
Abstract
Background
Wildlife repatriation represents an opportunity for parasites. Reintroduced hosts are expected to accumulate generalist parasites via spillover from reservoir hosts, whereas colonization with specialist parasites is unlikely. We address the question of how myxozoan parasites, which are characterized by a complex life-cycle alternating between annelids and fish, can invade a reintroduced fish species and determine the impact of a de novo invasion on parasite diversity. We investigated the case of the anadromous allis shad, Alosa alosa (L.), which was reintroduced into the Rhine approximately 70 years after its extinction in this river system.
Methods
We studied parasites belonging to the Myxozoa (Cnidaria) in 196 allis shad from (i) established populations in the French rivers Garonne and Dordogne and (ii) repatriated populations in the Rhine, by screening the first adults returning to spawn in 2014. Following microscopical detection of myxozoan infections general myxozoan primers were used for SSU rDNA amplification and sequencing. Phylogenetic analyses were performed and cloned sequences were analyzed from individuals of different water sources to better understand the diversity and population structure of myxozoan isolates in long-term coexisting vs recently established host-parasite systems.
Results
We describe Hoferellus alosae n. sp. from the renal tubules of allis shad by use of morphological and molecular methods. A species-specific PCR assay determined that the prevalence of H. alosae n. sp. is 100 % in sexually mature fish in the Garonne/Dordogne river systems and 22 % in the first mature shad returning to spawn in the Rhine. The diversity of SSU rDNA clones of the parasite was up to four times higher in the Rhine and lacked a site-specific signature of SNPs such as in the French rivers. A second myxozoan, Ortholinea sp., was detected exclusively in allis shad from the Rhine.
Conclusions
Our data demonstrate that the de novo establishment of myxozoan infections in rivers is slow but of great genetic diversity, which can only be explained by the introduction of spores from genetically diverse sources, predominantly via straying fish or by migratory piscivorous birds. Long-term studies will show if and how the high diversity of a de novo introduction of host-specific myxozoans succeeds into the establishment of a local successful strain in vertebrate and invertebrate hosts.
Similar content being viewed by others

Background
The allis shad, Alosa alosa (L.) is an anadromous clupeid fish whose original distribution covered the area from the coast of southern Scandinavia to that of northwestern Africa. This species has a pelagic marine existence but upon maturation (4 to 6 years) migrates to spawn in the higher middle watercourse of rivers [1]. The populations of allis shad decreased severely by the middle of the 20th Century and are regarded as endangered on a European level. A combination of anthropogenic factors, such as the construction of dams on rivers that prevent spawning migrations, the destruction of spawning grounds, over-fishing and increasing pollution were considered to be causal [2, 3]. Residential populations of allis shad were likewise extinct in the Rhine ecosystem by the middle of the 20th Century [4, 5] until their repatriation in the course of the EU-LIFE project “The reintroduction of Allis shad (Alosa alosa) in the Rhine system” (2007–2010) and the follow-on EU-LIFE+ project “Conservation and Restoration of the Allis shad in the Gironde and Rhine watersheds” (2011–2015). Within these projects, allis shad brood stock from a natural population in France were spawned in captivity and 10.66 million larvae reared in aquaculture facilities were released into the Rhine system, between 2008 and 2014 [6, 7]. Adult shad first returned to spawn in the Rhine six years after the release of larvae in these waters [7].
Wildlife repatriation after long periods of absence offers a great opportunity for obtaining real time data on the recolonization of hosts with parasites and for understanding local species diversity. Whilst generalist parasites can be obtained from reservoir hosts in a relatively short period of time, colonization with specialist parasites is dependent on the contact with members of established fish populations elsewhere [8]. Allis shad shows natal site fidelity which provides fitness benefits due to local adaption [9, 10]. However, some degree of straying has been observed, especially between neighboring populations [11].
Myxozoans are morphologically extremely reduced cnidarian parasites with complex life-cycles that require an invertebrate (predominantly annelid) definitive host and a vertebrate (mainly fish) intermediate host [12, 13]. In the present study, allis shad originating from natural populations in the Dordogne/Garonne river systems were found infected with a myxozoan species belonging to the genus Hoferellus Berg, 1898 in the renal tubules and collecting ducts of the kidney. In cyprinids, Hoferellus spp. show extreme host specificity [14]. Specialist parasites with an indirect life-cycle can only establish if (i) all hosts required for completion of the life-cycle are present and (ii) one of the hosts becomes infected in a given habitat. In this study, we describe Hoferellus alosae n. sp. from Alosa alosa and investigate parasite SSU rDNA clone diversity in established host-parasite populations in the Dordogne/Garonne watershed and in the first infected hosts returning to the Rhine system for spawning. We address the questions of genetic diversity of newly establishing myxozoan specialist parasite populations and the potential sources of infection.
Methods
Origin of fish and diagnostic methods
During 2012–2014, allis shad were obtained from well-established natural populations of two large rivers in south-western France, the Dordogne and the Garonne watersheds, which merge into a macrotidal estuary, the Gironde that empties into the Atlantic (Fig. 1). In the Rhine (Germany), reintroduced allis shad first returned in 2014, when adults and newly-established young-of-the-year populations were detected and sampled. In the present study, 196 wild allis shad were analyzed for myxozoan infections. The wild fish populations were represented by 110 adults and 23 young-of-the-year caught in the Garonne/Dordogne system in France, as well as 9 adults and 54 young-of-the-year from the Rhine in Germany (Fig. 1, Additional file 1: Table S1). Young-of-the-year were directly frozen or fixed in 100 % ethanol after capture. Adult allis shad caught in the Rhine were frozen immediately or necropsied within 24 h after capture. The captive brood stock, which originated from the Garonne and the Dordogne, was reared in two 10 m3 tanks for about 1 month at a breeding facility in Bruch, France, as part of the reintroduction program. Allis shad that died during this time period or fish euthanized at the end of the reproduction period were analyzed during the present study (Additional file 1: Table S1). Freshly dead or moribund fish were frozen or analyzed immediately on site. The diagnostic methods were dependent on the quality and quantity of available material. All freshly dead and sacrificed fish were sectioned using sterile instruments and a complete bacteriological, mycological, virological and parasitological screening including histology was performed within the framework of the project. For the study of Hoferellus spp. infections, squash preparations of the kidney were examined and screened using light microscopy (Zeiss, Axiostar plus, Oberkochen, Germany). Furthermore, 125 samples of anterior kidney were fixed in 5 % formalin as well as 54 whole allis shad in Bouin’s solution (Sigma-Aldrich, Taufkirchen, Germany). The Bouin-fixed samples were decolorized over 24 h [15] and afterwards dehydrated in an alcohol series together with the formalin-fixed kidney samples, transferred to xylene and embedded in paraffin at the Hessen State Laboratory, Germany. Paraffin blocks were sectioned (4 μm), stained with haematoxylin and eosin and analyzed using a Leica microscope (Leica DM 2500, Leica Mikrosysteme Vertrieb GmbH, Wetzlar, Germany).

Map of France and Germany showing the watersheds studied (Dordogne, Garonne and Rhine), stocking locations (A-G, see Hundt et al. [6], sampling places (red dots), dams (black bars)
Parasite morphology
Kidneys from 14 freshly dead or euthanized adult allis shad of the donor population (France) were transferred into a 1.5 ml collection tube with 500 μl distilled water containing 100 U/ml Penicillin-Streptomycin solution. The material was sent to the Biology Centre of the Czech Academy of Sciences and analyzed immediately. Plasmodia and spores were examined on an Olympus BX51 microscope equipped with an Olympus DP72 digital camera. Measurements of spores (n = 25) follow the guidelines by Lom & Arthur (1989) [16] and were taken on digital images, using ImageJ v.1_44p (Wayne Rasband, http://imagej.nih.gov/ij).
SSU rDNA sequence analyses and genetic variability between sites
Kidney samples of 100 allis shad (Additional file 1: Table S1) frozen at -20 °C or fixed in 100 % ethanol were used for molecular analyses. Kidneys were removed from ethanol and briefly dried on a paper towel. Thereafter they were placed in TNES urea buffer [17] and DNA was extracted using proteinase K and a simplified phenol-chloroform extraction method [18]. Two partial, overlapping myxozoan SSU rDNA sequences were amplified using nested PCR assays. Universal eukaryotic primers ERIB1 and ERIB10 [19] were used in the first round. The resulting PCR product was used in two nested PCRs, employing myxozoan-specific primers: (i) MyxGP2F [20] and ACT1R [21]; and (ii) Myxgen4F [22] and ERIB10 (see above). The following PCR cycling conditions were used: 95 °C for 3 min, thereafter 30 cycles of 94 °C for 1 min, 60 °C [first round PCR)/58 °C (nested PCRs) for 1 min, 68 °C for 2 min (first round PCR)/1 min (nested PCRs)], followed by a final elongation step at 68 °C for 10 min. PCRs were performed in 10 μl reactions using Titanium Taq DNA polymerase. PCR products were visualized on 1 % agarose gels stained with ethidium bromide, purified using a Gel/PCR DNA Fragments Extraction Kit (Geneaid Biotech Ltd., New Taipei City, Taiwan) and sequenced commercially (https://www.seqme.eu/en/). Almost complete SSU rDNA sequences of H. alosae were obtained from four fish each from the Garonne and the Dordogne whereas the sequences obtained from infected fish from the Rhine showed double peaks in the variable sections of the SSU rDNA, indicating mixed infections. Nested PCR products of these samples were subsequently cloned into the pDrive Vector (PCR Cloning Kit, Qiagen, Hilden, Germany) and plasmid DNA was isolated using the High Pure Plasmid Isolation Kit (Roche Applied Science, Penzberg, Germany). Twelve clones per nested PCR product (2 overlapping partial SSU rDNA amplicons, see above) per infected fish (2 individuals) from the Rhine were sequenced and analyzed (fish individuals indicated in Additional file 1: Table S1).
In order to estimate SSU rDNA genetic diversity and compare it between Hoferellus isolates from different fish and rivers, nested PCR products (primers MyxGP2F and ACT1R) were cloned as described above. In total, six clones per fish were sequenced from three allis shad from the Dordogne, three shad from the Garonne and two infected specimens from the Rhine. Clones were obtained from adult shad as infection was only detected in adults in the Rhine. However, to determine differences in parasite diversity between the riverine young-of-the-year and adults returning from the sea to the same river, the same number of clones (3 fish, 6 clones each) was obtained from juveniles in the Garonne. Sequences were aligned in Geneious v8.1.3. (http://www.geneious.com, [23]) using the MAFFT v7.017 algorithm [24] and the L-INS-i method, with a default gap opening penalty (–op = 1.53) and gap extension penalty (–ep = 0.0). The number and position of single nucleotide changes and of polymorphic sites was noted and the divergence calculated.
Phylogenetic analyses
The SSU rDNA sequences obtained for the Hoferellus isolates from the three rivers as well as that of an Ortholinea sp. SSU rDNA sequence from allis shad kidneys from the Rhine were aligned with 21 ingroup taxa, which were selected on the basis of their close relatedness (BLAST result on GenBank) and to represent all sub-clades within the “freshwater myxosporean clade” sensu Fiala [25]. Basal freshwater myxosporeans Myxidium lieberkuehni Biitschli, 1882 and Chloromyxum legeri Tourraine, 1931 were used as outgroup taxa. Sequences were aligned as stated above and maximum parsimony (MP) analysis was performed in PAUP* v4.b10 [26], using a heuristic search with random taxa addition, the ACCTRAN option, TBR swapping algorithm, all characters treated as unordered and gaps treated as missing data. Maximum likelihood (ML) analysis was performed in RAxML v7.2.8 [27] using the GTR + Γ model. Clade support values were calculated from 1000 bootstrap replicates with random sequence additions for both MP and ML analyses. Bayesian inference analysis (BI) was performed in MrBayes v3.2.2 [28] implemented in Geneious and using the GTR + Γ + I model. MrBayes was run to estimate posterior probabilities over 1,100,000 generations via 2 independent runs of 4 simultaneous Markov Chain Monte Carlo (MCMC) algorithms with every 200th tree saved. ‘Burn-in’ was set to 100,000 generations.
Diagnostic PCR assay
In order to estimate true prevalences of Hoferellus infections in allis shad we designed a diagnostic PCR assay on the basis of specific nucleotide differences in highly variable regions of the SSU rDNA gene region. Primers HaloF (5'-CTT TGC GGT TTA CCC CAG AGG-3') and HaloR (5'-AAT TTC GAC GCC CAT AGT TGC-3') were used in PCR (see cycling protocol above) using 56 °C as annealing temperature and 40 s for elongation. The resulting 865 bp PCR product was sequenced from 15 kidney isolates including all rivers (see Additional file 1: Table S1). Specificity of the PCR assay was tested on DNA isolates of phylogenetically related myxosporeans H. cyprini (Doflein, 1898), H. carassii Akhmerov, 1960, Hoferellus sp., H. anurae Mutschmann, 2004, H. gnathonemi Alama-Bermejo, Jirků, Kodádková, Pecková, Fiala & Holzer, 2016, Ortholinea orientalis (Shul’man & Shul’man-Albova, 1953) and Ortholinea sp. (present study). All samples obtained in this study were screened for potential H. alosae n. sp. infection.
Scanning electron microscopy
The spores used for scanning electron microscopy were gently spun and pipetted onto filter paper (Millipore Millex-HV size 0.45 μl). The filter paper was stuck on a stub using Tissue-Tek and rapidly frozen (< 10-3 K/s) in slushy nitrogen. After freezing, the sample was transferred to a high vacuum preparation chamber (ALTO 2500, Gatan). The surface of the sample was sublimated at -95 °C, for 1 min. After sublimation, the sample was coated with a mixture of platina and paladium at a temperature of -135 °C. The coated sample was analyzed on a Field Emission Scanning Electron Microscope (JSM-7401 F, JEOL). Images were obtained at an accelerating voltage of 1 kV using GB high mode.
Results
Phylum Cnidaria Hatschek, 1888
Class Myxosporea Bütschli, 1881
Order Bivalvulida Schulman, 1959
Suborder Variisporina Lom & Noble, 1984
Family Sphaerosporidae Davis, 1917
Genus Hoferellus Berg, 1898
Hoferellus alosae n. sp.
Type-host : Alosa alosa (L.) (Actinopterygii: Clupeiformes: Clupeidae), allis shad.
Type-locality : River Garonne, France (mainly 44°06'33"N, 0°51'14"E).
Other localities : River Dordogne, France (mainly 44°50'42"N, 0°37'59"E) and River Rhine, Germany (49°04'09"N, 8°25'55"E and 49°24'19"N, 8°29'40"E); for additional sites see Additional file 1: Table S1.
Type-material : Unstained spores, fixed for 1 h in neutral buffered formalin, washed and mounted in glycerol-gelatine; ethanol-fixed infected kidney tissue of A. alosa, DNA extracted from infected kidney tissue stored at -80 °C; 2 histological slides stained with haematoxylin and eosin are deposited in the Collection of the Laboratory of Fish Protistology, Institute of Parasitology, Biology Centre of the Czech Academy of Sciences, České Budějovice, Czech Republic (record number: IPCAS Prot 34; collection curator: Miloslav Jirků, miloslav.jirku@seznam.cz).
Location of sporogonic stages : Kidney tubules (predominantly in anterior kidney), ureters and urinary bladder, exceptionally in Bowman’s capsules.
Prevalence : Determined by PCR. Garonne (2012–2014): adults 100 % (33/33), young-of-the-year 63.6 % (14/22); Dordogne (2013): adults 100 % (10/10); Rhine (2014): adults 22.2 % (2/9), young-of-the-year 0 % (0/26).
Representative DNA sequences : Three SSU rDNA sequences are submitted to the GenBank database under accession numbers KU301050–KU301052.
ZooBank registration : To comply with the regulations set out in article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature (ICZN) [29], details of the new species have been submitted to ZooBank (www.zoobank.org). The Life Science Identifier (LSID) of the article is urn:lsid:zoobank.org:pub: 15E34957–2C16-43E8-B374-845148395D1B. The LSID for the new name Hoferellus alosae is urn:lsid:zoobank.org:act: 230BD64C-E2C6-41D6-8DFC-19DC4362F237.
Etymology : Species name ‘alosae’ refers to the species name of the host Alosa alosa.
Description
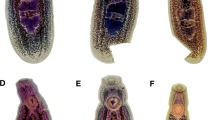
Mature spore. Mature spores subspherical in valvular view, ellipsoidal in sutural view, posteriorly rounded, measuring 9.1–10.3 (9.7 ± 0.4) μm in length, 7.7–9.2 (8.4 ± 0.5) μm in width, and 7.2–8.3 (7.7 ± 0.3) μm in thickness (n = 25 spores). Valves thickened at posterior end of spore, with 2 distinct but relatively small posterior valve projections (Fig. 2), occasionally with 3 to a maximum of 7 posterior filaments measuring 5–22 μm (Figs. 2 and 3b). Sutural line between valve cells straight, well marked; valves with 12 longitudinal ridges parallel to suture line, bifurcating in center of each valve, forming a distinct pattern (Fig. 3c, d). Polar capsules 2, equally sized, subspherical, 3.5–4.4 (4.0 ± 0.2) μm long, 2.4–3.6 (3.0 ± 0.3) μm wide (n = 25 spores). Polar filament with 5 turns. Sporoplasm in posterior part of spore, bi-nucleated.

Schematic line drawing of Hoferellus alosae n. sp. ex Alosa alosa

Morphological characteristics of Hoferellus alosae n. sp. spores and plasmodia. Light microscopy photomicrographs of a polysporous plasmodium with finger-like surface extensions (FE Plasm) and b spores with posterior filaments (FIL), one polar capsule with extruded polar filament (PF). c, d Scanning electron micrographs of spore surface showing suture between valves and longitudinal ridges and their characteristic pattern in the center of each valve. The arrow in c indicates capsular openings at the apical pole of the spore
Plasmodium. Plasmodia polymorphous (round, spherical or elongated), often with finger-like processes averaging 20 μm in length (Fig. 3a). Plasmodia di- to polysporous, without visible pansporoblast formation, measuring 25–71 × 18–53 μm.
Remarks
Five Hoferellus spp. have been found in nine clupeid species worldwide; of these four were reported from members of the genus Alosa but none of them from the allis shad (Table 1). Hoferellus donecii (Gasimagomedov, 1970) [30] and H. jurachni Moshu & Trombitsky, 2006 [31] differ considerably from H. alosae n. sp. with regard to spore shape and length as well as organization of the posterior spore appendages. Hoferellus caspialosum (Dogiel & Bychovsky 1939) [32] is smaller in size than H. alosae n. sp. Hoferellus caudatus (Parisi, 1910) [33] overlaps with H. alosae n. sp. with regard to most measurements. However, in contrast to H. alosae n. sp., this species was shown to consistently possess long posterior filaments, while H. alosae n. sp. was only occasionally found to have posterior filaments. Furthermore, the posterior end of the spore of H. caudatus is serrated whereas that of H. alosae n. sp. is smooth with only one small process on each valve. Hoferellus caudatus was described from an isolated, landlocked population of twaite shad Alosa agone (Scopoli, 1786) in Lake Como in Italy [33, 34], but was later reported from the anchovy Engraulis encrasicolus (L.) [35] and its Azov Sea-inhabiting subspecies E. encrasicolus maeoticus Pusanov, 1926 [36]. Considering the important difference in host habitat and the recently described strong host specificity of Hoferellus spp. in closely related cyprinids [12], it may be speculated that the records of Reshetnikova [35] and Naidenova [36] refer to a different parasite species in the anchovies. One could suspect a similar case for H. caspialosum which was described from the Caspian shad Alosa caspia caspia (Eichwald, 1838) by Dogiel & Bychovsky [32] but later reported from the Pontic shad Alosa immaculata Bennet, 1835 and the twait shad Alosa fallax (Lacépède, 1803) [37]. However, in contrast to A. alosa, the latter Alosa spp. are more closely related and their exact relationship is unresolved [38]. Unfortunately, SSU rDNA sequences are presently only available for H. alosae n. sp. from the allis shad.
Pathology and diagnostics
The gross examination of allis shad revealed no pathological or macroscopically visible changes of the kidney. In allis shad from the Garonne/Dordogne system the intensity of infection with Hoferellus alosae n. sp. was estimated as mild in 45 %, moderate in 47 % and severe in 9 % of cases, while the infection intensity of allis shad from the Rhine was always mild. Depending on the intensity of infection, the tubules were mildly to severely dilated, but generally no further pathological changes of the kidney tubules, the parenchyma or the excretory ducts were observed. In a single severe case the rupture of dilated renal tubules led to an inflow of H. alosae n. sp. in the surrounding kidney parenchyma causing an inflammation with infiltration of epitheloid cells.
The PCR assay designed in this study was specific for H. alosae n. sp. and did not amplify DNA of any phylogenetically related species screened in this study (see Methods).
SSU rDNA sequence diversity in host individuals and river systems
In allis shad from the Dordogne/Garonne river system, all sequences obtained by direct sequencing of PCR products and by sequencing of clones belonged to a single species, H. alosae n. sp. In contrast, in shad from the Rhine, H. alosae n. sp. was amplified from two individuals that had mixed infections of H. alosae n. sp. with a second myxozoan inhabiting the urinary tract of allis shad. This second myxozoan was later morphologically identified as Ortholinea sp., via spores detected in histological sections and the partial SSU rDNA sequence was submitted to GenBank under the accession number KU301053. Ortholinea sp. was also detected in a third fish from the Rhine (identified by SSU rDNA sequences from the nested myxozoan PCR assay), which had no mixed infection with H. alosae n. sp.
Almost full length SSU rDNA PCR products of H. alosae n. sp. in numerous fish from the Garonne did not exhibit variable positions or polymorphic sites, whereas those in allis shad from the Dordogne showed six consistent single nucleotide polymorphisms (SNPs) at positions 100 (A/G), 524 (C/T), 527 (C/T), 590 (A/G), 608 (C/T), 825 (C/T), in a 1923 bp alignment. The comparison of cloned partial SSU rDNA sequences (901 bp) for H. alosae n. sp. encompassing these sites, revealed two additional polymorphic sites (total of 8) and showed that clones from individual fish varied in only 3–7 base changes (1 SNP) in the Garonne, in 9–10 base changes (6 SNPs) in the Dordogne and in 15–18 base changes (2 SNPs) in the Rhine (summarized in Table 2; detailed in Additional files 2, 3 and 4: Tables S2, Tables S3 and Tables S4, see alignment in Additional file 5: Figure S1). Only a single SNP site overlapped between different rivers (position 655, rivers Rhine and Garonne). The genetic diversity comparison between young-of-the-year shad and adult fish returning to spawn (Garonne) showed a similar signature, with 3–7 base changes vs 3–5 base changes and the presence of a single, identical SNP (Table 2, Additional file 5: Figure S1). Sequence divergence between complete SSU rDNA sequences (PCR products of 19 fish; 1922 bp) was 0–1.09 % and that between partial cloned SSU rDNA isolates (66 sequences; 901 bp) was 0–1.44 %.
Phylogenetic relationships
BLAST results and subsequent pairwise sequence alignments indicated O. orientalis as the closest relative of H. alosae n. sp., with only 87–88 % SSU rDNA sequence identity. The second myxozoan SSU rDNA sequence belongs to Ortholinea sp. and was isolated from three fish in the Rhine. The sequence was almost identical to O. orientalis, with only 1.8–2.0 % sequence divergence over 912 bp. A consistent number of 15 nucleotide changes in the variable regions of the SSU rDNA suggest Ortholinea sp. is a different, but very closely related species. Phylogenetic analyses (MP/ML/BI; Fig. 4) showed that Ortholinea sp. clusters with O. orientalis and the three H. alosae n. sp. river isolates in a well-supported group (Fig. 4). This group furthermore clustered in polytomy with two clades composed of three Hoferellus (sensu lato) spp. (i.e. H. gilsoni (Debaisieux, 1925), H. anurae and H. gnathonemi) and of Ortholinea spp. + Myxobilatus gasterostei (Parisi, 1912) + Acauda hoffmani Whipps, 2011. Importantly, H. alosae n. sp. clustered outside the Hoferellus (sensu stricto) clade (comprising the type-species H. cyprini as well as H. carassii and Hoferellus sp. ex Cyprinus carpio L.) and hence is to be considered Hoferellus (sensu lato).

Bayesian inference (BI) tree showing the phylogenetic position of Hoferellus alosae n. sp. and Ortholinea sp. within the freshwater urinary bladder clade as defined by Fiala [25]. The new sequences are shown in bold and red. Myxidium lieberkuehni and Chloromyxum legeri were used as outgroup (OG). The short diagonal double-line represents a branch shortened to 50 % of its original length. Dashes at nodes represent nodal supports MP/ML < 50 and BI < 0.60 or node not present in the maximum parsimony (MP) or Maximum likelihood (ML) tree. Asterisk labels a node with maximum nodal supports (MP/ML = 100; BI = 1)
Discussion
Little is known about the population structure and dynamics of myxozoans, and information on the de novo establishment of myxozoans in watersheds that have been extirpated of obligatory fish host populations is missing to date. The repatriation program of the allis shad, A. alosa, in the Rhine allowed a unique first insight into the repopulation by myxozoans, and the diversity of new parasite settlements compared with watersheds where hosts and parasites have coexisted for a long period of time.
Baglinière & Élie [39] listed 16 species of Alosa native to the northern hemisphere and distributed through the western and eastern Atlantic coasts, the Mediterranean, Black and Caspian Seas, as well as Lake Volvi (Greece). The genus shows a large variation in life-history strategies (mostly anadromous, but also amphidromous, entirely marine and strictly freshwater) and a capacity to colonize new habitats thus making the genus Alosa an interesting model to study speciation and adaptation of the host itself and its parasites. To date, four Hoferellus spp. have been described from the urinary tract of Alosa spp. (Table 1). However, due to the different life strategies and hence geographical isolation (see also remarks in the species description) and strong host specificity in coelozoic myxozoans (e.g. [40–42]) with limited but convincing evidence also from the genus Hoferellus [14], parasite diversity in the genus Alosa may be larger than presently estimated. Unfortunately, sequence data are only available for H. alosae n. sp. from A. alosa (present study). Phylogenetic analyses of SSU rDNA sequences for Hoferellus spp. from different Alosa spp. and geographical localities in relation to host phylogeny would shed light on the diversity of species and the co-evolutionary history of Alosa spp. and their Hoferellus spp. parasites.
In myxozoans, species boundaries are difficult to determine as fish are unlikely infected by only one spore from a single parasite clone produced in one invertebrate host individual. SSU rDNA sequences have been widely used to describe and diagnose myxozoan species. rDNA occurs in a number of copies in eukaryotic cells [43], with around estimated 1000 copies in myxozoan rDNA [44–46]. Despite the concerted evolution of the rRNA gene [47], some degree of variation exists between these copies. Hence, rDNA sequences obtained from a single fish host show the full spectrum of such intragenomic heterogeneity as well as of intraspecific heterogeneity between ‘strains’ or genotypes. When assuming intragenomic heterogeneity as a constant in isolates of a single species, any additional variation can be ascribed to host- or site-specific variation. Comprehensive data on intraspecific variation of rDNA sequences are available for only a few myxozoan species, Myxobolus cerebralis Hofer, 1903 [48, 49], Tetracapsuloides bryosalmonae Canning, Curry, Feist, Longshaw & Okamura, 1999 [47, 48], Kudoa thyrsites (Gilchrist, 1924) [50], Ceratonova shasta (Noble, 1950) [51, 52] and Parvicapsula minibicornis Kent, Whitaker & Dawe, 1977 [53]. In most cases, variations have been ascribed to geographical differences between isolates. However, in the case of C. shasta, four sympatric genotypes were described, that showed little geographical structure in the parasite population but profound population isolation effects created by utilizing different vertebrate hosts. To some extent, population structuring by fish host was also evident in coho and Chinook salmon in P. minibicornis. In contrast to these species, H. alosae n. sp. seems to be host-specific and geographical isolation appears to be the main factor for SSU rDNA site variability.
Reports of allis shad populations in the Garonne and the Dordogne date back to the end of the 18th Century [54] and stocks are well-established despite a present decline [55]. Hoferellus alosae n. sp. SSU rDNA sequence variation is larger in hosts from the Dordogne (9–10 nucleotide positions and 6 SNPs) than in the Garonne (3–7 nucleotide positions, 1 SNP), potentially indicating a higher diversity in hosts from the Dordogne. This is surprising since the Garonne is longer (575 vs 472 km) and has a much larger number of tributaries, suggesting a higher diversity of invertebrate host habitats and populations of H. alosae n. sp. [56]. However, the distribution of susceptible oligochaete species is correlated to a variety of conditions [51, 57–59] and remains poorly understood. Independently, the presence of six SNP positions out of a total of 9–10 nucleotide changes indicates the establishment of a locally different ‘strain’ or genotype of H. alosae n. sp. in the Dordogne watershed when compared with that of the Garonne. Ceratonova shasta only showed three SNP SSU rDNA sites while P. minibicornis exhibited 17 SNPs [51, 53]. In contrast to these two species, our study was limited to a comparatively small sample set from three river systems (Garonne/Dordogne and Rhine) that revealed eight SNPs. The population dynamics of C. shasta and P. minibicornis are considerably different as a number of genotypes exist in different salmonid hosts and a polychaete definitive host is used in both cases [52, 53]. Hoferellus alosae n. sp. is expected to be very host-specific and to use an oligochaete definitive host, since it clusters in the ‘freshwater’ clade of myxozoans as defined by Fiala [25] whose members parasitize oligochaetes [60]. We expected to find more overlap in SNPs between the geographically close and estuary-linked French rivers. In contrast, our present data indicate a clear separation of H. alosae n. sp. populations in these rivers, which are characterized by a unique nucleotide signature for each river. The overlapping nucleotide signature pattern in young-of-the-year and adults in the Garonne indicate that straying of allis shad is limited, resulting in the establishment of local successful parasite genotypes as a consequence of long-term co-evolution of H. alosae n. sp. and allis shad in a specific watershed or microhabitat.
The SSU rDNA sequence variability pattern of H. alosae n. sp. in the first returners (2014) of allis shad repatriated in the Rhine was defined by only one (out of two) common SNP. The large number of individual changes in different clones (14–16) may indicate the existence of further SNP sites which cannot be identified at present. Likely, they represent those of close-by rivers, such as e.g. the Scheldt or the Meuse, where allis shad may be straying to or from. The large number of changes is likely a result of geographical distance from the French rivers. However, the nucleotide signatures (known SNPs and single nucleotide changes) vary to the same extent between the isolates from the Rhine as between Rhine and Garonne or Dordogne, rivers in which SNP signatures are site-specific and relatively homogenous. This strongly indicates that the infection in the two Rhine shad originates from at least two different sources and that genetic diversity in repatriated allis shad in the Rhine is higher than that found in established fish populations in the French rivers. Since the majority of the changes represents single nucleotide changes it may even be speculated that they represent multi-site signatures. The recovered isolates show a complexity that requires further analysis. In order to determine the diversity of H. alosae n. sp. isolates and populations in different watersheds as well as riverine microhabitats in more depth, the analysis of polymorphic microsatellite loci would be desirable as it allows a better fine scaling of the population structure in complex organisms like myxozoans that are characterized by clonal, vegetative and sexual reproduction. However, our findings underline that the analysis of parasites and their genetic diversity is a valuable tool to investigate biological characteristics and ecosystem variations.
The twaite shad, A. fallax shares A. alosa’s history of population decrease in the middle of the 20th Century but small populations can now be found restricted to the lower Rhine [61]. Though unlikely identical with H. caspialosum (originally described from geographically isolated A. caspia caspia, see Table 1 and Remarks section), the twaite shad in the Rhine may still harbor Hoferellus infection. In phylogenetic studies, A. alosa and A. fallax form clearly distinct lineages [38], even in watersheds where they hybridize [62]. The question arises whether the twaite shad can serve as a reservoir host for H. alosae n. sp. in the Rhine. Arguments that reject this hypothesis and the consequential existence of H. alosae n. sp.-infected oligochaete populations in the Rhine are: (i) Low infection rates of A. alosa [adults 22.2 % (2/9), young-of-the-year 0 % (0/26)] in contrast to the French rivers where 100 % of adults and 63.6 % of juveniles harbor infection, confirming the high prevalences typical for myxozoans in riverine habitats with effective dissemination of infective spore stages in the water current (e.g. [63]), and (ii) The high genetic diversity and lack of an identical genetic signature of H. alosae n. sp. isolates from the Rhine. Alosa fallax were not examined in the course of this project, however, these circumstances strongly suggest that A. fallax does not serve as a reservoir host for H. alosae n. sp. More likely, infection in the returning adults results from their contact with different H. alosae n. sp. enzootic watersheds. Alternatively, oligochaetes in parts of the river may become infected by spores released from straying allis shad originating from elsewhere or by spores released via the faeces of migratory piscivorous birds. The importance of avian vectors in the translocation of spores is unexplored to date but M. cerebralis spores are able to infect oligochaetes after avian intestinal passage and are released for several days after fish consumption [64]. New invasion and colonization events are generally associated with founder effects that reduce genetic variation in incipient populations [65]. However, due to the abundance and diversity of myxozoans in different aquatic habitats and a variety of source localities for de novo invasion this appears to be the opposite in this group. High SSU rDNA diversity in H. alosae n. sp. from the Rhine and the presence of another myxozoan, Ortholinea sp., which was absent from the French watersheds but present in three out of nine adult shad in the Rhine, suggests that myxozoan parasite introduction into parasite-free territory is characterized by a high diversity of species and ‘strains’. To our knowledge, this is the first time that initial parasite introduction data are available for myxozoans. Monitoring of the infections in repatriated allis shad returning to spawn over following years will allow to obtain real-time data on within-host competition and the survival and predominant establishment of the fittest in the Rhine.
Conclusion
To the best of our knowledge, this is the first study on a de novo introduction of a myxozoan species in a river where the host fish population was repatriated after 70 years of absence and loss of infection from the alternate invertebrate host. The first adults of allis shad returning to spawn in the Rhine showed low infection prevalence of H. alosae n. sp. when compared to established fish populations in France, where all adults were found infected. However, the diversity of H. alosae n. sp. clones in only two infected hosts varied as strongly between the Rhine isolates as between the Rhine and two rivers with established fish populations and H. alosae n. sp. infections in France. Additionally, a second species, Ortholinea sp., was only found in the Rhine. This comparatively high diversity in the newly established host population can only be explained by the introduction of spores from genetically diverse sources, most likely via straying hosts. The allis shad repatriation study in the Rhine presently offers the unique opportunity for a long-term study of a de novo settlement of this host-specific myxozoan and its succession from high diversity as a result of multiple introductions to a locally established, presumably successful strain, which characterize each of the French rivers.
Abbreviations
- BI:
-
Bayesian inference
- EU-LIFE:
-
European Union -L’Instrument Financier pour l’Environnement
- MP:
-
Maximum parsimony
- ML:
-
Maximum likelihood
- SNP:
-
Single nucleotide polymorphism
- SSU rDNA:
-
Small subunit ribosomal deoxyribonucleic acid
References
Baglinière J-L, M. Richard Sabatié, Rochard E, Alexandrino P, Aprahamian MW. The allis shad Alosa alosa: Biology, ecology, range and status of populations. Am Fish S S. 2003;5:85–102.
Aprahamian MW, Aprahamian CD, Baglinière JL, Sabatié R, Alexandrino P. Alosa alosa and Alosa fallax spp.: Literature review and bibliography. Warrington: Environment Agency; 2003.
Freyhof J. Freshwater fish diversity in Germany, threats and species extinction. In: Collares-Pereira MJ, Coelho MM, Cowx IG, editors. Conservation of freshwater fishes: Options for the future. Oxford: Fishing News Books; 2002. p. 3–22.
Beeck P. Konzeptstudie für ein wissenschaftlich begründetes Programm zur Wiederansiedlung des Maifischs im Rhein. Recklinghausen: Unveröffentlichte Studie im Auftrag der Landesanstalt für Ökologie, Bodenordnung und Forsten; 2004.
Maitland P, Hatton-Elli T. Ecology of the Allis and Twaite shad, Rivers Ecology, vol. 3. Peterborough: Conserving Natura 2000 Rivers Ecology Series No. 3; 2003. English Nature.
Scharbert A, Beeck P, Rochard E, Pierre RS, Jatteau P. The re-introduction of the Allis shad (Alosa alosa) to the Rhine system (LIFE06 NAT/D/000005). Düsseldorf: Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen; 2011.
Hundt M, Scharbert A, Weibel U, Kuhn G, Metzner K, Jatteau P, et al. First evidence of natural reproduction of the Allis shad Alosa alosa in the River Rhine following re-introduction measures. J Fish Biol. 2015;87(2):487–93.
Poulin R. Evolutionary Ecology of Parasites. 2nd ed. New Jersey: Princeton University Press; 2006.
Hendry AP. Selection against migrants contributes to the rapid evolution of ecologically dependent reproductive isolation. Evol Ecol Res. 2004;6(8):1219–36.
Tomas J, Augagneur S, Rochard E. Discrimination of the natal origin of young-of-the-year Allis shad (Alosa alosa) in the Garonne-Dordogne basin (south-west France) using otolith chemistry. Ecol Freshw Fish. 2005;14(2):185–90.
Martin J, Rougemont Q, Drouineau H, Launey S, Jatteau P, Bareille G, et al. Dispersal capacities of anadromous Allis shad population inferred from a coupled genetic and otolith approach. Can J Fish Aquat Sci. 2015;72(7):991–1003.
Lom J, Dyková I. Myxozoan genera: definition and notes on taxonomy, life-cycle terminology and pathogenic species. Folia Parasitol. 2006;53:1–36.
Okamura B, Gruhl A, Bartholomew JL. Myxozoan Evolution, Ecology and Development. Cham: Springer International Publishing; 2015.
Alama-Bermejo G, Jirků M, Kodádková A, Pecková H, Fiala I, Holzer AS. Species complexes and phylogenetic lineages of Hoferellus (Myxozoa, Cnidaria) including revision of the genus: A problematic case for taxonomy. Parasit Vectors. 2016;9(1):1–21.
Burck H-C. Histologische Technik - Leitfaden für die Herstellung mikroskopischer Präparate in Unterricht und Praxis. 6th ed. Stuttgart: Georg Thieme Verlag; 1988.
Lom J, Arthur JR. A guideline for the preparation of species descriptions in Myxosporea. J Fis Dis. 1989;12(2):151–6.
Asahida T, Kobayashi T, Saitoh K, Nakayama I. Tissue preservation and total DNA extraction from fish stored at ambient temperature using buffers containing high concentration of urea. Fisheries Sci. 1996;62:727–30.
Holzer AS, Sommerville C, Wootten R. Molecular relationships and phylogeny in a community of myxosporeans and actinosporeans based on their 18S rDNA sequences. Int J Parasitol. 2004;34(10):1099–111.
Barta JR, Martin DS, Liberator PA, Dashkevicz M, Anderson JW, Feighner SD, et al. Phylogenetic relationships among eight Eimeria species infecting domestic fowl inferred using complete small subunit ribosomal DNA sequences. J Parasitol. 1997;83:262–71.
Kent ML, Khattra J, Hervio DML, Devlin RH. Ribosomal DNA sequence analyses of isolates of the PKX myxosporean and their relationship to members of the genus Sphaerospora. J Aquat Anim Health. 1998;10:12–21.
Hallett SL, Diamant A. Ultrastructure and smallsubunit ribosomal DNA sequence of Henneguya lesteri n. sp. (Myxosporea), a parasite of sand whiting Sillago analis (Sillaginidae) from the coast of Queensland, Australia. Dis Aquat Org. 2001;46:197–212.
Diamant A, Whipps CM, Kent ML. A new species of Sphaeromyxa (Myxosporea: Sphaeromyxina: Sphaeromyxidae) in devil firefish, Pterois miles (Scorpaenidae), from the northern Red Sea: morphology, ultrastructure, and phylogeny. J Parasitol. 2004;90(6):1434–42.
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28(12):1647–9.
Katoh M, Kuma M. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002;30:3059–66.
Fiala I. The phylogeny of Myxosporea (Myxozoa) based on small subunit ribosomal RNA gene analysis. Int J Parasitol. 2006;36(14):1521–34.
Swofford DL. PAUP* Phylogenetic Analysis Using Parsimony (*and Other Methods) 4.0. b5. Sunderland: Sinauer Associates; 2003.
Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006;22(21):2688–90.
Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19(12):1572–4.
International Commission on Zoological Nomenclature. Amendment of articles 8, 9, 10, 21 and 78 of the International Code of Zoological Nomenclature to expand and refine methods of publication. Zootaxa. 2012;3450:1–7.
Gasimagomedov AA. New species of myxosporea in the fishes of the Caspian Sea. Sbornik Nauchnykh Soobshchenii Kafedry Fiziologii Cheloveka i Zhivomykh Respubliki Dagestan. Makkachkala. 1970;4:72–6.
Moshu AJ, Trombitsky ID. New parasites (Apicomplexa, Cnidospora) of some Clupeidae fishes from the Danube and Dniestr Basins. Eco-TIRAS International Environmental Association of River Keepers Leo Berg Educational Foundation Academician Leo Berg -103 Collection of Scientific Articles. Acad Leo Berg., Collect Sci Artic. 2006;130:95–103
Dogiel VA, Bychovsk BE. Parasites of fishes of the Caspian Sea. Trudy Kompleksnoi Izucheniyu Kaspiiskogo Morya. 1939;7(1):150 (in Russian).
Parisi B. Sphaerospora caudata n. sp. Zool Anz. 1910;36:253–4.
Parisi B. Sulla Sphaerospora caudata Parisi. Atti Soc Ital Sc Nat Mus Civ Stor Nat Milano. 1913;51:1–11.
Reshetnikova AV. Parasites of Certain Commercial Black Sea Fish. St.Petersburg: Zoological Institute of the Academy of Sciences; 1954.
Naidenova NN, Shulman SS, Donetz ZS. Class cnidosporidians - Cnidosporidia Doflein, 1901, emend. Shulman, 1967. In: Greze VN, Delyamure SP, Nikolaeva VM, editors. Identification of the parasites of vertebrates in the Black and Azov Seas. Kiev: Akademiya Nauk Ukrainskoy SSR, “Naukova Dumka”; 1975. p. 20–50 (In Russian).
Zaika VE. Parasite protozoa of the black sea fishes. Biol Morya. 1968;14:41–6 (in Russian).
Faria R, Weiss S, Alexandrino P. A molecular phylogenetic perspective on the evolutionary history of Alosa spp. (Clupeidae). Mol Phylogenet Evol. 2006;40(1):298–304.
Baglinière J-L, Élie P. Les aloses: (Alosa alosa et Alosa fallax sp.): écobiologie et variablité des populationes. Versailles Cedex: Editions Quae; 2000.
Heiniger H, Adlard RD. Molecular identification of cryptic species of Ceratomyxa Thelohan, 1892 (Myxosporea: Bivalvulida) including the description of eight novel species from apogonid fishes (Perciformes: Apogonidae) from Australian waters. Acta Parasitol. 2013;58(3):342–60.
Alama-Bermejo G, Raga JA, Holzer AS. Host-parasite relationship of Ceratomyxa puntazzi n. sp. (Myxozoa: Myxosporea) and sharpsnout seabream Diplodus puntazzo (Walbaum, 1792) from the Mediterranean with first data on ceratomyxid host specificity in sparids. Vet Parasitol. 2011;182(2-4):181–92.
Bartosova P, Fiala I, Jirku M, Cinkova M, Caffara M, Fioravanti ML, et al. Sphaerospora sensu stricto: taxonomy, diversity and evolution of a unique lineage of myxosporeans (Myxozoa). Mol Phylogenet Evol. 2013;68(1):93–105.
Torres-Machorro AL, Hernandez R, Cevallos AM, Lopez-Villasenor I. Ribosomal RNA genes in eukaryotic microorganisms: witnesses of phylogeny? FEMS Microbiol Rev. 2010;34(1):59–86.
Alama-Bermejo G, Sima R, Raga JA, Holzer AS. Understanding myxozoan infection dynamics in the sea: seasonality and transmission of Ceratomyxa puntazzi. Int J Parasitol. 2013;43(9):771–80.
Harada T, Kawai T, Jinnai M, Ohnishi T, Sugita-Konishi Y, Kumeda Y. Detection of Kudoa septempunctata 18S ribosomal DNA in patient fecal samples from novel food-borne outbreaks caused by consumption of raw olive flounder (Paralichthys olivaceus). J Clin Microbiol. 2012;50(9):2964–8.
True K, Purcell MK, Foott JS. Development and validation of a quantitative PCR to detect Parvicapsula minibicornis and comparison to histologically ranked infection of juvenile Chinook salmon, Oncorhynchus tshawytscha (Walbaum), from the Klamath River, USA. J Fish Dis. 2009;32(2):183–92.
Hillis DM, Scott KD. Ribosomal DNA: intraspecific polymorphism, concerted evolution, and phylogeny reconstruction. Syst Zool. 1988;37(1):63–6.
Whipps C, El-Matbouli M, Hedrick R, Blazer V, Kent M. Myxobolus cerebralis internal transcribed spacer 1 (ITS-1) sequences support recent spread of the parasite to North America and within Europe. Dis Aquat Org. 2004;60:105–8.
Lodh N, Kerans BL, Stevens L. The parasite that causes whirling disease, Myxobolus cerebralis, is genetically variable within and across spatial scales. J Eukaryot Microbiol. 2012;59(1):80–87. eng.
Whipps CM, Kent ML. Phylogeography of the cosmopolitan marine parasite Kudoa thyrsites (Myxozoa: Myxosporea). J Eukaryot Microbiol. 2006;53(5):364–73.
Atkinson SD, Bartholomew JL. Disparate infection patterns of Ceratomyxa shasta (Myxozoa) in rainbow trout (Oncorhynchus mykiss) and Chinook salmon (Oncorhynchus tshawytscha) correlate with internal transcribed spacer-1 sequence variation in the parasite. Int J Parasitol. 2010;40(5):599–604.
Atkinson SD, Bartholomew JL. Spatial, temporal and host factors structure the Ceratomyxa shasta (Myxozoa) population in the Klamath River basin. Infect Genet Evol. 2010;10(7):1019–26.
Atkinson SD, Jones SR, Adlard RD, Bartholomew JL. Geographical and host distribution patterns of Parvicapsula minibicornis (Myxozoa) small subunit ribosomal RNA genetic types. Parasitology. 2011;138(8):969–77.
Keith P, Allardi J. Endangered freshwater fish: the situation in France. In: Kirchhofer A, Müller DR, editors. Conservation of endangered freshwater fish in Europe. Basel: Birkhäuser; 1996. p. 35–54.
Rougier T, Lambert P, Drouineau H, Girardin M, Castelnaud G, Carry L, et al. Collapse of Allis shad, Alosa alosa, in the Gironde system (southwest France): environmental change, fishing mortality, or Allee effect? ICES J Mar Sci. 2012;69(10):1802–11.
Maasri A, Dumont B, Claret C, Archambaud-Suard G, Gandouin E, Franquet E. Tributaries under Mediterranean climate: their role in macrobenthos diversity maintenance. C R Biol. 2008;331(7):547–58.
Marcogliese DJ, Gendron AD, Cone DK. Impact of municipal effluents and hydrological regime on myxozoan parasite communities of fish. Int J Parasitol. 2009;39(12):1345–51.
Auer NA, Kahn JE. Abundance and distribution of benthic invertebrates, with emphasis on Diporeia, along the Keweenaw Peninsula, Lake Superior. J Great Lakes Res. 2004;30:340–59.
Nijboer RC, Wetzel MJ, Verdonschot PF. Diversity and distribution of Tubificidae, Naididae, and Lumbriculidae (Annelida: Oligochaeta) in the Netherlands: an evaluation of twenty years of monitoring data. Hydrobiologia. 2004;520(1-3):127–41.
Holzer AS, Wootten R, Sommerville C. The secondary structure of the unusually long 18S ribosomal RNA of the myxozoan Sphaerospora truttae and structural evolutionary trends in the Myxozoa. Int J Parasitol. 2007;37(11):1281–95.
Wiegerinck JAM, Boois IJ, Keeken OA, Willigen J. Jaarrapportage Passieve Vismonitoring Zoete Rijkswateren: fuik- en zalmsteekregistraties in 2010. Lelystad: IMARES - Institute for Marine Resources & Ecosystem Studies; 2011.
Alexandrino P, Faria R, Linhares D, Castro F, Le Corre M, Sabatié R, et al. Interspecific differentiation and intraspecific substructure in two closely related clupeids with extensive hybridization, Alosa alosa and Alosa fallax. J Fish Biol. 2006;69(sb):242–59.
Holzer AS, Sommerville C, Wootten R. Molecular studies on the seasonal occurrence and development of five myxozoans in farmed Salmo trutta L. Parasitology. 2006;132(2):193–205.
Koel TM, Kerans BL, Barras SC, Hanson KC, Wood JS. Avian piscivores as vectors for Myxobolus cerebralis in the Greater Yellowstone ecosystem. Trans Amer Fish Soc. 2010;139(4):976–88.
Carlson H, Templeton H. Genetic revolutions in relation to speciation phenomena: the founding of new populations. Annu Rev Ecol Syst. 1984;15:97–132.
Acknowledgements
The authors would like to thank Aline Chaumel, Syndicat Mixte d’Etude et d’Aménagement de la Garonne, Toulouse, France, Rudi Hell, Thomas Oswald, Dieter Herold and Thomas Höfer for providing of samples. Moreover, we would like to thank Kristina Dersch and Sivia Hopfinger from the Hessen State Laboratory, Giessen for their excellent technical assistance. Furthermore the authors are thankful to David Clavé, Association Migrateurs Garonne Dordogne (MIGADO), Le Passage d’Agen for his excellent support. We are particularly grateful for the cooperation with the LIFE+ project “Alosa alosa” (LIFE09 NAT/DE/000008) which was the prerequisite for the implementation of this study.
Funding
We acknowledge funding for molecular and ultrastructural research by the Czech Science Foundation (505/12/G112, European Center of Ichthyoparasitology), the Ministry of Education, Youth and Sports (LM2015062, Czech-BioImaging) and by the Institute of Parasitology of the Biology Centre of the Czech Academy of Sciences (RVO: 60077344).
Availability of data and materials
Data and other information supporting the results are presented in the article and its Additional files. Three SSU rDNA sequences of Hoferellus alosae n. sp. obtained from fish of each river were submitted to the GenBank database under the accession numbers KU301050–KU301052. The partial SSU rDNA sequence of the second myxozoan identified as Ortholinea sp. has the accession number KU301053.
Authors’ contributions
HW and ASH conceived and designed the study. HW, ASH and ML coordinated its design and execution; HW and ML were responsible for the collection of all samples from fish. HW and UE performed the histological analysis, PBS and HP the molecular analysis. HW and ASH drafted the manuscript and all authors read and approved its final version.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
All animal procedures were performed in accordance with European [Regulation (EC) No 614/2007 of the European Parliament and of the Council of 23 May 2007 (LIFE+)] and German legislation [Animal Welfare Law, Federal Regulation for the Protection of Species (BArtSchV)]. We declare that our study complies with the Convention for the Protection of the Marine Environment of the North-East Atlantic (the “OSPAR Convention”), the Convention on the Trade in Endangered Species of Wild Fauna and Flora and the IUCN Policy Statement on Research Involving Species at Risk of Extinction as well as the IUCN Guidelines for re-introductions.
Author information
Authors and Affiliations
Corresponding author
Additional files
Additional file 1: Table S1.
Summary of samples obtained from Alosa alosa. (DOCX 48 kb)
Additional file 2: Table S2.
SSU rDNA variability of Hoferellus alosae n. sp. clones from fish individuals in the Garonne. (DOCX 18 kb)
Additional file 3: Table S3.
SSU rDNA variability of Hoferellus alosae n. sp. clones from fish individuals in the Dordogne. (DOCX 17 kb)
Additional file 4: Table S4.
SSU rDNA variability of Hoferellus alosae n. sp. clones from fish individuals in the Rhine. (DOCX 16 kb)
Additional file 5: Figure S1.
Alignment of cloned SSU rDNA fragments of Hoferellus alosae n. sp. (901 bp) obtained from different fish individuals and river systems (see Additional file 1: Table S1 for origin and characteristics of samples). Single nucleotide polymorphic sites (SNPs) and individual nucleotide changes marked in color. (PNG 557 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

{kind=link}
Cite this article
Wünnemann, H., Holzer, A.S., Pecková, H. et al. Repatriation of an old fish host as an opportunity for myxozoan parasite diversity: The example of the allis shad, Alosa alosa (Clupeidae), in the Rhine. Parasites Vectors 9, 505 (2016). https://doi.org/10.1186/s13071-016-1760-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-016-1760-6