
Abstract
Background
Polysaccharides and poly-γ-glutamic acid (γ-PGA) are biomacromolecules that have been reported as bioflocculants, and they exhibit high flocculating activity in many industrial applications. Bacillus licheniformis CGMCC 2876 can produce polysaccharide and γ-PGA bioflocculants under different culture conditions. Several key genes are involved in the metabolic pathway of polysaccharides in B. licheniformis, but the impacts of the regulation of these genes on the production of polysaccharide bioflocculants have not been illustrated completely. To increase the bioflocculant production and identify the correlation between the synthesis of polysaccharides and γ-PGA in B. licheniformis, a few key genes were investigated to explore their influence on the synthesis of the bioflocculants.
Results
Overexpressing epsB from the eps gene cluster not only improved the bioflocculant crude yield by 13.98% but also enhanced the flocculating activity by 117.92%. The composition of the bioflocculant from the epsB recombinant strain was 28.95% total sugar, 3.464% protein and 44.03% γ-PGA, while in the original strain, these components represented 53.67%, 3.246% and 34.13%, respectively. In combination with an analysis of the transcriptional levels of several key genes involved in γ-PGA synthesis in B. licheniformis, we inferred that epsB played a key role in the synthesis of both polysaccharide and γ-PGA. The bioflocculant production of the epsB recombinant strain was further evaluated during batch fermentation in a 2 L fermenter; the flocculating activity reached 9612.75 U/mL, and the bioflocculant yield reached 10.26 g/L after 72 h, representing increases of 224% and 36.62%, respectively, compared with the original strain. Moreover, we found that the tandem expression of phosphoglucomutase (pgcA) and UTP-glucose-1-phosphate uridylyltransferase (gtaB1) could enhance the crude yield of the bioflocculant by 20.77% and that the overexpression of epsA could enhance the bioflocculant yield by 23.70% compared with the original strain.
Conclusions
This study provides a new method to greatly increase the bioflocculant production in B. licheniformis, and it demonstrates the correlation between the biosynthesis of polysaccharide and γ-PGA during EPS fermentation by regulating the expression of EpsB.
Similar content being viewed by others

Background
Polysaccharides are biomacromolecules that are produced by a wide variety of microorganisms, and they have recently attracted increasing attention because of their unique characteristics, such as their biodegradability, nontoxicity and negligible secondary pollution [1]. Polysaccharides have been reported as bioflocculants for many potential applications, such as for wastewater treatment and drinking water purification and in the food additive and fermentation industries [2, 3]. Although polysaccharide bioflocculants have many advantages, the low yield and high cost of their production as flocculants have largely limited their industrial applications [4].
Poly-γ-glutamic acid (γ-PGA) is a naturally occurring macromolecular polyamide that consists of d/l-glutamate monomers [5, 6]. γ-PGA exhibits many favourable features such as biodegradability, water solubility, edibility and nontoxicity to humans and the environment [7]. Like polysaccharides, γ-PGA can also be used as a bioflocculant in many applications, such as food, cosmetics and agriculture [5].
Bacillus licheniformis is a gram-positive, spore-forming soil bacterium that can secrete many kinds of biopolymers that can serve as antibiotics, biochemicals and consumer products, among other purposes [8, 9]. In our previous research, a bioflocculant-producing bacterium was isolated and identified as B. licheniformis [China General Microbiological Culture Collection Centre (CGMCC) 2876] [10], and it could produce polysaccharide and γ-PGA under different culture conditions. When using glucose as the sole carbon source, B. licheniformis mainly produces extracellular polysaccharides [11], whereas when using trisodium citrate and glycerol as the carbon sources, this strain mainly secretes γ-PGA [12]. Both polysaccharides and γ-PGA have shown high flocculating activities in industrial applications, so the ability to increase the flocculating activities of bioflocculants produced by B. licheniformis has huge commercial potential [13].
In this study, we mainly addressed bioflocculants that use glucose as the sole carbon source, and we used genetic engineering as a means to improve the flocculating activity and crude yield of the bioflocculants. In our previous research, we suggested the hypothetical metabolic pathways of polysaccharides in B. licheniformis and speculated on the key genes involved in polysaccharide synthesis; these genes are distributed in different locations in the genome and are responsible for the conversion of glucose into the subunits of polysaccharides [11]. Among the genes, crr encodes the protein component of glucose-specific phosphotransferase, whose function is to catalyse d-glucose to glucose-6-phosphate; pgcA encodes phosphoglucomutase, which is involved in converting glucose-6-phosphate to glucose-1-phosphate; then, glucose-1P is converted into UDP-glucose by UTP-glucose-1-phosphate uridylyltransferase, which is encoded by gtaB; further, epsA and epsB are two gene segments of the eps gene cluster, which encodes a series of glycosyltransferases that have different substrate specificities and are all important in the process of synthesizing polysaccharide repeating units. The length of the polysaccharide chain is determined by EpsA and EpsB, and then, the polysaccharides are exported under the action of EpsA [14,15,16]. These genes are all key genes in the main route for synthesizing EPS on the basis of the polysaccharide metabolic pathways that we have presented. Several studies have reported that regulating the expressions of genes can influence the polysaccharide production by the studied strains. In Staphylococcus aureus and Bacillus subtilis, the glycolipid production is related to the α-phosphoglucomutase PgcA, the α-glucose-1-phosphate uridyltransferase GtaB [17,18,19] and the glycosyltransferases YpfP and UgtP [20, 21]. The absence of genes pgcA, gtaB or ypfP prevents the strains from producing glycolipids [22]. Tang reported a linear relationship between the activity of phosphoglucomutase (PGM) and the production of EPS [23]. In Lactococcus lactis, overexpression of the phosphoglucomutase gene and UDP-glucose pyrophosphorylase gene can increase the production of UDP-monosaccharide [24], and in Streptococcus thermophilus LY03, the overexpression of these two genes can lead to a nearly 100% increase in the polysaccharide yield [25]. In Streptococcus pneumoniae, galU (glucose-1-phosphate uridylyltransferase gene) knockout mutants are unable to synthesize a detectable capsule polysaccharide [26]. Further, our previous research also indicated that the overexpression of UDP-glucose pyrophosphorylase gene gtaB2 in B. licheniformis could markedly increase both the yield of the polysaccharide bioflocculant and its flocculating activities [11]. In addition, many researchers have reported the importance of the eps gene cluster during polysaccharide synthesis, and it exhibits conspicuous homology across different strains [27, 28]. In Methylobacillus sp. strain 12S, EpsA regulates the transcription of methanolan synthesis genes, and the constitutive expression of epsA in strain 12S increases EPS production; further, researchers have also found that EpsB appears to act as a glucosyltransferase to catalyse the initial transfer of glucose moieties [29]. Alexander K. W. Elsholz reported that in Bacillus subtilis, the EPS synthesis depended on a tyrosine kinase that consisted of a membrane component (EpsA) and a kinase component (EpsB). Moreover, the tyrosine kinase-mediated self-regulation could control the exopolysaccharide production in bacteria [30, 31]. In summary, these genes are all important in the synthesis of EPS, but the impacts of regulating these genes on the production of polysaccharides in B. licheniformis have not been determined before. In this report, we focused on the effects of overexpressing single or tandem key genes on the synthesis of polysaccharide bioflocculants, and ultimately, we obtained a high-yield recombinant strain of B. licheniformis by overexpressing epsB.
Results and discussion
The construction of recombinant bacteria
Using electroporation followed by screening on tetracycline-resistant culture plates, several recombinant strains were obtained and called B. licheniformis-phY300-epsB, B. licheniformis-phY300-epsA, B. licheniformis-phY300-pgcA, B. licheniformis-phY300-pgcA-gtaB1, B. licheniformis-phY300-gtaB1, and B. licheniformis-phY300-crr.
A transcriptional analysis using qRT-PCR showed that the transcriptional levels of all of these key genes in the transformants were markedly increased compared with those of the original strain (Fig. 1).

qPCR analysis of key genes in wild-type strain and recombinant strains. a crr; b pgcA; c gtaB1; d epsA; e epsB; f tandem gene pgcA-gtaB1
Comparison between the original strain and recombinant B. licheniformis on the production of the bioflocculants
As shown in Fig. 2, the crude yield and flocculating activity of the polysaccharide bioflocculant produced by B. licheniformis were 7.51 g/L and 2340.52 U/mL, respectively, in the EPS medium for 56 h. The yield and flocculating activity of the bioflocculants produced by the recombinant strains B. licheniformis-phY300-pgcA, B. licheniformis-phY300-gtaB1 and B. licheniformis-phY300-crr were not improved compared with the original strain, demonstrating that solely overexpressing these three genes did not have an obvious impact on the polysaccharide bioflocculant production of B. licheniformis. In contrast, many reports have shown that the phosphoglucomutase gene (pgcA) and the UDP-glucose pyrophosphorylase gene (gtaB1) were key genes of the biosynthetic pathway of polysaccharide in Bacillus sp. [24, 25]. We also found that overexpressing these two genes simultaneously increased the bioflocculant crude yield to 9.07 g/L, which was 20.77% greater than that of the original strain. Thus, regulating the expression of genes pgcA and gtaB1 at the same time could promote the production of EPS in B. licheniformis. Moreover, the bioflocculant crude yield of B. licheniformis-phY300-epsA reached 9.29 g/L, which was an increase of 23.70% compared with that of the original strain. Remarkably, overexpressing epsB not only improved the bioflocculant crude yield by 13.98% to 8.56 g/L but also enhanced the flocculating activity by 117.92%, to 5100.50 U/mL. Thus, overexpressing epsB could dramatically improve the production of bioflocculants in B. licheniformis.

The comparison of original and recombinant B. licheniformis on flocculating activity and yield of bioflocculant
The composition of the bioflocculants produced by B. licheniformis-phY300-epsB
Scanning electron microscope (ZEISS SIGMA, USA) images of the bioflocculants produced by B. licheniformis-phY300-epsB are shown in Fig. 3. The purified bioflocculant was a white floccus solid after lyophilisation and did not exhibit obvious differences from the bioflocculant produced by the original strain. Meanwhile, the compositions of the bioflocculants produced by the original and epsB recombinant strains were markedly different. The total sugar content in the bioflocculant produced by B. licheniformis-phY300-epsB was 28.95%, the protein content was 3.464%, and the γ-PGA content reached 44.03%. In contrast, in the bioflocculant produced by the original strain, the content of total sugar was 53.67%, the protein content was 3.246%, and the γ-PGA content was 34.13% (Fig. 4). In our previous study, we found that EpsB contained the structural domain of the tyrosine kinase CpsD, which had a negative regulatory role on the formation of capsular polysaccharides in Streptococcus aureus [15] according to an analysis of the NCBI Conserved Domains. Therefore, we inferred that epsB had a negative regulatory effect on the synthesis of polysaccharides in B. licheniformis and thereby decreased the flocculating activity [11]. However, although the most recent results showed that overexpressing epsB indeed decreased the polysaccharide content synthesized by B. licheniformis, the flocculating activity of the bioflocculant was greatly improved compared with that of the original strain because overexpressing epsB produced a marked increase in the γ-PGA content. These results indicated that epsB could enhance the synthesis of γ-PGA during EPS fermentation. Moreover, we also found that the content of the other components in the bioflocculant produced by B. licheniformis-phY300-epsB increased from 8.95 to 23.56%. This result may have been another reason for the improvement in the flocculating activity, but the exact compositions of the active moieties need to be further investigated.

The SEM image of bioflocculant produced by B. licheniformis strains. a bioflocculant produced by epsB recombinant strain. Scale bar = 20 μm; b bioflocculant produced by epsB recombinant strain. Scale bar = 3 μm; c bioflocculant produced by original strain

The composition of bioflocculant produced by B. licheniformis. a wild-type strain; b epsB recombinant strain
The effect of overexpressing epsB on the biosynthetic pathway of γ-PGA
To determine the reason for the increase in the γ-PGA content of the polysaccharide bioflocculant when overexpressing epsB, the transcriptional levels of a few of the main genes involved in the synthesis of γ-PGA were assayed using quantitative real-time PCR; each sample was performed in triplicate (the related primers are listed in Additional file 1: Table S1). The results are shown in Fig. 5; the gene expression levels of pgsA, pgsB, icd, rocA and nrgB were increased in the epsB recombinant strain compared with the wild-type strain, while the expression levels of genes ccpA and ccpN were reduced in the epsB recombinant strain. Many studies have reported that pgsA and pgsB are synthetase genes that are involved in the biosynthesis of γ-PGA [32,33,34,35]; therefore, we inferred that overexpressing epsB could induce the transcription and improve the expression of γ-PGA synthetase in B. licheniformis, thereby resulting in an increase in the γ-PGA synthesis. The TCA cycle is an important pathway that is involved in glutamic acid biosynthesis, and isocitrate dehydrogenase, encoded by icd, is a rate-limiting enzyme in the process of acetyl-CoA conversion to α-ketoglutaric acid [32]. Moreover, arginine can be transformed to glutamic acid, an important pathway for the biosynthesis of glutamic acid, and the gene rocA is involved in this process [32, 36]. Thus, overexpressing epsB could also increase the expression of isocitrate dehydrogenase and facilitate the conversion of acetyl-CoA to α-ketoglutaric acid; meanwhile, overexpressing epsB could promote the transformation of arginine to glutamic acid. These processes would all eventually increase the γ-PGA synthesis. In contrast, in B. licheniformis, the carbon metabolism regulatory proteins CcpA and CcpN and the nitrogen metabolism regulatory protein NrgB played core roles in the transition between the metabolism of the polysaccharide and γ-PGA. CcpA and CcpN could co-enhance glycolysis and suppress the carbon flux in the TCA cycle, consequently slowing the synthesis of γ-PGA; simultaneously, CcpN could cut off the carbon flux from glycerol metabolism and further reduce the γ-PGA production [37]. The synthesis of γ-PGA was also influenced by NrgB, which could transform the major nitrogen metabolic flux between NH4 + and glutamate, facilitate ammonium utilization and promote glutamine synthesis; these effects are all beneficial for γ-PGA synthesis [37]. Our results also showed that overexpressing epsB could reduce the expression of CcpA and CcpN while increasing the expression of NrgB, all of which improved the γ-PGA synthesis (Fig. 6).

Comparison of the expression level of key genes involved in γ-PGA synthesis between wild-type and epsB recombinant strain

The γ-PGA and polysaccharide synthesis pathways in B. licheniformis
The fermentation production of epsB recombinant bacteria
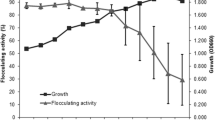
To evaluate the production of B. licheniformis-phY300-epsB in a more stable condition, batch fermentation was carried out in a 2 L fermenter. As shown in Fig. 7, the cells entered the stationary phase after 24 h of cultivation; at the same time, the glucose in the medium was used up, and then, the bacteria began to accumulate secondary metabolites. In addition, the flocculating activity of the fermentation broth began to increase rapidly until it peaked at 9612.75 U/mL after 60 h; this value was increased by 224% compared with the original strain under the same conditions and was 88.47% higher than the same strain cultured in the flask. Afterwards, the flocculating activity began to decline gradually. The final yield of the bioflocculant reached 10.26 g/L after alcohol precipitation and lyophilization; the yield was 36.62% higher than the original strain under the same conditions and 19.86% higher than the same strain cultured in the flask.

Bacillus licheniformis-phY300-epsB fermentation curve in 2 L fermenter
Conclusions
In this study, we investigated the influence of overexpressing several key genes involved in polysaccharide biosynthesis on the flocculating activity and crude yield of polysaccharide bioflocculants in B. licheniformis. The results showed that overexpressing epsA enhanced the yield of the polysaccharide bioflocculants by 23.70%, and overexpressing the tandem gene pgcA-gtaB1 enhanced the yield of the polysaccharide bioflocculants by 20.77% compared with that of the original strain, indicating that regulating the expression of epsA and pgcA-gtaB1 promoted polysaccharide biosynthesis in B. licheniformis. Notably, overexpressing epsB from the eps gene cluster not only improved the flocculating activity of the bioflocculants by 117.92% to 5100.50 U/mL, compared with the wild-type strain, but also enhanced the crude yield of the bioflocculant by 13.98% to 8.56 g/L. Subsequently, focusing on the epsB recombinant strain B. licheniformis-pHY300-epsB, by analysing the composition of the bioflocculant produced by B. licheniformis-phY300-epsB, we found that overexpressing epsB increased the content of γ-PGA and decreased the total sugar content in the bioflocculant. By comparing the transcriptional levels of a few key genes involved in the biosynthesis of γ-PGA in B. licheniformis-phY300-epsB and the wild-type strain, we found that the transcriptional levels of pgsA, pgsB, icd, rocA and nrgB increased to different degrees, whereas the transcriptional levels of ccpA and ccpN decreased. Thus, we could determine that epsB played a key role in the biosynthesis of both the polysaccharides and γ-PGA. Moreover, overexpressing epsB markedly enhanced the γ-PGA synthesis during EPS fermentation and consequently improved the bioflocculant production. During batch fermentation in a 2 L fermenter, the flocculating activity of B. licheniformis-phY300-epsB reached 9612.75 U/mL, and the bioflocculant yield reached 10.26 g/L after 72 h, increasing 224% and 36.62%, respectively, compared with those of the original strain. These results reveal major potential for industrial production.
In conclusion, this study provides a new method to greatly increase the bioflocculant production in B. licheniformis, and it demonstrates the correlation between the biosynthesis of polysaccharides and γ-PGA during EPS fermentation by regulating the expression of EpsB. All of these results provide meaningful biological information for further research.
Methods
Bacterial strain and culture medium
The bacterium B. licheniformis CGMCC 2876 was isolated in our laboratory and deposited at the China General Microbiological Culture Collection Center (Beijing, China). The LB medium contained the following components (g/L): tryptone, 10; yeast extract, 5; and NaCl, 10, at pH 7.2. The preculture medium contained the following components (g/L): glucose, 10; urea, 0.5; MgSO4·7H2O, 0.2; KH2PO4, 0.1; K2HPO4, 0.1; NaCl, 0.1; and yeast extract, 0.5. The EPS medium consisted of the following components (g/L): glucose, 13.9; urea, 2.67; MgSO4, 0.048; KH2PO4, 5.6; K2HPO4, 1.4; NaCl, 2; and yeast extract, 0.6. For polysaccharide production, the initial pH of each medium was 7.2. The strain was first cultured at 200 rpm for 17 h in a 250 mL Erlenmeyer flask containing 50 mL of preculture medium at 37 °C. Subsequently, 4% of the seed culture (v/v) was inoculated at 200 rpm for 56 h in a 250 mL Erlenmeyer flask with 50 mL of EPS medium at 37 °C for polysaccharide production.
Gene overexpression in B. licheniformis
The key genes were amplified using their corresponding primers (Table 1), and they were then cloned into the E. coli–Bacillus shuttle vector pHY300PLK-PamyL-TTamyL, which was constructed in our previous study. The recombinant expression vectors for genes epsB, epsA, pgcA, gtaB1 and crr were designated as pHY300-epsB, pHY300-epsA, pHY300-pgcA, pHY300-gtaB1, and pHY300-crr, respectively. The tandem gene fragments pgcA and gtaB1 were amplified using the primers pgcA-S and pgcA-A and gtaB1-S and gtaB1-A, respectively (Table 1), and the PCR products were spliced using a pEASY-Uni Seamless Cloning and Assembly Kit (TransGen Biotech, China). Using the assembled fragment as a template, the tandem gene pgcA-gtaB1 was amplified using the pgcA forward primer and gtaB1 reverse primer. Then, the tandem gene was cloned into pHY300PLK-PamyL-TTamyL by restriction digestion using KpnI and SpeI. These recombinant expression vectors were separately transformed into B. licheniformis cells by electroporation, and the transformants were screened on tetracycline-resistant culture plates.
Quantitative real-time PCR
The total RNA was extracted from B. licheniformis when the OD600 reached 3 by using the MiniBEST Universal RNA Extraction Kit (Takara, Japan).The RNA was quantified using a NanoDrop 2000 spectrophotometer (Thermo, USA). Using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, USA) for reverse transcription, the obtained cDNA was diluted to 100 ng/μL for further real-time PCR analyses performed on the stepOne Real-Time PCR System (Applied Biosystems, USA). The RT-PCR reaction was performed using Transtart Top Green qPCR Supermix Kit (TransGen Biotech, China) in a 20 μL mixture. Quantitative real-time PCR was performed in triplicate for each sample, and the relevant primers are listed in Table 2.
Purification of the biopolymer and determination of the flocculating activity
After 56 h of fermentation, the culture broths were centrifuged at 9000 rpm for 15 min to remove the cells. Three volumes of ethanol were added to the supernatant to precipitate the crude products. Then, the crude products were dissolved using distilled water and lyophilized to obtain the purified products.
The flocculating activity (FA) was measured according to the method described by Kurane et al. [38]. First, 40 mL of 1% (wt/vol) kaolin and 2.5 mL of CaCl2 solution (10 g/L) were mixed thoroughly with 1 mL of the sample and then incubated for 5 min at room temperature. By measuring the decrease in turbidity (OD550) of the upper phase, the FA was calculated using the following equation: FA (U/mL) = (A − B)/A × 100 × D per mL, where A and B are the optical densities of the control and sample at 550 nm, respectively, and D is the dilution factor of the cell-free culture broth. Each sample was measured in triplicate.
Analysis of the composition of the polysaccharide bioflocculant
The phenol–sulfuric acid method [39] was used to determine the total sugar content in the bioflocculant by measuring the optical densities of the samples at 490 nm. The protein content was measured using the Bradford method with the Bio-Rad Protein Assay Kit (Bio-Rad, USA) at OD595. The γ-PGA content in the polysaccharide bioflocculant was determined by HPLC with an Agilent 1200 HPLC system and an Agilent HC-C18 column (25 cm × 4.6 mm). The purified products were dissolved in 6 M HCl to hydrolyse the γ-PGA. The mixtures were maintained at 105 °C for 24 h and then neutralized and metered volumetrically. Afterwards, the samples were characterized using HPLC for qualitative and quantitative analysis. The operating conditions were as follows: 0.1 mol/L KH2PO4 (containing 5% methanol and with the pH adjusted to 2.5 using phosphoric acid) was used as the mobile phase at a flow rate of 1.0 mL/min; the injected volume was 20 μL, the column temperature was maintained at 30 °C, and the UV detection wave length was 210 nm. Pure sodium glutamate was used as the standard compound.
Production of the polysaccharide bioflocculant from a batch culture in a 2 L fermenter
To cultivate larger volumes of the recombinant strain, 56 mL seed cultures of B. licheniformis-pHY300-epsB were cultivated in a seed medium in 250 mL shaker flasks for 17 h and then transferred into a 2 L fermenter system containing 1.4 L of the EPS medium. The pH, sterile air rate and the dissolved oxygen (DO) were maintained at 7.2, 2 vvm and 30%, respectively. The fermentation was carried out at 37 °C for 72 h to determine the final bioflocculant crude yield and flocculating activity.
Abbreviations
- EPS:
-
extracellular polysaccharides
- γ-PGA:
-
poly-gamma-glutamic acid
- TCA:
-
tricarboxylic acid cycle
- UTP:
-
uridine triphosphate
- UDP:
-
uridine diphosphate
- HPLC:
-
high-performance liquid chromatography
References
Rasulov BA, Pattaeva MA, Yili A, Aisa HA. Polysaccharide-based bioflocculant template of a diazotrophic Bradyrhizobium japonicum 36 for controlled assembly of AgCl nanoparticles. Int J Biol Macromol. 2016;89:682–8.
Sheng GP, Yu HQ, Li XY. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: a review. Biotechnol Adv. 2010;28:882–94.
Subramanian SB, Yan S, Tyagi RD, Surampalli RY, Lohani BN. Isolation and molecular identification of extracellular polymeric substances (EPS) producing bacterial strains for sludge settling and dewatering. J Environ Sci Health A Tox Hazard Subst Environ Eng. 2008;43:1495–503.
Salehizadeh H, Yan N. Recent advances in extracellular biopolymer flocculants. Biotechnol Adv. 2014;32:1506–22.
Shih I-L, Van Y-T. The production of poly-(γ-glutamic acid) from microorganisms and its various applications. Bioresour Technol. 2001;79:207–25.
Ashiuchi M, Misono H. Biochemistry and molecular genetics of poly-γ-glutamate synthesis. Appl Microbiol Biotechnol. 2002;59:9–14.
Feng J, Gu Y, Quan Y, Cao M, Gao W, Zhang W, Wang S, Yang C, Song C. Improved poly-gamma-glutamic acid production in Bacillus amyloliquefaciens by modular pathway engineering. Metab Eng. 2015;32:106–15.
Schallmey M, Singh A, Ward OP. Developments in the use of Bacillus species for industrial production. Can J Microbiol. 2004;50:1–17.
Nithya V, Murthy PSK, Halami PM. Development and application of active films for food packaging using antibacterial peptide of Bacillus licheniformis Me1. J Appl Microbiol. 2013;115:475–83.
Xiong Y, Wang Y, Yu Y, Li Q, Wang H, Chen R, He N. Production and characterization of a novel bioflocculant from Bacillus licheniformis. Appl Environ Microbiol. 2010;76:2778–82.
Chen Z, Liu P, Li Z, Yu W, Wang Z, Yao H, Wang Y, Li Q, Deng X, He N. Identification of key genes involved in polysaccharide bioflocculant synthesis in Bacillus licheniformis. Biotechnol Bioeng. 2017;114:645–55.
Yan S, Yao H, Chen Z, Zeng S, Xi X, Wang Y, He N, Li Q. Poly-γ-glutamic acid produced from Bacillus licheniformis CGMCC 2876 as a potential substitute for polyacrylamide in the sugarcane industry. Biotechnol Prog. 2015;31:1287–94.
Zhuang X, Wang Y, Li Q, Yan S, He N. The production of bioflocculants by Bacillus licheniformis using molasses and its application in the sugarcane industry. Biotechnol Bioprocess Eng. 2012;17:1041–7.
Stingele F, Vincent SJF, Faber EJ, Newell JW, Kamerling JP, Neeser J-R. Introduction of the exopolysaccharide gene cluster from Streptococcus thermophilus Sfi6 into Lactococcus lactis MG1363: production and characterization of an altered polysaccharide. Mol Microbiol. 1999;32:1287–95.
Morona JK, Morona R, Paton JC. Comparative genetics of capsular polysaccharide biosynthesis in Streptococcus pneumoniae types belonging to serogroup 19. J Bacteriol. 1999;181:5355–64.
Sau S, Bhasin N, Wann ER, Lee JC, Foster TJ, Lee CY. The Staphylococcus aureus allelic genetic loci for serotype 5 and 8 capsule expression contain the type-specific genes flanked by common genes. Microbiology. 1997;143:2395–405.
Gründling A, Schneewind O. Genes required for glycolipid synthesis and lipoteichoic acid anchoring in Staphylococcus aureus. J Bacteriol. 2007;189:2521–30.
Lazarevic V, Soldo B, Médico N, Pooley H, Bron S, Karamata D. Bacillus subtilis α-phosphoglucomutase is required for normal cell morphology and biofilm formation. Appl Environ Microbiol. 2005;71:39–45.
Lu M, Kleckner N. Molecular cloning and characterization of the pgm gene encoding phosphoglucomutase of Escherichia coli. J Bacteriol. 1994;176:5847–51.
Jorasch P, Wolter FP, Zähringer U, Heinz E. A UDP glucosyltransferase from Bacillus subtilis successively transfers up to four glucose residues to 1,2-diacylglycerol: expression of ypfP in Escherichia coli and structural analysis of its reaction products. Mol Microbiol. 1998;29:419–30.
Kiriukhin MY, Debabov DV, Shinabarger DL, Neuhaus FC. Biosynthesis of the glycolipid anchor in lipoteichoic acid of Staphylococcus aureus RN4220: role of YpfP, the diglucosyldiacylglycerol synthase. J Bacteriol. 2001;183:3506–14.
Reichmann NT, Gründling A. Location, synthesis and function of glycolipids and polyglycerolphosphate lipoteichoic acid in Gram-positive bacteria of the phylum Firmicutes. FEMS Microbiol Lett. 2011;319:97–105.
Tang Y-J, Zhong J-J. Exopolysaccharide biosynthesis and related enzyme activities of the medicinal fungus, Ganoderma lucidum, grown on lactose in a bioreactor. Biotechnol Lett. 2002;24:1023–6.
Kleerebezem M, van Kranenburg R, Tuinier R, Boels IC, Zoon P, Looijesteijn E, Hugenholtz J, de Vos WM. Exopolysaccharides produced by Lactococcus lactis: from genetic engineering to improved rheological properties? In: Konings WN, Kuipers OP, In’t Veld JHJH, editors. Lactic acid bacteria: genetics, metabolism and applications: proceedings of the sixth symposium on lactic acid bacteria: genetics, metabolism and applications, 19–23 September 1999, Veldhoven, The Netherlands. Dordrecht: Springer; 1999. p. 357–65.
Levander F, Svensson M, Rådström P. Enhanced exopolysaccharide production by metabolic engineering of Streptococcus thermophilus. Appl Environ Microbiol. 2002;68:784–90.
Mollerach M, López R, García E. Characterization of the galU gene of Streptococcus pneumoniae encoding a uridine diphosphoglucose pyrophosphorylase: a gene essential for capsular polysaccharide biosynthesis. J Exp Med. 1998;188:2047–56.
Veith B, Herzberg C, Steckel S, Feesche J, Maurer KH, Ehrenreich P, Bäumer S, Henne A, Liesegang H, Merkl R. The complete genome sequence of Bacillus licheniformis DSM13, an organism with great industrial potential. J Mol Microbiol Biotechnol. 2004;7:204–11.
Yan S, Wang N, Chen Z, Wang Y, He N, Peng Y, Li Q, Deng X. Genes encoding the production of extracellular polysaccharide bioflocculant are clustered on a 30-kb DNA segment in Bacillus licheniformis. Funct Integr Genom. 2013;13:425–34.
Yoshida T, Ayabe Y, Yasunaga M, Usami Y, Habe H, Nojiri H, Omori T. Genes involved in the synthesis of the exopolysaccharide methanolan by the obligate methylotroph Methylobacillus sp. strain 12S. Microbiology. 2003;149:431–44.
Elsholz AK, Wacker SA, Losick R. Self-regulation of exopolysaccharide production in Bacillus subtilis by a tyrosine kinase. Genes Dev. 2014;28:1710–20.
Gerwig J, Kiley TB, Gunka K, Stanley-Wall N, Stülke J. The protein tyrosine kinases EpsB and PtkA differentially affect biofilm formation in Bacillus subtilis. Microbiology. 2014;160:682–91.
Wang J, Yuan H, Wei X, Chen J, Chen S. Enhancement of poly-γ-glutamic acid production by alkaline pH stress treatment in Bacillus licheniformis WX-02. J Chem Technol Biotechnol. 2016;91:2399–403.
Ashiuchi M. Microbial production and chemical transformation of poly-γ-glutamate. Microb Biotechnol. 2013;6:664.
Kimura K, Tran LS, Do TH, Itoh Y. Expression of the pgsB encoding the poly-gamma-dl-glutamate synthetase of Bacillus subtilis (natto). Biosci Biotechnol Biochem. 2009;73:1149–55.
Sung M-H, Park C, Kim C-J, Poo H, Soda K, Ashiuchi M. Natural and edible biopolymer poly-γ-glutamic acid: synthesis, production, and applications. Chem Rec. 2005;5:352–66.
Yu W, Chen Z, Shen L, Wang Y, Li Q, Yan S, Zhong C-J, He N. Proteomic profiling of Bacillus licheniformis reveals a stress response mechanism in the synthesis of extracellular polymeric flocculants. Biotechnol Bioeng. 2016;113:797–806.
Yu W, Chen Z, Ye H, Liu P, Li Z, Wang Y, Li Q, Yan S, Zhong CJ, He N. Effect of glucose on poly-γ-glutamic acid metabolism in Bacillus licheniformis. Microb Cell Fact. 2017;16:22.
Kurane R, Hatamochi K, Kakuno T, Kiyohara M, Hirano M, Taniguchi Y. Production of a bioflocculant by Rhodococcus erythropolis S-1 grown on alcohols. Biosci Biotechnol Biochem. 1994;58:428–9.
Yokoi H, Yoshida T, Mori S, Hirose J, Hayashi S, Takasaki Y. Biopolymer flocculant produced by an Enterobacter sp. Biotechnol Lett. 1997;19:569–73.
Authors’ contributions
PZL, ZC, LJY, QBL and NH designed the experiments. PZL and ZC preformed the experiments. PZL, ZC, LJY and NH analyzed the results. PZL wrote the manuscript which was reviewed and approved by all authors. All authors read and approved the final manuscript.
Acknowledgements
This work was financially supported by the National Natural Science Foundation of China (51378444, 21676221), the University of Science and Technology in Fujian Province in the cooperative major project (2015H6004) and Xiamen Southern Oceanographic Center (15GYY024NF03). We also gratefully acknowledge the Analysis and Testing Center, School of Life Sciences, Xiamen University and the Fujian Provincial Scientific and AJE for language polishing work.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data generated or analysed during this study are included in this published article (and in Additional file 1).
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
National Natural Science Foundation of China, Award Number: 51378444, Recipient: Ning He Ph.D.
National Natural Science Foundation of China, Award Number: 21676221, Recipient: Ning He Ph.D.
University of Science and Technology in Fujian Province in the cooperative major project, Award Number: 2015H6004, Recipient: Ning He Ph.D.
Xiamen Southern Oceanographic Center, Award Number: 15GYY024NF03, Recipient: Ning He Ph.D.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional file
12934_2017_775_MOESM1_ESM.docx
Additional file 1: Table S1. Provides the related primers used in the Fig. 5.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Liu, P., Chen, Z., Yang, L. et al. Increasing the bioflocculant production and identifying the effect of overexpressing epsB on the synthesis of polysaccharide and γ-PGA in Bacillus licheniformis . Microb Cell Fact 16, 163 (2017). https://doi.org/10.1186/s12934-017-0775-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-017-0775-9