
Abstract
Background
The CDH1 gene codes for the epithelial-cadherin (E-cad) protein, which is embedded in the plasma membrane of epithelial cells to form adherens junctions. E-cad is known to be essential for maintaining the integrity of epithelial tissues, and the loss of E-cad has been widely considered a hallmark of metastatic cancers enabling carcinoma cells to acquire the ability to migrate and invade nearby tissues. However, this conclusion has come under scrutiny.
Methods
To assess how CDH1 and E-cad expression changes during cancer progression, we analyzed multiple large transcriptomics, proteomics, and immunohistochemistry datasets on clinical cancer samples and cancer cell lines to determine the CDH1 mRNA and E-cad protein expression profiles in tumor and normal cells.
Results
In contrast to the textbook knowledge of the loss of E-cad during tumor progression and metastasis, the levels of CDH1 mRNA and E-cad protein are either upregulated or remain unchanged in most carcinoma cells compared to normal cells. In addition, the CDH1 mRNA upregulation occurs in the early stages of tumor development and the levels remain elevated as tumors progress to later stages across most carcinoma types. Furthermore, E-cad protein levels are not downregulated in most metastatic tumor cells compared to primary tumor cells. The CDH1 mRNA and E-cad protein levels are positively correlated, and the CDH1 mRNA levels are positively correlated to cancer patient’s survival. We have discussed potential mechanisms underlying the observed expression changes in CDH1 and E-cad during tumor progression.
Conclusions
CDH1 mRNA and E-cadherin protein are not downregulated in most tumor tissues and cell lines derived from commonly occurring carcinomas. The role of E-cad in tumor progression and metastasis may have previously been oversimplified. CDH1 mRNA levels may serve as a reliable biomarker for the diagnosis of some tumors (such as colon and endometrial carcinomas) due to the marked upregulation of CDH1 mRNA in the early stages of tumor development of these carcinomas.
Similar content being viewed by others

Background
CDH1 is a gene that codes for the epithelial-cadherin (E-cad) protein, which is embedded in the plasma membrane of epithelial cells forming the tissues that cover the body surfaces and line the walls of cavities, channels, and glands [1, 2]. E-cad is a calcium-dependent cell–cell adhesion protein that forms homophilic interactions in adjacent epithelial cells establishing adherens junctions [3, 4]. This protein plays a major role in embryonic development and morphogenesis [5, 6]. In its inactive form, E-cad contains a short signal sequence for import to the endoplasmic reticulum (ER), a 130 amino acid pro-peptide, a single transmembrane domain, a 150 amino acid cytoplasmic domain, and a 550 amino acid ectodomain [7, 8]. E-cad is activated after cleavage of the pro-peptide in the presence of calcium ions [9]. In the extracellular matrix (ECM), the ectodomains of E-cad on adjacent cells bind each other to form adherens junctions, while the cytoplasmic domain of E-cad interacts with β-catenin, which in turn binds α-catenin connecting to the actin cytoskeleton of the cell leading to the stabilization and integrity of the epithelial tissues [10, 11].
The loss of E-cad expression has been considered a hallmark of cancer progression and metastasis [12, 13] via loss of heterozygosity of the chromosomal region 16q22.1 containing the CDH1 locus, nonsense mutations [14], or promoter methylation [15]. E-cad activity as a tumor suppressor manifests via its loss during epithelial-mesenchymal transition (EMT) and/or regulation during metastatic progression, where its loss leads to increased tumor cell migration and invasion [16]. E-cad also plays a role in primary tumor development, progression [17, 18], and metastatic colonization [19]. The loss of E-cad expression is thought to disrupt adherens junctions leading to the acquisition of motility/invasiveness of metastasizing tumor cells. Although some carcinoma cells undergo EMT, many carcinoma cells neither fully lose the ability to produce E-cad nor undergo a mesenchymal‐to‐epithelial transition (MET) during metastasis [20,21,22]. Assertions as to the necessity of EMT and its reverse MET in metastasis have been controversial [23,24,25] as many metastatic tumor cells still express E-cad [26,27,28,29].
In this study, we analyzed multiple large transcriptomics, proteomics, and immunohistochemistry datasets on clinical cancer samples and cancer cell lines to determine the levels of CDH1 mRNA and E-cad protein in different carcinomas during tumor progression. Strikingly, the levels of CDH1 mRNA and E-cad protein were not reduced in most of the examined tumors, even in the later stages of cancer compared to respective healthy tissues. The only exception to this trend was kidney cancer, which exhibited significantly lower levels of CDH1 mRNA and E-cad protein, the pattern normally described in textbooks. The observations presented in this study demonstrate that the changes in E-cad expression during tumor progression and metastasis are more complex than widely believed.
Methods
Analysis of CDH1 mRNA levels in cancer clinical samples and cancer cell lines
The Gene Expression Profiling Interactive Analysis 2 (GEPIA2) web server (http://gepia2.cancer-pku.cn/#index) [30, 31] was used to analyze levels of CDH1 mRNA in the tumors of interest. GEPIA2 is a resource for gene expression analysis compiling tumor and normal samples from The Cancer Genome Atlas (TCGA), a database containing samples from 11,000 patients, and the Genome-Typing Expression (GTEx), a database containing 948 post-mortem donors and approximately 17,382 RNA sequencing (RNA-seq) samples across 54 tissue sites and 2 cell lines [32]. CDH1 mRNA levels between tumor and normal samples for breast, colon, lung, ovarian, pancreatic, endometrial, kidney, liver, and head/neck tissues were analyzed using one-way analysis of variance (ANOVA). A change was considered significantly different if i) the log2 fold change was larger than 1 (representing an actual fold change of 2), ii) the q-value was smaller than 0.01 or the p-value was smaller than 0.05, and iii) the samples were available in both the TCGA and GTEx databases. The CDH1 mRNA expression in tumor subtypes relative to corresponding normal tissues was analyzed with the same parameter as discussed above. CDH1 mRNA expression across specific tumor stages was performed using a one-way ANOVA, in which expression was compared among the major stages of specific carcinomas. Only the major stages containing enough samples for statistical analysis were analyzed.
The Cancer Dependency Map (DepMap, https://depmap.org/portal/), which systematically identifies genetic and pharmacologic dependencies and biomarkers for 1,072 cancer cell lines from various lineages [33], was used to analyze the CDH1 mRNA profile in cell lines derived from breast, colorectal, endometrial, head/neck, kidney, lung, liver, pancreatic, and ovarian carcinomas. The CDH1 mRNA levels in the kidney, breast, liver, colorectal, ovarian, pancreatic, endometrial, and lung carcinomas cell lines were analyzed using the Cancer Cell Line Encyclopedia (CCLE) RNA-seq data available on the European Molecular Biology Laboratory’s European Bioinformatics Institute (EMBL-EBI) database (https://www.ebi.ac.uk/gxa/experiments/E-MTAB-2770/) with the expression value set at 0.5 and the data reported as transcripts per million (TPM) [34]. The results were reported in TPM where low/no expression was defined as 0–10 TPM, medium expression was defined as 11–100 TPM, and high expression was defined as 101–2,120 TPMs. The cell lines used are listed in Supplementary Tables—Additional file 2 (the same for other sections that were involved in using cell lines), and the sources of the cell lines were described in the respective studies.
CDH1 promoter methylation analysis in tissue samples
The University of Alabama at Birmingham Cancer Data Analysis Portal (UALCAN, http://ualcan.path.uab.edu/analysis-prot.html), an interactive web source for analyzing cancer OMICS data with a focus on transcriptomics and proteomics, and the TCGA methylation datasets (http://ualcan.path.uab.edu/analysis.html) were used to assess promoter methylation in kidney carcinomas compared to normal samples, and among the major stages of kidney carcinoma [35, 36]. The beta value, the ratio of methylated probe intensity to the total probe intensity (the sum of methylated and unmethylated probe intensity), was reported. The beta values of CpG probes ([TSS200, TSS1500]) located up to 1,500 bp upstream of the CDH1 gene’s start site were plotted. Beta values range from 0 (unmethylated) to 1 (fully methylated).
Analysis of E-cad protein levels in clinical cancer samples, cancer cell lines, and normal samples
The expression levels of E-cad protein in cancer and normal tissues were determined using the Clinical Proteomic Tumor Analysis Consortium (CPTAC) Confirmatory/Discovery dataset available on the UALCAN web-server (http://ualcan.path.uab.edu/analysis-prot.html) [37]. The log2 spectral count ratios obtained from CPTAC were normalized within each sample profile and then normalized across samples. The results were presented using Z-values, the standard deviation from the median across samples for any given carcinoma compared to normal tissues. Differential expression of E-cad with p-values smaller than 0.05 was considered significant.
The expression of E-cad in cancer cell lines was determined using the proteomics data available in the DepMap portal (https://depmap.org/portal/interactive/) produced from the quantitative analyses of protein expression in 375 cancer cell lines in the CCLE [38]. The protein expression is reported as values closely related to log2-transformed ratios to the bridge, a sample of 10 cell lines from the CCLE selected for maximal protein expression diversity to help with the normalization of protein levels in the cell lines analyzed [38]. A cell line was said to have a high expression of E-cad if the log2-transformed values were higher than 0, and a low expression if the log2-transformed values were lower than 0. In this context, 0 means that there is no difference between the expression of E-cad in the cell line of interest compared to the levels of E-cad in the bridge mixture.
To analyze whether the levels of E-cad were different between metastatic and primary carcinoma cell lines, the E-cad expression data was downloaded from the DepMap portal (Proteomics data) [38], then cancer cell lines derived from breast, colon, head/neck, lung, ovarian, and endometrial carcinomas were separated into primary tumor cell lines and metastatic cell lines. The mean E-cad expression for the defined groups in specific lineages was determined and analyzed using one-way ANOVA using the PSI-Plot software. Analyses for kidney and liver carcinoma cell lines were not performed because of the lack of enough metastatic cell lines to calculate statistical significance. Metastatic and primary cell lines from all lineages were compared also using one-way ANOVA; this comparison included metastatic and primary cell lines derived from kidney and liver cancer as well. p-value less than 0.05 represented significant differential expression between primary and metastatic carcinoma cell lines.
Analysis of immunohistochemistry staining of E-cad in clinical cancer tissue samples
E-cad expression in immunohistochemically stained with CAB072856 antibody, and pathologist-certified/annotated images for breast, colon, head/neck, kidney, lung, liver, pancreatic, ovarian, and endometrial carcinomas, and corresponding normal tissue microarrays stored on the Human Protein Atlas (HPA) (https://www.proteinatlas.org/) was assessed [39]. The HPA summarized the results as high, medium, and low/not detected E-cad staining intensity.
Determination of the relationship between CDH1 mRNA levels and E-cad protein levels in cancer cell lines
Pearson Correlation Analysis and Spearman Correlation Analysis were carried out using the DepMap portal (https://depmap.org/portal/interactive/) to compare the CDH1 mRNA and E-cad protein levels in 9 individual carcinoma cell lineages, including endometrial, head/neck, pancreatic, ovarian, liver, kidney, colorectal, breast, and lung tissues [33, 38]. The relationship between CDH1 mRNA levels and E-cad protein levels in all the carcinoma cell lines was also determined.
Determination of the relationship between CDH1 mRNA levels in carcinoma tissues and cancer patient’s survival
GEPIA2 (http://gepia2.cancer-pku.cn/#index) was used to assess the relationship between CDH1 mRNA levels in carcinoma tissues and patient’s overall survival (OS) and disease-free survival (DFS) [30, 31]. GEPIA2 employs the Log-rank test, also known as the Mantel-Cox test, to assess the null hypothesis that there is no significant difference in survival among the different groups being compared [30, 31]. Cancer patients with defined types of cancer were divided equally into two groups based on the median level of CDH1 mRNA: the high CDH1 mRNA group represented half of the patients with tumors expressing higher levels of CDH1 mRNA, and the low CDH1 mRNA group represented half of the patients with tumors expressing lower levels of CDH1 mRNA relative to the median CDH1 mRNA level in the group. A p-value less than 0.05 was considered significant, meaning that the null hypothesis was rejected and that there was a significant difference in the survival between the two groups.
Statistical analysis
The difference in CDH1 mRNA expression between carcinoma and normal tissues, between carcinoma subtypes and normal tissues, across major cancer stages, and between metastatic and primary cancer cell lines were analyzed with one-way ANOVA. Survival analyses were performed using a Log-rank test (Mantel-Cox test) [30, 31]. The difference in E-cad expression and promoter methylation between carcinoma types and normal tissues, and between stages and normal tissues were analyzed using a t-test [35,36,37, 40]. The RNA seq data and proteomic data on the sites have been normalized by the authors of the sites. The correlation between CDH1 mRNA and E-cad protein expression was analyzed using both Pearson correlation analysis and Spearman correlation analysis [33, 38].
Results
CDH1 mRNA is upregulated or unchanged in most carcinoma tissues
The loss of E-cad expression is considered a hallmark of cancer invasion and metastasis potentially via a role in EMT [12]. To determine whether the reported loss of E-cad expression in some cancers [14, 15, 41] is consistent with CDH1 transcription in clinical tissue samples, we assessed mRNA levels in carcinoma and normal tissues in various parts of the body from the TCGA and GTEx databases using the GEPIA2 web server (http://gepia2.cancer-pku.cn/#index). Surprisingly, most carcinoma tissues did not show significant reductions in CDH1 mRNA levels compared to normal tissues (Fig. 1). Among the 9 types of commonly occurring carcinomas analyzed, CDH1 mRNA levels in 6 of them (67%) were upregulated (Fig. 1A), 2 of them (22%) were unchanged (Fig. 1B), and only one of them (11%) was downregulated (Fig. 1C) compared to corresponding normal tissues. Thus, most carcinoma samples examined did not exhibit a reduction in CDH1 mRNA levels.

CDH1 mRNA is either upregulated or remains unchanged in most carcinoma tissues. A, CDH1 mRNA is significantly upregulated in tissues derived from endometrial (number of normal tissues, N = 91; number of tumor tissues, T = 174), pancreatic (N = 171; T = 179), ovarian (N = 88; T = 426), lung (N = 347; T = 483), colon (N = 349; T = 275), and breast (N = 281; T = 1,085) carcinomas compared to the corresponding normal tissues. B, CDH1 mRNA remains unchanged in the liver (N = 160; T = 369) and head/neck (HNSC) (N = 44; T = 519) carcinomas compared to the corresponding normal tissues. C, CDH1 mRNA is significantly downregulated in kidney carcinoma (N = 100; T = 523) compared to the normal samples. Grey, normal; red, carcinoma. **, p < 0.01
We then assessed CDH1 mRNA levels in different subtypes or stages of carcinomas in which CDH1 mRNA was upregulated or unchanged. The levels of CDH1 mRNA were significantly upregulated in all subtypes of breast carcinoma (Fig. 2A), colon carcinoma (Fig. 2B), pancreatic carcinoma (Supplementary Fig. 1A—Additional file 1), and lung carcinoma (Supplementary Fig. 1B—Additional file 1) and remained mostly unchanged in the subtypes of head/neck carcinoma and liver carcinoma (Fig. 2C-D). Further analysis demonstrated that the CDH1 mRNA levels were not significantly changed across the major cancer stages in breast carcinoma (Fig. 2E), colon carcinoma, pancreatic carcinoma, lung carcinoma, and endometrial carcinoma (Supplementary Fig. 1C-F—Additional file 1). Among the cancers examined, the levels of CDH1 mRNAs only exhibited significant downregulation between stage 2 and stage 3 of ovarian cancer (Fig. 2F). These results suggest that the CDH1 mRNA upregulation occurs in the early stages of tumor development and the levels remain high as tumors progress to later stages across most carcinoma subtypes except for ovarian cancer, where downregulation of CDH1 mRNA is observed when the tumors progress from stage 2 to stage 3 (Fig. 2F). Since CDH1 mRNA is markedly upregulated in some types of tumors, for example, colon carcinoma (26x) and endometrial carcinoma (148x), compared to corresponding normal tissues (Fig. 1A), with further research, CDH1 mRNA levels may be established as a diagnostic biomarker for early detection of these carcinomas.

CDH1 mRNA is either upregulated or remains unchanged in tumor subtypes and major stages of most carcinoma tissues. The levels of CDH1 mRNA were significantly upregulated in all breast cancer subtypes luminal B (T = 194), luminal A (T = 415), HER2 (T = 66), and basal-like (T = 135) compared to the normal tissues (N = 291) (A) and in all colon cancer subtypes: MSS (T = 175), MSI-L (T = 48) and MSI-H (T = 48) compared to the normal tissues (N = 349) (B). C, CDH1 mRNA levels remain unchanged in the four subtypes of head/neck carcinoma subtypes: mesenchymal (T = 75), classical (T = 49), basal (T = 87), and atypical (T = 67) compared to the normal tissues (N = 44). D, CDH1 mRNA is significantly upregulated in iCluster 1 (T = 53) but remains unchanged in iCluster 2 (T = 55) and iCluster 3 (T = 63) of liver carcinoma compared to the normal tissues (N = 160). Grey, normal; red, carcinoma in (A)-(D). The levels of CDH1 mRNA are not significantly different across the major stages of breast carcinoma (p-value = 0.265) (E) but are significantly downregulated when ovarian carcinoma progresses from stage 2 to stage 3 (p-value = 0.015) (F). *, p < 0.05, **, p < 0.01
CDH1 mRNA is highly expressed in most carcinoma cell lines while non-epithelial cancer cell lines exhibit lower expression of CDH1 mRNA
To further assess the transcription of the CDH1 gene, we analyzed CDH1 mRNA levels in different cancer cell lines stored on the DepMap portal (https://depmap.org/portal/). Consistent with the data on tumor tissues (Fig. 1), CDH1 mRNA expression in most carcinoma cell lines derived from commonly occurring carcinomas was in the range of moderate to high (Fig. 3A, Table 1). For example, among 55 breast cancer cell lines, 34 (62%) cell lines exhibited high levels of CDH1 mRNA, 4 (7%) exhibited medium levels, and 17 (31%) cell lines showed low or no detection of CDH1 mRNA (Fig. 3B, Table 1).

CDH1 mRNA is highly expressed in most carcinoma cell lines and is expressed at low levels in non-epithelial cancer cell lines. A, CDH1 mRNA is highly expressed in carcinoma cell lines but is expressed at low levels in non-epithelial cancer cell lines. B, CDH1 expression in breast cancer cell lines, shown as a representative of CDH mRNA expression in most carcinoma cell lines. C, CDH1 mRNA levels remain unchanged in most non-epithelial tumor tissues compared to the corresponding normal tissues (cancer name in black) except for skin cutaneous melanoma (SKCM) which demonstrated a significant downregulation (green; q < 0.01), and testicular germ cell tumor (TGCT) and thymoma (THYM) which demonstrated an upregulation of CDH1 mRNA compared to the corresponding normal tissues (red; q < 0.01). DLBCL, diffuse large B-cell lymphoma; GBM, glioblastoma multiforme; UVM, uveal melanoma; LAML, acute myeloid leukemia; LGG, brain lower grade glioma; MESO, mesothelioma; PCPG, pheochromocytoma and paraganglioma; SARC, sarcoma
While E-cad plays an essential role in cell–cell adhesion in epithelial tissues [3, 10, 11, 42], CDH1 is also expressed in non-epithelial cells [43]. Consistent with its non-essential role in cell–cell adhesion in non-epithelial tissues/cells, CDH1 levels were lower than those in carcinoma cells (Fig. 3A). In addition, CDH1 mRNA levels did not significantly change in most of the tested non-epithelial cancers (including lymphoid neoplasm diffuse large B-cell lymphoma, glioblastoma multiforme, acute myeloid leukemia, brain lower-grade glioma, mesothelioma, pheochromocytoma and paraganglioma, sarcoma, and uveal melanoma) compared to corresponding normal tissues (Fig. 3C). The exceptions were testicular germ cell tumors and thymoma, in which CDH1 mRNA was upregulated, and skin cutaneous melanoma, in which CDH1 mRNA was downregulated compared to corresponding normal tissue (Fig. 3C).
E-cad protein is not downregulated in most carcinoma tissues and carcinoma cell lines
We next determined how E-cad protein changed in carcinoma tissues since higher mRNA levels do not always result in higher protein levels [44,45,46,47,48,49]. Consistent with CDH1 mRNA expression in most carcinoma tissues (Fig. 1) and carcinoma cell lines (Fig. 3A-B), E-cad protein was either significantly upregulated or remained unchanged in most carcinoma tissues compared to corresponding normal tissue (Fig. 4A-B). Among the tested carcinoma tissues, E-cad levels were significantly upregulated in breast, endometrial, ovarian, and lung carcinomas (Fig. 4A), and remained unchanged in colon and head/neck carcinomas (Fig. 4B); the levels of E-cad were significantly downregulated only in kidney, pancreatic, and liver carcinomas (Fig. 4C).

E-cad protein is either significantly upregulated or remains unchanged in most carcinoma tissues. A, E-cad protein expression is significantly upregulated in breast carcinoma (N = 25; T = 125), endometrial carcinoma (N = 31; T = 100), ovarian carcinoma (N = 25; T = 100), and lung carcinoma (N = 111; T = 111) compared to their corresponding normal tissues. B, E-cad protein expression remains unchanged in colon carcinoma (N = 100; T = 97) and head/neck carcinoma compared to the corresponding normal tissues (N = 71; T = 108). C, E-cad protein expression is downregulated in kidney carcinoma (N = 84; T = 110), pancreatic carcinoma (N = 74; T = 137), and liver carcinoma (N = 165; T = 165) compared to their corresponding normal tissues. Blue, normal; red, carcinoma in (A)-(C). E-cad protein is significantly upregulated in the different subtypes: luminal (n = 64), HER2 + (n = 10), and TNBC (n = 16) (D), and in the major stages: stage 1 (n = 4), stage 2 (n = 74), and stage 3 (n = 32) (E) of breast carcinomas compared to the normal tissues (n = 18). F, E-cad expression is significantly upregulated in IDC (n = 93) but downregulated in ILC (n = 10) compared to the normal tissues (n = 18). *, p < 0.05, **, p < 0.01
In carcinomas in which E-cad was upregulated/unchanged, this expression pattern was reflected in most cancer subtypes and stages (Fig. 4E and Supplementary Fig. 2—Additional file 1). For example, E-cad expression was significantly upregulated in luminal (estrogen receptor (ER) + , progesterone receptor (PR) ± , human epidermal growth factor 2 (HER2)-, and low levels of Ki-67 protein), and HER2 positive breast cancer (ER-, PR-, and HER2 +), and remained unchanged in triple-negative breast cancer (ER-, PR-, HER2-) (TNBC) compared to normal tissues (Fig. 4D). In addition, E-cad levels were mostly upregulated or remained unchanged during tumor progression in most carcinomas (Table 2). For example, the levels of E-cad were significantly upregulated in stage 2, and stage 3 and remained unchanged in stage 1 of breast carcinoma compared to normal tissue (Fig. 4E). Similarly, E-cad levels were predominately upregulated or remained unchanged in the major cancer stages of endometrial, lung, ovarian, head/neck, and colon carcinomas (Supplementary Fig. 2—Additional file 1). Interestingly, E-cad is significantly downregulated in infiltrating lobular carcinoma (ILC) of breast cancer, consistent with the observations from other groups [14, 50,51,52], but upregulated in infiltrating ductal carcinoma (IDC) (Fig. 4F), suggesting that E-cad levels may serve as a marker to differentiate ILC from IDC breast cancer.
In agreement with the data from tissues, analysis of the CCLE proteomics database in the DepMap portal revealed that most of the tested carcinoma cell lines expressed elevated levels of E-cad protein. Specifically, 67% of breast carcinoma cell lines, 77% of colorectal carcinoma cell lines, 60% of pancreatic carcinoma cell lines, 77% of endometrial carcinoma cell lines, and 100% of head/neck carcinoma cell lines expressed elevated levels of E-cad. Most of the cell lines derived from kidney, liver, lung, and ovarian carcinomas exhibited low levels of E-cad (Table 3).
We also assessed the scanned images of E-cad immunohistochemistry staining of different carcinoma tissues stored on the HPA website (https://www.proteinatlas.org/humanproteome/pathology) [39]. All the representative samples for colorectal, pancreatic, endometrial, ovarian, and liver carcinomas, 82% of breast cancer, 84% of lung cancer, and 75% of head/neck carcinoma samples exhibited medium to high levels of E-Cad staining, which were comparable to those of corresponding normal tissues (Fig. 5A-C, and E). Kidney carcinoma was the only tumor that showed lower E-cad staining compared to normal tissues (Fig. 5D-E), consistent with its CDH1 mRNA expression profile (Fig. 1C). These immunohistochemistry staining results are also consistent with the E-cad protein expression profiles revealed by the mass spectrometry analysis, which demonstrated that E-cad expression was upregulated or remained unchanged in most carcinoma tissues (Fig. 4A-B) and was expressed at higher levels in most of the carcinoma cell lines analyzed (Table 3).

Immunohistochemistry staining reveals that E-cad protein is not downregulated in most carcinoma tissues. Immunohistochemistry staining of E-cad in breast carcinoma (A), head/neck carcinoma (B), pancreatic carcinoma (C), and kidney carcinoma (D), and their corresponding normal tissues. E, Most carcinoma tissues exhibit medium to high intensity of E-cad staining except for kidney carcinoma, which exhibits little or no E-cad staining in most of the tumor tissues
E-cad protein is not downregulated when most primary carcinomas progress to metastatic tumors
Since the E-cad protein is postulated to play a critical role during the transition from primary tumors to metastatic tumors in carcinomas [12, 13, 28, 53, 54], we assessed how the E-cad levels differed in the cell lines derived from metastatic tumors compared to those derived from primary tumors using the proteomics data on the CCLE available in the DepMap portal. The levels of E-cad were not significantly different between the metastatic cell lines and primary cells lines in any of the specific lineages (Fig. 6A), nor were they significantly different when all the metastatic cell lines were compared to all the primary tumor cell lines encompassing all 9 lineages (Fig. 6B). Thus, on average, the levels of E-cad protein in metastatic carcinoma cell lines remained unchanged compared to primary tumor cell lines, suggesting that E-cad protein is not downregulated in commonly occurring carcinomas when primary tumors progress to metastatic tumors. It is also possible that the cells are able to revert back to an epithelial phenotype after settling down in the new sites due to the MET process.

E-cad protein levels are not significantly changed in the carcinoma cell lines derived from metastatic tumors compared to the carcinoma cell lines derived from primary tumors. A, E-cad protein levels are not significantly different in the carcinoma cell lines derived from metastatic tumors compared to the carcinoma cell lines derived from primary tumors of pancreatic (11 metastatic (M); 7 primary (P)), lung (49 M; 30 P), ovarian (8 M; 9 P), colorectal (9 M; 17 P), head/neck (4 M; 6 P), endometrial (2 M; 11 P), and breast (16 M; 14 P) carcinomas. B, The E-cad level in the combined metastatic carcinoma cell lines was not statistically different from the level in the combined primary tumor cell lines (101 M; 117 P)
CDH1 mRNA and E-cad protein are indeed downregulated in certain cancers
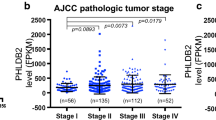
Suppression of CDH1 gene expression in cancer cells has been widely reported [55,56,57,58], particularly in cancer cell line-based studies [41, 59, 60]. Indeed, we found that the levels of CDH1 mRNA and E-cad protein were downregulated in certain carcinomas, particularly kidney carcinoma (Figs. 1C and 4C). When kidney carcinoma was divided into four subtypes based on the differential expression of 500 genes and 500 microRNAs [61], CDH1 mRNA levels were significantly downregulated in cluster 2 (m2), cluster 3 (m3), and cluster 4 (m4), but remained unchanged in cluster 1 (m1) compared to normal tissues (Fig. 7A). In addition, the CDH1 mRNA levels were significantly downregulated during tumor progression in kidney tumors, with lower levels in the advanced stage 3, in which tumors have intruded into veins and lymph nodes, and the lowered levels of CDH1 mRNA persisted in stage 4, in which tumors have grown in tissues outside the kidney and in distant organs (Fig. 7B) [62]. The decreased levels of CDH1 mRNA in advanced stages coincide with the invasion of surrounding vessels and tissues, suggesting that the lower levels of CDH1 mRNA in kidney carcinoma are closely related to tumor invasion. These results are consistent with the CDH1 mRNA levels in the kidney carcinoma cell lines, which were at the lower end of the levels in the carcinoma cell lines examined (Fig. 3A). In addition, 10 out of the 14 examined kidney carcinoma cell lines (71.4%) exhibited low or no detection of CDH1 mRNA, and only 4 out of the 14 (28.6%) kidney carcinoma cell lines exhibited moderate to high levels of CDH1 mRNA (Fig. 7C and Table 1).

CDH1 mRNA is downregulated in kidney tumors and the downregulation potentially results from promoter methylation. A, The CDH1 mRNA levels are significantly decreased in 3 of the 4 subtypes of kidney cancer: m2 (n = 85), m3 (n = 93), and m4 (n = 85) and remain unchanged in m1 (n = 145) compared to the normal tissues (n = 100) (grey, normal; red, carcinoma). B, CDH1 mRNA is significantly downregulated across the major stages of kidney carcinoma with a decrease in CDH1 mRNA between stage 2 and stage 3 (p-value = 0.00025). C, Most kidney carcinoma cell lines exhibit low or no CDH1 mRNA expression. D, CDH1 promoter methylation in kidney carcinoma (n = 324) is significantly higher than in the normal tissues (n = 160). E, CDH1 promoter methylation is significantly increased in stage 1 (n = 160), stage 2 (n = 31), stage 3 (n = 73), and stage 4 (n = 58) compared to the normal tissues (n = 160). F, E-cad protein expression is significantly decreased in stage 1 (n = 52), stage 2 (n = 13), stage 3 (n = 33), and stage 4 (n = 12) of kidney carcinoma compared to the normal tissues (n = 84). G, E-cad protein expression is significantly decreased in stage 1 (n = 4), stage 2 (n = 85), stage 3 (n = 35), and stage 4 (n = 7) of pancreatic cancer compared to the normal tissues (n = 74). *, p < 0.05, **, p < 0.01
Promoter methylation is a major mechanism in suppressing tumor suppressor gene expression [55,56,57,58]. We examined the CDH1 promoter methylation using the UALCAN web server (http://ualcan.path.uab.edu/analysis.html). CDH1 promoter methylation was significantly higher in kidney primary tumors than in normal tissues (Fig. 7D). In addition, CDH1 promoter methylation is significantly elevated in all stages of kidney carcinoma compared to normal samples (Fig. 7E). These results demonstrate that promoter methylation plays a critical role in suppressing the transcription of the CDH1 gene in kidney carcinoma.
Consistent with the lower levels of CDH1 mRNAs in kidney carcinoma (Fig. 7A-C), analysis using the UALCAN web server revealed that the levels of E-cad protein in kidney cancer were significantly downregulated in tumors compared to normal tissue (Fig. 4C) and E-cad was not detectable in the immunohistochemistry staining of kidney carcinoma (Fig. 5D), demonstrating that the lower levels of CDH1 mRNA result in lower levels of E-cad protein [37]. This relationship suggests that the expression of E-cad protein in kidney carcinoma is regulated at the transcription level, most likely due to CpG methylation (Fig. 7D-E), allelic deletion of 16q22.1 containing the E-cad locus, or nonsense mutations [14]. E-cad protein levels were also significantly decreased in all stages of kidney carcinoma compared to normal tissue (Fig. 7F), suggesting that the loss of E-cad expression occurs initially in the early stages of tumor development and is maintained through later stages.
Decreased levels of E-cad protein were also detected in pancreatic carcinoma (Fig. 4C). The downregulation of E-cad protein was observed in all stages of pancreatic carcinoma compared to the normal tissue (Fig. 7G). These results are intriguing because transcriptomics data revealed upregulation of CDH1 mRNA in pancreatic carcinoma tissues compared to normal tissue (Fig. 1A) and higher CDH1 mRNA levels in pancreatic cancer cell lines compared to other carcinoma cell lines (Fig. 3A). The E-cad immunohistochemistry staining also revealed that the E-cad protein was expressed at elevated levels in all the pancreatic carcinoma samples analyzed (Fig. 5C). The conflicting results warrant more careful examinations of CDH1 mRNA levels and E-cad protein levels in pancreatic cancer tissues and cell lines in the future.
CDH1 mRNA levels and E-cad protein levels in carcinomas are positively correlated, and the CDH1 mRNA levels are correlated to cancer patient’s survival
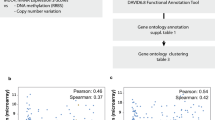
The relationship between the levels of CDH1 mRNA and E-cad protein in the carcinoma cell lines from nine different lineages stored on DepMap portal was analyzed using Pearson correlation analysis and Spearman correlation analysis. CDH1 mRNA levels were strongly positively correlated with E-cad protein levels for eight of the nine lineages of carcinoma cell lines (Table 4). The exception was the kidney carcinoma lineage, in which the cell lines exhibited a weak correlation between CDH1 mRNA levels and E-cad protein levels (Table 4). When all the cell lines were analyzed together, CDH1 mRNA levels and E-cad levels exhibited a strong positive correlation (Fig. 8A). These results suggest that, at least at the cell line level, the expression of E-cad is regulated at the transcriptional level in most carcinoma cell types. To determine whether the same observation can be made at the tissue level, a database with proteomics and transcriptomics analyses of carcinoma tissue samples would be beneficial.

Positive correlation between CDH1 mRNA levels and E-cad protein levels in carcinoma cell lines. A, The CDH1 mRNA levels and E-cad protein levels in carcinoma cell lines are positively correlated (R = 0.845). CDH1 mRNA and E-cad protein data from tumor cell lines derived from the endometrium (n = 14), head/neck (n = 10), pancreas (n = 19), ovary (n = 17), lung (n = 77), liver (14), kidney (n = 12), colon (n = 29), and breast (n = 30) were used in the plot. CDH1 mRNA and E-cad data were obtained from Expression 22Q1 Public and Proteomics datasets in the DepMap portal (https://depmap.org/portal/). B and C, OS and DFS of breast, colon, lung, pancreatic, ovarian, and endometrial carcinoma patients who carry tumors with upregulated CDH1 mRNA (High = 1,298; Low = 1,298) based on CDH1 mRNA expression. C and D, OS and DFS of kidney carcinoma patients who carry tumors with downregulated CDH1 (High = 258; Low = 258) based on CDH1 mRNA expression. F and G, OS and DFS of liver and head/neck carcinoma patients who carry tumors with the unchanged expression of CDH1 (High = 441; Low = 441) based on CDH1 mRNA expression
We performed cancer patient survival analysis to assess whether the CDH1 mRNA expression is correlated to cancer patient’s survival. For the patients with carcinomas that had upregulated CDH1 mRNA, including endometrial, pancreatic, ovarian, lung, colon, and breast carcinomas (Fig. 1A), cancer patients with tumors that expressed higher levels of CDH1 mRNA (n = 1,298) fared significantly better than those with tumors that expressed lower levels of CDH1 mRNA (n = 1,298) with better OS and DFS (p = 0.0018 and 1.3E-09, respectively) (Fig. 8B-C). For the patients with carcinomas that had downregulated CDH1 mRNA, including kidney cancer (Fig. 1C), CDH1 mRNA expression had a significant effect on OS but not DFS (High, n = 258 vs Low, n = 258; p = 3.7E-06 and 0.11, respectively) (Fig. 8D-E). For the patients diagnosed with carcinomas that had unchanged levels of CDH1 mRNA compared to normal tissues, including liver and head/neck carcinomas (Fig. 1B), tumor CDH1 mRNA levels were not significantly correlated to OS but were significantly correlated to DFS (High, n = 441 vs Low, n = 441; p, = 0.56 and 1.2E-05, respectively) (Fig. 8F-G). Overall, these results demonstrate that higher levels of CDH1 mRNA expression are correlated with better survival of carcinoma patients.
Discussion
The loss of E-cad has been widely considered a hallmark of metastatic cancers and critical for metastasizing tumor cells to break away from the epithelial tissues to invade the tumor stroma [12,13,14]. This observation was established primarily with the help of invasive lobular breast cancer (ILC) tissues, in which the loss of E-cad has been shown to play a key role [14, 50,51,52]. Our analysis of clinical cancer tissues revealed that CDH1/E-cad expression was downregulated only in a few types or subtypes of tumors among the large group of tumor types or subtypes examined; ILC happens to be a subtype of breast tumors in which E-cad expression was downregulated (Fig. 4F). Another major exception is kidney carcinoma, which exhibited the well-described loss of E-cad expression (Figs. 1C and 7). In agreement with recent debates on the role of E-cad in tumor progression and metastasis [63, 64], our analysis demonstrates that CDH1 mRNA and E-cad protein are not downregulated in the majority of carcinomas (Figs. 1, 4, and 5) or during tumor progression in most carcinomas (Figs. 2 and 4E, and Supplementary Figs. 1 and 2—Additional file 1). For a more detailed analysis of the role of E-cad in EMT in tumor progression, tumor samples exhibiting hybrid EMT or partial EMT [65,66,67,68] may be required. Because it is difficult to obtain data on the hybrid EMT or partial EMT samples from cancer patients, at least in large quantities, our studies cannot provide insight into the role of E-cad in complex phenomena like hybrid EMT or partial EMT in metastasis, which shows a limitation of this type of study.
It is interesting to note that CDH1 mRNA and/or E-Cad were upregulated in most cancers in the early stages of tumor development and the levels remained elevated as tumors progressed to later stages across most carcinoma types (Figs. 2 and 4E, and Supplementary Figs. 1 and 2—Additional file 1). These results suggest that most carcinomas may require higher levels of E-cad expression for tumor formation and tumor progression in earlier stages of tumor development, and this requirement needs to be maintained even after metastasis has occurred. One possibility is that the upregulation of CDH1/E-cad expression in carcinoma cells is an adaptive response to the abnormal signaling inside tumor cells, which is known to result in increasingly altered cell–cell adhesion and actin cytoskeleton rearrangement during tumor formation, progression, and invasion [69,70,71]. For example, it has been shown that tumor cells can upregulate proteins that are directly related to the rearrangement of the actin cytoskeleton [72, 73] and that there is rearrangement (but not loss) of E-cad-based adherens junctions during neoplastic transformation [69]. Tumor cells may respond to these types of changes in cell–cell adhesion and actin cytoskeleton rearrangement by expressing more E-cad to restore the altered cell–cell adhesion and epithelial tissue integrity during tumor formation, progression, and invasion. It has also been shown that E-cad plays an important role in preventing anoikis, the induction of apoptosis after the loss of attachment to the ECM and neighboring cells [71, 74]. To prevent anoikis induced by truncation of the cytoplasmic domain of E-cad which results in disruption of the binding of the domain to β-catenin, a linker protein that connects the actin cytoskeleton to the cytoplasmic domain of E-cad [74, 75], tumor cells may be required to upregulate E-cad [76, 77].
Since CDH1/E-cad upregulation is widespread in carcinomas (Figs. 1, 4, and 5) and the levels remain elevated as tumors progressed to later stages across most carcinoma types (Figs. 2 and 4E, and Supplementary Figs. 1 and 2—Additional file 1), the effect of higher levels of CDH1 mRNA on carcinoma patient’s survival (Fig. 8) suggest that the role of E-cad on carcinoma development and progression is more complex than previously thought and warrants further investigation. Although the Log-rank tests allowed us to establish a positive correlation between CDH1 mRNA levels and cancer patient’s survival (Fig. 8), the survival tests did not allow us to conclude whether CDH1 expression is functionally linked to cancer patient’s survival, showing another limitation of this study. To gain further insight into the potential value of CDH1 mRNA levels in cancer prognosis and the role of CDH1/E-cad in carcinoma development and progression, future studies should consider additional clinical data, such as median survival time, age, and tumor stages. In addition, since CDH1 mRNA is markedly upregulated in some types of tumors, such as colon and endometrial carcinoma (Fig. 1A), from the early stages of tumor development (Fig. 4E, and Supplementary Fig. 2A-C—Additional file 1), it is worth further investigation to determine whether CDH1 mRNA levels can serve as a reliable biomarker for early diagnosis of these carcinomas.
It is well established that metastatic carcinoma cells invade the stroma and migrate in single cells or collectively in groups [78]. In single-cell invasion/migration, single cells acquire the ability to break away from the primary tumor tissues through the loss of E-cad [79, 80]. In contrast, in collective cell invasion/migration, most of the tumor cells localized in the interior of a cell cluster maintain elevated levels of E-cad expression and only the tumor cells on the edge of the cluster express low levels of E-cad, which allow the cluster of cells to break away from the primary carcinoma tissues [23, 27, 81, 82]. Most previous studies designed to investigate tumor EMT and metastasis normally used in-vitro 2-dimensional cell culture (2D) or 3D scaffold cell culture with a focus on single-cell invasion/migration [53, 59, 83, 84]. Results from these types of studies may not reflect the situation in the collective cell invasion/migration [63, 64, 85, 86]. Our findings that CDH1/E-cad expression is not significantly downregulated when primary tumors progress into metastatic tumors (Fig. 6), which are consistent with the observations from other groups [13, 20, 28, 29], suggest that single-cell invasion/migration may not be the preferred mode of invasion/migration, and collective invasion/migration might be the predominant form of invasion/migration for most carcinomas, a notion that is supported by several studies monitoring metastatic tumors in circulation [81, 87, 88]. Furthermore, after metastatic carcinoma cells settle down in a new place, the metastatic carcinoma cells re-acquire epithelial cell phenotypes via MET [89, 90]. It is also possible that MET can contribute to the elevated or unchanged levels of E-cad in metastatic cancer cells.
Conclusion
CDH1 mRNA and E-cadherin protein are not downregulated in most carcinoma tissues and carcinoma cell lines tested in this study. Thus, the role of E-cad in tumor progression and metastasis may have previously been oversimplified. Because CDH1 mRNA is markedly upregulated in the early stages of tumor development of some types of tumors, such as colon and endometrial carcinomas, CDH1 mRNA levels may serve as a reliable biomarker for the early diagnosis of these carcinomas.
Availability of data and materials
The datasets analyzed during the current study are available in the Gene Expression Profiling Interactive Analysis 2 (GEPIA2) (http://gepia2.cancer-pku.cn/#index), the University of Alabama at Birmingham Cancer Data Analysis (UALCAN) portal.
(http://ualcan.path.uab.edu/analysis-prot.html and http://ualcan.path.uab.edu/analysis.html), the Human Protein Atlas (HPA) (https://www.proteinatlas.org/), and the Cancer Dependency Map (DepMap) portal (https://depmap.org/portal/interactive/).
Abbreviations
- CCLE:
-
Cancer Cell Line Encyclopedia
- DepMap:
-
Cancer Dependency Map
- E-cad:
-
E-cadherin
- DFS:
-
Disease-free survival
- OS:
-
Overall survival
- ECM:
-
Extracellular matrix
- EMT:
-
Epithelial-mesenchymal transition
- GEPIA2:
-
Gene Expression Profiling Interactive Analysis 2
- GTEx:
-
Genome-Typing Expression
- TPM:
-
Transcripts per million
- TCGA:
-
The Cancer Genome Atlas
- UALCAN:
-
The University of Alabama at Birmingham Cancer Data Analysis Portal
References
Ganz T. Epithelia: not just physical barriers. Proc Natl Acad Sci U S A. 2002;99(6):3357–8.
Gloushankova NA, Rubtsova SN, Zhitnyak IY. Cadherin-mediated cell-cell interactions in normal and cancer cells. Tissue Barriers. 2017;5(3):e1356900.
Coopman P, Djiane A. Adherens Junction and E-Cadherin complex regulation by epithelial polarity. Cell Mol Life Sci. 2016;73(18):3535–53.
Boggon TJ, Murray J, Chappuis-Flament S, Wong E, Gumbiner BM, Shapiro L. C-cadherin ectodomain structure and implications for cell adhesion mechanisms. Science. 2002;296(5571):1308–13.
Takeichi M. The cadherins: cell-cell adhesion molecules controlling animal morphogenesis. Development. 1988;102(4):639–55.
Halbleib JM, Nelson WJ. Cadherins in development: cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 2006;20(23):3199–214.
Overduin M, Harvey TS, Bagby S, Tong KI, Yau P, Takeichi M, Ikura M. Solution structure of the epithelial cadherin domain responsible for selective cell adhesion. Science. 1995;267(5196):386–9.
Parisini E, Higgins JM, Liu JH, Brenner MB, Wang JH. The crystal structure of human E-cadherin domains 1 and 2, and comparison with other cadherins in the context of adhesion mechanism. J Mol Biol. 2007;373(2):401–11.
Cailliez F, Lavery R. Cadherin mechanics and complexation: the importance of calcium binding. Biophys J. 2005;89(6):3895–903.
Le Bras GF, Taubenslag KJ, Andl CD. The regulation of cell-cell adhesion during epithelial-mesenchymal transition, motility and tumor progression. Cell Adh Migr. 2012;6(4):365–73.
Yamada S, Pokutta S, Drees F, Weis WI, Nelson WJ. Deconstructing the cadherin-catenin-actin complex. Cell. 2005;123(5):889–901.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74.
Lambert AW, Pattabiraman DR, Weinberg RA. Emerging Biological Principles of Metastasis. Cell. 2017;168(4):670–91.
Berx G, Cleton-Jansen AM, Nollet F, de Leeuw WJ, van de Vijver M, Cornelisse C, van Roy F. E-cadherin is a tumour/invasion suppressor gene mutated in human lobular breast cancers. EMBO J. 1995;14(24):6107–15.
Liu J, Sun X, Qin S, Wang H, Du N, Li Y, Pang Y, Wang C, Xu C, Ren H. CDH1 promoter methylation correlates with decreased gene expression and poor prognosis in patients with breast cancer. Oncol Lett. 2016;11(4):2635–43.
Pecina-Slaus N. Tumor suppressor gene E-cadherin and its role in normal and malignant cells. Cancer Cell Int. 2003;3(1):17.
Mendonsa AM, Na TY, Gumbiner BM. E-cadherin in contact inhibition and cancer. Oncogene. 2018;37(35):4769–80.
Yu W, Yang L, Li T, Zhang Y. Cadherin Signaling in Cancer: Its Functions and Role as a Therapeutic Target. Front Oncol. 2019;9:989.
Onder TT, Gupta PB, Mani SA, Yang J, Lander ES, Weinberg RA. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Res. 2008;68(10):3645–54.
Jolly MK, Ware KE, Gilja S, Somarelli JA, Levine H. EMT and MET: necessary or permissive for metastasis? Mol Oncol. 2017;11(7):755–69.
Grigore AD, Jolly MK, Jia D, Farach-Carson MC, Levine H. Tumor Budding: The Name is EMT Partial EMT. J Clin Med. 2016;5(5):51.
Lou Y, Preobrazhenska O. auf dem Keller U, Sutcliffe M, Barclay L, McDonald PC, Roskelley C, Overall CM, Dedhar S: Epithelial-mesenchymal transition (EMT) is not sufficient for spontaneous murine breast cancer metastasis. Dev Dyn. 2008;237(10):2755–68.
Fischer KR, Durrans A, Lee S, Sheng J, Li F, Wong ST, Choi H, El Rayes T, Ryu S, Troeger J, et al. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature. 2015;527(7579):472–6.
Shamir ER, Pappalardo E, Jorgens DM, Coutinho K, Tsai WT, Aziz K, Auer M, Tran PT, Bader JS, Ewald AJ. Twist1-induced dissemination preserves epithelial identity and requires E-cadherin. J Cell Biol. 2014;204(5):839–56.
Zheng X, Carstens JL, Kim J, Scheible M, Kaye J, Sugimoto H, Wu CC, LeBleu VS, Kalluri R. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature. 2015;527(7579):525–30.
Bukholm IK, Nesland JM, Karesen R, Jacobsen U, Borresen-Dale AL. E-cadherin and alpha-, beta-, and gamma-catenin protein expression in relation to metastasis in human breast carcinoma. J Pathol. 1998;185(3):262–6.
Park SY, Jang H, Kim SY, Kim D, Park Y, Kee SH. Expression of E-Cadherin in Epithelial cancer cells Increases cell motility and directionality through the localization of ZO-1 during collective cell migration. Bioengineering (Basel). 2021;8(5):65.
Yang J, Weinberg RA. Epithelial-mesenchymal transition: at the crossroads of development and tumor metastasis. Dev Cell. 2008;14(6):818–29.
Petrova YI, Schecterson L, Gumbiner BM. Roles for E-cadherin cell surface regulation in cancer. Mol Biol Cell. 2016;27(21):3233–44.
Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–102.
Tang Z, Kang B, Li C, Chen T, Zhang Z. GEPIA2: an enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019;47(W1):W556–60.
Consortium GT. The Genotype-Tissue Expression (GTEx) project. Nat Genet. 2013;45(6):580–5.
Ghandi M, Huang FW, Jane-Valbuena J, Kryukov GV, Lo CC, McDonald ER 3rd, Barretina J, Gelfand ET, Bielski CM, Li H, et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature. 2019;569(7757):503–8.
Barretina J, Caponigro G, Stransky N, Venkatesan K, Margolin AA, Kim S, Wilson CJ, Lehar J, Kryukov GV, Sonkin D, et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature. 2012;483(7391):603–7.
Chandrashekar DS, Bashel B, Balasubramanya SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi B, Varambally S. UALCAN: a portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia. 2017;19(8):649–58.
Chandrashekar DS, Karthikeyan SK, Korla PK, Patel H, Shovon AR, Athar M, Netto GJ, Qin ZS, Kumar S, Manne U, et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia. 2022;25:18–27.
Chen F, Chandrashekar DS, Varambally S, Creighton CJ. Pan-cancer molecular subtypes revealed by mass-spectrometry-based proteomic characterization of more than 500 human cancers. Nat Commun. 2019;10(1):5679.
Nusinow DP, Szpyt J, Ghandi M, Rose CM, McDonald ER 3rd, Kalocsay M, Jane-Valbuena J, Gelfand E, Schweppe DK, Jedrychowski M, et al. Quantitative proteomics of the cancer cell line encyclopedia. Cell. 2020;180(2):387-402 e316.
Uhlen M, Bjorling E, Agaton C, Szigyarto CA, Amini B, Andersen E, Andersson AC, Angelidou P, Asplund A, Asplund C, et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol Cell Proteomics. 2005;4(12):1920–32.
Monsivais D, Vasquez YM, Chen F, Zhang Y, Chandrashekar DS, Faver JC, Masand RP, Scheurer ME, Varambally S, Matzuk MM, et al. Mass-spectrometry-based proteomic correlates of grade and stage reveal pathways and kinases associated with aggressive human cancers. Oncogene. 2021;40(11):2081–95.
Wendt MK, Taylor MA, Schiemann BJ, Schiemann WP. Down-regulation of epithelial cadherin is required to initiate metastatic outgrowth of breast cancer. Mol Biol Cell. 2011;22(14):2423–35.
Drees F, Pokutta S, Yamada S, Nelson WJ, Weis WI. Alpha-catenin is a molecular switch that binds E-cadherin-beta-catenin and regulates actin-filament assembly. Cell. 2005;123(5):903–15.
Campbell K, Casanova J. A role for E-cadherin in ensuring cohesive migration of a heterogeneous population of non-epithelial cells. Nat Commun. 2015;6:7998.
Koussounadis A, Langdon SP, Um IH, Harrison DJ, Smith VA. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Sci Rep. 2015;5:10775.
Hanna J, Guerra-Moreno A, Ang J, Micoogullari Y. Protein degradation and the pathologic basis of disease. Am J Pathol. 2019;189(1):94–103.
Ramazi S, Zahiri J: Posttranslational modifications in proteins: resources, tools and prediction methods. Database (Oxford) 2021, 2021:baab012
Vaklavas C, Blume SW, Grizzle WE. Translational dysregulation in cancer: molecular insights and potential clinical applications in biomarker development. Front Oncol. 2017;7:158.
Van Drie JH. Protein folding, protein homeostasis, and cancer. Chin J Cancer. 2011;30(2):124–37.
Yang H, Hu HY. Sequestration of cellular interacting partners by protein aggregates: implication in a loss-of-function pathology. FEBS J. 2016;283(20):3705–17.
Gamallo C, Palacios J, Suarez A, Pizarro A, Navarro P, Quintanilla M, Cano A. Correlation of E-cadherin expression with differentiation grade and histological type in breast carcinoma. Am J Pathol. 1993;142(4):987–93.
Moll R, Mitze M, Frixen UH, Birchmeier W. Differential loss of E-cadherin expression in infiltrating ductal and lobular breast carcinomas. Am J Pathol. 1993;143(6):1731–42.
Rasbridge SA, Gillett CE, Sampson SA, Walsh FS, Millis RR. Epithelial (E-) and placental (P-) cadherin cell adhesion molecule expression in breast carcinoma. J Pathol. 1993;169(2):245–50.
Cano A, Perez-Moreno MA, Rodrigo I, Locascio A, Blanco MJ, del Barrio MG, Portillo F, Nieto MA. The transcription factor snail controls epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat Cell Biol. 2000;2(2):76–83.
Huang RY, Guilford P, Thiery JP. Early events in cell adhesion and polarity during epithelial-mesenchymal transition. J Cell Sci. 2012;125(Pt 19):4417–22.
Graff JR, Greenberg VE, Herman JG, Westra WH, Boghaert ER, Ain KB, Saji M, Zeiger MA, Zimmer SG, Baylin SB. Distinct patterns of E-cadherin CpG island methylation in papillary, follicular, Hurthle’s cell, and poorly differentiated human thyroid carcinoma. Cancer Res. 1998;58(10):2063–6.
Graff JR, Gabrielson E, Fujii H, Baylin SB, Herman JG. Methylation patterns of the E-cadherin 5’ CpG island are unstable and reflect the dynamic, heterogeneous loss of E-cadherin expression during metastatic progression. J Biol Chem. 2000;275(4):2727–32.
Nass SJ, Herman JG, Gabrielson E, Iversen PW, Parl FF, Davidson NE, Graff JR. Aberrant methylation of the estrogen receptor and E-cadherin 5’ CpG islands increases with malignant progression in human breast cancer. Cancer Res. 2000;60(16):4346–8.
Lombaerts M, van Wezel T, Philippo K, Dierssen JW, Zimmerman RM, Oosting J, van Eijk R, Eilers PH, van de Water B, Cornelisse CJ. et al. E-cadherin transcriptional downregulation by promoter methylation but not mutation is related to epithelial-to-mesenchymal transition in breast cancer cell lines. Br J Cancer. 2006, 94(5):661–671.
Raeber GP, Lutolf MP, Hubbell JA. Molecularly engineered PEG hydrogels: a novel model system for proteolytically mediated cell migration. Biophys J. 2005;89(2):1374–88.
EslamiAmirabadi H, Tuerlings M, Hollestelle A, SahebAli S, Luttge R, van Donkelaar CC, Martens JWM, den Toonder JMJ. Characterizing the invasion of different breast cancer cell lines with distinct E-cadherin status in 3D using a microfluidic system. Biomed Microdevices. 2019;21(4):101.
Cancer Genome Atlas Research N. Comprehensive molecular characterization of clear cell renal cell carcinoma. Nature. 2013;499(7456):43–9.
Gu T, Zhao X. Integrating multi-platform genomic datasets for kidney renal clear cell carcinoma subtyping using stacked denoising autoencoders. Sci Rep. 2019;9(1):16668.
Cheung KJ, Ewald AJ. A collective route to metastasis: Seeding by tumor cell clusters. Science. 2016;352(6282):167–9.
Friedl P, Gilmour D. Collective cell migration in morphogenesis, regeneration and cancer. Nat Rev Mol Cell Biol. 2009;10(7):445–57.
Brabletz S, Schuhwerk H, Brabletz T, Stemmler MP. Dynamic EMT: a multi-tool for tumor progression. EMBO J. 2021;40(18): e108647.
Nieto MA, Huang RY, Jackson RA, Thiery JP. Emt: 2016. Cell. 2016;166(1):21–45.
Pastushenko I, Blanpain C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019;29(3):212–26.
Yang J, Antin P, Berx G, Blanpain C, Brabletz T, Bronner M, Campbell K, Cano A, Casanova J, Christofori G, et al. Guidelines and definitions for research on epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2020;21(6):341–52.
Ayollo DV, Zhitnyak IY, Vasiliev JM, Gloushankova NA. Rearrangements of the actin cytoskeleton and E-cadherin-based adherens junctions caused by neoplasic transformation change cell-cell interactions. PLoS One. 2009;4(11):e8027.
Padmanaban V, Krol I, Suhail Y, Szczerba BM, Aceto N, Bader JS, Ewald AJ. E-cadherin is required for metastasis in multiple models of breast cancer. Nature. 2019;573(7774):439–44.
Na TY, Schecterson L, Mendonsa AM, Gumbiner BM. The functional activity of E-cadherin controls tumor cell metastasis at multiple steps. Proc Natl Acad Sci U S A. 2020;117(11):5931–7.
Yamaguchi H, Condeelis J. Regulation of the actin cytoskeleton in cancer cell migration and invasion. Biochim Biophys Acta. 2007;1773(5):642–52.
Barkan D, El Touny LH, Michalowski AM, Smith JA, Chu I, Davis AS, Webster JD, Hoover S, Simpson RM, Gauldie J, et al. Metastatic growth from dormant cells induced by a col-I-enriched fibrotic environment. Cancer Res. 2010;70(14):5706–16.
Vallorosi CJ, Day KC, Zhao X, Rashid MG, Rubin MA, Johnson KR, Wheelock MJ, Day ML. Truncation of the beta-catenin binding domain of E-cadherin precedes epithelial apoptosis during prostate and mammary involution. J Biol Chem. 2000;275(5):3328–34.
Fouquet S, Lugo-Martinez VH, Faussat AM, Renaud F, Cardot P, Chambaz J, Pincon-Raymond M, Thenet S. Early loss of E-cadherin from cell-cell contacts is involved in the onset of Anoikis in enterocytes. J Biol Chem. 2004;279(41):43061–9.
Masszi A, Di Ciano C, Sirokmany G, Arthur WT, Rotstein OD, Wang J, McCulloch CA, Rosivall L, Mucsi I, Kapus A. Central role for Rho in TGF-beta1-induced alpha-smooth muscle actin expression during epithelial-mesenchymal transition. Am J Physiol Renal Physiol. 2003;284(5):F911-924.
Tian YC, Fraser D, Attisano L, Phillips AO. TGF-beta1-mediated alterations of renal proximal tubular epithelial cell phenotype. Am J Physiol Renal Physiol. 2003;285(1):F130-142.
Krakhmal NV, Zavyalova MV, Denisov EV, Vtorushin SV, Perelmuter VM. Cancer Invasion: Patterns and Mechanisms. Acta Naturae. 2015;7(2):17–28.
Naber HP, Drabsch Y, Snaar-Jagalska BE, ten Dijke P, van Laar T. Snail and Slug, key regulators of TGF-beta-induced EMT, are sufficient for the induction of single-cell invasion. Biochem Biophys Res Commun. 2013;435(1):58–63.
Lintz M, Munoz A, Reinhart-King CA: The Mechanics of Single Cell and Collective Migration of Tumor Cells. J Biomech Eng. 2017;139(2):021005–9.
Aceto N, Bardia A, Miyamoto DT, Donaldson MC, Wittner BS, Spencer JA, Yu M, Pely A, Engstrom A, Zhu H, et al. Circulating tumor cell clusters are oligoclonal precursors of breast cancer metastasis. Cell. 2014;158(5):1110–22.
Hou JM, Krebs MG, Lancashire L, Sloane R, Backen A, Swain RK, Priest LJ, Greystoke A, Zhou C, Morris K, et al. Clinical significance and molecular characteristics of circulating tumor cells and circulating tumor microemboli in patients with small-cell lung cancer. J Clin Oncol. 2012;30(5):525–32.
Peela N, Sam FS, Christenson W, Truong D, Watson AW, Mouneimne G, Ros R, Nikkhah M. A three dimensional micropatterned tumor model for breast cancer cell migration studies. Biomaterials. 2016;81:72–83.
Batlle E, Sancho E, Franci C, Dominguez D, Monfar M, Baulida J. Garcia De Herreros A: The transcription factor snail is a repressor of E-cadherin gene expression in epithelial tumour cells. Nat Cell Biol. 2000;2(2):84–9.
Nabeshima K, Inoue T, Shimao Y, Kataoka H, Koono M. Cohort migration of carcinoma cells: differentiated colorectal carcinoma cells move as coherent cell clusters or sheets. Histol Histopathol. 1999;14(4):1183–97.
Wang X, Enomoto A, Asai N, Kato T, Takahashi M. Collective invasion of cancer: Perspectives from pathology and development. Pathol Int. 2016;66(4):183–92.
Cheung KJ, Padmanaban V, Silvestri V, Schipper K, Cohen JD, Fairchild AN, Gorin MA, Verdone JE, Pienta KJ, Bader JS, et al. Polyclonal breast cancer metastases arise from collective dissemination of keratin 14-expressing tumor cell clusters. Proc Natl Acad Sci U S A. 2016;113(7):E854-863.
Friedl P, Locker J, Sahai E, Segall JE. Classifying collective cancer cell invasion. Nat Cell Biol. 2012;14(8):777–83.
Chao YL, Shepard CR, Wells A. Breast carcinoma cells re-express E-cadherin during mesenchymal to epithelial reverting transition. Mol Cancer. 2010;9:179.
Christiansen JJ, Rajasekaran AK. Reassessing epithelial to mesenchymal transition as a prerequisite for carcinoma invasion and metastasis. Cancer Res. 2006;66(17):8319–26.
Acknowledgements
We thank Jianhong Zhou for helpful discussion and for making part of Fig. 8.
Funding
This work was supported by the Arkansas Breast Cancer Research Program and Arkansas Biosciences Institute (to YD), and the American Dissertation Fellowship from the University of Arkansas (to BSM).
Author information
Authors and Affiliations
Contributions
BSM and YD conceptualized and developed the methodology. BSM and SA collected data/evidence. BSM analyzed the data. BSM wrote the original manuscript. BSM, SA, and YD revised and edited the manuscript. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplementary Figure 1.
CDH1 mRNAis significantly upregulated in some carcinoma subtypes and remains unchanged across major tumor stages of most carcinomas. CDH1 mRNA levels are significantly upregulated in the two subtypes of pancreatic carcinoma: classical (T=86) and basal (T=65) compared to the normal tissues (N=171) (A) and in the three subtypes of lung carcinoma: proximal inflammatory (T=78), proximal proliferative (T=58), and terminal respiratory unit (T=68) compared to the normal tissues (N=347) (B). Grey, normal; red, carcinoma in (A) and (B). The levels of CDH1 mRNA remain unchanged across the major stages of colon (C), pancreatic (D), lung (E), and endometrial (F) carcinomas. **, p < 0.01. Supplementary Figure 2. E-cad protein is either upregulated or remains unchanged in most major tumor stages. E-cad levels are either significantly upregulated or remain unchanged in distinct stages of endometrial carcinoma (A), lung carcinoma (B), ovarian carcinoma (C), head/neck carcinoma (D), and colon carcinoma (E and F) except for stage 1 of head/neck carcinoma (D) compared to the corresponding normal tissues. Numbers of tissues used in the analysis are: endometrial carcinoma: normal, n=31, stage 1,n=74, stage 2, n=8, stage 3, n=15; lung cancer: normal, n=11, stage 1, n=59, stage 2, n=30, stage 3, n=21; ovarian cancer: normal, n=25, stage 1, n=2, stage 3, n=75, stage 4, n=16; head/neck carcinoma: normal, n=71, stage 1, n=7, stage 2, n=25, stage 3, n=75, stage 4, n=4; colon carcinoma: normal, n=100, mucinous, n=19, non-mucinous, n=77, stage 1, n=10, stage 2, n=39, stage 3, n=40, stage 4, n=8. *, p < 0.05, **, p < 0.01.
Additional file 2: Supplementary Table 1.
List of carcinoma cell lines used for CDH1 mRNA expression analysis in Table 1. Supplementary Table 2. List of carcinoma cell lines used for E-cad expression analysis in Table 3. Supplementary Table 3. List of carcinoma cell lines used for Pearson analysis and Spearman analysis in Table 4. Supplementary Table 4. List of cancer cell lines used for CDH1 mRNA expression analysis across different types of cancers in Fig. 3A. Supplementary Table 5. List of carcinoma cell lines used for E-cad proteomics analysis comparing metastic vs primary tumor derived cells lines in Fig. 6. Supplementary Table 6. List of carcinoma cell lines used for the E-cad and CDH1 mRNA correlation analysis in Fig. 8A.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Sicairos, B., Alam, S. & Du, Y. A comprehensive analysis of different types of databases reveals that CDH1 mRNA and E-cadherin protein are not downregulated in most carcinoma tissues and carcinoma cell lines. BMC Cancer 23, 441 (2023). https://doi.org/10.1186/s12885-023-10916-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-023-10916-0