
Abstract
The vagina hosts a community of microorganisms known as the vaginal microbiota. This community is relatively stable and straightforward, with Lactobacillus species being the most dominant members. The vaginal microbiota has various functions that are essential for maintaining human health and balance. For example, it can metabolise dietary nutrients, produce growth factors, communicate with other bacteria, modulate the immune system, and prevent the invasion of harmful pathogens. When the vaginal microbiota is disrupted, it can lead to diseases and infections. The observed disturbance is distinguished by a reduction in the prevalence of Lactobacillus and a concurrent rise in the number of other bacterial species that exhibit a higher tolerance to low oxygen levels. Gynecologic cancers are a group of cancers that affect the female reproductive organs and tissues, such as the ovaries, uterus, cervix, vagina, vulva, and endometrium. These cancers are a major global health problem for women. Understanding the complex interactions between the host and the vaginal microorganisms may provide new insights into the prevention and treatment of gynecologic cancers. This could improve the quality of life and health outcomes for women.
Similar content being viewed by others

Introduction—what is the scope of this review?
The word “microbiota” was first used in the early 1900s, when various microorganisms, such as bacteria, viruses, and yeasts, lived in different parts of the human body [1]. Despite their frequent interchangeability, microbiota and microbiome have subtle differences in meaning. For instance, “microbiota” refers to microorganisms living in a particular habitat. On the other hand, the term “microbiome” refers to a more comprehensive notion that comprises not only the actual microbes but also their genomes, or genetic material, and the environments in which they live [2]. The microbiota is critical in maintaining human health and balance through many processes. These include several processes, such as acquiring energy from nutritional sources, promoting growth hormone synthesis, intercellular communication among bacteria, augmenting immune system activity, inhibiting pathogen colonisation and resisting their displacement of the epithelium or mucosal peithelium [3, 4].
The vaginal microbiota refers to a complex assemblage of microorganisms that inhabit the vaginal environment. Compared to other parts of the body, the vaginal microbiota is comparatively less varied and is often dominated by the Lactobacillus species [5, 6]. The probiotic activity is attributed to individual Lactobacillus species and their multi-microbial interaction [7]. Through competition with other bacteria for resources and the release of chemicals that make the vaginal environment inhospitable to other potentially harmful germs, the vaginal microbiota plays a crucial role in the health condition of the female reproductive system [8, 9]. Vaginal microbial dysbiosis contributes to the pathogenic process of illnesses. Frequently, there is an observed transition from a state characterised by a predominance of Lactobacillus to one marked by an elevation in facultative and anaerobic microorganisms [10, 11]. Treatments aimed at altering vaginal microbiota have been very successful in curing a variety of illnesses, which supports the viability of using vaginal microbiota in the management of gynecologic disorders [12].
Gynecologic cancers are a significant global health issue for women. The phrase “gynecological cancers” encompasses a range of malignant diseases that primarily affect the reproductive tissues and organs in females, such as the ovaries, uterus, cervix, vagina, vulva, and endometrium. Frequently, there is a transition from a state characterised by a predominance of Lactobacillus to one marked by an escalation in facultative and anaerobic microorganisms [13]. Gynecologic malignancies, which comprise about 14.4% of newly diagnosed cancer cases in women globally, have a substantial impact on both mortality and morbidity. The elements mentioned significantly influence the quality of life (QoL) experienced by survivors [14]. Patients have a diverse array of symptoms, including psychological anguish, menopausal complications, exhaustion, sleep disturbances, bladder and bowel dysfunction, lymphedema, and sexual impairments [15].
Vaginal microbiota
The identification of vaginal microbiota dates back to 1892, when Albert Doderlein, a German obstetrician and gynecologist, observed the presence of elongated, robust, mobile, rod-shaped bacteria in vaginal fluid. These bacteria were characterised as gram-positive and non-spore-producing [16]. After discovering its ability to produce lactic acid, Doderlein’s bacillus was called Lactobacillus. Doderlein also observed that puerperal fever was connected to the lack of his bacillus in vaginal fluid and that it inhibited Staphylococcus growth [17, 18]. The findings of this study have shown that Lactobacillus is the predominant bacterial species in the vaginal microbiota and has a crucial role in maintaining women’s health [19].
The composition of the bacterial community in the vagina has been comprehensively determined to mainly consist of lactobacilli, which may be categorised into five unique groups referred to as community state types (CST). The dominant species in each of the classed vaginal microbiota states, known as CSTs I, II, III, IV, and V, are Lactobacillus crispatus, L. gasseri, L. iners, a polymicrobial flora consisting of Lactobacillus and bacteria associated with bacterial vaginosis (BVAB), and L. jensenii, respectively [19, 20]. Females usually exhibit CST I, III, and IV, which have been extensively investigated in scholarly studies. Conversely, CST II and V are less commonly seen in women [21]. Indeed, studies conducted by DiGiulo et al. and van de Wijgert et al. have shown that the vaginal microbiota of women in good health may be partly categorised into CST II and V [22, 23]. Gajer et al. (2012) further categorised CST IV, a notable deficiency in a specific Lactobacillus species, into two subgroups, CST IV-A and CST IV-B. According to the findings of Gajer et al., it has been shown that CST IV-B has a much higher abundance of BVAB. In contrast, CST IV-A generally consists of a modest proportion of L. iners and anaerobic bacteria such as Corynebacterium, Finegoldia, Streptococcus, or Anaerococcus [19, 24].
CST I is often linked to the preservation of vaginal health, which is crucial for preventing infections such as bacterial vaginosis, as well as other health concerns such as female infertility, early birth, and miscarriage. The main contributing factor to this phenomenon is its ability to produce lactic acid, hydrogen peroxide, and bacteriocins, creating an inhospitable environment for certain pathogenic microorganisms [25,26,27]. CST II, like CST I, generates lactic acid and plays a role in maintaining a favorable vaginal milieu. However, it serves a more dynamic and intermediate function in terms of protection [28]. The efficacy of CST III in protecting bacterial vaginosis and problems related to pregnancy is comparatively lower when compared to other species of Lactobacillus. There exists a correlation between the presence of L. iners and a vaginal environment that is susceptible to dysbiosis [29]. According to Witkin et al., many factors are believed to contribute to this phenomenon. One such factor is the ability of L. iners to produce a distinct isomeric form of lactic acid (L-lactic acid) that lacks the required power to hinder the invasion of pathogenic bacteria [30]. The absence of D-lactic acid seems to have a role in the deterioration of the extracellular matrix, hence facilitating the mobility of pathogenic microbes [31]. Moreover, it was also shown that L. iners generates inerolysin, a pore-forming protein often seen in disease-causing bacteria, which might increase its adhesive ability [32]. The ability of L. iners to generate inerolysin might be one of the most significant determinants impacting its capability to get nutrients from the vaginal environment [33]. To thrive in the bacterial vaginosis environment, L. iners can enhance the expression of inerolysin and mucin and stimulate the creation of glycerol and the expression of relevant metabolic enzymes. This allows the bacteria to obtain nutrients from external sources effectively [34, 35]. CST IV is correlated with vaginal dysbiosis, perhaps rendering women more susceptible to recurring infections. This association may be attributed to the likely creation of biofilm as well as an elevated risk of contracting sexually transmitted diseases and bacterial vaginosis [36]. The cytotoxic protein vaginolysin, dependent on cholesterol, is secreted in the highest amounts by CST IV, particularly by G. vaginalis [37]. Another group of bacteria associated with bacterial vaginosis, Fusobacteria and G. vaginalis, secrete the enzyme sialidase. This enzyme causes mucus breakdown, making the cervical epithelium more susceptible to viral infections [38, 39]. Finally, CST V is seen as a vaginal community state type that promotes a favorable and stable vaginal environment, akin to CST I [40].
Research has shown that the incidence of CST varies based on an individual’s ethnic origin. CST IV was primarily identified in black and Hispanic women, but the other four groups (I, II, III, and V) were mainly separated from white and Asian women. Additionally, women’s vaginal pH varied by ethnic group, with Hispanic and black women having higher vaginal pHs than Asian and white women [20, 41].
Extensive interplay occurs between the host immune system and the microbiota. The recruitment of natural killer cells, macrophages, and T and B lymphocytes occurs as a result of microbial activation of toll-like receptors located on the vaginal epithelial cells. The chemokines include interleukins such as IL-1β, IL-8, and IL-10, as well as tumor necrosis factor-a (TNF-α). The study revealed that women who exhibited a dominant CST IV had significantly higher levels of IL-1α, IL-1β, and IL-8 than those with dominant CST I communities. Additionally, women with dominant L. iners communities (CST III) had moderate amounts of IL-8 compared to those with CST I. Chemokines may exhibit variability due to transitions between cervical microbial community state types (CSTs). For instance, women transitioning from CST I to CST III and CST IV demonstrated increased levels of IL-1α, IL-1β, and TNF-α with time [42, 43]. Table 1 lists the various CST categories.
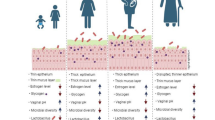
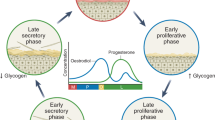
The vaginal microbiome of women is a multifaceted and ever-changing micro-ecosystem that transforms their lifespan and menstrual cycle. The vaginal mucosa is stratified into a squamous, non-keratinized epithelium and cervicovaginal discharge [8, 44]. Researchers must examine the human microbiome to determine whether microbial populations are consistent or vary by individual to understand the human body. Human health and sickness risk may be linked to core microbiota alterations [45]. The female vaginal microbiota may alter throughout several life phases, such as childhood, prepuberal, puberty, adulthood and menopause. In actuality, the prevalent elements that guided the temporal variations in human vaginal microbiota were menstruation, vaginal douching, uncontrolled antibiotic use, and hormonal fluctuations [46]. In childhood, vaginal pH is neutral, and several bacteria species, including gram-negative anaerobic, gram-positive anaerobic, and aerobic bacteria, predominate in the vaginal environment. Following the period of childhood, the vaginal epithelial cells undergo stimulation by the hormone estrogen, resulting in the production of glycogen. This process ultimately leads to the establishment of a dominant population of lactobacilli in the healthy vagina throughout the reproductive years. In the menopause stage, there is a loss in estrogen levels, resulting in a subsequent reduction in glycogen synthesis. The hormonal fluctuation leads to a decrease in the population of lactobacilli. Nevertheless, even during menopause, Lactobacillus spp. persists as a prominent presence in the vaginal microbiome, although with increased diversity in comparison to earlier stages of life [47, 48] (Fig. 1).
Recent research has undertaken a comparative examination of the vaginal microbiota’s makeup at three unique locations inside the vagina: the cervix, mid-vagina, and introitus. These studies have shown that the female vaginal ecosystem consists of a wide range of more than 200 phylotypes, with the most common taxa belonging to the phyla Firmicutes, Bacteroidetes, Actinobacteria, and Fusobacteria [49].

Alteration of the female vaginal microbiota throughout several life phases
Vaginal microbiota in gynecological cancers
Gynecologic cancer encompasses malignancies that arise from the female reproductive organs. There are five main categories of gynecologic cancer, including cervical, ovarian, endometrial, vaginal, and vulvar cancer [50]. The primary occupants of the vaginal microbiome consist of organisms belonging to the Firmicutes phylum and Lactobacillus genus. Evidence suggests a correlation between the development of gynecological malignancies and alterations in the microbial community, namely a reduction in commensal bacteria and an elevation in anaerobic bacteria [51, 52].
Cervical cancer
Cervical cancer, characterised by the malignant conversion of cells inside the cervix, which serves as the passage between the vagina and the uterus, has emerged as a notable public health issue in the contemporary era. The onset of this condition occurs when healthy cells undergo uncontrolled proliferation, resulting in the development of malignant tumors that can spread and cause harm to neighbouring vital organs [53]. In the year 2018, cervical cancer had a global impact on more than 500,000 women, leading to an estimated 300,000 fatalities. Cervical cancer ranks as the fourth most frequent kind of cancer among women [54]. According to projections made by the World Health Organization (WHO), it was estimated that 2020 there would be around 604,000 newly reported cases and 342,000 fatalities attributed to this particular ailment. If no significant intervention is implemented, it is projected that the worldwide prevalence of cervical cancer will increase to around 700,000 cases by the year 2030, resulting in an estimated 400,000 deaths. These findings indicate a rise of 21% in incidence rates and a 27% increase in fatality rates when compared to the data from 2018 [55].
The vaginal microbiota can influence the acquisition and long-term presence of the Human Papillomavirus (HPV), a viral infection that is linked to the development of cervical cancer [56]. HPV is generally recognized as the primary risk factor associated with the onset of cervical cancer since over 90% of cervical cancer cases are potentially linked to an HPV infection [57, 58]. HPV types 16 and 18 have been recognized as the most high-risk kinds and are linked to gynecologic malignancies and cervical cancer in women worldwide [59]. The known scientific literature has firmly proven the association between papillomaviruses and the transformative effects of the viral E6 and E7 oncoproteins [60, 61]. The protein encoded by the E6 oncogene can elicit cellular changes in the host cells. This phenomenon primarily targets the p53 protein, and its suppression may result in the development of malignant tumors and hinder the process of programmed cell death, known as apoptosis [62]. Conversely, the E7 protein has a complementary activity to the Retinoblastoma protein (PRb). The expression of E7 leads to the release of E2F, hence inducing DNA synthesis [63].
The cervical microenvironment is a multifaceted structure that includes immune cells and unique bacteria, which collaborate to orchestrate localized immune responses [64, 65]. Considerable research efforts have been dedicated to examining the intricate attributes of the cervical microenvironment to comprehend the interaction between the vaginal microbiota, HPV infection, various phases of cervical intraepithelial neoplasia (CIN), and cervical cancer. The research above indicates that an augmentation in bacterial variety and a reduction in lactobacilli may result in immunological dysregulation [66,67,68]. The phenomenon above gives rise to a milieu that is favorable for the proliferation of tumors, hence emphasizing the noteworthy influence of the vaginal microbiota as a contributing element in the progression of cervical cancer. The prevailing consensus in the scientific community is that while human HPV plays a significant role in the development of cervical cancer, it is not the only factor responsible for its occurrence [69].
Chen et al. studied Chinese cohorts to examine the connections between the vaginal microbiome, cervical cancer, CIN, and HPV infection. The study contrasted the makeup of vaginal microorganisms. Regardless of CINs, it was discovered that HPV infection enhanced the variety of vaginal microorganisms. Vaginal bacterial variety and richness were more significant in patients with cervical cancer. Among all the categories, Lactobacillus was the most prevalent. Lactobacillus, Gardnerella, and Atopobium decreased in response to HPV infection, but Prevotella, Bacillus, Anaerococcus, Sneathia, Megasphaera, Streptococcus, and Anaerococcus increased. The severity of CIN may have been influenced by a reduction in G. vaginalis and an increase in Bacillus and Anaerococcus. Megasphaera was significantly linked to HPV in the absence of CINs or malignant tumors. Prevotella amnii was the group’s most prevalent bacterium with low-grade squamous intraepithelial lesions (LSIL). At the family level, Prevotella timonensis, Shuttleworthia, and Streptococcaceae were linked to high-grade squamous lesions (HSIL) [70].
Mengying Wu et al. looked into whether the vaginal microbiota influences cervical squamous intraepithelial neoplasia development. They investigated sixty-nine women who came to the Obstetrics and Gynecology Hospital at Fudan University. Three groups of women were studied: those without intra-epithelial lesions or cancer, those with LSIL, and those with HSIL. The purpose of the study was to determine the makeup of vaginal bacteria in each group. In every group, Lactobacillus predominated. There were more streptococci and prevotella in the HSIL group. The development of cervical illness has been associated with the incidence of high-risk HPV infection. Those without intra-epithelial lesions or cancer had higher concentrations of the Pseudomonadales order, the Peptostreptococcaceae family, and other bacteria than those with squamous neoplasia. Comparing the LSIL and HSIL groups to the non-malignant group, Delftia enrichment is seen [71].
A 2021 research looked at the relationship between the emergence of SIL-related cervical cancer and the cervical microbiota of women who are of reproductive age. The research looked at 94 patients’ cervical mucous microbiota using 16 S rDNA. Alpha diversity was greater in severe cervical pathology, with less Lactobacillus and more anaerobes. Beta diversity varied greatly. Sneathia of HSIL and Porphyromonas, Prevotella, and Campylobacter of cervical cancer were marker genera. Prevotella had opposing functional pathway correlations with Lactobacillus [72].
The composition of vaginal bacteria in Mexican women with precancerous SIL was investigated by Nieves-Ramírez et al. The species richness rose, and the presence of SIL brought about compositional changes. The microbiota of HPV-positive women was altered even in the absence of SIL. According to research that used multivariate association with linear models (MaAsLin), HPV infection was associated with two operational taxonomic units (OTUs) reduction in L. iner and an increase in Brachybacterium conglomeratum and Brevibacterium aureum [73].
In five groups—normal, healthy, HR-HPV infections, LSIL, HSIL, and cervical cancer—Wei et al. looked at 59 Chinese women. Following HR-HPV infection and cervical lesions, there was a drop in Lactobacillus and an increase in the diversity of vaginal microbiota species. Actinobacteria, however, were significantly more prevalent in the four disease groups compared to the control group. Cervical cancer may also arise as a result of L. iners. After analyzing the whole process, Gardnerella, Atopobium, and Dialister impact HR-HPV persistence and etiology of cervical cancer [74].
Li et al.‘s 2023 study investigated cervical cancer, female vaginal microbiota, and immunological variables. Gardnerella, Prevotella, and Lactobacillus are the main bacteria in the vaginal flora. Prevotella, Ralstonia, Gardnerella, and Sneathia are more common in the group with cervical cancer than in the HPV-negative group. CIN with HPV positivity has a higher prevalence of Gardnerella, Prevotella, and Sneathia than non-CIN. The HPV-negative group has a preference for Atopobium and Lactobacillus [75]. Table 2 compiles the studies conducted on vaginal microbiota in cervical cancer patients.
Ovarian cancer
The development of abnormal cell proliferation inside the ovaries characterises ovarian cancer. Cellular proliferation occurs rapidly, leading to the infiltration and subsequent degradation of normal bodily tissues. The female reproductive system comprises a pair of ovaries situated bilaterally next to the uterus [76]. Ovarian cancer is one of the leading causes of death for women with gynecological cancers. It also comes in at number five among cancers diagnosed in females [77]. In the year 2020, the incidence of ovarian cancer was estimated to be about 21,750 cases, accounting for around 1.2% of the total number of cancer cases. The anticipated fatality rate linked to the occurrence is 13,940. The projected relative survival rate over five years is estimated to be 48.6%. The study determined that the average incidence rate per 100,000 people, after adjusting for age using the 2000 US standard population, was 11.1 throughout the period spanning from 2012 to 2016. The demographic group exhibiting the highest incidence rate is people who identify as non-Hispanic white, with a rate of 11.6 per 100,000. Subsequently, American Indians and Alaska Natives have a rate of 10.3 per 100,000, followed by Hispanics with a rate of 10.1 per 100,000. Non-Hispanic blacks and Asian and Pacific Islanders also show varying rates [78].
The etiology of ovarian cancer is not fully comprehended, despite the presence of established risk factors including family history, age, inflammation, reproductive variables, benign gynecologic diseases, gynecologic surgery, and genetic susceptibility linked to mutations in the BRCA1 and BRCA2 genes [79, 80]. According to recent studies, the pathogenesis of ovarian cancer is significantly influenced by the vaginal microbiota.
Zhou et al. investigated the diversity and composition of the microbiota in normal distal fallopian tube tissues (25 samples) and ovarian cancer tissues (25 samples) using high-throughput sequencing techniques that targeted the 16 S rRNA gene. Compared to the normal distal fallopian tube tissues, the high-throughput sequencing study revealed a substantial decrease in diversity and richness indices within the ovarian cancer tissues. Furthermore, there was a notable increase in the relative abundance of two phyla in ovarian cancer cases: Firmicutes and Proteobacteria. The research cited above suggests that changes in the makeup of microbes may have a role in the development of ovarian cancer [81].
Two groups of women, ages 18–87, from the Czech Republic, Germany, Italy, Norway, and the United Kingdom were involved in the Nené et al. study. Patients with epithelial ovarian cancer and healthy, benign gynecological controls comprised the first group. The second group included wild-type BRCA1 and BRCA2 controls (healthy individuals with benign gynecological diseases) and women with BRCA1 mutations but no ovarian cancer. Every person’s cervicovaginal samples were sequenced for 16 S rRNA. It was calculated what percentage of lactobacilli species in the cervicovaginal microbiota offer a protective low vaginal pH. At the same time, community-type O samples had fewer than 50% lactobacilli, and community-type L samples contained at least 50%. In both groups, there was a stronger correlation seen in younger persons between the community type O microbiota and the status of ovarian cancer or BRCA1 mutation. Age-matched controls were less likely to have a community-type O microbiota than those with ovarian cancer and BRCA1 mutation carriers under 50. This risk would increase if more first-degree relatives developed cancer. Ovarian cancer risk factors were significantly correlated with the community-type O cervicovaginal microbiota, including age and BRCA1 germline mutations. According to the research, restoring a community-type L microbiota may help prevent ovarian cancer [82].
The retrospective investigation by Jacobson et al. found that ovarian cancer patients had a lower prevalence of vaginal communities dominated by Lactobacillus. This discovery was made in light of previously collected data on cancer-free women of comparable age [83].
Huang et al. examined 30 patients in 2022 to compare the intratumor microbiomes of epithelial benign ovarian tumors (EBOTs) with epithelial ovarian cancer (EOC). For this, high-throughput sequencing was used. Additionally, according to the study, Propionibacterium acnes may hasten the evolution of EOC. Compared to EBOT tissues, EOC tissues exhibit a more varied and abundant intratumor microbiome, according to high-throughput sequencing. EOC tissues had a higher abundance of Actinomycetales, Acinetobacter, Streptococcus, Ochrobacterium, and Pseudomonas. The study discovered that the P. acnes strain had a significant impact on the development of EOC [84].
In the comprehensive study conducted by Yu et al., intraoperative swabs were procured from the fallopian tube and other surgical sites to serve as controls. The research study included a total of 187 participants, consisting of 81 individuals diagnosed with ovarian cancer and 106 individuals without cancer. 1001 swabs were collected and submitted to 16 S rRNA gene PCR and sequencing to investigate the microbial composition. The study discovered a total of 84 bacterial species that have the potential to constitute the microbiota of the fallopian tube. A notable alteration was seen in the microbiome composition of individuals diagnosed with ovarian cancer in comparison to those without the disease. Certain bacterial strains, Streptococcus parasanguinis, and Neisseriaceae, were seen to be more prevalent in samples collected from the fallopian tubes and ovarian surfaces of individuals diagnosed with ovarian cancer. In contrast, several bacterial species, such as Ruminiclostridium, Dialister invisus, and Bacteroides dorei, were found solely in individuals diagnosed with ovarian cancer, although with a very low abundance [85]. Table 3 summarises the studies conducted on vaginal microbiota in ovarian cancer patients.
Endometrial cancer
Endometrial cancer is a malignancy that originates from the endometrium, which is the epithelial lining of the uterus or womb. The phenomenon under consideration is the outcome of an atypical proliferation of cells with the capacity to infiltrate or disseminate to distant anatomical sites [86, 87]. Endometrial cancer is a prevalent malignancy that affects a significant population of women worldwide [88]. In 2018, it was anticipated that this disease would cause 63,230 new instances of endometrial cancer and 11,350 deaths. According to this, endometrial cancer ranks as the fifth most common cause of cancer-related death in the US and the fourth most common cancer among women [89]. In terms of endometrial cancer incidence, white women exhibited the highest rates across various ethnic groups, with a recorded incidence of 24.8 cases per 100,000 individuals. Conversely, the death rate for black women was found to be twice as high as that of white women, with rates of 7.3 cases per 100,000 individuals compared to 3.9 cases per 100,000 individuals, respectively [90].
Endometrial cancer can be caused by several things, such as environmental factors, genetic predisposition, hormonal imbalances (especially involving estrogen and progesterone), heavy periods, being overweight, or old age. Recent research indicates that the vaginal microbiome has the potential to influence the development of endometrial cancer via direct or indirect mechanisms. This effect could happen by interacting with vulnerable endometrial tissue or by making different metabolites and inflammatory factors [52, 91].
In research conducted in 2019, Walsh et al. identified Porphyromonas somerae as the predominant microbial marker associated with endometrial cancer. Targeted qPCR was used to validate this finding, suggesting that it may be useful in identifying endometrial cancer in high-risk, asymptomatic women. It is essential to do more studies to investigate P. somerae and its related network’s possible role in the development of endometrial cancer, given the pathogenic qualities shown in the context of tissue infections and ulcers [92].
The study performed in 2022 by Barczyński et al. included a cohort of 48 female participants who presented with various medical conditions, including endometrial cancer, endometrial atypical hyperplasia, and benign gynecological problems. The research used real-time PCR to determine the bacterial species present in the samples. About specific species, it was shown that the incidence of Mobiluncus curtisii and Fusobacterium nucleatum was significantly elevated in vaginal samples. In contrast, the occurrence of Gardnerella vaginalis and Fannyhessea vaginae was mostly seen in samples obtained from the endocervical canal. It is of significance to observe that patients who have been diagnosed with endometrial cancer have a greater prevalence of vaginal microbiota as compared to their endocervical canal. However, the female participants in the control group had comparable levels of isolated bacteria in both the vaginal and endocervical canal swabs. The study showed a significant decrease in the prevalence of Lactobacillus spp. and Bifidobacterium spp. in cancer patients compared to the control group. Moreover, a notable disparity was seen in the number of microorganisms obtained from endocervical canal swabs between women belonging to the control group and those diagnosed with endometrial cancer [93].
Semertzidou et al. performed a comprehensive investigation of many anatomical locations within the reproductive system, including the vagina, cervix, endometrium, fallopian tubes, and ovaries, in a cohort consisting of 61 individuals. A microbial continuity was seen in the vaginal tract of the majority of women without cancer, as shown by the consistent presence of bacterial species often found in this region. A correlation has been shown between endometrial cancer and a reduction in the presence of bacteria in both the cervicovaginal and rectal regions. Subsequently, there was a decline in the relative prevalence of Lactobacillus species, namely L. crispatus. Furthermore, an increase in the variety of bacteria was seen in conjunction with the expansion of several bacterial taxa, such as Porphyromonas, Prevotella, Peptoniphilus, and Anaerococcus, in both the lower genital tract and endometrium of persons who have been diagnosed with endometrial cancer [94].
The study done by Barczyński et al. included a cohort of 96 participants who had surgical interventions for a range of illnesses, such as benign uterine disorders, precancerous endometrial lesions, and endometrial cancer. The researchers used real-time PCR analysis to detect and measure the abundance of 19 commonly occurring bacteria in samples collected from the vaginal fornix and endocervical canal. Microbial presence was observed in 88.5% of vaginal samples and 83.3% of cervical samples. The prevalence of L. iners was higher among those with benign diseases, but Dialister pneumosintes and Mobiluncus curtisii were found to be more prevalent among cancer patients. The prevalence of these two bacteria was notably higher in vaginal samples obtained from individuals with endometrial cancer, suggesting a possible role in the initiation or promotion of carcinogenesis. Nevertheless, the precise process remains elusive, requiring further investigation [95]. Table 4 summarises the studies conducted on vaginal microbiota in endometrial cancer patients.
Vaginal cancer
Vaginal cancer is precisely characterised as a medical condition that lacks any indication of cervical or vulvar cancer or a documented history of either during the preceding five-year period. The majority of vaginal lesions (about 80–90%) are derived from cervical or vulvar lesions, as well as other nearby locations such as the endometrium, bladder, rectosigmoid, or ovary [96, 97]. Vaginal cancer is a very uncommon kind of malignancy, accounting for around 1–2% of all gynecologic cancers [98]. According to a survey provided by the American Cancer Society, it is projected that around 8,180 new instances of vaginal cancer will be detected in the year 2021. The projected fatality rate is estimated to be around 1530 individuals among the total number of diagnosed cases [99].
Squamous cell carcinoma is the predominant histological subtype of primary vaginal cancer, comprising about 90% of the total incidence. Additional, less prevalent variants include adenocarcinoma, clear cell adenocarcinoma, melanoma, sarcoma, and lymphoma affecting the vaginal region [100]. Vaginal cancer is correlated with several risk factors, such as infection with human papillomavirus (HPV) and herpes simplex virus (HSV), exposure to diethylstilbestrol, chronic irritation, prior radiation therapy, and an imbalance in the homeostatic equilibrium of the vaginal microbiota [52, 101].
A correlation has been shown between the composition of the vaginal microbiota and the occurrence of vaginal intraepithelial neoplasia (VAIN). The progression of VAIN and the development of vaginal cancer have been associated with changes in the composition of the vaginal microbiota, including an increased presence of Atopobium, Gardnerella, Enterococcus, Clostridium, and Allobaculum. Additionally, an elevated viral load of HPV-16, 52, and 58 has been identified as a contributing factor [9].
Vulvar cancer
Vulvar cancer is considered a relatively uncommon malignancy within the field of gynecology, accounting for around 4% of all malignancies affecting the female genital system [102]. Based on the figures presented by the International Agency for Research on Cancer (IARC), the yearly incidence of vulvar cancer exceeds 45,000 cases, with about 50.1% of these occurrences seen in countries with high-income economies [103]. This particular neoplasia is accountable for an estimated annual mortality rate of 17,000 individuals, primarily concentrated in high-income nations (40.8% of cases) [102].
Squamous cell carcinoma accounts for around 90% of vulvar malignancies. Less common histologic subtypes include basal cell carcinoma, verrucous carcinoma, Bartholin’s gland adenocarcinoma, extramammary Paget’s disease, and vulvar melanoma [102]. Various risk factors have been identified about the development of vulvar cancer. These include advancing age, infection with HPV, tobacco smoking, inflammatory vulvar diseases, previous pelvic radiation, and compromised immune function [104]. The presence of mucosal HPVs has been associated with the development of vulvar cancer in young women who have previously had genital warts, cervical dysplasia, and immunosuppression. The majority of vulvar cancer patients who test positive for HPV carry the HPV-16 strain, accounting for around 80–90% of cases. The remainder of individuals exhibit HPV18 or HPV33 [105]. Figure 2 illustrates the Alteration of the female vaginal microbiota throughout gynecological cancers.

Alteration of the female vaginal microbiota throughout gynecological cancers
Conclusions
Epidemiological research has shown a significant correlation between the makeup of the vaginal microbiota and the incidence of gynecological cancers. However, our understanding of the host’s defensive mechanisms against these microorganisms remains limited. Additional investigation is required to clarify the functional implications of these microbial communities on the vaginal microenvironment, particularly concerning tumor formation. Future investigations using extensive clinical datasets, in conjunction with in vitro models encompassing both human and animal participants, will assume a pivotal role in understanding the exact impact of these microorganisms on the genesis and advancement of gynecological malignancies. This will provide a more comprehensive comprehension of these bacteria’ functions in the beginning and progression of gynecological malignancies. In summary, a deeper understanding of the intricate interplay between the host and microorganisms in the vaginal area holds promise for uncovering novel strategies in the realm of cancer prevention and therapy. Consequently, this would enhance women’s overall welfare and physical state.
Data availability
No datasets were generated or analysed during the current study.
References
Hou K, Wu Z-X, Chen X-Y, Wang J-Q, Zhang D, Xiao C, et al. Microbiota in health and diseases. Signal Transduct Target Therapy. 2022;7(1):135.
Marchesi JR, Ravel J. The vocabulary of microbiome research: a proposal. Springer; 2015. pp. 1–3.
Petrillo F, Pignataro D, Lavano MA, Santella B, Folliero V, Zannella C, et al. Current evidence on the ocular surface microbiota and related diseases. Microorganisms. 2020;8(7):1033.
Song Q, Wang Y, Huang L, Shen M, Yu Y, Yu Q, et al. Review of the relationships among polysaccharides, gut microbiota, and human health. Food Res Int. 2021;140:109858.
Kwon MS, Lee HK. Host and microbiome interplay shapes the vaginal microenvironment. Front Immunol. 2022;13:919728.
Gupta P, Singh MP, Goyal K. Diversity of vaginal microbiome in pregnancy: deciphering the obscurity. Front Public Health. 2020;8:326.
Pacha-Herrera D, Erazo-Garcia MP, Cueva DF, Orellana M, Borja-Serrano P, Arboleda C, et al. Clustering analysis of the multi-microbial consortium by Lactobacillus species against vaginal dysbiosis among Ecuadorian women. Front Cell Infect Microbiol. 2022;12:863208.
Chen X, Lu Y, Chen T, Li R. The female vaginal microbiome in health and bacterial vaginosis. Front Cell Infect Microbiol. 2021;11:631972.
Gholiof M, Luca A-D, Wessels JM. The female reproductive tract microbiotas, inflammation, and gynecological conditions. Front Reproductive Health. 2022;4:963752.
Saraf VS, Sheikh SA, Ahmad A, Gillevet PM, Bokhari H, Javed S. Vaginal microbiome: normalcy vs dysbiosis. Arch Microbiol. 2021;203:3793–802.
Li X, Xu F, Sha X, Chen X, hui Pan Q, He X, et al. Biochemical indicators and vaginal microecological abnormalities indicate the occurrence of intrauterine adhesions. Med Microecology. 2022;11:100050.
Han Y, Liu Z, Chen T. Role of vaginal microbiota dysbiosis in gynecological diseases and the potential interventions. Front Microbiol. 2021;12:643422.
Keyvani V, Kheradmand N, Navaei ZN, Mollazadeh S, Esmaeili S-A. Epidemiological trends and risk factors of gynecological cancers: an update. Med Oncol. 2023;40(3):93.
Gil-Ibanez B, Davies-Oliveira J, Lopez G, Díaz-Feijoo B, Tejerizo-Garcia A, Sehouli J. Impact of gynecological cancers on health-related quality of life: historical context, measurement instruments, and current knowledge. Int J Gynecologic Cancer. 2023:ijgc–2023.
Hashemipour M, Boroumand H, Mollazadeh S, Tajiknia V, Nourollahzadeh Z, Borj MR, et al. Exosomal microRNAs and exosomal long non-coding RNAs in gynecologic cancers. Gynecol Oncol. 2021;161(1):314–27.
Tachedjian G, O’Hanlon DE, Ravel J. The implausible in vivo role of hydrogen peroxide as an antimicrobial factor produced by vaginal microbiota. Microbiome. 2018;6(1):1–5.
Lash AF, Kaplan B. A study of Döderlein’s vaginal bacillus. J Infect Dis. 1926:333–40.
Thomas S. Döderlein’s bacillus: Lactobacillus acidophilus. J Infect Dis. 1928:218–27.
Gajer P, Brotman RM, Bai G, Sakamoto J, Schütte UM, Zhong X, et al. Temporal dynamics of the human vaginal microbiota. Sci Transl Med. 2012;4(132):ra13252–52.
Ravel J, Gajer P, Abdo Z, Schneider GM, Koenig SS, McCulle SL et al. Vaginal microbiome of reproductive-age women. Proceedings of the National Academy of Sciences. 2011;108(supplement_1):4680-7.
Doyle R, Gondwe A, Fan Y-M, Maleta K, Ashorn P, Klein N, Harris K. A Lactobacillus-deficient vaginal microbiota dominates postpartum women in rural Malawi. Appl Environ Microbiol. 2018;84(6):e02150–17.
DiGiulio DB, Callahan BJ, McMurdie PJ, Costello EK, Lyell DJ, Robaczewska A, et al. Temporal and spatial variation of the human microbiota during pregnancy. Proc Natl Acad Sci. 2015;112(35):11060–5.
Van De Wijgert JH, Borgdorff H, Verhelst R, Crucitti T, Francis S, Verstraelen H, Jespers V. The vaginal microbiota: what have we learned after a decade of molecular characterization? PLoS ONE. 2014;9(8):e105998.
Oerlemans E, Ahannach S, Wittouck S, Dehay E, De Boeck I, Ballet N, et al. Impacts of menstruation, community type, and an oral yeast probiotic on the vaginal microbiome. Msphere. 2022;7(5):e00239–22.
Chee WJY, Chew SY, Than LTL. Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. Microb Cell Fact. 2020;19(1):203.
Zheng J, Wittouck S, Salvetti E, Franz CM, Harris HM, Mattarelli P, et al. A taxonomic note on the genus Lactobacillus: description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int J Syst Evol MicroBiol. 2020;70(4):2782–858.
De Seta F, Campisciano G, Zanotta N, Ricci G, Comar M. The vaginal community state types microbiome-immune network as key factor for bacterial vaginosis and aerobic vaginitis. Front Microbiol. 2019:2451.
Atassi F, Pho Viet Ahn DL, Lievin-Le Moal V. Diverse expression of antimicrobial activities against bacterial vaginosis and urinary tract infection pathogens by cervicovaginal microbiota strains of Lactobacillus gasseri and Lactobacillus crispatus. Front Microbiol. 2019;10:2900.
Cocomazzi G, De Stefani S, Del Pup L, Palini S, Buccheri M, Primiterra M, et al. The impact of the female genital microbiota on the outcome of assisted Reproduction treatments. Microorganisms. 2023;11(6):1443.
Witkin SS, Mendes-Soares H, Linhares IM, Jayaram A, Ledger WJ, Forney LJ. Influence of vaginal bacteria and D-and L-lactic acid isomers on vaginal extracellular matrix metalloproteinase inducer: implications for protection against upper genital tract infections. MBio. 2013;4(4). https://doi.org/10.1128/mbio. 00460 – 13.
Beghini J, Linhares I, Giraldo P, Ledger W, Witkin S. Differential expression of lactic acid isomers, extracellular matrix metalloproteinase inducer, and matrix metalloproteinase-8 in vaginal fluid from women with vaginal disorders. BJOG: Int J Obstet Gynecol. 2015;122(12):1580–5.
Ragaliauskas T, Plečkaitytė M, Jankunec M, Labanauskas L, Baranauskiene L, Valincius G. Inerolysin and vaginolysin, the cytolysins implicated in vaginal dysbiosis, differently impair molecular integrity of phospholipid membranes. Sci Rep. 2019;9(1):10606.
Rampersaud R, Planet PJ, Randis TM, Kulkarni R, Aguilar JL, Lehrer RI, Ratner AJ. Inerolysin, a cholesterol-dependent cytolysin produced by Lactobacillus iners. J Bacteriol. 2011;193(5):1034–41.
Macklaim JM, Fernandes AD, Di Bella JM, Hammond J-A, Reid G, Gloor GB. Comparative meta-RNA-seq of the vaginal microbiota and differential expression by Lactobacillus iners in health and dysbiosis. Microbiome. 2013;1(1):12.
Zheng N, Guo R, Wang J, Zhou W, Ling Z. Contribution of Lactobacillus iners to vaginal health and diseases: a systematic review. Front Cell Infect Microbiol. 2021;11:792787.
Spurbeck RR, Arvidson CG. Lactobacillus jensenii surface-associated proteins inhibit Neisseria gonorrhoeae adherence to epithelial cells. Infect Immun. 2010;78(7):3103–11.
Nowak RG, Randis TM, Desai P, He X, Robinson CK, Rath J, et al. Higher levels of a cytotoxic protein, vaginolysin, in Lactobacillus-deficient community state types at the vaginal mucosa. Sex Transm Dis. 2018;45(4):e14.
Sharifian K, Shoja Z, Jalilvand S. The interplay between human papillomavirus and vaginal microbiota in cervical cancer development. Virol J. 2023;20(1):73.
Borgdorff H, Gautam R, Armstrong SD, Xia D, Ndayisaba GF, van Teijlingen NH, et al. Cervicovaginal microbiome dysbiosis is associated with proteome changes related to alterations of the cervicovaginal mucosal barrier. Mucosal Immunol. 2016;9(3):621–33.
Ma B, Forney LJ, Ravel J. Vaginal microbiome: rethinking health and disease. Annu Rev Microbiol. 2012;66:371–89.
Günther V, Allahqoli L, Watrowski R, Maass N, Ackermann J, von Otte S, Alkatout I. Vaginal microbiome in reproductive medicine. Diagnostics. 2022;12(8):1948.
Smith SB, Ravel J. The vaginal microbiota, host defence and reproductive physiology. J Physiol. 2017;595(2):451–63.
Byrne EH, Doherty KE, Bowman BA, Yamamoto HS, Soumillon M, Padavattan N, et al. Cervicovaginal bacteria are a major modulator of host inflammatory responses in the female genital tract. Immunity. 2015;42(5):965–76.
Amabebe E, Anumba DO. The vaginal microenvironment: the physiologic role of lactobacilli. Front Med. 2018;5:181.
Dekaboruah E, Suryavanshi MV, Chettri D, Verma AK. Human microbiome: an academic update on human body site specific surveillance and its possible role. Arch Microbiol. 2020;202:2147–67.
Hickey RJ, Zhou X, Pierson JD, Ravel J, Forney LJ. Understanding vaginal microbiome complexity from an ecological perspective. Translational Res. 2012;160(4):267–82.
Punzón-Jiménez P, Labarta E. The impact of the female genital tract microbiome in women health and reproduction: a review. J Assist Reprod Genet. 2021;38(10):2519–41.
Tuniyazi M, Zhang N. Possible therapeutic mechanisms and future perspectives of vaginal microbiota transplantation. Microorganisms. 2023;11(6):1427.
Srinivasan S, Hoffman NG, Morgan MT, Matsen FA, Fiedler TL, Hall RW, et al. Bacterial communities in women with bacterial vaginosis: high resolution phylogenetic analyses reveal relationships of microbiota to clinical criteria. PLoS ONE. 2012;7(6):e37818.
Corpus CU, Vulva UOV. Gynecologic cancer incidence, United States—2012–2016. Centers for Disease Control and Prevention, US Department of Health and Human Services; 2019.
Champer M, Wong A, Champer J, Brito I, Messer P, Hou J, Wright J. The role of the vaginal microbiome in gynaecological cancer. BJOG: Int J Obstet Gynecol. 2018;125(3):309–15.
Łaniewski P, Ilhan ZE, Herbst-Kralovetz MM. The microbiome and gynaecological cancer development, prevention and therapy. Nat Reviews Urol. 2020;17(4):232–50.
Dau TN, Pham HA, Nguyen TB, Phan TTH, FINANCIAL AND MEDICAL BARRIERS AND, MOTIVATIONS OF CERVICAL CANCER SCREENING AMONG ETHNIC MINORITY WOMEN IN VIETNAM. J Pharm Negat Results. 2022:6282–99.
Membrilla-Beltran L, Cardona D, Camara-Roca L, Aparicio-Mota A, Roman P, Rueda-Ruzafa L. Impact of Cervical Cancer on Quality of Life and Sexuality in Female survivors. Int J Environ Res Public Health. 2023;20(4):3751.
Singh D, Vignat J, Lorenzoni V, Eslahi M, Ginsburg O, Lauby-Secretan B, et al. Global estimates of incidence and mortality of cervical cancer in 2020: a baseline analysis of the WHO Global Cervical Cancer Elimination Initiative. Lancet Global Health. 2023;11(2):e197–206.
Alimena S, Davis J, Fichorova RN, Feldman S. The vaginal microbiome: a complex milieu affecting risk of human papillomavirus persistence and cervical cancer. Curr Probl Cancer. 2022;46(4):100877.
Bonab FR, Baghbanzadeh A, Ghaseminia M, Bolandi N, Mokhtarzadeh A, Amini M, et al. Molecular pathways in the development of HPV-induced cervical cancer. EXCLI J. 2021;20:320.
Pappa KI, Lygirou V, Kontostathi G, Zoidakis J, Makridakis M, Vougas K, et al. Proteomic analysis of normal and cancer cervical cell lines reveals deregulation of cytoskeleton-associated proteins. Cancer Genomics Proteomics. 2017;14(4):253–66.
Mahmoodi P, Fani M, Rezayi M, Avan A, Pasdar Z, Karimi E, et al. Early detection of cervical cancer based on high-risk HPV DNA‐based genosensors: a systematic review. BioFactors. 2019;45(2):101–17.
Chiantore MV, Mangino G, Iuliano M, Capriotti L, Di Bonito P, Fiorucci G, Romeo G. Human papillomavirus and carcinogenesis: novel mechanisms of cell communication involving extracellular vesicles. Cytokine Growth Factor Rev. 2020;51:92–8.
Li Y, Yu T, Yan H, Li D, Yu T, Yuan T et al. Vaginal microbiota and HPV infection: novel mechanistic insights and therapeutic strategies. Infect drug Resist. 2020:1213–20.
Li S, Hong X, Wei Z, Xie M, Li W, Liu G, et al. Ubiquitination of the HPV oncoprotein E6 is critical for E6/E6AP-mediated p53 degradation. Front Microbiol. 2019;10:2483.
Zhou L, Ng DS-C, Yam JC, Chen LJ, Tham CC, Pang CP, Chu WK. Post-translational modifications on the retinoblastoma protein. J Biomed Sci. 2022;29(1):1–16.
Audirac-Chalifour A, Torres-Poveda K, Bahena-Román M, Téllez-Sosa J, Martínez-Barnetche J, Cortina-Ceballos B, et al. Cervical microbiome and cytokine profile at various stages of cervical cancer: a pilot study. PLoS ONE. 2016;11(4):e0153274.
Cascardi E, Cazzato G, Daniele A, Silvestris E, Cormio G, Di Vagno G, et al. Association between cervical microbiota and HPV: could this be the key to complete cervical cancer eradication? Biology. 2022;11(8):1114.
Mitra A, MacIntyre DA, Lee Y, Smith A, Marchesi JR, Lehne B, et al. Cervical intraepithelial neoplasia disease progression is associated with increased vaginal microbiome diversity. Sci Rep. 2015;5(1):16865.
Garrett WS. Cancer and the microbiota. Science. 2015;348(6230):80–6.
Curty G, de Carvalho PS, Soares MA. The role of the cervicovaginal microbiome on the genesis and as a biomarker of premalignant cervical intraepithelial neoplasia and invasive cervical cancer. Int J Mol Sci. 2019;21(1):222.
Kwasniewski W, Wolun–Cholewa M, Kotarski J, Warchol W, Kuzma D, Kwasniewska A, Gozdzicka–Jozefiak A. Microbiota dysbiosis is associated with HPV–induced cervical carcinogenesis. Oncol Lett. 2018;16(6):7035–47.
Chen Y, Qiu X, Wang W, Li D, Wu A, Hong Z, et al. Human papillomavirus infection and cervical intraepithelial neoplasia progression are associated with increased vaginal microbiome diversity in a Chinese cohort. BMC Infect Dis. 2020;20:1–12.
Wu M, Gao J, Wu Y, Li Y, Chen Y, Zhao F et al. Characterization of vaginal microbiota in Chinese women with cervical squamous intra-epithelial neoplasia. Int J Gynecologic Cancer. 2020;30(10).
Wu S, Ding X, Kong Y, Acharya S, Wu H, Huang C, et al. The feature of cervical microbiota associated with the progression of cervical cancer among reproductive females. Gynecol Oncol. 2021;163(2):348–57.
Nieves-Ramírez M, Partida-Rodríguez O, Moran P, Serrano-Vázquez A, Pérez-Juárez H, Pérez-Rodríguez M, et al. Cervical squamous intraepithelial lesions are associated with differences in the vaginal microbiota of Mexican women. Microbiol Spectr. 2021;9(2):e00143–21.
Wei B, Chen Y, Lu T, Cao W, Tang Z, Yang H. Correlation between vaginal microbiota and different progression stages of cervical cancer. Genet Mol Biology. 2022;45:e20200450.
Li X, Wu J, Wu Y, Duan Z, Luo M, Li L et al. Imbalance of vaginal microbiota and immunity: two main accomplices of Cervical Cancer in Chinese women. Int J Women’s Health. 2023:987–1002.
Pattanaik S, Sharma D. Impact of ovarian cancer on hormone imbalance and its remedies with ayurvedic medicines. J Med Plants. 2022;10(2):34–9.
Height T, Ovarian, Epithelial. Fallopian Tube, and Primary Peritoneal Cancer Treatment (PDQ®) Health Professional Version.
Siegel Rebecca L, Miller Kimberly D, Ahmedin J. Cancer statistics, 2020. Ca Cancer J Clin. 2020;70(1):7–30.
Reid BM, Permuth JB, Sellers TA. Epidemiology of ovarian cancer: a review. Cancer Biology Med. 2017;14(1):9.
Liu Y-B, Mei Y, Long J, Zhang Y, Hu D-L, Zhou H-H. RIF1 promotes human epithelial ovarian cancer growth and progression via activating human telomerase reverse transcriptase expression. J Experimental Clin Cancer Res. 2018;37:1–15.
Zhou B, Sun C, Huang J, Xia M, Guo E, Li N, et al. The biodiversity composition of microbiome in ovarian carcinoma patients. Sci Rep. 2019;9(1):1691.
Nené NR, Reisel D, Leimbach A, Franchi D, Jones A, Evans I, et al. Association between the cervicovaginal microbiome, BRCA1 mutation status, and risk of ovarian cancer: a case-control study. Lancet Oncol. 2019;20(8):1171–82.
Jacobson D, Moore K, Gunderson C, Rowland M, Austin R, Honap TP, et al. Shifts in gut and vaginal microbiomes are associated with cancer recurrence time in women with ovarian cancer. PeerJ. 2021;9:e11574.
Huang Q, Wei X, Li W, Ma Y, Chen G, Zhao L, et al. Endogenous Propionibacterium acnes promotes ovarian Cancer Progression via regulating hedgehog signalling pathway. Cancers. 2022;14(21):5178.
Yu B, Liu C, Proll S, Mannhardt E, Liang S, Srinivasan S et al. Identification of fallopian tube microbiota and its association with ovarian cancer: a prospective study of intraoperative swab collections from 187 patients. medRxiv. 2023:2023.06. 28.23291999.
Walker JL, Nuñez ER. Endometrial cancer. Cancer Screening: CRC; 2021. pp. 531–56.
PDQATE B. Endometrial Cancer treatment (PDQ (R)): Patient Version. PDQ Cancer Information Summaries Bethesda: National Cancer Institute (US). 2002.
Zhang S, Gong T-T, Liu F-H, Jiang Y-T, Sun H, Ma X-X, et al. Global, regional, and national burden of endometrial cancer, 1990–2017: results from the global burden of disease study, 2017. Front Oncol. 2019;9:1440.
Brooks RA, Fleming GF, Lastra RR, Lee NK, Moroney JW, Son CH, et al. Current recommendations and recent progress in endometrial cancer. Cancer J Clin. 2019;69(4):258–79.
Terplan M, Schluterman N, McNamara EJ, Tracy JK, Temkin SM. Have racial disparities in ovarian cancer increased over time? An analysis of SEER data. Gynecol Oncol. 2012;125(1):19–24.
Dossus L, Rinaldi S, Becker S, Lukanova A, Tjonneland A, Olsen A, et al. Obesity, inflammatory markers, and endometrial cancer risk: a prospective case–control study. Endocrine-related Cancer. 2010;17(4):1007.
Walsh DM, Hokenstad AN, Chen J, Sung J, Jenkins GD, Chia N, et al. Postmenopause as a key factor in the composition of the Endometrial Cancer Microbiome (ECbiome). Sci Rep. 2019;9(1):19213.
Barczyński B, Frąszczak K, Korona-Głowniak I, Malm A, Kotarski J. 2022-RA-1627-ESGO vaginal microbiome in patients with endometrial cancer. BMJ Specialist Journals; 2022.
Semertzidou A, Whelan E, Smith A, Ng S, Brosens J, Marchesi J et al. Microbial signatures and continuum in endometrial cancer and benign patients. 2022.
Barczyński B, Frąszczak K, Grywalska E, Kotarski J, Korona-Głowniak I. Vaginal and cervical microbiota composition in patients with endometrial Cancer. Int J Mol Sci. 2023;24(9):8266.
Adams TS, Cuello MA. Cancer of the vagina. Int J Gynecol Obstet. 2018;143:14–21.
Ng QJ, Namuduri RP, Yam KL, Lim-Tan SK. Vaginal metastasis presenting as postmenopausal bleeding. Singapore Med J. 2015;56(8):e134.
Adhikari P, Vietje P, Mount S. Premalignant and malignant lesions of the vagina. Diagn Histopathology. 2017;23(1):28–34.
Sinno AK, Saraiya M, Thompson TD, Hernandez BY, Goodman MT, Steinau M, et al. Human papillomavirus genotype prevalence in invasive vaginal cancer from a registry-based population. Obstet Gynecol. 2014;123(4):817.
Kulkarni A, Dogra N, Zigras T. Innovations in the management of vaginal cancer. Curr Oncol. 2022;29(5):3082–92.
Bhat R. Vaginal Cancer: Epidemiology and Risk Factors. Preventive Oncology for the Gynecologist. 2019:309 – 14.
Pedrão PG, Guimarães YM, Godoy LR, Possati-Resende JC, Bovo AC, Andrade CEMC, et al. Management of early-stage vulvar cancer. Cancers. 2022;14(17):4184.
Organization WH. Global Cancer Observatory-estimated number of new cases in 2020, worldwide, females, all ages. Ginebra, Suiza: WHO; 2020. Acceso 5 de julio de 2022.
Canavan TP, Cohen D. Vulvar cancer. Am Family Phys. 2002;66(7):1269–75.
Wahid M, Dar SA, Jawed A, Mandal RK, Akhter N, Khan S, et al. editors. Microbes in gynecologic cancers: causes or consequences and therapeutic potential. Seminars in cancer biology. Elsevier; 2022.
Acknowledgements
None.
Funding
This paper was not funded.
Author information
Authors and Affiliations
Contributions
KJ, MK, EFS, MR: design of study. KJ, MK, EFS, MR: acquisition of data. KJ, MK, EFS, MR: evaluation of data, preparation of the manuscript. KJ, MK, EFS, MR: assessment of data. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Javadi, K., Ferdosi-Shahandashti, E., Rajabnia, M. et al. Vaginal microbiota and gynecological cancers: a complex and evolving relationship. Infect Agents Cancer 19, 27 (2024). https://doi.org/10.1186/s13027-024-00590-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13027-024-00590-7