stages (3 stages: mean length ± SE, early [1st instar; 1.5 ± 0.2 mm], intermediate [2nd/3rd instar; 3.0 ± 0.2 mm], late [4th instar; 5.1 ± 0.2 mm]). Critical thermal limits (CTmin and CTmax) were measured randomly across all treatments, following modified protocols by Nyamukondiwa et al. [59]. A set of ten individual organisms at a time were each placed in a series of ten insulated double-jacketed chambers, connected to a programmable water bath (Lauda Eco Gold, Lauda DR.R. Wobser GMBH and Co. KG, Germany). The water bath contained a 1:1 water:propylene glycol ratio to sufficiently cater to sub-zero cooling temperatures. Each ‘organ pipe’ was filled with 50 mL matured tap water to house an individual animal, which was then given 10 min to stabilize at the 28 °C temperature, i.e. equivalent to climate chamber rearing conditions. A thermocouple (type T 36 SWG) connected to a digital thermometer (53/54IIB, Fluke Corp., USA) was inserted into a central ‘organ pipe’ (the control pipe) to monitor the water temperature experienced by the test animals. Temperature was ramped up (CTmax) or down (CTmin) (from benign 28 °C) at 0.25 °C min−1 following described protocols [59, 60]. The experiment was repeated twice (i.e. two runs per set of ten animals; n = 20) per treatment in keeping with Nyamukondiwa and Terblanche [61]. Here, we defined CTLs as the temperature at which an animal lost coordinated muscle function or responses resulting from a slight prodding using a thermally inert object congruous to Nyamukondiwa et al. [59]. For CTmax, this loss of coordinated muscle function always coincides with lethal temperatures and mortality such that recovery is not possible. However, for CTmin, recovery often occurs and thus the trait is not always lethal [27, 40].
Microclimate data recordings
Microclimate temperature data were recorded from a sunlit (1) temporary clay-lined pond (123 m length × 95 m width × 1.5 m depth; 022° 52′ 16.0 S; 027° 47′ 42.7 E) and (2) temporary rock pool (2.4 m length × 1.7 m width × 13 cm depth; 22° 35′ 46.07″ S; 27° 07′ 16.46″ E), using programmable data logger probes and software (HOBOware Pro, version 3.7.16, Bourne, MA, USA) (0.5 °C accuracy; 1 h sampling frequency) during the period between August 2019 and February 2020. This was to determine the thermal fluctuations in these temporary wetland aquatic habitats that represent the dominant available natural breeding habitats for predators and mosquito prey in the region [62]. These temporary wetlands have been observed to host all larval species and the three predators tested. A data logger probe was placed on sediments at the bottom of the pond/pool during the dry period and monitored throughout the wet phases to reflect temperature variations associated with these environments, although species abundances were not monitored. The mean temperature for the wet phase, between both habitats, was used as the habitat temperature (Thab).
Statistical analysis
Data analysis was performed using R, version 3.6.3. The residuals were first checked for normality and variance homogeneity using Shapiro-Wilks and Levene’s tests, respectively, and were found to violate normality and variance homogeneity assumptions. Therefore, a one-way Kruskal-Wallis nonparametric test was employed. The CTLs (CTmax and CTmin) were considered separately as dependent variables, while the different prey species, instar stages and predator species were the independent factors. Statistically significant effects were examined pairwise post hoc using Dunn's test. We thus fit two models to our data, whereby the two CTLs (CTmax or CTmin) of predators and prey (early, intermediate and late instars) were compared.
The thermal breadths (warming tolerance [WT] and cooling tolerance [CT]) for the predators and the larvae were calculated as described by Machekano et al. [63]:
and,
where CTmax and CTmin were the CTLs for the predators and larval prey whereas Thab was the mean daily habitat (clay-lined pond and rock pool data combined mean) temperature, reflective of natural conditions likely experienced by both predators and the larval prey in these dominant temporary wetland environments. The standard errors (SE) between CTLs and the Thab for each species were also calculated.
Results
Critical thermal maxima (CTmax) differed significantly among predators and prey across instar stages (χ2 = 229.43, df = 11, p < 0.001) (Fig. 1a). Among predators, L. falcifera exhibited the lowest CTmax, which was the highest for the notonectids E. chinai and A. sardea, yet only significantly for E. chinai. Among prey, Ae. aegypti generally consistently exhibited the highest CTmax, whilst An. quadriannulatus was the lowest and Cx. pipiens intermediate. Whilst the CTmax of first instar prey were all statistically similar, Ae. aegypti exhibited significantly higher CTmax than An. quadriannulatus at intermediate and late instar stages, but was statistically similar to Cx. pipiens. Within species, prey responses were in turn dependent on their instar stage, with first instar stages consistently exhibiting the significantly lowest CTmax, whilst the later instar stages were not statistically different (Fig. 1a). As such, the extent of predator-prey CTmax mismatch was greatest when considering later instar stages.

Critical thermal limits (a CTmax and b CTmin) of three mosquito predators (Lovenula falcifera [Lf], Enithares chinai [Ec] and Anisops sardea [As]) and three larval prey (Aedes aegypti [Aa], Anopheles quadriannulatus [Aq] and Culex pipiens [Cp]) at the first, second/third and fourth instar stages. Group medians with different letters are statistically different from each other (n = 20 per experimental group). In the boxplots, the box gives the interquartile ranges and the whiskers show the largest and smallest values up to 1.5 × interquartile range. The horizontal line in each box shows the median
Similarly, CTmin differed significantly across predator and prey types (χ2 = 233.63, df = 11, p < 0.001) (Fig. 1b). Among predators, the notonectid E. chinai exhibited the greatest cold tolerance (lowest CTmin temperature) compared to the other species, with CTmin significantly lower than L. falcifera or A. sardea. For prey, responses were inconsistent among species considering instar stage. For first instars, CTmin values were always statistically similar, whilst for second–third and fourth instars Cx. pipiens CTmin was the highest, and significantly higher than An. quadriannulatus. Within prey species, first instar stages consistently exhibited significantly higher cold tolerance (lower CTmin), except between the first and second–third instars of An. quadriannulatus (Fig. 1b). Accordingly, in contrast to CTmax, cold tolerance mismatches between predators and prey tended to be greatest for early instars.
The mean daily Thab obtained from the wet phases of the clay-lined and rock pool temporary wetland was 25.2 °C (minimum: 21.9 °C; maximum: 29.7 °C), and this was further used to determine the thermal breadths of mosquito predators and the larval prey (Table 1; Fig. 2). These results showed an overall trend of higher CTLs and wider thermal breadths for mosquito larval prey compared to their predators (Table 1). The notonectid E. chinai had the greatest thermal breadths compared to the other predator species at both temperature extremes. In turn, the WT of L. falcifera was narrower than that of A. sardea, but the same species (L. falcifera) exhibited a wider CT (Table 1). Amongst larval mosquitoes, Ae. aegypti always had wider WT thermal breadths at matched instar stages. For CT, Ae. aegypti had the greatest breadths for the first instar stage alone; second/third and fourth instar stages were greater than in Cx. pipiens, but lower than in An. quadriannulatus. For all species, first instar stages had a narrower WT, but broader CT than later instars. Generally, aside from first instars, An. quadriannulatus had a narrower thermal breadth for WT and a wider CT breadth than Cx. pipiens (Table 1). Enithares chinai was an exception among predators, with wider WT and CT that was more similar to several larval mosquito stages (Table 1).

Microclimatic data showing mean daily maximum (Max), minimum (Min) and average (Ave) temperature (°C) of a a clay-lined pond and b a rock pool during their wet phase between August 2019 and February 2020
Discussion
Biological control of mosquito larvae using aquatic predators is a sustainable and environmentally friendly approach in reducing disease vector populations [64, 65]. Dissociations in thermal tolerances between predators and prey may, however, adversely affect predator-prey interactions leading to compromised foraging impacts. Our results showed varied thermal tolerance amongst mosquito larval prey, with early instars exhibiting compromised heat tolerance but having higher cold tolerance. Furthermore, the container-breeding specialist (Ae. aegypti) always had a wider WT (thermal breadth), indicating greater tolerance of heat stress. The wholly aquatic copepods had narrower thermal breadth, which was more compromised in terms of WT compared to other semi-aquatic notonectids. Overall, there was a mismatch between predators and the mosquito larval prey at both low- and high-temperature extremes. Predators showed lower thermal fitness and activity windows, suggesting that they may be impacted by temperature extremes earlier and more negatively than their mosquito prey. In keeping with Hunsicker et al. [66], these results potentially represent a loss of predator optimal ecosystem services provision and a mosquito larval prey proliferation cost in aquatic ecosystems with shifting environments. This has subsequent negative implications on increased vector population and associated disease risks amid rapidly shifting climate environments [67].
Aedes aegypti had a wider thermal window compared to the other mosquito species, having higher thermal limits to activity especially on the warming extreme. The species is highly invasive in tropical and subtropical regions of the world [46]. This could be linked to its behavioural profile of associating with thermally heterogeneous transient or temporary microhabitats, although CTLs may vary across mosquito species, space and methodological context [68, 69]. The species is found in human dwellings and thrives in diverse artificial containers as breeding habitats [70, 71]. Given that this species specializes in small water body utilization for oviposition (e.g. tyres, tins, gutters, flowerpots), Ae. aegypti have likely adapted (e.g. through transgenerational plasticity; [72]) and have the ability to withstand extreme temperatures in small environments associated with less thermal inertia. With the highest thermal tolerance and breadth compared to its predators and other mosquito prey larvae tested, Ae. aegypti has potential to thrive in conditions where other vector species would otherwise be compromised. Moreover, during this time, mismatch between Ae. aegypti and its predators could allow a rapid mosquito population growth owing to a lack of natural biotic suppression. This may further promote invasion success across the globe, with likely increased risks of associated pathogens and their diseases [73]. This has implications for emerging and reemerging diseases in vulnerable societies across rural-urban spheres with overall negative implications on public health and livelihoods. However, additional population regulatory effects of increased intraspecific competition should not be ignored.
Amongst the prey species assessed here, generally, Cx. pipiens was intermediate heat (CTmax) and warming tolerant. Culex pipiens can breed in highly compromised water quality habitats (e.g. water treatment plants, sewage ponds, septic tanks) [74], and must oviposit directly onto water in contrast to Ae. aegypti, which produce dormant eggs. These environments provide a thermal variability cushion to the aquatic life stages compared to relatively smaller microhabitats that are likely not as buffered. Conversely, the results showed that An. quadriannulatus was generally the most cold tolerant, and generally least heat tolerant, except at first instars. Activity at low temperatures means An. quadriannulatus can still maintain key life history traits (e.g. swimming, foraging and development) during winter periods. Although the largely zoophilic An. quadriannulatus is not presently considered a malaria vector to humans [51], the species is susceptible to Plasmodium infection [52, 53]. This may have future implications for spatial emerging–re-emerging infections to humans, given potential shifts in behavioural [75, 76] and feeding preferences associated with such species [77, 78]. While An. quadriannulatus does not vector malaria, current thermal activity limit results are likely reasonable ecological proxies for more competent congeneric malaria vectors. To this end, we recommend further research considering thermal profiles of malaria-implicated vectors regionally, and diverse mosquito predators, alongside actual predator-prey performance experiments, in changing climatic conditions owing to the risk of emerging and reemerging diseases [79].
The results of this study generally reported that mosquitoes had wider thermal windows than their predators. Owing to this temperature regime mismatch, mosquito larvae are likely to proliferate against their predators, suggesting reduced efficacy of predator-prey interaction under shifting aquatic environments [80]. Enithares chinai heat tolerance (CTmax) however synchronized with first instar stages of mosquitoes. However, Buxton et al. [14] showed that, at this size, there are some prey refuge effects, with the notonectid consuming second/third and fourth instar larvae more efficiently. This suggests that any observed thermal synchrony between E. chinai and first instar larvae may still offer limited biocontrol potential. Moreover, this study found that the wholly aquatic copepods are highly likely to be impacted in the warming temperature regimes as opposed to the semi-aquatic, air breathing predatory notonectids and the larval prey. Copepods’ life history traits are negatively affected by escalating temperatures as demonstrated by Lee et al. [81]. This has an overall implication for biocontrol in shifting environments, with the need to further identify diverse species, additively combine predators for sustainable mosquito regulation [82] and monitor (physiological-mediated traits) in time and space given the varied habitat temperature exposure. Although the species used here were a representation of a single location, the direction of the conclusions drawn is likely more broadly applicable for the studied taxa, all of which are widespread in southern Africa, and in certain instances other parts of Africa (e.g. Lovenula spp.) or even globally (e.g. Ae. aegypti). It would, however, be useful for future studies to investigate temperature effects across space and closely related taxa in search of unifying patterns. Similarly, future studies should also investigate predator-prey thermal interaction effects using more controlled predator and prey developmental stages (juveniles), since thermal fitness might vary across insect ontogeny. The relationship between CTLs and optimal temperatures also needs to be explored, within the context of predator-prey behaviour and interaction outcomes.
Based on the predator-prey dissociations exhibited here, whether predator thermal fitness can co-evolve symmetrically with their prey in changing aquatic environments remains a key question [83]. Field data have shown that, during the hydroperiod, performance of predators and mosquito larvae was within thermal breadths with no compromised activity on both extremes. Although pond temperature extremes could be seasonal and short-lived because of hydroperiods, more investigations on predator life history traits are warranted, especially the dormant egg physiology and hatching phenology consequent to the extreme hot and dry phases evidenced from clay-lined ponds and rock pools [84, 85]. Furthermore, the current study only measured basal thermal traits, and further exploration of other physiological-mediated traits driving the fate of predator-prey interactions within aquatic heterogeneous systems is needed. In particular, behavioural microclimate selection often drives invertebrate vulnerability to shifting climates [86]. Thus, the role of behaviour in modulating thermal fitness and how this may reshape predator-prey interactions also ought to be investigated.
Critical thermal limits have widely been used in assessing insect responses to climate change [27, 87, 88], including aquatic invertebrates [60, 89]. As such, these assays have gained attention in explaining the fate of trophic interactions, e.g. coevolved predator-prey and host-natural enemy association under high temperature stress [34, 35]. It is nevertheless critical to directly investigate and establish predator-prey interaction dynamics under varied temperatures [90, 91] and how optimal performance may relate to CTLs. This will allow for a more effective assessment of constraints of biological control associated with thermal stress prior to organismal loss of physiological function, e.g. through thermal performance curves (see discussions in [37]). While CTLs are only a measure of the fate of an organism at extreme temperatures [92, 93], they can still be useful in predictive models associated with population dynamics [27, 37, 94]. In addition, the outcomes have ecological implications not only for the long-term effects of global change [95], but also for the short- to medium term whereby organisms may be exposed to unexpected extreme acute temperatures, such as cold snaps and heat waves [21, 67, 96]. Although some organisms can survive these harsh conditions, some may succumb to them, with implications for community dynamics. In this context, although feeding rates of mosquito natural enemies can relate positively to temperature [97], as temperatures exceed thermal optimums a unimodal feeding relationship may arise [98, 99], which could alleviate prey from predation pressure and promote their proliferation.
Ectothermic organisms also often adaptively remodel their phenotypes to better survive stressful environments through plasticity [89, 100], a near ubiquitous mechanism in insects. Thus, it is likely that the limited thermal tolerance and breadths for predators recorded here may be compensated for through higher phenotypic adjustments [89, 101,102,103]. Nevertheless, the presence, magnitude and extent to which phenotypic plasticity may cushion organisms against climate change remains debatable [103, 104]. Thus, the exact extent to which plasticity may change the direction of interactions reported here warrants future investigation. Unraveling these physiological mechanisms will foster improved understanding of predator longevity and success, paramount to sustaining mosquito biological control under climate change. This is significant in maintaining the integrity and efficacy of biocontrol agent’s life history traits in aquatic habitats given the increased extreme temperature means and fluctuations with high intensities under global change [19, 20, 67, 104, 105].
Conclusion
Our results demonstrate a mismatch of thermal activity limits (CTLs) and thermal breadths (WT and CT) between key predators and their mosquito prey. Larval mosquitoes had significantly higher activity limits and wider thermal windows relative to their predator antagonists. This thermal mismatch may mean asynchrony in predator-prey phenologies in shifting habitats, consequently altering the aquatic ecosystem trophic community structures and functioning. Predators are thus projected to reduce interactive foraging strength towards an increasingly thermally fit prey, giving vectors an advantage by proliferating in aquatic habitats. Implications for increasing temperature stress remain a challenge in predicting mosquito biocontrol using natural enemies, and more so under shifting aquatic habitats. In future research, the role of plastic thermal compensation in building resilience under climate change should be explored. Conservation of mosquito predators, coupled with the use of other complementary biological control strategies in an integrated approach may help reduce disease vector populations and associated public health concerns.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Holomuzki JR, Feminella JW, Power ME. Biotic interactions in freshwater benthic habitats. J N Am Benthol Soc. 2010;29:220–44.
Arribas R, Touchon JC, Gomez-Mestre I. Predation and competition differentially affect the interactions and trophic niches of a Neotropical amphibian guild. Front Ecol Evol. 2018;6:28.
Zuharah WF, Lester PJ. The influence of aquatic predators on mosquito abundance in animal drinking troughs in New Zealand. J Vector Ecol. 2010;35:347–53.
Roux O, Robert V. Larval predation in malaria vectors and its potential implication in malaria transmission: an overlooked ecosystem service? Parasite Vector. 2019;12:217.
Lebl K, Zittra C, Silbermayr K, Obwaller A, Berer D, Brugger K, Rubel F. Mosquitoes (Diptera: Culicidae) and their relevance as disease vectors in the city of Vienna, Austria. Parasitol Res. 2015;114:707–13.
Tandina F, Doumbo O, Traoré SF, Parola P, Robert V. Mosquitoes (Diptera: Culicidae) and mosquito–borne diseases in Mali, West Africa. Parasite Vector. 2018;11:1–12.
Foster WA, Walker ED. Mosquitoes (Culicidae). In: Medical and veterinary entomology. Cambridge: Academic Press; 2019. p. 261–325.
Kocher DK, Jamwal S, Kansal M, Bakshi DK, Kocher DK. Coexistence of copepods and mosquito larvae in different standing water bodies of Punjab, India. Scholars Acad J Biosci. 2014;2:640–2.
Chandra G, Bhattacharjee I, Chatterjee SN, Ghosh A. Mosquito control by larvivorous fish. Indian J Med Res. 2008;127:13.
Ohba SY, Huynh TT, Kawada H, Le LL, Ngoc HT, Hoang SL, Takagi M. Heteropteran insects as mosquito predators in water jars in southern Vietnam. J Vector Ecol. 2011;36:170–4.
Acquah-Lamptey D, Brandl R. Effect of a dragonfly (Bradinopyga strachani Kirby, 1900) on the density of mosquito larvae in a field experiment using mesocosms. Web Ecol. 2018;18:81–9.
Cuthbert RN, Dalu T, Wasserman RJ, Callaghan A, Weyl OL, Dick JT. Calanoid copepods: an overlooked tool in the control of disease vector mosquitoes. J Med Entomol. 2018;55:1656–8.
Buxton M, Cuthbert RN, Dalu T, Nyamukondiwa C, Wasserman RJ. Complementary impacts of heterospecific predators facilitate improved biological control of mosquito larvae. Biol Control. 2020a;144:104216.
Buxton M, Cuthbert RN, Dalu T, Nyamukondiwa C, Wasserman RJ. Predator density modifies mosquito regulation in increasingly complex environments. Pest Manag Sci. 2020b. https://doi.org/10.1002/ps.5746.
Abrahams MV, Mangel M, Hedges K. Predator-prey interactions and changing environments: who benefits? Philos Trans R Soc B. 2007;362:2095–104.
Broitman BR, Szathmary PL, Mislan KAS, Blanchette CA, Helmuth B. Predator-prey interactions under climate change: the importance of habitat vs body temperature. Oikos. 2009;118:219–24.
Karuitha M, Bargul J, Lutomiah J, Muriu S, Nzovu J, Sang R. Mbogo CLarval habitat diversity and mosquito species distribution along the coast of Kenya. Wellcome Open Res. 2019;4:175.
Wilke AB, Chase C, Vasquez C, Carvajal A, Medina J, Petrie WD, Beier JC. Urbanization creates diverse aquatic habitats for immature mosquitoes in urban areas. Sci Rep. 2019;9:1–11.
Meehl GA, Tebaldi C. More intense, more frequent, and longer lasting heat waves in the 21st century. Science. 2004;305:994–7.
Thuiller W. Climate change and the ecologist. Nature. 2007;448:550–2.
Stillman JH. Heat waves, the new normal: summertime temperature extremes will impact animals, ecosystems, and human communities. Physiology. 2019;34:86–100.
Buxton M, Cuthbert RN, Dalu T, Nyamukondiwa C, Wasserman RJ. Cattle–induced eutrophication favours disease–vector mosquitoes. Sci Total Environ. 2020c;715:136952.
Dida GO, Gelder FB, Anyona DN, Abuom PO, Onyuka JO, Matano AS, Ofulla AV. Presence and distribution of mosquito larvae predators and factors influencing their abundance along the Mara River, Kenya and Tanzania. Springerplus. 2015;4:136.
Scholes RJ. Climate change and ecosystem services. Wiley Interdiscip Rev Clim Change. 2016;7:537–50.
Johansson F, Brodin T. Effects of fish predators and abiotic factors on dragonfly community structure. J Freshw Ecol. 2003;18:415–23.
Gillooly JF, Brown JH, West GB, Savage VM, Charnov EL. Effects of size and temperature on metabolic rate. Science. 2001;293:2248–51.
Chown SL, Nicolson S. Insect physiological ecology: mechanisms and patterns. Oxford: University Press; 2004.
Teoh ML, Chu WL, Phang SM. Effect of temperature change on physiology and biochemistry of algae: a review. Malays J Sci. 2010;29:82–97.
Burton T, Einum S. The old and the large may suffer disproportionately during episodes of high temperature: evidence from a keystone zooplankton species. Conserv Physiol. 2020;8:coaa038.
Domenici P, Claireaux G, McKenzie DJ. Environmental constraints upon locomotion and predator-prey interactions in aquatic organisms: an introduction. Philos Trans R Soc B. 2007;362:1929–36.
Vucic-Pestic O, Roswitha B, Ehnes RB, Rall BC, Brose U. Warming up the system: higher predator feeding rates but lower energetic efficiencies. Glob Chang Biol. 2011;17:1301–10.
Wasserman RJ, Vink TJF, Dalu T, Froneman PW. Fish predation regimes modify benthic diatom community structures: experimental evidence from an in situ mesocosm study. Austral Ecol. 2015;40:806–15.
Hance T, van Baaren J, Vernon P, Boivin G. Impact of extreme temperatures on parasitoids in a climate change perspective. Annu Rev Entomol. 2007;52:107–26.
Machekano H, Mvumi BM, Nyamukondiwa C. Loss of coevolved basal and plastic responses to temperature may underlie trophic level host–parasitoid interactions under global change. Biol Control. 2018;118:44–54.
Mutamiswa R, Chidawanyika F, Nyamukondiwa C. Thermal plasticity potentially mediates the interaction between host Chilo partellus Swinhoe (Lepidoptera: Crambidae) and endoparasitoid Cotesia flavipes Cameron (Hymenoptera: Braconidae) in rapidly changing environments. Pest Manag Sci. 2018;74:1335–45.
Bowler K, Terblanche JS. Insect thermal tolerance: what is the role of ontogeny, ageing and senescence? Biol Rev. 2008;83:339–55.
Chidawanyika F, Mudavanhu P, Nyamukondiwa C. Global climate change as a driver of bottom-up and top-down factors in agricultural landscapes and the fate of host–parasitoid interactions. Front Ecol Evol. 2019;7:1–13.
Jiang L, Morin PJ. Temperature-dependent interactions explain unexpected responses to environmental warming in communities of competitors. J Anim Ecol. 2004;73:569–76.
Deutsch CA, Tewksbury JJ, Huey RB, Sheldon KS, Ghalambor CK, Haak DC, Martin PR. Impacts of climate warming on terrestrial ectotherms across latitude. Proc Natl Acad Sci. 2008;105:6668–72.
Terblanche JS, Hoffmann AA, Mitchell KA, Rako L, le Roux PC, Chown SL. Ecologically relevant measures of tolerance to potentially lethal temperatures. J Exp Biol. 2011;214:3713–25.
Andersen JL, Manenti T, Sørensen JG, MacMillan HA, Loeschcke V, Overgaard J. How to assess Drosophila cold tolerance: chill coma temperature and lower lethal temperature are the best predictors of cold distribution limits. Funct Ecol. 2015;29:55–65.
Mutamiswa R, Machekano H, Chidawanyika F, Nyamukondiwa C. Thermal resilience may shape population abundance of two sympatric congeneric Cotesia species (Hymenoptera: Braconidae). PLoS ONE. 2018;13:e0191840.
Thomas CD, Cameron A, Green RE, Bakkenes M, Beaumont LJ, Collingham YC, Hughes L. Extinction risk from climate change. Nature. 2004;427:145–8.
Agosta SJ, Joshi KA, Kester KM. Upper thermal limits differ among and within component species in a tritrophic host–parasitoid–hyperparasitoid system. PLoS ONE. 2018;13:e0198803.
Cornel AJ, Lee Y, Almeida APG, Johnson T, Mouatcho J, Venter M, Braack L. Mosquito community composition in South Africa and some neighboring countries. Parasite Vector. 2018;11:331.
Buxton M, Lebani K, Nyamukondiwa C, Wasserman RJ. First record of Aedes (Stegomyia) aegypti (Linnaeus, 1762) (Diptera: Culicidae) in Botswana. Bioinvasions Rec. 2019;8:551–7.
Coetzee M, Kyalo D, Irish S, Snow R. Updated list of Anopheles species (Diptera: Culicidae) by country in the Afrotropical Region and associated islands. Zootaxa. 2020;4743:401–49.
Tolle MA. Mosquito-borne diseases. Curr Prob Pediatr Ad. 2009;39:97–140.
Braack L, de Almeida APG, Cornel AJ, Swanepoel R, De Jager C. Mosquito-borne arboviruses of African origin: review of key viruses and vectors. Parasite Vector. 2018;11:29.
Papa A. Emerging arboviruses of medical importance in the Mediterranean region. J Clin Virol. 2019;115:5–10.
Habtewold T, Povelones M, Blagborough AM, Christophides GK. Transmission blocking immunity in the malaria non–vector mosquito Anopheles quadriannulatus species A. PLoS Pathog. 2008;4:e1000070.
Takken W, Eling W, Hooghof J, Dekker T, Hunt R, Coetzee M. Susceptibility of Anopheles quadriannulatus theobald (Diptera: Culicidae) to Plasmodium falciparum. Trans R Soc Trop Med Hyg. 1999;93:578–80.
Lobo NF, Laurent BS, Sikaala CH, Hamainza B, Chanda J, Chinula D, Krishnankutty SM, Mueller JD, Deason NA, Hoang QT, Boldt HL. Unexpected diversity of Anopheles species in Eastern Zambia: implications for evaluating vector behavior and interventions using molecular tools. Sci Rep. 2015;5:17952.
Ravelo AC, Andreasen DH, Lyle M, Lyle AO, Wara MW. Regional climate shifts caused by gradual global cooling in the Pliocene epoch. Nature. 2004;429:263–7.
Seidel DJ, Fu Q, Randel WJ, Reichler TJ. Widening of the tropical belt in a changing climate. Nat Geosci. 2008;1:21.
Wu L, Cai W, Zhang L, Nakamura H, Timmermann A, Joyce T, Chang P. Enhanced warming over the global subtropical western boundary currents. Nat Clim Change. 2012;2:161–6.
Engelbrecht F, Adegoke J, Bopape MJ, Naidoo M, Garland R, Thatcher M, Gatebe C. Projections of rapidly rising surface temperatures over Africa under low mitigation. Environ Res Lett. 2015;10:085004.
Sørensen JG, Loeschcke V. Larval crowding in Drosophila melanogaster induces Hsp70 expression, and leads to increased adult longevity and adult thermal stress resistance. J Insect Physiol. 2001;47:1301–7.
Nyamukondiwa C, Weldon CW, Chown SL, le Roux PC, Terblanche JS. Thermal biology, population fluctuations and implications of temperature extremes for the management of two globally significant insect pests. J Insect Physiol. 2013;59:1199–211.
Stillman JH. A comparative analysis of plasticity of thermal limits in porcelain crabs across latitudinal and intertidal zone clines. Int Congr Ser. 2004;1275:267–74.
Nyamukondiwa C, Terblanche JS. Thermal tolerance in adult Mediterranean and Natal fruit flies (Ceratitis capitata and Ceratitis rosa): effects of age, gender and feeding status. J Therm Biol. 2009;34:406–14.
Chirebvu E, Chimbari MJ. Characteristics of Anopheles arabiensis larval habitats in Tubu village, Botswana. J Vector Ecol. 2015;40:129–38.
Machekano H, Mutamiswa R, Nyamukondiwa C. Evidence of rapid spread and establishment of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) in semi-arid Botswana. Agric Food Secur. 2018;7:48.
Benelli G, Jeffries CL, Walker T. Biological control of mosquito vectors: past, present, and future. Insects. 2016;7:52.
Sapari NB, Manan TSBA, Yavari S. Sustainable control of mosquito by larval predating Micronecta polhemus Niser for the prevention of mosquito breeding in water retaining structures. Int J Mosq Res. 2019;6:31–7.
Hunsicker ME, Ciannelli L, Bailey KM, Zador S, Stige LC. Climate and demography dictate the strength of predator-prey overlap in a subarctic marine ecosystem. PLoS ONE. 2013;8:e66025.
Xu C, Kohler TA, Lenton TM, Svenning JC, Scheffer M. Future of the human climate niche. Proc Natl Acad Sci. 2020. https://doi.org/10.1073/pnas.1910114117.
Bayoh MN, Lindsay SW. Temperature-related duration of aquatic stages of the Afrotropical malaria vector mosquito Anopheles gambiae in the laboratory. Med Vet Entomol. 2004;18:174–9.
Ruybal JE, Kramer LD, Kilpatrick AM. Geographic variation in the response of Culex pipiens life history traits to temperature. Parasite Vector. 2016;9:1–9.
Ngugi HN, Mutuku FM, Ndenga BA, Musunzaji PS, Mbakaya JO, Aswani P, LaBeaud AD. Characterization and productivity profiles of Aedes aegypti (L.) breeding habitats across rural and urban landscapes in western and coastal Kenya. Parasite Vector. 2017;10:331.
Abilio AP, Abudasse G, Kampango A, Candrinho B, Sitoi S, Luciano J, David MR. Distribution and breeding sites of Aedes aegypti and Aedes albopictus in 32 urban/peri–urban districts of Mozambique: implication for assessing the risk of arbovirus outbreaks. Plos Neglect Trop D. 2018;12:e0006692.
Sgro CM, Terblanche JS, Hoffmann AA. What can plasticity contribute to insect responses to climate change? Annu Rev Entomol. 2016;61:433–51.
Powell JR. Mosquito-borne human viral diseases: why Aedes aegypti? Am J Trop Med Hyg. 2018;98:1563–5.
Weitzel T, Jawień P, Rydzanicz K, Lonc E, Becker N. Culex pipiens sl and Culex torrentium (Culicidae) in Wrocław area (Poland): occurrence and breeding site preferences of mosquito vectors. Parasitol Res. 2015;114:289–95.
Pates HV, Takken W, Curtis CF, Jamet H. Zoophilic Anopheles quadriannulatus species B found in a human habitation in Ethiopia. Ann Trop Med Parasit. 2006;100:177–9.
Seyoum A, Sikaala CH, Chanda J, Chinula D, Ntamatungiro AJ, Hawela M, Miller JM, Russell TL, Briët OJ, Killeen GF. Human exposure to anopheline mosquitoes occurs primarily indoors, even for users of insecticide–treated nets in Luangwa Valley, South-east Zambia. Parasite Vector. 2012;5:101.
Dekker T, Takken W. Differential responses of mosquito sibling species Anopheles arabiensis and An. quadriannulatus to carbon dioxide, a man or a calf. Med Vet Entomol. 1998;12:136–40.
Chinula D, Hamainza B, Chizema E, Kavishe DR, Sikaala CH, Killeen GF. Proportional decline of Anopheles quadriannulatus and increased contribution of An. arabiensis to the An. gambiae complex following introduction of indoor residual spraying with pirimiphos-methyl: an observational, retrospective secondary analysis of pre-existing data from south-east Zambia. Parasite Vector. 2018;11:1–7.
Sharma V, Sharma M, Dhull D, Sharma Y, Kaushik S, Kaushik S. Zika virus: an emerging challenge to public health worldwide. Can J Microbiol. 2020;66:87–98.
Grigaltchik VS, Webb C, Seebacher F. Temperature modulates the effects of predation and competition on mosquito larvae. Ecol Entomol. 2016;41:668–75.
Lee HW, Ban S, Ikeda T, Matsuishi T. Effect of temperature on development, growth and reproduction in the marine copepod Pseudocalanus newmani at satiating food condition. J Plankton Res. 2003;25:261–71.
Cuthbert RN, Dalu T, Wasserman RJ, Weyl OL, Froneman PW, Callaghan A, Dick JT. Additive multiple predator effects of two specialist paradiaptomid copepods towards larval mosquitoes. Limnologica. 2019;79:125727.
Collins M, Knutti R, Arblaster J, Dufresne JL, Fichefet T, Friedlingstein P, Shongwe M. Long-term climate change: projections, commitments and irreversibility. In: Climate change 2013—the physical science basis: contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge: Cambridge University Press; 2013. p. 1029–136.
Shadrin NV, Anufriieva EV, Amat F, Eremin OY. Dormant stages of crustaceans as a mechanism of propagation in the extreme and unpredictable environment in the Crimean hypersaline lakes. Chin J Oceanol Limn. 2015;33:1362–7.
Svensson BG, Tallmark B, Petersson E. Habitat heterogeneity, coexistence and habitat utilization in five backswimmer species (Notonecta spp.; Hemiptera, Notonectidae). Aquat Insect. 2000;22:81–98.
Pincebourde S, Woods HA. There is plenty of room at the bottom: microclimates drive insect vulnerability to climate change. Curr Opin Insect Sci. 2020. https://doi.org/10.1016/j.cois.2020.07.001.
Hoffmann AA, Chown SL, Clusella-Trullas S. Upper thermal limits in terrestrial ectotherms: how constrained are they? Funct Ecol. 2013;27:934–49.
Terblanche JS, Deere JA, Clusella-Trullas S, Janion C, Chown SL. Critical thermal limits depend on methodological context. Proc R Soc B. 2007;274:2935–43.
Stillman JH. Acclimation capacity underlies susceptibility to climate change. Science. 2003;301:65.
Mondal RP, Chandra G, Bandyopadhyay S, Ghosh A. Effect of temperature and search area on the functional response of Anisops sardea (Hemiptera: Notonectidae) against Anopheles stephensi in laboratory bioassay. Acta Trop. 2017;166:262–7.
DeLong JP, Lyon S. Temperature alters the shape of predator-prey cycles through effects on underlying mechanisms. PeerJ. 2020;8:e9377.
Lyons CL, Coetzee M, Terblanche JS, Chown SL. Thermal limits of wild and laboratory strains of two African malaria vector species, Anopheles arabiensis and Anopheles funestus. Malar J. 2012;11:226.
Lyons CL, Oliver SV, Hunt RH, Coetzee M. the influence of insecticide resistance, age, sex, and blood feeding frequency on thermal tolerance of wild and laboratory phenotypes of Anopheles funestus (Diptera: Culicidae). J Med Entomol. 2016;53:394–400.
Hamblin AL, Youngsteadt E, López-Uribe MM, Frank SD. Physiological thermal limits predict differential responses of bees to urban heat–island effects. Biol Lett. 2017;13:20170125.
Cuthbert RN, Dick JT, Callaghan A, Dickey JW. Biological control agent selection under environmental change using functional responses, abundances and fecundities; the Relative Control Potential (RCP) metric. Biol Control. 2018;121:50–7.
Ma CS, Ma G, Pincebourde S. Survive a warming climate: insect responses to extreme high temperatures. Annu Rev Entomol. 2020;66:8.1-8.22.
Englund G, Öhlund G, Hein CL, Diehl S. Temperature dependence of the functional response. Ecol Lett. 2011;14:914–21.
Whitman DW. Acclimation. In: Whitman DW, Ananthakrishnan TN, editors. Phenotypic plasticity of insects. Mechanisms and consequences. Enfield: Science Publishers; 2009. p. 675–739.
Culler LE, Ayres MP, Virginia RA. In a warmer Arctic, mosquitoes avoid increased mortality from predators by growing faster. Proc R Soc B. 2015;282:20151549.
Calosi P, Bilton DT, Spicer JI. Thermal tolerance, acclimatory capacity and vulnerability to global climate change. Biol Lett. 2008;4:99–102.
Gunderson AR, Stillman JH. Plasticity in thermal tolerance has limited potential to buffer ectotherms from global warming. Proc R Soc B-Biol Sci. 2015;282:20150401.
van Heerwaarden B, Kellermann V, Sgrò CM. Limited scope for plasticity to increase upper thermal limits. Funct Ecol. 2016;30:1947–56.
Gunderson AR, Dillon ME, Stillman JH. Estimating the benefits of plasticity in ectotherm heat tolerance under natural thermal variability. Funct Ecol. 2017;31:1529–39.
Walther GR, Roques A, Hulme PE, Sykes MT, Pyšek P, Kühn I, Czucz B. Alien species in a warmer world: risks and opportunities. Trends Ecol Evol. 2009;24:686–93.
Carrington LB, Armijos MV, Lambrechts L, Barker CM, Scott TW. Effects of fluctuating daily temperatures at critical thermal extremes on Aedes aegypti life-history traits. PLoS ONE. 2013;8:e58824.
Acknowledgements
Gratitude is extended to the Department of Physics and Astronomy (BIUST) for the use of equipment and the South African Environmental Observation Network (NRF-SAEON) for use of Onset HOBO loggers. The Ministry of Environment, Natural Resources Conservation and Tourism (Botswana) is thanked for issuing a research permit (ENT 8/36/4XXXXII[14]). In addition, we acknowledge the contributions made by Murphy Tladi, Monamodi Kesamang, Bame Segaiso and Precious Mpofu in data collection and processing.
Funding
MB, CN and RJW acknowledge funding from the Botswana International University of Science and Technology (BIUST) for providing infrastructure and funding [REF: DVC/2/1/13 XI and DVC/RDI/2/1/7 V (18)] for the study. RC and TD acknowledge funding from the Alexander von Humboldt Foundation and University of Venda (SES/18/ERM/10) + National Research Foundation (117700), respectively.
Author information
Authors and Affiliations
Contributions
Project conceptualization and management: MB CN TD RC RJW. Data curation: MB. Formal analysis: MB RC. Investigation: MB CN RJW. Visualization and validation: MB CN TD RC RJW. Writing: original draft: MB CN RJW. Writing, review and editing: MB CN TD RC RJW.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
All authors declare no conflict of interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Buxton, M., Nyamukondiwa, C., Dalu, T. et al. Implications of increasing temperature stress for predatory biocontrol of vector mosquitoes. Parasites Vectors 13, 604 (2020). https://doi.org/10.1186/s13071-020-04479-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-020-04479-3