
Abstract
Background
Microencapsulation technology is the fundamental delivery system for encapsulating the natural bioactive compounds especially phenolic in order to developing bioavailability, stability and controlling release. This study was conducted to determine the antibacterial and health-promoting potential of the phenolic rich extract (PRE)-loaded microcapsules obtained from Polygonum bistorta root as a dietary phytobiotic in mice challenged by enteropathogenic Escherichia coli (E. coli).
Method
The PRE was obtained from Polygonum bistorta root using fractionation by different polarity solvents and the highest PRE was encapsulated by the combination of modified starch, maltodextrin, and whey protein concentrate as wall materials using a spray dryer. Then, the physicochemical characterization (particle size, zeta potential, Morphology and polydispersity index) of microcapsules have been assessed. For the invivo study, 30 mice at five treatment were designed and antibacterial properties were analyzed. Furthermore, relative fold changes in the ileum population of E. coli was investigated using Real time PCR.
Results
The encapsulation of PRE resulted in the production of phenolic enriched extract-loaded microcapsules (PRE-LM) with a mean diameter of 330 nm and relatively high entrapment efficiency (87.2% w/v). The dietary supplementation of PRE-LM improved weight gain, liver enzymes, gene expression, morphometric characteristics of the ileum and decreased the population of E. coli present in the ileum significantly (p < 0.05).
Conclusion
Our funding suggested PRE-LM as a promising phytobiotic against E. coli infection in mice.
Similar content being viewed by others
Background
Nowadays polyphenols have been applied as a natural phytobiotic and antioxidant supplement to fight enteropathogens and improve gut health and function [1] Phenolic compounds are universally distributed in plants and phenolics such as gallic acid, syringic acid, ferulic acid, catechin, ellagic acid, naringin, and chrysin are known for their antimicrobial and antioxidant activities [2]. Phenolic compounds' biological activity are also greatly affected by their interactions with other biomolecules. For instance, the solubility and bioactivity of tannins may also be altered by the presence of various molecules, including polysaccharides and proteins. The strong interactions with other biomolecules are likely to interfere with the bioavailability and biological activities of phenolic compounds [3].
Polygonum bistorta L. (Polygonacea family) is known as Bistort or Snakeroot, which is perennial herbaceous flowering with a slender stem distributed in Europe, Asia, North America, and India. It is a popular medicinal plant in many traditional medicines for its medicinal properties such as controlling bleeding, astringent, diuretic, and expectorant. Among the different aerial parts, the root has been widely used for treating dysentery, sepsis, exogenous heat, cough, and microbial infections [4, 5]. The former investigation has confirmed the main bioactive compound constituents in this plant [5, 6]. A study was performed by Pirvu et al. [7] indicated the presence of caffeic acid, gallic acid [8], ellagic acid, and ferulic acid [9] as the main phenolic compounds, and rutin [10], hyperine, quercitrin, isoquercitrin [11], miquelianin [9] as the important flavonol glycosides identified in this herbal medicine [7]. These natural compounds are known for their antimicrobial activities. Similar research elucidated the appreciable antimicrobial potential of various parts of this plant against pathogenic bacteria such as Paenibacillus larvae, Melissococcus plutonius and Bacillus subtilis [12]. A recent experiment has been done by Pawłowska et al. [13] indicated that the Flavon-3-ols and galloylglucose derivatives as natural products possess antimicrobial activity against skin pathogens [13]. While the various type of bioactive compounds especially phenolic compounds have been found in the root of Polygonum bistorta L. therefore, it could be an interesting source for natural phytochemicals with health-promoting properties in the medicinal, cosmeceuticals, nutraceuticals, and pharmaceutical industries. Despite the various biological activities of the phenolic compounds for the treatment of diseases, their less bioavailability, low absorption, and sensitivity to oxidation are the main problems of these compounds [14, 15]. Therefore, the encapsulation of these phytochemicals is suggested to overcome these limitations facilitate their release, and enhance their bioavailability [16].
The encapsulation technology has great attention and interest in the delivery of bioactive compounds as this technology could help in maintaining all features of bioactive compounds such as potent and strong protection. This technology can be categorized into three parts: physical methods like spray drying, a chemical procedure like molecular inclusion, and a physicochemical process such as liposome encapsulation [17,18,19]. Among all described techniques spray drying is widely performed for encapsulation of bioactive compounds. In this technology, different types of carriers are commonly used including starch, maltodextrin, gum, and whey protein concentrate. In the present study, we attempted to encapsulate the phenolic-rich extract obtained from Polygonum bistorta L. root and to determine its antioxidant, and antibacterial activities in the mouse model of enteropathogenic Escherichia coli infection.
Materials and method
Materials
The dry roots of Polygonum bistorta were purchased from the herbal market of Shirvan, North Khorasan, IRAN. The voucher spiceman was deposited in islamic azad university herbarium by 10,536 IAUM code number. For encapsulation, the wall material such as modified starch (MS) (HI-CAP® 100, Ingredion, Humberg, Germany) and maltodextrin (MD) (DE = 18–20; Foodchem, China) and whey protein concentrate 80% (w/w) (WPC) (Hilmar, CA, USA) was purchased. The foodborne pathogen Escherichia coli (E. coli O157: H7) was purchased from the microbial culture collection of the Razi Vaccine and Serum Research Institute, Karaj, Iran. The RNeasy mini kit, QIAamp DNA Stool Mini Kit was obtained from Qiagen Germany. The cDNA synthesis kit and SYBER green were bought from Biofact, Korea. All the other solvents and chemicals not mentioned here were purchased from Merck, Germany.
Obtaining phenolic-enriched extract
The root part of the plant was ground to the fine powder using laboratory grinder. For obtaining of phenolic-enriched extract, 100 g of dried root powder was added into the 2L round bottom flask and 1000 mL of methanol (80% v/v), and 50 mL of hydrochloric acid (6 M) were added. The mixture was refluxed (90 °C, 2 h) [20]. The obtained extract was filtered and the solvent was evaporated (60 °C) by using a rotary evaporator (Buchi, Flawil, Switzerland) [21]. To obtain the phenolic-enriched extract (PRE) the dried extract was re-extracted with different polarity solvents including hexane, chloroform, ethyl acetate, n-butanol, and water, respectively. For extract partitioning, 250 mL from each solvent was used and re-extraction was performed twice in a separating funnel. Finally, the obtained extract was filtered and concentrated using a rotary evaporator (Buchi. Switzerland). The total phenolic content of each extract was determined colorimetrically using a visible spectrophotometer (Novaspec II Visiblespectro, Japan) at 765 by Folin-ciocalteu assay [21], and results were reported as mg gallic acid equivalent (GA eq.) /g dry crude extract. The extract with the highest concentration of phenolic compounds is called phenolic-enriched extract (PRE) and is used for further experiments.
Encapsulation process
A pilot plant scale spray dryer (Buchi, B-191, Switzerland) with counter-current airflow was utilized to encapsulate the PRE obtained from P. bistorta root [22]. The maltodextrin (70% w/w), modified starch (20% w/w), and whey protein concentrate (10% w/w) was used as wall materials according to our early studies [23,24,25]. Briefly, the PRE (8 g), tween 80 (3 g), and the 32 g of wall material with the respective ratio of 1:4 were added to 100 ml of water and mixed using a stirrer at 4 °C overnight for complete rehydration. The solution was homogenized using Ultra Turrax homogenizer (IKA, Germany) before spry drying at 10,000 rpm for 5 min. The temperature of the spray dryer was fixed between the range of 130 °C (inlet temperature) and 80 °C (outlet temperature), the pressure was 0.4 kg/cm2 and the flow rate was adjusted to 8 mL/min. The dried powder obtained is called PRE-loaded microcapsules (PRE-LM) and it was used for further investigations.
Characterization of microcapsules
The particle size, zeta potential, and polydispersity index (PDI) of the capsules containing PRE of P. bistorta root were tested by Zetasizer (Malvern Instruments Ltd., UK). The morphology evaluation of microcapsules was carried out by Scanning Electron Microscopy (SEM). The total phenolic compounds in the capsules were determined using the Folin–Ciocalteau assay [21].
Entrapment efficiency
The entrapment efficiency of PRE-LM was determined as described earlier with slight modifications [26, 27]. The dried microcapsules (100 mg) were dispersed in 1.5 ml of 80% (v/v) methanol and centrifuged (12,000 rpm, 5 min). The total phenolic compounds were determined in the supernatant by Folin–Ciocalteau assay. The pellet (precipitant) was re-extracted using 1.5 ml of 80% (v/v) methanol using a sonicator 60 kHz for the 30 s. The total phenolic compounds were determined in the extract obtained from the pellet. The experiment was repeated three times. Finally, the entrapment efficiency (EE) was calculated using the below formula.
HPLC analysis of phenolic compounds
The profiling of natural phenolic compounds present in the PRE-LM was determined using high-performance liquid chromatography (HPLC) with a UV–Vis photodiode array (DAD) detector (Agilent-1200 series). The phenolic compounds present in the PRE-LM were extracted using 80% (v/v) methanol applying sonication for the complete extraction of phenolic compounds from microcapsules. The extract was used for HPLC analysis. The C18 analytical column (Intersil ODS-3 5um 4.6 × 150 mm Gl Science Inc. USA) was applied for identification of phenolic compounds. It equilibrated with 85% of deionized water as solvent A and 15% of acetonitrile as solvent B. Then the ratio of solvent B was enhanced to 85% in 50 min followed by decreasing solvent B to 15% in 55 min. This ratio was kept to 50 min for the next analysis. Gallic acid, syringic acid, vanillic acid, salicylic acid, caffeic acid, pyrogallol, catechin, cinnamic acid, ellagic acid, naringin, chrysin, and ferulic acid were the bioactive phenolic standards used in this research [28].
Antioxidant and antibacterial activities
The antioxidant potential of the PRE-LM was evaluated using DPPH radical scavenging activity. Vitamin C was used as antioxidant standard [29]. The antibacterial properties of PRE-LM were determined against E. coli (O157: H7) and data was reported as Minimum Inhibitory Concentration (MIC) using microdilution method and the positive control was oxytetracycline [30]. All the experiments mentioned here were run in triplicates.
Animal trial
The 30 Balb/c male mice with a respective weight range of 28–30 g were provided by the Razi Vaccine and Serum Research Institute of Mashhad, Mashhad, Iran. To acclimatize the animal, they were kept in cages for 10 days (12 h light /dark cycle) and received food and water. After that, they were randomly divided into five groups as described in Table 1. All mice received experimental treatments for 4 weeks and the oral infection was performed through a gavage needle using 108 CFU of E. coli once on day 21. Animals were monitored daily for general health and the amount of food eaten. At the end of the experiment (Day 28), the mice were euthanized with pentobarbital-HCL (50 mg/kg, i.p.) and sacrificed. The blood, liver, and ileum samples were collected immediately and used for liver enzyme analysis, lipid peroxidation assay, gene expression analysis, and morphometric evaluation of the ileum.
Liver enzymes and lipid peroxidation assay
The biochemical assay of serum for liver enzymes including alkaline phosphatase (ALP), alanine aminotransferase (ALT), and aspartate aminotransferase (AST) was carried out on an automated chemistry analyzer (Roche, Hitachi 902 analyzer, Japan). The lipid peroxidation of the liver for each treatment was evaluated using the determination of malondialdehyde (MDA) content in the liver tissue lysate by thiobarbituric acid reactive substances as described earlier [31].
Gene expression analysis
The ileum tissues were separated and kept in liquid nitrogen at the end of in vivo study. Then, the RNA was extracted using the RNeasy Mini kit (Qiagen, Hilden, Germany) and immediately proceed to cDNA synthesis using a cDNA synthesis kit (Biofact Korea). The real-time PCR program was adjusted at 95 °C for 5 min (1X), 95 °C for 25 s, then 58 °C for the 30 s, and 72 °C for 30 s (35X). Finally, the expressions of claudin, occludin, and mucin as major biomarker genes in nutrient absorption and defense system in the ileum were determined. The expressions of genes were normalized to β-actin as a reference gene [32]. The list of primers used in the current study are Claudin-1 (F: tccttgctgaatctgaaca; R: agccatccacatcttctg); Claudin-2 (F: gtcatcgcccatcagaagat; R: actgttggacagggaaccag); Occludin (F: actcctccaatggacaagtg; R: ccccacctgtcgtgtagtct); Mucin-2 (F: gatggcacctacctcgttgt; R: gtcctggcacttgttggaat) and Beta-actin (F: gctgagagggaaatcgtgcgtg; R: ccagggaggaagaggatgcgg) [33, 34].
Quantification of E. coli in the ileum digesta
The major sites of microbial fermentation, propagation, and colonization of enteropathogens in the monogastric are the ileum, which is why in this study the population of E. coli was analyzed only in the ileum section. The entire ileum digesta was taken and mixed and 100 mg of the digesta was used for the DNA extraction using QIAamp DNA Stool Mini Kit (Germany). The quantitative real-time PCR was applied using the SYBR Green PCR Master Mix (Biofact, Korea). The fold changes for the E. coli in the ileum digesta was determined using the 2−∆∆Ct method as described earlier [35, 36]. The primer used in this study are study are E. coli (O157:H7) ( F: ttaccagcgataccaagagc; R: caacatgaccgatgacaagg) [37] and Total bacteria (F: cggcaacgagcgcaaccc; R: ccattgtagcacg tgtgtagcc) [38].
Statistical analysis
The ANOVA test, using the GLM procedure of SAS (Ver. 9.1) was considered to analyze the obtained results. The significance was confirmed by Duncan's multiple range test. The p-value of < 0.05 was used as the criterion for a statistically significant difference. All experiments were done in triplicate and the results are presented as mean values ± standard deviation (mean ± SD) or mean values + standard error of the means (SEM).
Results and discussion
Extract partitioning
The application of solvents with different polarities for the partitioning of phenolic compounds from P. bistorta root produced different partitions with various amounts of phenolic compounds. The highest amount of phenolic content among partitions was observed in ethyl acetate extract with respective values of 256.7 ± 3.67 mg/g GAE followed by n-butanol (61.2 ± 5.03) > water (34.1 ± 4.27) > chloroform (21.7 ± 3.63) > hexane (18.7 ± 4.82) mg/g dried extract, respectively. Based on these results, the ethyl acetate extract was used for further evaluations as a phenolic enriched extract (PRE). These results indicated that the polarity of solvents could influence the extraction of phenolic compounds. The earlier publications reported the phenolic compounds as moderately polar compounds hence they tended to accumulate in the medium polar solvent such as ethyl acetate. The results obtained in this experiment were similar to other reports which observed the highest phenolic content in the ethyl acetate extract among other solvents such as hexane, chloroform, n-butanol, and water [39,40,41]. A similar observation was made by Abdelwahab et al. [42] and Kaur et al. [43] who observed the highest phenolic rich extract in the ethyl acetate extract when various polarity solvents were used for the extraction of Orthosiphon stamineus and Chukrasia tabularis respectively.
Characteristics of PRE-LM
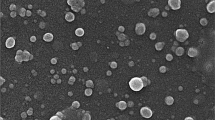
Table 2 illustrated the physicochemical traits of PRE-LM of P. bistorta roots. The encapsulation efficiency was ~ 87.2% (w/w) and surface phenolic compounds were 7.1% (w/w). The results indicated that the spray-dried technique could produce microcapsules with 330.3 nm in size with an acceptable PDI range (0.21) indicating the homogenous dispersion [44]. The total phenolic content of capsules was 172.6 ± 9.24 mg GA eq./g dry microcapsules. Furthermore, the SEM is applied to determine the surface morphology of materials and it is known as a valuable method for monitoring the size and formation of inclusion complexes. The SEM photomicrographs revealed that microcapsules are spherical and mostly with uniform and smooth cover with minimum fractures and signs of collapse (Fig. 1). The size of microcapsules in the SEM micrograph was consistent with the results obtained by Zetasizer. The encapsulation of plant bioactive compounds could protect the phenolics from oxidation, evaporation, and facilitate precision dosing. It is a low-cost and popular method for microencapsulation of bioactive compounds in the industry [45]. Consistent with the results obtained in this study, the early studies reported the encapsulation efficiency ranged from ~ 65 up to 95% (w/w) with the particle size ranging from 100 nm up to 4 µm. The early reports revealed the successful microencapsulation of polyphenols extracted from olive leaves, citrus processing by-products, grape pomace, and pomegranate peel and indicated that the encapsulation of phenolic compounds enhanced the solubility, functionality and shelf life of bioactive phenolic compounds [45,46,47,48].

Scanning electron micrographs of microcapsules containing phenolic enriched extract-loaded microcapsules (PRE-LM) obtained from P. bistorta root
HPLC analysis of PRE-LM
The HPLC profiling of PRE-LM from P. bistorta root indicated various phenolic compounds including gallic acid, syringic acid, ferulic acid, catechin, ellagic acid, naringin, and chrysin with the concentration ranging from 195 to 911 µg/g DW (Table 3). The HPLC chromatogram in Fig. 2 shows the various phenolic compounds in PRE-LM of P. bistorta.

The RP-HPLC chromatogram of various phenolic compounds detected in phenolic enriched extract-loaded microcapsules (PRE-LM) obtained from P. bistorta root. The analysis was performed in triplicates (n = 3)
Biological activities of PRE-loaded microcapsules
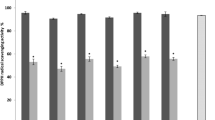
The antioxidant and antibacterial properties of PRE-LM have been shown in Table 4. The microcapsules manifest appreciable antioxidant by preventing the DPPH free radicals with IC50 values of 38.1 ± 4.18 µg/ml but the activities were lower than Vitamin C (17.1 ± 2.37 µg/ml) as the reference antioxidant. Besides that, the antibacterial potential of PRE-LM against the E. coli indicated that MIC values for the microcapsules and oxytetracycline as a reference antibiotic were 52.7 ± 2.86 and 19.2 ± 3.06 µg/ml, respectively. The moderate antibacterial and antioxidant activity of PRE-LM is probably attributed to the presence of phenolic compounds including gallic acid, syringic acid, ferulic acid, catechin, ellagic acid, naringin, and chrysin. It is difficult to say which phenolic compound mainly contributed in the antibacterial activity but most probably the synergistic effects of phenolic compounds may have resulted in the observed antibacterial activity. The mechanisms of action of phenolic compounds on bacterial cell have been partially attributed to inhibition of virulence factors such as enzymes and toxins, damage to the bacterial membrane, and suppression of bacterial biofilm formation [49]. The early studies reported the significant roles of these phenolic compounds in the antibacterial and antioxidant activity of plant extract as there was a direct relationship between the phenolic compounds with biological activities of plant extracts [50, 51].
Animal trial
Table 5 shows the average daily weight gain and feed intake of mice receiving different treatments during the 28 days of study. It can be inferred from the results that the mice receiving normal food (T1) possessed the average daily weight gain with the value of 155.9 mg. In contrast, the mice challenged with enteropathogenic E. coli showed the lowest average daily weight gain (126.3 mg). It was interesting that the mice that received 150 and 300 mg phenolic compounds (GA eq.) /Kg BW through dietary supplementing of microcapsules significantly (p < 0.05) improved the average daily gain with respective values of 134.2 and 162.1 mg. Besides, the daily feed intake data was in harmony with bodyweight alterations. The mice challenging with enteropathogenic E. coli significantly (p < 0.05) reduced the mice's appetite. The inclusion of microcapsules containing phenolic compounds improved the feed intake in a dose-dependent manner similar to the tetracycline as a reference antibiotic.
Liver enzymes and lipid peroxidation status
The obtained results of liver enzymes (ALP, ALT, AST) together with lipid peroxidation have been summarized in Table 6. The results illustrated that the mice challenged by E. coli infection (T2) have significantly (p < 0.05) elevated the values of these parameters. Meanwhile, the groups receiving the different concentrations of microcapsules (T3 and T4) showed lower levels of liver enzymes and lipid peroxidation values significantly (p < 0.05).
From this result, it is postulated that the infecting mice with E. coli reduced the appetite and induced weight loss due to gastroenteritis. The E. coli induced gastroenteritis through adherence to the intestinal mucosa and initiating the production of enterotoxin and cytotoxins [52]. The enterotoxins and cytotoxins reduced the appetite and increased the liver enzymes and lipid peroxidation, resulting in systemic inflammation and oxidative stress induction. The phytobiotic used in this study could improve the weight gain and alleviate the liver enzymes production and lipid peroxidation in the mice challenged by E. coli infection. This observation is most probably attributed to the presence of antimicrobial, antioxidant, and anti-inflammatory phenolic compounds (gallic acid, syringic acid, ferulic acid, catechin, ellagic acid, naringin, and chrysin) present in the phytobiotic [53]. These results were consistent with earlier reports indicating the potential of phenolic compounds in reducing the side effects of photogenic bacterial infection in humans and animals [54,55,56].
Ileum morphometric analysis
The results of the morphometric analysis of ileum including villus height, villus width, crypt depth, and the number of the goblet cell are presented in Table 7. The overall results demonstrated that the mice group infected with E. coli (T2) showed a reduction in the villus height, villus width, and increase in the crypt depth as compared to the uninfected group (T1). The number of goblet cells as a part of the immune system increased significantly (p < 0.05) to confront the enteropathogens. Besides, dietary inclusion of microcapsules (150 and 300 mg phenolic compounds /Kg BW) containing phenolic compounds obtained from P. bistorta root improved the villus height and width together with the reduction in the crypt depth. The microcapsules containing phenolic compounds behaved similarly to the oxytetracycline as reference antibiotics. These results were in agreement with the results of Awad et al. [57] and Fasina et al. [58] who reported the impair in the villus height, villus width, and crypt depth upon the challenge of broiler chicken with enterophategenic bacteria including C. jejuni or Salmonella typhimurium infection, respectively. Further, another study conducted by Mohiti-Asli and Ghanaatparast-Rashti [59] revealed that the inclusion of phenolic compounds in the diet of broiler chicken challenged by E. coli improved the morphometric parameters of the small intestine and reduced the presence of E. coli.
Gene expression analysis
The Claudin-1 and Occludin genes are major constituents of the tight junction complexes in the ileum that regulate the permeability of epithelia. Moreover, The Mucin-2 is a gene responsible for the production of related-mucin proteins, polymerizes into a gel of which 80% (w/w) by weight is oligosaccharide side-chains that are insoluble mucus barrier that serves to protect the ileum epithelium against enteropathogens. The expression of these main marker genes involved in permeability and protection showed that enteropathogenic E. coli suppressed the expression of claudin-1, claudin-2, occludin, and mucin-2 genes significantly (p < 0.05). On the other hand, dietary inclusion of phenolic compounds in the form of microcapsules significantly (p < 0.05) up-regulated the expression of these genes (Table 8). These results revealed that infection with E. coli interrupted the absorption of nutrients through ileum and reduced the mucin production as a defense mechanism in the ileum against enteropathogenic E. coli. These results augur well with the results of average daily weight gain.
E. coli population analysis
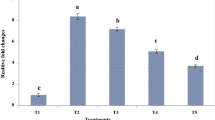
As shown in Fig. 3, the highest E. coli population was found in the ileum digesta of the mice challenged by E. coli. The dietary addition of microcapsules containing phenolic compounds of P. bistorta root significantly (p < 0.05) reduced the E. coli population in the ileum digesta (Fig. 3). The microcapsules containing phenolic compounds behaved similar to the oxytetracycline in reducing the E. coli population in the ileum digesta devoid of their differences in the applied concentrations. Hence the microcapsules containing phenolic compounds obtained from P. bistorta root could play a role as a natural phytobiotic. The release of bioactive phenolic compounds in the ileum section could appear either through the diffusion process or intestinal degradation of wall materials used for the synthesis of microcapsules.

Relative fold changes in the ileum population of E. coli. T1: Normal diet, T2: Normal diet + E. coil infection, T3: Normal diet + E. coil infection + supplemented with microcapsules (150 mg phenolic compounds (GA eq.) /Kg BW, T4: Normal diet + E. coil infection + supplemented with microcapsules (300 mg phenolic compounds (GA eq.) /Kg BW, T5: Normal diet + E. coil infection + supplemented with oxytetracycline 5 mg /Kg BW. Different letters above charts indicate statistically significant differences among groups (P < 0.05). The analysis was performed in triplicates (n = 3)
Conclusion
In this experiment, the phenolic rich extract (PRE) from the root of Polygonum bistorta encapsulated in the protecting wall materials (10% WPC, 20% MS, and 70% MD) to form PRE-loaded spherical microcapsules with the mean diameter of 330 nm, PDI of 0.21 and relatively high entrapment efficiency (87.2% w/w). The synthesized microcapsules contained total phenolics of 172.6 ± 9.24 mg GA eq./g dry microcapsules. The microcapsules contained gallic acid, syringic acid, ferulic acid, catechin, ellagic acid, naringin, and chrysin. The in vitro evaluation revealed the antioxidant and antibacterial activity of microcapsules. The dietary supplementation of microcapsules at the concentrations of 150 and 300 mg total phenolic GA eq./Kg BW/day improved weight gain, liver enzymes, lipid peroxidation, gene expression, morphometric characteristics of the ileum and decreased the population of E. coli present in the ileum significantly (p < 0.05). Consequently, the synthesized microcapsules appeared to be a promising natural antibiotic-alternative called phytobiotic against E. coli infection in mice.
Availability of data and materials
The datasets applied during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ZnO-NPs:
-
Zinc oxide nanoparticle
- F.B.S:
-
Fasting Blood Sugar
- A.S.T:
-
Aspartate aminotransferase
- A.L.T:
-
Alanine aminotransferase
- TNF-α:
-
Tumour necrosis factor-alpha
- iNOS:
-
Inducible nitric oxide synthase
- C.A.T:
-
Catalase
- M.D.A:
-
Malondialdehyde
References
Ndomou SCH, Djikeng FT, Teboukeu GB, Doungue HT, Foffe HAK, Tiwo CT, Womeni HMJJoA. Research F: Nutritional value, phytochemical content, and antioxidant activity of three phytobiotic plants from west Cameroon. J Agri Food Res. 2021;3:100105.
Cushnie T, Lamb A. Antimicrobial activity of flavonoids. Int J Antimicrob Agents. 2005;26(5):343–56.
Shamsudin S, Selamat J, Abdul Shomad M, Ab Aziz MF, Haque Akanda MJJoFQ. Antioxidant properties and characterization of heterotrigona itama honey from various botanical origins according to their polyphenol compounds. 2022;2022:1–14.
Voronkova M, Vysochina G. Bistorta Scop. genus (Polygonaceae) chemical composition and biological activity. Chem Sustain Dev. 2014;22:207–12.
Intisar A, Zhang L, Luo H, Kiazolu JB, Zhang R, Zhang W. Anticancer constituents and cytotoxic activity of methanol-water extract of Polygonum bistorta L. Afr J Tradit Complement Altern Med. 2013;10(1):53–9.
Wang KJ, Zhang YJ, Yang CR. Antioxidant phenolic compounds from rhizomes of Polygonum paleaceum. J Ethnopharmacol. 2005;96(3):483–7.
Pirvu L, Sha’at F, Miclea LC, Savopol T, Neagu G, Udeanu DI, Moisescu MG. Polygonum bistorta L herba et flores polyphenols profile, antioxidant properties and cytotoxic effect on murine fibroblast cell line NIH3T3. FARMACIA. 2017;65(4):571–6.
Wang Y, Du H, Xie M, Ma G, Yang W, Hu Q, Pei FJFP, Life S. Characterization of the physical properties and biological activity of chitosan films grafted with gallic acid and caffeic acid A comparison study. Food Packag Shelf Life. 2019;22:100401.
Salem AM, Mohammaden TF, Ali MA, MohamedHasan EAHFJLs. Ellagic and ferulic acids alleviate gamma radiation and aluminium chloride-induced oxidative damage. Life Sci. 2016;160:2–11.
Negahdari R, Bohlouli S, Sharifi S, Maleki Dizaj S, Rahbar Saadat Y, Khezri K, Jafari S, Ahmadian E, Gorbani Jahandizi N, Raeesi SJPR. Therapeutic benefits of rutin and its nanoformulations. Phytother Res. 2021;35(4):1719–38.
Kim CH, Kim JE, Song Y-JJM. Antiviral activities of quercetin and isoquercitrin against human herpesviruses. Molecules. 2020;25(10):2379.
Cecotti R, Carpana E, Falchero L, Paoletti R, Tava A. Determination of the volatile fraction of Polygonum bistorta L. at different growing stages and evaluation of its antimicrobial activity against two major honeybee (Apis mellifera) pathogens. Chem Biodivers. 2012;9(2):359–69.
Pawłowska KA, Hałasa R, Dudek MK, Majdan M, Jankowska K, Granica S. Antibacterial and anti-inflammatory activity of (bistort Bistorta) officinalis aqueous extract and its major components. Justification of the usage of the medicinal plant material as a traditional topical agent. J Ethnopharmacol. 2020;260:113077.
Haminiuk CW, Maciel GM, Plata-Oviedo MS, Peralta RM. Phenolic compounds in fruits–an overview. Int J Food Sci Technol. 2012;47(10):2023–44.
Chen L, Gnanaraj C, Arulselvan P, El-Seedi H, Teng H. A review on advanced microencapsulation technology to enhance bioavailability of phenolic compounds: based on its activity in the treatment of type 2 diabetes. Trends Food Sci Technol. 2019;85:149–62.
Comunian TA, Boillon MR, Thomazini M, Nogueira MS, de Castro IA, Favaro-Trindade CS. Protection of echium oil by microencapsulation with phenolic compounds. Food Res Int. 2016;88:114–21.
Ozkan G, Franco P, De Marco I, Xiao J, Capanoglu E. A review of microencapsulation methods for food antioxidants: Principles, advantages, drawbacks and applications. Food Chem. 2019;272:494–506.
Paulo F, Santos L. Design of experiments for microencapsulation applications: A review. Mater Sci Eng C Mater Biol Appl. 2017;77:1327–40.
Aguiar J, Estevinho BN, Santos L. Microencapsulation of natural antioxidants for food application–The specific case of coffee antioxidants–A review. Trends Food Sci Technol. 2016;58:21–39.
Karimi E, Mehrabanjoubani P, Es-Haghi A, Chamani J. Phenolic compounds of endemic buxus plants in caspian hyrcanian forest buxus hyrcana pojark and their biological activities. Pharma Chem J. 2019;53(8):741–7.
Karimi E, Ghorbani Nohooji M, Habibi M, Ebrahimi M, Mehrafarin A, Khalighi-Sigaroodi F. Antioxidant potential assessment of phenolic and flavonoid rich fractions of Clematis orientalis and Clematis ispahanica (Ranunculaceae). Nat Prod Res. 2018;32(16):1991–5.
Sablania V, Bosco SJD, Rohilla S, Shah MA. Microencapsulation of Murraya koenigii L leaf extract using spray drying. J Food Meas Character. 2018;12(2):892–901.
Moharreri M, Vakili R, Oskoueian E, Rajabzadeh G. Phytobiotic role of essential oil-loaded microcapsules in improving the health parameters in Clostridium perfringens-infected broiler chickens. Ita J Animal Sci. 2021;20(1):2075–85.
Moharreri M, Vakili R, Oskoueian E, Rajabzadeh G. Effects of microencapsulated essential oils on growth performance and biomarkers of inflammation in broiler chickens challenged with salmonella enteritidis. J Saudi Soc Agri Sci. 2022;21(5):349–57.
Moharreri M, Vakili R, Oskoueian E, Rajabzadeh G. Evaluation of microencapsulated essential oils in broilers challenged with salmonella enteritidis: a focus on the body’s antioxidant status, gut microbiology, and morphology. Arch Razi Ins. 2022;77(2):629–39.
Oskoueian E, Abdullah N, Zulkifli I, Ebrahimi M, Karimi E, Goh YM, Oskoueian A, Shakeri M. Cytoprotective effect of palm kernel cake phenolics against aflatoxin B1-induced cell damage and its underlying mechanism of action. BMC Complement Altern Med. 2015;15(1):1.
Yi X, Zheng Q, Ding B, Pan M-h, Chiou Ys, Li L, Li ZJL. Liposome-whey protein interactions and its relation to emulsifying properties. LWT. 2019;99:505–12.
Karimi E, Mehrabanjoubani P, Keshavarzian M, Oskoueian E, Jaafar HZ, Abdolzadeh A. Identification and quantification of phenolic and flavonoid components in straw and seed husk of some rice varieties (Oryza sativa L.) and their antioxidant properties. J Sci Food Agri. 2014;94(11):2324–30.
Karimi E, Mehrabanjoubani P, Homayouni-Tabrizi M, Abdolzadeh A, Soltani M. Phytochemical evaluation, antioxidant properties and antibacterial activity of Iranian medicinal herb Galanthus transcaucasicus Fomin. J Food Meas Character. 2018;12(1):433–40.
Banothu V, Neelagiri C, Adepally U, Lingam J, Bommareddy K. Phytochemical screening and evaluation of in vitro antioxidant and antimicrobial activities of the indigenous medicinal plant Albizia odoratissima. Pharma Biol. 2017;55(1):1155–61.
Garcia YJ, Rodríguez-Malaver AJ, Peñaloza N. Lipid peroxidation measurement by thiobarbituric acid assay in rat cerebellar slices. J Neurosci Methods. 2005;144(1):127–35.
Kathirvel E, Chen P, Morgan K, French SW, Morgan TR. Oxidative stress and regulation of anti-oxidant enzymes in cytochrome P4502E1 transgenic mouse model of non-alcoholic fatty liver. J Gastroenterol Hepatol. 2010;25(6):1136–43.
Volynets V, Rings A, Bárdos G, Ostaff MJ, Wehkamp J, Bischoff SC. Intestinal barrier analysis by assessment of mucins, tight junctions, and α-defensins in healthy C57BL/6J and BALB/cJ mice. Tissue Barriers. 2016;4(3):e1208468.
Barroso FAL, de Jesus LCL, de Castro CP, Batista VL, Ferreira Ê, Fernandes RS, de Barros ALB, Leclerq SY, Azevedo V, Mancha-Agresti P. Intake of Lactobacillus delbrueckii (pExu: hsp65) prevents the inflammation and the disorganization of the intestinal mucosa in a mouse model of mucositis. Microorganisms. 2021;9(1):107.
Feng Y, Gong J, Yu H, Jin Y, Zhu J, Han Y. Identification of changes in the composition of ileal bacterial microbiota of broiler chickens infected with Clostridium perfringens. Veterinary Microbiol. 2010;140(1–2):116–21.
Oskoueian E, Abdullah N, Oskoueian A. Effects of flavonoids on rumen fermentation activity, methane production, and microbial population. BioMed Res Int. 2013;2013:349129.
Si C, Kun-Lun H, Wen-Tao X, Yuan L, Yun-Bo L. Real-time quantitative PCR detection of Escherichia coli O157: H7. Chin J Agri Biotechnol. 2007;4(1):15–9.
Denman SE, McSweeney CS. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol Ecol. 2006;58(3):572–82.
Mariem S, Hanen F, Inès J, Mejdi S, Riadh K. Phenolic profile, biological activities and fraction analysis of the medicinal halophyte Retama raetam. South Afr J Botany. 2014;94:114–21.
Ju HY, Chen SC, Wu KJ, Kuo HC, Hseu YC, Ching H, Wu CR. Antioxidant phenolic profile from ethyl acetate fraction of Fructus Ligustri Lucidi with protection against hydrogen peroxide-induced oxidative damage in SH-SY5Y cells. Food Chem Toxicol. 2012;50(3):492–502.
Olatunji OJ, Chen H, Zhou Y. Effect of the polyphenol rich ethyl acetate fraction from the leaves of lycium chinensemill. On oxidative stress, dyslipidemia, and diabetes mellitus in streptozotocin-nicotinamide induced diabetic rats. Chem Biodivers. 2017;14(10):e1700277.
Abdelwahab SI, Mohan S, Mohamed Elhassan M, Al-Mekhlafi N, Mariod AA, Abdul AB, Abdulla MA, Alkharfy KM. Antiapoptotic and antioxidant properties of Orthosiphon stamineus benth Cat’s Whiskers: intervention in the Bcl-2-mediated apoptotic pathway. Evid-Based Complement Altern Med. 2010;2011:156765.
Kaur R, Arora S, Singh B. Antioxidant activity of the phenol rich fractions of leaves of Chukrasia tabularis A. Juss Biores Technol. 2008;99(16):7692–8.
Hasan M, Belhaj N, Benachour H, Barberi-Heyob M, Kahn C, Jabbari E, Linder M. Arab-Tehrany EJIjop: Liposome encapsulation of curcumin: physico-chemical characterizations and effects on MCF7 cancer cell proliferation. Int J Pharm. 2014;461(1–2):519–28.
Paulo F, Santos L. Deriving valorization of phenolic compounds from olive oil by-products for food applications through microencapsulation approaches: a comprehensive review. Crit Rev Food Sci Nutr. 2020;61:1–26.
Popović DA, Milinčić DD, Pešić MB, Kalušević AM, Tešić ŽL, Nedović VA: 12 - Encapsulation technologies for polyphenol-loaded microparticles in food industry. In: Green Food Processing Techniques. edn. Edited by Chemat F, Vorobiev E: Academic Press; 2019;335–367.
Rahaiee S, Assadpour E, Faridi Esfanjani A, Silva AS, Jafari SM. Application of nano/microencapsulated phenolic compounds against cancer. Adv Colloid Interface Sci. 2020;279:102153.
Munin A, Edwards-Lévy F. Encapsulation of natural polyphenolic compounds; a review. Pharmaceutics. 2011;3(4):793–829.
Bouarab-Chibane L, Forquet V, Lantéri P, Clément Y, Léonard-Akkari L, Oulahal N, Degraeve P, Bordes C. Antibacterial properties of polyphenols: characterization and QSAR (Quantitative structure–activity relationship) models. Front Microbiol. 2019;10:829.
Lang G, Buchbauer G. A review on recent research results 2008–2010 on essential oils as antimicrobials and antifungals A review. Flav Fragr J. 2012;27(1):13–39.
Solórzano-Santos F, Miranda-Novales MG. Essential oils from aromatic herbs as antimicrobial agents. Curr Opinion Biotechnol. 2012;23(2):136–41.
Stahl M, Graef FA, Vallance BA: Mouse Models for Campylobacter jejuni Colonization and Infection. In: Campylobacter jejuni. edn.: Springer; 2017: 171–188.
Sarı A, Şahin H, Özsoy N, Çelik BÖ. Phenolic compounds and in vitro antioxidant, anti-inflammatory, antimicrobial activities of Scorzonera hieraciifolia Hayek roots. South Afr J Botany. 2019;125:116–9.
Castillo S, Dávila-Aviña J, Heredia N, Garcia S. Antioxidant activity and influence of Citrus byproduct extracts on adherence and invasion of Campylobacter jejuni and on the relative expression of cadF and ciaB. Food Sci Biotechnol. 2017;26(2):453–9.
Silvan JM, Mingo E, Martinez-Rodriguez AJ. Grape seed extract (GSE) modulates campylobacter pro-inflammatory response in human intestinal epithelial cell lines. Food Agri Immunol. 2017;28(5):739–53.
Saha P, Talukdar AD, Nath R, Sarker SD, Nahar L, Sahu J, Choudhury MD. Role of natural phenolics in hepatoprotection: A mechanistic review and analysis of regulatory network of associated genes. Front Pharmacol. 2019;10:509.
Awad W, Dublecz F, Hess C, Dublecz K, Khayal B, Aschenbach J, Hess MJPs. Campylobacter jejuni colonization promotes the translocation of Escherichia coli to extra-intestinal organs and disturbs the short-chain fatty acids profiles in the chicken gut. Poult Sci. 2016;95(10):2259–65.
Fasina Y, Hoerr F, McKee S, Conner DJAd. Influence of Salmonella enterica serovar Typhimurium infection on intestinal goblet cells and villous morphology in broiler chicks. Avian Dis. 2010;54(2):841–7.
Mohiti-Asli M, Ghanaatparast-Rashti MJJoAAR. Comparing the effects of a combined phytogenic feed additive with an individual essential oil of oregano on intestinal morphology and microflora in broilers. J Appl Ani Res. 2018;46(1):184–9.
Acknowledgements
The authors are grateful to the Islamic Azad University of Mashhad for the laboratory facilities.
Funding
There has been no financial support for this work.
Author information
Authors and Affiliations
Contributions
Zahra kadkhoda Mezerji, Reza Boshrouyeh, Seyedehfarnaz Hafezian Razavi, Shaghayegh Ghajari, Hasti Hajiha, Negin Shafaei: Study design, experimental work, formal analysis, and writing original draft; Ehsan Karimi and Ehsan Oskoueian.: analysis, methodology, project administration, supervision, review, and editing of the original draft; All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal handling methods were accomplished as per the Islamic Azad University of Mashhad, IRAN regulations with their prior approval for using the animals (IR.IAU.MSHD.REC.1399.019.). All methods and procedures were carried out in accordance with the relevant guidelines and regulations. All methods are reported in accordance with ARRIVE guidelines.
Consent to publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Mezerji, Z.k., Boshrouyeh, R., Razavi, S.H. et al. Encapsulation of Polygonum bistorta root phenolic compounds as a novel phytobiotic and its protective effects in the mouse model of enteropathogenic Escherichia coli infection. BMC Complement Med Ther 23, 49 (2023). https://doi.org/10.1186/s12906-023-03868-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-023-03868-2