
Abstract
Background
Single nucleotide polymorphisms occurring in the Plasmodium falciparum multidrug resistant gene 1 (pfmdr1) are known to be associated with aminoquinoline resistance and, therefore, represent key P. falciparum markers for monitoring resistance both in susceptible groups (children under 5 years old and pregnant women) and in the general population. This study aimed to determine prevalence and factors associated with the carriage of pfmdr1 N86Y, Y184F and D1246Y polymorphisms among pregnant women in a setting of high malaria transmission in Burkina Faso.
Methods
Plasmodium falciparum isolates were collected at the first antenatal care visit (ANC-1) as well as at delivery from pregnant women participating in the COSMIC trial (NTC01941264), which assessed malaria preventive interventions during pregnancy in the Nanoro Health District. Here, pregnant women received intermittent preventive treatment with sulfadoxine-pyrimethamine (IPTp-SP) and malaria infections and/or diseases were treated using artemether-lumefantrine (AL) during the trial. Parasite DNA was extracted from dried blood spots and the presence of pfmdr1 mutations at positions 86, 184 and 1246 was determined using nested PCR, followed by restriction fragment length polymorphism (RFLP) analysis.
Results
A prevalence of 13.2% (20/151) and 12.1% (14/116) of the pfmdr1 86Y mutant allele was found at ANC-1 and at delivery, respectively, while no mutant allele was observed for Y184F and D1246Y codons at both ANC-1 and at delivery. There were no significant factors associated with pfmdr1 86Y mutant allele carriage at ANC-1. However, malaria infections at delivery with a parasite density above the median (2237.2 (IQR: 613.5–11,425.7) parasites/µl) was associated with an increase risk of pfmdr1 86Y mutant allele carriage (AOR = 5.5 (95% CI 1.07–28.0); P = 0.04). In contrast, both three or more IPTp-SP doses (AOR = 0.25 (95% CI 0.07–0.92); P = 0.04) and one or more AL treatment (AOR = 0.25 (95% CI 0.07–0.89); P = 0.03) during pregnancy were associated with a significant reduce risk of pfmdr1 86Y mutant allele carriage at delivery.
Conclusion
These findings suggest that both high coverage of IPTp-SP and the use of AL for the treatment of malaria infection/disease during pregnancy select for pfmdr1 N86 wild-type allele at delivery.
Similar content being viewed by others
Background
Malaria in pregnancy (MiP) is a significant public health problem, with substantial adverse effects on both mother and fetus, including maternal anaemia, fetal loss, premature delivery, intrauterine growth retardation, and delivery of low birth-weight infants, which is a risk factor for death [1,2,3,4]. MiP current control measures used in most endemic countries, according to WHO recommendations, include the use of insecticide treated nets (ITNs), intermittent preventive treatment with sulfadoxine-pyrimethamine (IPTp-SP) and effective case management of malaria, which, since 2010, includes the use of artemisinine-based combination therapy (ACT) [5, 6].
Current efforts of malaria control during pregnancy rely mostly on the effectiveness of anti-malarial drugs used for both IPTp and case management. Indeed, although there are other limiting factors, including low attendance rate of antenatal services [7,8,9,10], low coverage and compliance to the preventive treatment by pregnant women [11,12,13] and inadequate protection of fewer than three SP doses where malaria transmission is intense [12, 14], the major hindrance of the effectiveness of IPTp-SP policy is the spread of Plasmodium falciparum resistance to SP [15,16,17,18,19,20]. Given the number of studies reporting the increased resistance of P. falciparum to SP, there has been several responses: (i) the increase of IPTp-SP doses by WHO in 2012 [21]; (ii) the evaluation of alternative drugs to SP [22, 23]; and, (iii) the assessment of alternative or improved strategies to IPTp-SP [24]. However, none of the alternative drugs or strategies tested has prompted the replacement of the IPTp-SP policy [25, 26]. Reducing the burden of malaria during pregnancy in high transmission setting remains challenging.
Since 2015, WHO recommends the use of ACT for the treatment of P. falciparum uncomplicated malaria during the second and third trimester of pregnancy [27, 28], and such recommendation has already been adopted and implemented by all sub-Saharan African countries [29]. However, there is limited knowledge on the effect of ACT, treatments such as artemether-lumefantrine (AL), on the selection of P. falciparum resistance markers during pregnancy that can affect the treatment outcome. Indeed, available information in non-pregnant women have shown that the P. falciparum multidrug resistance 1 (pfmdr1) N86 and D1246 alleles might be associated with AL resistance [30, 31]. In addition, the combination of N86, 184F and D1246, forming the ‘NFD’ haplotype, led to a decreased susceptibility to AL and treatment with AL selects for such a haplotype [32,33,34]. There is therefore a need for pharmacovigilance studies to monitor any delayed parasite clearance by AL and to assess risk factors associated with the carriage of P. falciparum resistance markers.
Between 2013 and 2016, a multi-centre, cluster-randomized, controlled trial (COSMIC) was conducted in three West African countries with high (Burkina Faso, Benin) and low (The Gambia) malaria transmission, to assess the protective efficacy of adding community-scheduled screening and treatment of malaria during pregnancy (CSST) to standard IPTp-SP (CSST/IPTp-SP) [35, 36]. The CSST/IPTp-SP strategy was based on monthly active follow-up by community health workers using rapid diagnostic tests (RDTs). The aim of the combined CSST/IPTp-SP strategy was to provide an opportunity to detect and treat malaria infections during pregnancy with AL and reduce the prevalence of placental malaria [35]. As part of the COSMIC trial, it has shown a high prevalence of the triple dhfr mutation with presence of quintuple mutants (triple dhfr and double dhps) in Burkina Faso, confirming concerns about the efficacy of IPTp-SP in the near future [37]. This study aimed to determine the prevalence and factors associated with the carriage of pfmdr1 polymorphisms (pfmdr1 N86Y7, Y184F, D1246Y) among pregnant women within the COSMIC trial in Burkina Faso.
Methods
Study site
This study was conducted in Nanoro Health District (NHD) in Burkina Faso between 2013 and 2016. NHD is a rural setting located in the centre-west region of the country, 85 km from the capital city, Ouagadougou. Malaria transmission in the region is endemic and highly seasonal. There is year-round malaria transmission with high transmission season occurring from July to December, corresponding to the wet period in the country. Most malaria cases are due to P. falciparum.
Study participants and sample size
The samples analysed in this study were collected from pregnant women enrolled in a clinical trial assessing the effectiveness of MiP preventive treatments, known as COSMIC trial (Clinical Trials.gov Identifier: NCT1941264). In the COSMIC trial, the protective efficacy of adding CSST during pregnancy to the standard IPTp-SP (CSST/IPTp-SP, intervention arm) was compared to IPTp-SP alone (control arm) [35]. CSST intervention was implemented by community health workers through monthly screening using RDTs and treatment of malaria infection with AL. AL treatment was given for both malaria episodes and infections detected by RDTs in the intervention arm, whereas in the control arm (standard IPTp-SP alone), AL was given for clinical episode treatment only, according to the national guidelines.
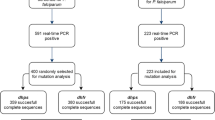
For the present analysis, a total of 324 dried blood spots (DBS) collected from finger prick at enrolment (first antenatal care visit (ANC-1)) and at delivery were selected from participants in Burkina Faso based on light microscopy (LM) results as shown in Fig. 1: (i) all available DBS collected at delivery from pregnant women with a P. falciparum infection (N = 162); (ii) all available DBS collected at ANC-1 from pregnant women who experienced P. falciparum infection both at recruitment and delivery (N = 42); and, (iii) a random selection of DBS collected at ANC-1 from pregnant women with a P. falciparum infection (N = 120).

Flow diagram of samples selection
Laboratory methods
DNA extraction
Plasmodium falciparum genomic DNA was extracted using the QIAamp 96 DNA Blood Kit (Qiagen, Germany) following the manufacturer’s instructions.
PCR-RLFP
pfmdr1 86Y, 184F and 1246Y mutations were determined using nested PCR followed by restriction fragment length polymorphism (RFLP) analysis as previously described [38]. PCR products were digested with AflIII (NEB), DraI (NEB), and EcoRV (NEB) for the determination of pfmdr1 N86Y, Y184F, and D1246Y alleles, respectively (Table 1). The digested products were visualized by electrophoresis using 2.5% agarose gel for 2 h at 80 V. The nested PCR and digestion reactions were run using 3D7 as wild-type control for the three SPNs and V1 (for N86Y) and 7G8 (for Y184F and D1246Y) as mutant controls.
Sulfadoxine-pyrimethamine resistance markers
The assessment of P. falciparum mutations in the dhfr gene (codons N51, C59, S108) and the dhps gene (codons A437, K540) was performed as part of the COSMIC trial [35, 37] and, re-analysed in this manuscript. The dhfr and dhps genes were amplified by nested PCR and products were sequenced to identify the targeted mutations as previously reported [37].
Statistical analysis
Data were analysed using STATA version 14.0 (StataCorp, USA). The pfmdr1 genotype profile was determined according to the presence or absence of wild/mutant alleles. Samples in which both wild and mutant alleles were detected were considered as mutant allele carriers. Differences between samples collected at ANC-1 and at delivery were estimated using the Chi-square test for proportions. Factors associated with pfmdr1 86Y mutant allele carriage were assessed by univariate and multivariable logistic regression analyses. Variables with P values ≤ 0.10 in univariate analyses were included in the multivariable models. The investigated variables included: MiP preventive strategy, parasitaemia, age, gravidity, number of malaria episodes, number of IPTp-SP doses received during pregnancy, AL treatment during pregnancy, haemoglobin levels at delivery, and bed-net usage. P values less than 0.05 were considered statistically significant.
Results
Characteristics of study population
The characteristics of the study participants are shown in Table 2. The mean age of pregnant women infected both at ANC-1 and at delivery (21.5 ± 5.6 years) was significantly lower than that of women infected only at ANC-1 (23.9 ± 5.7 years) and of women infected only at delivery (26.5 ± 6.1 years) (P < 0.001). The median parasite density was 980.5 (IQR: 412.5–2,949.2) parasites/µl and 2,237.2 (IQR: 613.5–11,425.7) parasites/µl at ANC-1 and at delivery, respectively (P = 0.02). Among women infected at ANC-1, the median parasite density was significantly higher among those infected both at ANC-1 and at delivery (1,752.5 (IQR: 558.5–5,373.5) parasites/µl) compared to those infected only at ANC-1 (903 (IQR: 324.5–2,308.5) parasites/µl; P = 0.01). Among women infected at delivery, there was no significant difference of the median parasite density between those infected only at delivery and those infected both at ANC-1 and at delivery (P = 0.84). There was no significant difference of the proportion of women who benefited from CSST intervention between the three groups (P = 0.40). The proportion of primigravid and secondigravid women infected both at ANC-1 and at delivery was significantly higher than that of women infected only at ANC-1 and those infected only at delivery (P < 0.001).
Among pregnant women infected at ANC-1, there was a significant difference of bed-net use between those infected both at ANC-1 and at delivery (72.7% (24/42)) and those only infected at ANC-1 (92.8% (103/120); P = 0.004). Such a difference was not observed among women infected at delivery (P = 0.30). Among pregnant women who experienced malaria infection at delivery, 69% (29/42) of those infected both at ANC-1 and at delivery received at least one AL treatment during pregnancy against 44.2% (53/120) of those infected only at delivery (P = 0.02). There was a significant difference of the time of AL treatment to delivery between women infected only at delivery and that of women infected both at ANC-1 and at delivery (P = 0.05). The proportion of women who received at least three IPTp-SP doses was not significantly different between those infected only at delivery and that of those infected both at ANC-1 and at delivery (P = 0.21). The carriage of triple dhfr 51/59/108 mutation was similar among women infected only at delivery and those infected both at ANC-1 and at delivery (P = 0.60). Only two women were found to carry a double dhps 437/540 mutant allele at delivery. Of note, data on dhfr-dhps mutations among the study popualtion were obtained from the main COSMIC trial [36, 37] and re-analysed in this manuscript.
Plasmodium falciparum resistance markers genotyping success rate
In total, the success rate for pfmdr1 D1246Y genotyping was 92.0% (298/324), with rates of 90.7% (147/162) and 93.2% (151/162) for samples collected at ANC-1 and at delivery, respectively. The pfmdr1 Y184F and N86Y genotyping showed similar success rates as the samples underwent the same nested PCR procedure. The success rate for the samples collected at ANC-1 was 93.2% (151/162), while the success rate for the samples collected at delivery was 71.6% (116/162), giving a total success rate of 82.4% (267/324) for both Y184F and N86Y codons.
Prevalence of pfmdr1 alleles among the study population
The mutant pfmdr1 86Y allele was detected among the study participants with a prevalence of 13.2% (20/131) and 12.1% (14/116) at ANC-1 and at delivery, respectively (P = 0.77). Among women who experienced malaria infection both at ANC-1 and at delivery, the prevalence of the mutant pfmdr1 86Y allele was 7.9% (3/38) and 16.1% (5/31) at ANC-1 and at delivery, respectively (P = 0.25). There was no significant difference of the proportion of pfmdr1 86Y allele carriage between women infected at ANC-1 and those infected at delivery for both women who received the standard IPTp-SP treatment (P = 0.63) and those who received the CSST/IPTp-SP treatment (P = 0.38). By contrast, no mutant alleles corresponding to pfmdr1 Y184F and D1246Y codons were observed in the samples collected from either ANC-1 or delivery (Table 3).
Factors associated with pfmdr1 86Y mutant allele carriage at ANC-1 and at delivery
Univariate and multivariable logistic regression analyses were performed to assess the factors associated with the pfmdr1 86Y mutant allele at ANC-1 and at delivery (Tables 4 and 5). None of the variables investigated in the univariate analysis was significantly associated with the carriage of pfmdr1 86Y mutant allele at ANC-1 (P > 0.1). Consequently, no multivariable analysis was undertaken for further assessment (Table 4). At delivery, univariate analyses showed a tendency of an increase risk of pfmdr1 86Y mutant allele carriage with parasite density above the median (P = 0.08). By contrast, there was a tendency towards reduced risk of pfmdr1 86Y mutant allele carriage among women who received at least three IPTp-SP doses during pregnancy (P = 0.06) and those who received at least one AL treatment during pregnancy (P = 0.10). In multivariable analyses, these associations were confirmed as infections at delivery with a parasite density more than the median (2,237.2 (IQR: 613.5–11,425.7) parasites/µl) was associated with an increase risk of pfmdr1 86Y mutant allele carriage (AOR = 5.5 (95% CI 1.07–28.0); P = 0.04). In addition, both three or more IPTp-SP doses (AOR = 0.25 (95% CI 0.07–0.92); P = 0.04) and one or more AL treatment (AOR = 0.25 (95% CI 0.07–0.89); P = 0.03) during pregnancy were associated with a significant reduce risk of pfmdr1 86Y mutant allele carriage at delivery. None of the other variables of interest, including MiP preventive strategy, triple dhrf 51/59/108 mutation, timing of AL treatment to delivery, gravidity, and malaria infection both at ANC-1 and at delivery, showed a significant association with pfmdr1 86Y mutant allele carriage (Table 5).
Discussion
Despite the widespread implementation of IPTp-SP to prevent MiP, pregnant women in endemic countries often experience peripheral and/or placental malaria infection at delivery [15, 39, 40]. Although WHO revised IPTp-SP guidelines and increased the SP dose, which has been shown to improve birth outcomes [41, 42], IPTp-SP strategy is still threatened by increasing Plasmodium falciparum resistance to SP. Consequently, there is a need to develop new alternative or improved strategies as part of IPTp-SP policy. In line with the latter, CSST of MiP, in addition to standard IPTp-SP (CSST/IPTp-SP), was tested in Burkina Faso, Benin and The Gambia as an intervention to improve maternal health and birth outcomes in areas of different malaria transmission intensity (COSMIC trial, NCT1941264) [35]. In such a context, the current study was conducted to determine prevalence and factors associated with the carriage of pfmdr1 polymorphisms among pregnant women participating in the COSMIC trial in Burkina Faso.
Mutations in the gene-encoding pfmdr1 are known to be associated with aminoquinoline resistance [43], and therefore represent key P. falciparum markers for monitoring resistance in both susceptible groups (children under 5 years and pregnant women) and the general population. In this study, the analysis was focused on: (i) mutations in pfmdr1 N86Y and D1246Y codons, which have been associated with resistance to chloroquine and amodiaquine [44,45,46], whereas wild-type sequences in these alleles were associated with reduced sensitivity to lumefantrine [44, 47, 48]; and, (ii) mutation in the pfmdr1 Y184F codon, which was associated with altered sensitivity to artemisinins and mefloquine [49]. A prevalence of 13.2 and 12.1% of the pfmdr1 86Y mutant allele was found at ANC-1 and at delivery, respectively. No mutant allele was observed for pfmdr1 Y184F and pfmdr1 D1246Y codons at both ANC-1 and at delivery. The observed prevalence of these mutations at positions 86, 184 and 1246 in this study lack comparable data in the country as previous studies differ with regard to study population (children vs adults vs pregnant women), Plasmodium falciparum isolates (clinical vs asymptomatic infections), and study periods [50,51,52,53]. However, looking at available reports in the study area, these results showed a higher prevalence of the pfmdr1 86Y mutant allele in pregnant women at ANC-1 (13.2%) compared to that in patients with uncomplicated malaria two years before (pre-treatment prevalence of 8.3% in 2010–2012) [50]. An early study conducted by the time of adoption of ACT as a first-line treatment for uncomplicated falciparum malaria in the country (2005), reported a higher prevalence of pfmdr1 86Y mutant allele (35–40%) in children aged 6–59 months with uncomplicated malaria [51]. Surprisingly, no pfmdr1 184F mutant allele was detected among the study population while a prevalence of about 50% was reported among P. falciparum uncomplicated malaria patients in 2010–2012 [50]. In the same study, only three pfmdr1 1246Y mutant alleles were detected in 660 isolates, corresponding to a prevalence of 0.4%.
It has been shown that wild-type sequences of pfmdr1 N86Y, Y184F and D1246Y codons are selected by prior use of AL treatment in malaria patients [47, 48, 51,52,53]. By contrast, selection of the pfmdr1 184F mutant allele has been observed in prior therapy with AL in malaria patients in Uganda [47]. To explore the potential selection of wild-type/mutant sequences of pfmdr1 polymorphisms following AL treatments during pregnancy, the prevalence of pfmdr1 86Y mutant alleles at ANC-1 was compared to that at delivery. No significant difference of the pfmdr1 86Y mutant allele was found neither in the general study population (P = 0.77) nor in sub-groups represented by women infected both at ANC-1 and at delivery (P = 0.25), women who received the standard IPTp-SP (P = 0.63) and women who benefitted from additional screening and treatment of P. falciparum infections (asymptomatic infections) using AL (P = 0.38).
To further assess the potential selection of pfmdr1 N86Y mutant/wild-type alleles by specific factors, factors associated with pfmdr1 86Y mutant allele carriage at ANC-1 and at delivery were investigated. Among the variables evaluated, none was significantly associated with pfmdr1 86Y mutant allele carriage at ANC-1. However, Plasmodium falciparum infection at delivery with high parasitaemia was significantly associated with nearly 5.5 times increase risk of pfmdr1 86Y mutant allele carriage at delivery after adjusting by confounding factors (P = 0.04). By contrast, uptake of at least three IPTp-SP doses and at least one AL treatment were found to be significant protective factors against pfmdr1 86Y mutant allele carriage at delivery in multivariate analyses (75% reduction for both with P = 0.04 and P = 0.03, respectively). These results suggest a benefit in reducing the risk of aminoquinoline resistance marker carriage by high coverage of IPTp-SP doses, which has been shown to reduce parasite load in P. falciparum infection during pregnancy [54, 55]. Moreover, these findings suggest a positive selection of pfmdr1 N86 wild-type allele at delivery following AL treatment during pregnancy in women receiving IPTp-SP, similar to that reported in non-pregnant women in Southeast Asia (Thailand) and East Africa (Kenya) [30, 31] and in patients of all age groups in West Africa (Burkina Faso) [50].
Studies have demonstrated that recent AL use has more of an impact on pfmdr1 N86 wild-type allele prevalence than less recent AL use, as lumefantrine levels decline over time, resulting in less selection [56, 57]. On the other hand, it has been shown that AL could select for pfmdr1 N86 wild-type allele a few months post-treatment in children [58]. This could be explained by genetic variations or other factors including the administration of AL with fatty foods, leading some individuals to exhibit longer artemether or lumefantrine half-lives than other, allowing longer periods of selection [58, 59]. Although, no evidence of an impact of the timing of AL treatment to delivery on the selection of pfmdr1 N86 wild-type allele was observed in this study, the existence of a specific selective window in pregnant women should not be ruled out given the limited number of pfmdr1 86Y mutant allele carriers. In addition, the sub-group of women who experienced malaria infection both at ANC-1 and at delivery did not show a significant association with pfmdr1 86Y mutant allele carriage at delivery and no significant difference was found for triple dhfr 51/59/108 mutations carriage in this sub-group compared to those infected only at delivery (P = 0.6), suggesting new infection parasite population after IPTp-SP and eventually AL treatments. In this regard, the lack of Plasmodium falciparum genotyping to distinguish recrudescent parasites to new infection parasites could be seen as another limit of this study. Future investigations on factors associated with pfmdr1 gene polymorphisms selection in pregnant women living in endemic countries should include large sample size and parasites population genotyping.
Conclusion
This study showed that the uptake of three or more IPTp-SP doses and one or more AL treatments are significantly associated with a reduced risk of pfmdr1 86Y mutant allele carriage in pregnant women at delivery. These findings suggest that both high coverage of IPTp-SP and the use of AL for the treatment of malaria infection/disease during pregnancy select for pfmdr1 N86 wild-type allele at delivery.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Abbreviations
- ANC-1:
-
First antenatal care
- MiP:
-
Malaria in pregnancy
- IPTp-SP:
-
Intermittent preventive treatment during pregnancy with sulfadoxine-pyrimethamine
- CSST:
-
Community-based scheduled screening and treatment of malaria during pregnancy
- ITN:
-
Insecticide treated net
- IQR:
-
Interquartile range
- LM:
-
Light microscopy
- OR:
-
Odds ratio
- AOR:
-
Adjusted odds ratio
References
Desai M, ter Kuile FO, Nosten F, McGready R, Asamoa K, Brabin B, et al. Epidemiology and burden of malaria in pregnancy. Lancet Infect Dis. 2007;7:93–104.
Moya-alvarez V, Abellana R, Cot M. Pregnancy-associated malaria and malaria in infants: an old problem with present consequences. Malar J. 2014;13:271.
Coulibaly SO, Gies S, D’Alessandro U. Malaria burden among pregnant women living in the rural district of Boromo, Burkina Faso. Am J Trop Med Hyg. 2007;77(6 Suppl):56–60.
Sangeeta V, Pushpalatha S. Severe maternal anemia and neonatal outcome. Sch J Appl Med Sci. 2014;2(IC):303–9.
WHO. A strategic framework for malaria prevention and control during pregnancy in the African Region. Brazzaville: WHO Regional Office for Africa; 2004.
WHO. Guidelines for the treatment of malaria. 2nd ed. Geneva: World Health Organization; 2010.
Finlayson K, Downe S. Why do women not use antenatal services in low- and middle-income countries? A meta-synthesis of qualitative studies. PLoS Med. 2013;10:e1001373.
Simkhada B, Teijlingen E. Factors affecting the utilization of antenatal care in developing countries: systematic review of the literature. J Adv Nurs. 2008;61:244–60.
Bulatao RA, Ross JA. Rating maternal and neonatal health services in developing countries. Bull World Health Organ. 2002;80:721–7.
Myer L, Harrison A. Why do women seek antenatal care late? Perspectives from rural South Africa. J Midwifery Womens Health. 2003;48:268–72.
Gies S, Coulibaly SO, Ouattara FT, Ky C, Brabin BJ, D’Alessandro U. A community effectiveness trial of strategies promoting intermittent preventive treatment with sulphadoxine-pyrimethamine in pregnant women in rural Burkina Faso. Malar J. 2008;7:180.
Valea I, Tinto H, Drabo MK, Huybregts L, Henry M, Roberfroid D, et al. Intermittent preventive treatment of malaria with sulphadoxine-pyrimethamine during pregnancy in Burkina Faso : effect of adding a third dose to the standard two-dose regimen on low birth weight, anaemia and pregnancy outcomes. Malar J. 2010;9:324.
van Eijk AM, Hill J, Alegana VA, Kirui V, Gething PW, ter Kuile FO, et al. Coverage of malaria protection in pregnant women in sub-Saharan Africa: a synthesis and analysis of national survey data. Lancet Infect Dis. 2011;11:190–207.
Maiga OM, Kayentao K, Traoré BT, Djimde A, Traoré B, Traoré M, et al. Superiority of 3 over 2 doses of intermittent preventive treatment with sulfadoxine-pyrimethamine for the prevention of malaria during pregnancy in mali: a randomized controlled trial. Clin Infect Dis. 2011a;53:215–23.
Harrington WE, Mutabingwa TK, Kabyemela E, Fried M, Duffy PE. Intermittent treatment to prevent pregnancy malaria does not confer benefit in an area of widespread drug resistance. Clin Infect Dis. 2011;53:224–30.
Harrington W, McGready R, Muehlenbachs A, Fried M, Nosten F, Duffy P. Intermittent preventive treatment in pregnancy with sulfadoxine-pyrimethamine: the times they are a-changin’. Clin Infect Dis. 2012;55:1025–6.
Tahita M, Tinto H, Erhart A, Kazienga A, Fitzhenry R, VanOvermeir C, et al. Prevalence of the dhfr and dhps mutations among pregnant women in rural Burkina Faso five years after the introduction of intermittent preventive treatment with sulfadoxine-pyrimethamine. PLoS ONE. 2015;10:e0137440.
Cisse M, Awandare G, Soulama A, Tinto H, Hayette M, Guiguemdé T. Recent uptake of intermittent preventive treatment during pregnancy with sulfadoxine–pyrimethamine is associated with increased prevalence of Pfdhfr mutations in Bobo-Dioulasso, Burkina Faso. Malar J. 2017;16:38.
Geiger C, Compaore G, Coulibaly B, Sie A, Dittmer M, Sanchez C, et al. Substantial increase in mutations in the genes pfdhfr and pfdhps puts sulphadoxine-pyrimethamine-based intermittent preventive treatment for malaria at risk in Burkina Faso. Trop Med Int Health. 2014;19:690–7.
Parikh S, Rosenthal PJ. Intermittent preventive therapy for malaria in pregnancy: is sulfadoxine-pyrimethamine the right drug? Clin Pharmacol Ther. 2010;87:160–2.
Desai M, Hill J, Fernandes S, Walker P, Pell C, Gutman J, et al. Prevention of malaria in pregnancy. Lancet Infect Dis. 2018;18:e119–32.
González R, Pons-Duran C, Piqueras M, Aponte JJ, ter Kuile FO, Menéndez C. Mefloquine for preventing malaria in pregnant women. Cochrane Database Syst Rev. 2018;3:CD011444.
Henry M, Florey L, Youll S, Gutman JR. An analysis of country adoption and implementation of the 2012 WHO recommendations for intermittent preventive treatment for pregnant women in sub-Saharan Africa 11 Medical and Health Sciences 1117 Public Health and Health Services. Malar J. 2018;17:364.
Awine T, Belko MM, Oduro AR, Oyakhirome S, Tagbor H, Chandramohan D, et al. The risk of malaria in Ghanaian infants born to women managed in pregnancy with intermittent screening and treatment for malaria or intermittent preventive treatment with sulfadoxine/pyrimethamine. Malar J. 2016;15:46.
WHO Malaria Policy Advisory Committee and Secretariat. Malaria policy advisory committee to the WHO : conclusions and recommendations of September 2013 meeting. Malar J. 2013;12:456.
WHO Malaria Policy Advisory Committee and Secretariat. Malaria policy advisory committee to the WHO: conclusions and recommendations of seventh biannual meeting (March 2015). Malar J. 2015;14:295.
D’Alessandro U, Hill J, Tarning J, Pell C, Webster J, Gutman J, et al. Treatment of uncomplicated and severe malaria during pregnancy. Lancet Infect Dis. 2018;18:e133–46.
WHO. Guidelines for the treatment of malaria. 3rd ed. Geneva: World Health Organization; 2015.
Nambozi M, Tinto H, Mwapasa V, Tagbor H, Kabuya JBB, Hachizovu S, et al. Artemisinin-based combination therapy during pregnancy: Outcome of pregnancy and infant mortality: A cohort study. Malar J. 2019;18:105.
Pickard AL, Wongsrichanalai C, Purfield A, Kamwendo D, Emery K, Zalewski C, et al. Resistance to antimalarials in Southeast Asia and genetic polymorphisms in pfmdr1. Antimicrob Agents Chemother. 2003;47:2418–23.
Mwai L, Kiara SM, Abdirahman A, Pole L, Rippert A, Diriye A, et al. In vitro activities of piperaquine, lumefantrine, and dihydroartemisinin in Kenyan Plasmodium falciparum isolates and polymorphisms in pfcrt and pfmdr1. Antimicrob Agents Chemother. 2009;53:5069–73.
Kavishe RA, Paulo P, Kaaya RD, Kalinga A, Van Zwetselaar M, Chilongola J, et al. Surveillance of artemether-lumefantrine associated Plasmodium falciparum multidrug resistance protein-1 gene polymorphisms in Tanzania. Malar J. 2014;13:264.
Brabin BJ. An analysis of malaria in pregnancy in Africa. Bull World Health Organ. 1983;61:1005–16.
LeClair M, Conrad M, Baliraine F, Al E. Optimization of a ligase detection reaction-fluorescent microsphere assay for characterization of resistance-mediating polymorphisms in African samples of Plasmodium falciparum. J Clin Microbiol. 2013;51:2564–70.
Scott S, Mens PF, Tinto H, Nahum A, Ruizendaal E, Pagnoni F, et al. Community-based scheduled screening and treatment of malaria in pregnancy for improved maternal and infant health in The Gambia, Burkina Faso and Benin: study protocol for a randomized controlled trial. Trials. 2014;15:340.
Ic COSM. Community-based malaria screening and treatment for pregnant women receiving standard intermittent preventive treatment with sulfadoxine-pyrimethamine: a multicentre (The Gambia, Burkina Faso and Benin) cluster randomised controlled trial. Clin Infect Dis. 2019;68:586–96.
Ruizendaal E, Tahita MC, Geskus RB, Versteeg I, Scott S, Alessandro U, et al. Increase in the prevalence of mutations associated with sulfadoxine – pyrimethamine resistance in Plasmodium falciparum isolates collected from early to late pregnancy in Nanoro. Burkina Faso Malar J. 2017;16:179.
Dokomajilar C, Nsobya SL, Greenhouse B, Rosenthal PJ, Dorsey G. Selection of Plasmodium falciparum pfmdr1 alleles following therapy with artemether-lumefantrine in an area of Uganda where malaria is highly endemic. Antimicrob Agents Chemother. 2006;50:1893–5.
Natama HM, Rovira-Vallbona E, Sorgho H, Somé MA, Traoré-Coulibaly M, Scott S, et al. Additional screening and treatment of malaria during pregnancy provides further protection against malaria and nonmalarial fevers during the first year of life. J Infect Dis. 2018;217:1967–76.
Asante KP, Owusu-Agyei S, Cairns M, Dodoo D, Boamah EA, Gyasi R, et al. Placental malaria and the risk of malaria in infants in a high malaria transmission area in Ghana: A prospective cohort study. J Infect Dis. 2013;208:1504–13.
Maiga OM, Kayentao K, Traoré BT, Djimde A, Traoré B, Traoré M, et al. Superiority of 3 over 2 doses of intermittent preventive treatment with sulfadoxine- pyrimethamine for the prevention of malaria during pregnancy in Mali: a randomized controlled trial. Clin Infect Dis. 2011b;53:215–23.
Kayentao K, Garner P, Van Eijk AM, Naidoo I, Roper C, Mulokozi A, et al. Intermittent preventive therapy for malaria during pregnancy using 2 vs 3 or more doses of sulfadoxine-pyrimethamine and risk of low birth weight in Africa: Systematic review and meta-analysis. JAMA. 2013;309:594–604.
Petersen I, Eastman R, Lanzer M. Drug-resistant malaria: molecular mechanisms and implications for public health. FEBS Lett. 2011;585:1551–62.
Conrad MD, Mota D, Foster M, Tukwasibwe S, Legac J, Tumwebaze P, et al. Impact of intermittent preventive treatment during pregnancy on Plasmodium falciparum drug resistance – mediating polymorphisms in Uganda. J Infect Dis. 2017;216:1008–17.
Holmgren G, Hamrin J, Svärd J, Mårtensson A, Gil JP, Björkman A. Selection of pfmdr1 mutations after amodiaquine monotherapy and amodiaquine plus artemisinin combination therapy in East Africa. Infect Genet Evol. 2007;7:562–9.
Tumwebaze P, Conrad MD, Walakira A, LeClair N, Byaruhanga O, Nakazibwe C, et al. Impact of antimalarial treatment and chemoprevention on the drug sensitivity of malaria parasites isolated from Ugandan children. Antimicrob Agents Chemother. 2015;59:3018–30.
Conrad MD, Leclair N, Arinaitwe E, Wanzira H, Kakuru A, Bigira V, et al. Comparative impacts over 5 years of artemisinin-based combination therapies on Plasmodium falciparum Polymorphisms that modulate drug sensitivity in Ugandan children. J Infect Dis. 2014;210:344–53.
Sisowath C, Stro J, Ma A, Msellem M, Obondo C, Bjo A, et al. In vivo selection of Plasmodium falciparum pfmdr1 86N coding alleles by artemether-lumefantrine (Coartem). J Infect Dis. 2005;191:1014–7.
Achieng AO, Muiruri P, Ingasia LA, Opot BH, Juma DW, Yeda R, et al. Temporal trends in prevalence of Plasmodium falciparum molecular markers selected for by artemether–lumefantrine treatment in pre-ACT and post-ACT parasites in western Kenya. Int J Parasitol Drugs Drug Resist. 2015;5:92–9.
Sondo P, Derra K, Diallo Nakanabo S, Tarnagda Z, Kazienga A, Zampa O, et al. Artesunate-amodiaquine and artemether-lumefantrine therapies and selection of Pfcrt and Pfmdr1 alleles in Nanoro. Burkina Faso PLoS One. 2016;11:e0151565.
Baraka V, Tinto H, Valea I, Fitzhenry R, Delgado-Ratto C, Mbonye MK, et al. In vivo selection of Plasmodium falciparum pfcrt and pfmdr1 variants by artemether-lumefantrine and dihydroartemisinin-piperaquine in Burkina Faso. Antimicrob Agents Chemother. 2015;59:734–7.
Somé AF, Séré YY, Dokomajilar C, Zongo I, Rouamba N, Greenhouse B, et al. Selection of known Plasmodium falciparum resistance-mediating polymorphisms by artemether-lumefantrine and amodiaquine-sulfadoxine-pyrimethamine but not dihydroartemisinin-piperaquine in Burkina Faso. Antimicrob Agents Chemother. 2010;54:1949–54.
Somé AF, Sorgho H, Zongo I, Bazié T, Nikiéma F, Sawadogo A, et al. Polymorphisms in K13, pfcrt, pfmdr1, pfdhfr, and pfdhps in parasites isolated from symptomatic malaria patients in Burkina Faso. Parasite. 2016;23:60.
Agbor L, Apinjoh T. Evidence of Plasmodium falciparum resistance to sulphadoxine-pyrimethamine (SP) in pregnant women along the slope of mount Cameroon. BMJ Glob Health. 2017;2:A1-67.
Bouyou-Akotet MK, Mawili-Mboumba DP, Kendjo E, Moutandou Chiesa S, Tshibola Mbuyi ML, Tsoumbou-Bakana G, et al. Decrease of microscopic Plasmodium falciparum infection prevalence during pregnancy following IPTp-SP implementation in urban cities of Gabon. Trans R Soc Trop Med Hyg. 2016;110:333–42.
Stepniewska K, White NJ. Pharmacokinetic determinants of the window of selection for antimalarial drug resistance. Antimicrob Agents Chemother. 2008;52:1589–96.
Kay K, Hastings IM. Measuring windows of selection for anti-malarial drug treatments. Malar J. 2015;14:292.
Baliraine FN, Rosenthal PJ. Prolonged selection of pfmdr1 polymorphisms after treatment of falciparum malaria with artemether-lumefantrine in Uganda. J Infect Dis. 2011;204:1120–4.
Mwesigwa J, Parikh S, McGee B, German P, Drysdale T, Kalyango JN, et al. Pharmacokinetics of artemether-lumefantrine and artesunate-amodiaquine in children in Kampala, Uganda. Antimicrob Agents Chemother. 2010;52:1589–96.
Acknowledgements
We express our sincere gratitude to the study participants. We are also grateful to nurses, field workers, field supervisors, data managers and laboratory workers at the Clinical Research Unit of Nanoro (CRUN) in Burkina Faso. Our special thank goes to Esmée Ruizendaal for kindly providing data on dhfr-dhps mutations. We acknowledge the research teams at the Malariology Unit at the Institute of Tropical Medicine (ITM), Belgium, and Centre d’Epidémiologie, Biostatistique et Recherche Clinique, Ecole de Santé Publique, Université Libre de Bruxelles (ULB) in Belgium.
Funding
The main study "COSMIC", was supported by European community’s Seventh Framework Programme under Grant Agreement No. 305662. The current analysis is funded by “Projets d’Opportunité” of “Académie de Recherche et d’Enseignement Supérieur” (ARES-POP), Belgium.
Author information
Authors and Affiliations
Contributions
HMN, RT, SS, HT and FKS conceived and designed the study. HMN and DLCB performed the genotyping experiments. MCT and HT coordinated the main COSMIC trial. HMN and DLCB analysed the data and interpreted the results with the input of RT, SS and FKS. MCT, HT and FSK provided reagents/materials/analysis tools. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The main trial protocol (COSMIC, NCT1941264) was reviewed and approved by the Institutional ethics committee at Centre Muraz, Bobo Dioulasso, Burkina Faso (reference number A20-2013/CE-CM). Written informed consent was obtained from all pregnant women.
Consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Natama, H.M., Toussaint, R., Bazié, D.L.C. et al. Prevalence and factors associated with carriage of Pfmdr1 polymorphisms among pregnant women receiving intermittent preventive treatment with sulfadoxine-pyrimethamine (IPTp-SP) and artemether-lumefantrine for malaria treatment in Burkina Faso. Malar J 19, 399 (2020). https://doi.org/10.1186/s12936-020-03473-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-020-03473-5