
Abstract
Background
Ichang papeda (Citrus ichangensis), a wild perennial plant of the Rutaceae family, is a cold-hardy plant. WRKY transcription factors are crucial regulators of plant growth and development as well as abiotic stress responses. However, the WRKY genes in C. ichangensis (CiWRKY) and their expression patterns under cold stress have not been thoroughly investigated, hindering our understanding of their role in cold tolerance.
Results
In this study, a total of 52 CiWRKY genes identified in the genome of C. ichangensis were classified into three main groups and five subgroups based on phylogenetic analysis. Comprehensive analyses of motif features, conserved domains, and gene structures were performed. Segmental duplication plays a significant role in the CiWRKY gene family expansion. Cis-acting element analysis revealed the presence of various stress-responsive elements in the promoters of the majority of CiWRKYs. Gene ontology (GO) analysis and protein-protein interaction predictions indicate that the CiWRKYs exhibit crucial roles in regulation of both development and stress response. Expression profiling analysis demonstrates that 14 CiWRKYs were substantially induced under cold stress. Virus-induced gene silencing (VIGS) assay confirmed that CiWRKY31, one of the cold-induced WRKYs, functions positively in regulation of cold tolerance.
Conclusion
Sequence and protein properties of CiWRKYs were systematically analyzed. Among the 52 CiWRKY genes 14 members exhibited cold-responsive expression patterns, and CiWRKY31 was verified to be a positive regulator of cold tolerance. These findings pave way for future investigations to understand the molecular functions of CiWRKYs in cold tolerance and contribute to unravelling WRKYs that may be used for engineering cold tolerance in citrus.
Similar content being viewed by others


Background
Citrus is an important source of human nutrition with high economic value; however, different biotic and abiotic stresses threaten production and limit its distribution [1]. Ichang papeda (Citrus ichangensis), a wild plant of the Rutaceae family, originates from the southwestern and middle-west regions of China and has a sporadic distribution at high elevations or humid conditions in mountainous regions [2]. It exhibits superior tolerance to extreme frosts and damp environmental conditions [3], making it a valuable germplasm for investigating stress response mechanisms. In this regard, identification of cold-responsive genes of C. ichangensis is of great importance to improve the stress resilience of citrus.
Fluctuations in the biotic and abiotic environment are detected and transmitted through various signaling pathways, ultimately leading to alterations in gene expression [4]. Being immobile plants cannot evade unfavorable circumstances by relocating to more suitable locations. They have to rely on particular physiological or developmental modifications to react and adapt to harsh environments [5]. The WRKY transcription factors, one of the most extensive families of regulatory proteins in plants, play crucial roles in governing the plant response to stressful conditions [6, 7]. The WRKY proteins are defined by the presence of highly conserved seven amino acid sequence (WRKYGQK), known as the WRKY domain surrounded by a C2H2 or C2HC zinc finger motif [8]. Both the peptide sequence and zinc finger motifs are essential for the sequence-specific recognition and high affinity of the WRKYs to their canonical cis-acting element W-box sequence (T)TGAC(C/T) [9].
Based on the zinc-finger motif structure and the number of WRKY domains, WRKY proteins are classified into three major groups [10]. Group I include proteins containing two WRKY domains with a C2H2 motif, whereas Group II members possess a single WRKY domain along with a C2H2 motif. In addition, the Group II proteins can be further categorized into five subgroups, IIa, IIb, IIc, IId, and IIe. Group III WRKY proteins possess a single WRKY domain and a C2CH motif.
As one of the largest families of TFs with diverse regulatory functions, the WRKY genes are closely associated with the regulation of stress responses in higher plants [11]. It has been reported that most plant WRKY proteins function as positive regulators in environmental adaptation. For example, VqWRKY31 of grapevine directly regulates STS9 and STS48, two genes involved in stilbene synthesis, to regulate powdery mildew resistance [12]. CdWRKY2 of bermudagrass has been demonstrated to play a role in regulation of sucrose biosynthesis, in conjunction with the CBF signaling pathway, to increase cold tolerance [13]. In addition, some plant WRKY proteins have been reported to function as negative regulators of stress defense. For instance, OsWRKY63 of rice negatively regulated chilling tolerance through the OsWRKY63-OsWRKY76-OsDREB1B signaling cascade [14]. In another study, overexpression of OsWRKY10 resulted in the accumulation of reactive oxygen species (ROS) in both rice chloroplasts and apoplasts [15]. Moreover, OsWRKY5 acts as a repressor against ABA-triggered drought resilience [16]. Additionally, ZmWRKY20 and ZmWRKY115 reduced the capacity of maize to tolerate high salinity by repressing ZmbZIP111 expression [17].
Since WRKY family genes play a significant role in plant stress resistance [18], structural characteristic and functions of WRKY transcription factors from various species have been identified so far. For example, 74 WRKY genes have been identified in Arabidopsis [19], 103 in rice [20], 188 in soybean [21], and 58 in maize [22]. Genome-wide characterization of WRKYs has also been conducted in horticultural plants, which revealed presence of 81 WRKY genes in tomato [23], 61 in cucumber [24], 127 in apple [25], 103 in pear [26], and 54 in pineapple [27]. However, there is limited information on the number, characteristics, evolutionary connections, and expressions of WRKY genes in C. ichangensis, which to some extent impedes our understanding on the role of WRKY in regulation of cold tolerance in Ichang papeda.
In this study, 52 WRKY genes were identified in C. ichangensis (names as CiWRKYs), which were categorized into three main groups and five subfamilies. A comprehensive analysis was conducted to characterize the exon-intron, motif composition, subcellular localization, gene duplication, chromosome distribution, and phylogenetic and synteny analyses. Gene Ontology (GO) annotations and promoter analysis were also carried out to better understand the biological functions of CiWRKYs. Moreover, the expression patterns of CiWRKYs under cold treatment were assessed using RT-qPCR, and the transcript abundance of 14 most cold-responsive genes was examined based on the RNA-seq data of C. ichangensis. Finally, the role of CiWRKY31 in cold tolerance was experimentally verified by using VIGS (virus-induced gene silencing). Taken together, this study uncovers the functional roles of WRKY genes in C. ichangensis under cold stress and provides valuable insights for exploring specific CiWRKY genes to enhance cold tolerance in Citrus.
Materials and methods
Plant materials and cold treatment
C. ichangensis seeds harvested from the Citrus Breeding Centre at Huazhong Agriculture University were sown in soil pots and grown for three months at 25 °C. For cold treatment, 3-month-old plants were placed in a low-temperature incubator set at 4 °C. The leaves were collected at specific time points (0 h, 6 h, 24 h, 72 h, and 120 h), immediately frozen in liquid nitrogen and stored at − 80 °C for further analysis.
Sequence retrieval
The Citrus ichangensis genome sequence was downloaded from the Citrus Pan-genome to Breeding Database (http://citrus.hzau.edu.cn/) [28]. The WRKY domain (PF03106) hidden Markov model file was acquired from the Pfam protein family database (http://pfam.xfam.org/) [29]. The putative CiWRKY genes were identified using the HMMER3.3.2 program (https://www.ebi.ac.uk/Tools/hmmer/search/hmmscan) [30]. Based on the filtered results from the HMMER database, all identified genes were then investigated utilizing two online software tools. SMART (http://smart.embl.de/) [31] was used to verify that the candidate CiWRKY proteins containing the conserved WRKY domains. The ExPASy website was utilized to evaluate the amino acid length, molecular weight, and isoelectric point of the protein (https://web.expasy.org/protparam/). The PSORT software was employed for the prediction of subcellular localization (https://www.genscript.com/psort.html).
Sequence analysis
The WRKY domains of the identified C. ichangensis WRKY proteins were aligned with Clustal default parameters to generate protein sequence alignments. The inferred amino acid sequences within the WRKY domains were subsequently manually refined with the GeneDoc software. The exon-intron arrangement of CiWRKY genes was obtained by comparing the predicted coding sequences (CDS) with their corresponding full-length counterparts using the online software Gene Structure Display Server (GSDS) (http://gsds.cbi.pku.edu.cn). The conserved motifs of CiWRKY proteins were examined using the MEME online software (https://meme-suite.org/meme/). The gene structure and conserved motif were illustrated utilizing the TBtools software [32].
Phylogenetic analysis
The peptide sequences of the AtWRKY proteins were obtained from The Arabidopsis Information Resource (TAIR, https://www.arabidopsis.org/). The Clustal software was used to align all conserved WRKY domains. Phylogenetic tree was constructed using the neighbor-joining (NJ) method in MEGA 7.0, with 1,000 bootstrap replications [33], and a high-quality image of the phylogenetic tree was generated by iTOL (https://itol.embl.de/).
Chromosomal distribution and gene duplication analysis
The physical location of CiWRKY genes chromosome was obtained using the Rag Tag software (Unpublished data). Gene duplication was assessed by MCScanX with default parameters using WRKY gene annotations and the whole-genome sequences of C. ichangensis and Arabidopsis thaliana, rice (Oryza Sativa), apple, and grape [34]. Tbtools software was utilized in constructing the circos plot.
Cis-acting element analysis in CiWRKYs promoters
The promoter sequences 2000 bp upstream of CiWRKY genes were obtained by utilizing the “Sequence Fetch” tool from the Citrus Pan-genome to Breeding Database. Plant CARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) was utilized to analyze the ten most representative cis-acting elements involved in stress response of the CiWRKY promoters. Visualization was performed by the TBtools software.
Protein interaction analysis and GO annotations
The program STRING 12.0 (https://cn.string-db.org/) was utilized for the prediction of protein-protein interactions, with a confidence score of > 0.4. Gene ontology (GO) annotation was performed by the phyper function in the R package and GO terms with a Q-value ≤ 0.05 were selected as overrepresented. The bubble diagram was generated using the Dr. Tom Multi-omics Data Mining System (https://biosys.bgi.com).
RNA extraction and qRT-PCR analysis
RNA was extracted from the harvested samples using a commercially available RNA extraction kit (RN33; Aidlab Biotech Co. Ltd, Beijing, China). The extracted RNA was transformed into complementary DNA (cDNA) using HiScript III RT SuperMix for qPCR (+ gDNA wiper) (Vazyme, Nanjing, China). RT-qPCR was performed on an ABI7500 system (Applied Biosystems, Foster City, CA, USA) using AceQ SYBR Green Master Mix (Vazyme, Nanjing, China). The thermocycling conditions were 95 °C for 5 min, followed by 40 cycles of 95 °C for 10 min, 58 °C for 30 s, and 95 °C for 30 s. The 10 µl reaction mixture consisted of 5 µl of 2×SYBR Green PCR Master Mix, 0.2 µl of 10 mM primers, and 200 ng of cDNA. Three replicates were performed for each sample. ACTIN was used as an internal reference gene, and the relative expression level was calculated using the 2-▵▵CT method [35]. The primer sequences used in this study are listed in Table S8. Expression heatmap visualization was performed by the TBtools software. Transcript profile of cold-responsive CiWRKY genes was carried out from an earlier transcriptome dataset [36].
Virus-Induced Gene Silencing (VIGS)
A 300-bp partial segment of CiWRKY31 was amplified using primers containing BamHI and SmaI restriction sites, then ligated to the pTRV2 vector (tobacco rattle virus 2) to create the pTRV2-CiWRKY31 construct. The plasmid was introduced into the Agrobacterium tumefaciens strain GV3101 by heat shock. The fusion construct (pTRV2-CiWRKY31) and control vectors (pTRV1 or pTRV2) were separately mixed (construction: control; 1:1) and then infiltrated into 1-month-old Ichang papeda seedlings [37]. After 3 d of growth in darkness at 25 °C, the infiltrated plants were transferred to soil and grew for 1 month. Genomic PCR was conducted to detect positive transgenic lines, while RT-qPCR was utilized to assess the transcript levels of CiWRKY31 in the VIGS plant.
Cold tolerance assay of VIGS plants and physiological measurements
The VIGS and control plants were treated at − 4 °C for 8 h and leaves were collected for further analysis of physiological indexes. Electrolyte leakage was measured following the method described in a previous study [38]. Chlorophyll fluorescence was assessed utilizing an IMAGING-PAM chlorophyll fluorometer (Walz, Germany), and the Fv/Fm ratio was calculated with the Imaging Win Gege software.
Statistical analysis
The experiments were performed using a completely randomized design. Statistical differences were analyzed using the Statistical Package for the Social Sciences (SPSS) software with a one-way Analysis of variance (ANOVA) method based on the LSD test, taking P < 0.05 (*), P < 0.01 (**), and P < 0.001(***) as statistically significant.
Results
Identification of WRKYs in C. ichangensis
A total of 52 putative CiWRKY genes were obtained by HMM search against the Citrus ichangensis genome. SMART software tools were then used to ensure the accuracy and completeness of the HMMER search results. The 52 gene models were handpicked and designated as CiWRKY genes owing to the verification of fully intact WRKY domains. The CiWRKYs were then annotated and renamed (Table S1) through comparative analyses by blasting against the peptide sequences of Arabidopsis WRKY genes [18]. Their CDS ranged from 240 bp to 2736 bp, with corresponding polypeptide between 79 and 911 amino acids. The theoretical isoelectric points of the CiWRKY proteins varied between 5.01 and 9.83, and their molecular weights ranged from 8.87 kDa to 10.2 kDa. Furthermore, subcellular localization prediction suggested that all the CiWRKY proteins are located in the nucleus, in line with their typical function as transcription factors.
Multiple sequence alignment, classification and phylogenetic analysis of CiWRKYs
To gain insight into the properties of the CiWRKYs, multiple sequence alignments were conducted to assess the structural features of CiWRKY proteins. A fragment of 60 amino acids containing the WRKY domains in each protein was selected for analysis (Fig. S1). Specifically, a total of 50 members were found to possess the WRKYGK domains, indicating high degree of sequence conservation among the CiWRKYs. However, it is important to note that CiWRKY50 showed a slight variation in a single amino acid. In addition, the protein sequence of CiWRKY52 was shorter and did not exhibit a precise alignment with that of other CiWRKY members.
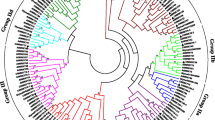
Then, based on the category of Arabidopsis WRKY proteins, all CiWRKY members were classified into three groups (Group I, II and III) (Fig. 1A). Ten proteins possessing two WRKY domains was categorized into Group I, while seven CiWRKY proteins were classified in Group III. Group II comprised 35 CiWRKY proteins and was further classified into five subgroups, with 3, 8, 14, 5, and 5 members in subgroups IIa-IIe, respectively (Fig. 1B). Next, a phylogenetic tree was constructed using the protein sequences of CiWRKYs and AtWRKYs (Fig. 1C), which better illustrates the evolutionary relationship among the WRKY members in the two species.

Classification and phylogenetic analysis of CiWRKY genes. (A-B) Proportion (A) and number (B) of Citrus ichangensis WRKY genes in each group or subgroup. (C) The phylogenetic tree between CiWRKYs and AtWRKYs, created using MEGA by the Neighbor-Joining (NJ) method with 1,000 bootstrap replicates. All members were clustered into Groups I, Group II (IIa, IIb, IIc, IId, IIe), and Group III
Gene structure and conserved motif analysis of CiWRKYs
Gene structural diversity can provide insight into the evolution of multigene families [39]. In order to gain a more comprehensive understanding of the structural characteristics of CiWRKYs, we conducted analyses of conserved motifs, conserved domains and exon-intron structures to provide a comprehensive overview. A total of 10 conserved motifs were identified in the C. ichangensis genome using the MEME tool (Table S2). As is shown in Fig. 2A, motifs 1, 2, and 4 were extensively distributed in CiWRKY proteins. Motifs 3 and 9 were detected exclusively in the Group I members, whereas motifs 6, 7, 8, and 10 were only present in the Group II members. Of note, motif 10 was uniquely detected in subgroup IId.
Conserved domain analysis demonstrated that among the 52 CiWRKY proteins, 42 (80.8%) possessed one WRKY domain while the remaining 10 (19.2%) had two WRKY domains. Except the conserved WRKY domains, five other distinct domains were also identified in the CiWRKYs, including the Globin-like subfamily and bZIP subfamily domains (Fig. 2B).
Furthermore, we examined the exon-intron structure of each CiWRKY gene to obtain a more comprehensive understanding of the evolution of C. ichangensis WRKY family. As shown in Fig. 2C, all CiWRKY genes contain one to eleven exons, in which one exon was observed in two genes, two exons in five genes, three exons in 23 genes, four and five exons in eight genes, six exons in five genes, and 11 exons in one gene. Genes belonging to the same group typically exhibit the similar exon-intron structure. For instance, all members of Group III contain three exons and two introns. Overall, the arrangement of exons and intron phases is consistent with the alignment clustering of CiWRKY genes.

Gene structure and conserved motif analysis of CiWRKYs. (A) Motif composition of the CiWRKY proteins. The 10 types of motifs are displayed in boxes with different colors. The details of motif sequence were illustrated in supplemental Table S2. (B) Conserved domains in the CiWRKYs. The six conserved domains are shown by boxes with different colors. (C) Gene structure of CiWRKYs. Green and yellow boxes indicate the UTR and CDS regions, respectively; the black lines indicate introns. Sizes of the genes can be estimated by scale at the bottom
Chromosomal distribution and synteny analysis of CiWRKYs
To identify potential gene duplication events and the phylogenetic relationships among the CiWRKY genes, the syntenic blocks within the C. ichangensis genome were examined (Fig. 3; Table S3). A total of 22 CiWRKY gene pairs were identified and situated on distinct chromosomes, suggesting that segmental duplications occurred in these regions, which may contribute to the CiWRKY family expansion. Distribution of the CiWRKY genes was not equal among the nine chromosomes, with the largest number (12) in chromosome 5 and the least (2) in chromosome 3. Four comparative syntenic maps were established between C. ichangensis and A. thaliana, rise, apple and grape. A total of 38 CiWRKY genes showed a syntenic relationship with counterparts in A. thaliana (Fig. 4A), 19 with rice (Fig. 4B), 44 with apple (Fig. 4C) and 43 with grape (Fig. 4D) (S11–14). The orthologous pairs between C. ichangensis and A. thaliana, rice, apple, and grape were 64, 37, 168, and 78, respectively. Certain CiWRKY genes were found to be orthologous with at least two syntenic gene pairs (particularly with apple WRKY genes), such as CiWRKY75 (Ci036300) and CiWRKY57 (Ci016440), suggesting that these genes may play an important role during the evolutionary process.

Chromosomal distribution and synteny analysis of CiWRKYs. The circle diagram illustrated the chromosomal location of CiWRKY genes and their syntenic relationships. Gray lines represented synteny blocks within the C. ichangensis genome, and the red lines represented CiWRKY gene pairs. The exact position of these WRKY genes was presented in supplemental Table S3

Synteny analysis of WRKY genes between C. ichangensis and other plants. (A–D) Synteny between C. ichangensis and A. thaliana (A), rice (B), apple (C) and grape (D). The collinear blocks were shown with gray lines in the background, while the syntenic WRKY gene pairs were highlighted in red lines. The horizontal columns depict chromosomes with their corresponding chromosome numbers shown beside the diagram. The exact position of these WRKY genes was presented in supplemental Table S11–14
Cis-acting elements analysis in the promoters of CiWRKYs
Cis-acting elements, significant regulators and indicators of gene function, have been comprehensively studied [40]. To investigate the diverse potential functions of CiWRKY genes, the Plant CARE website was used to analyze 2-kb promoter region of the CiWRKY genes. Each CiWRKY gene contained various cis-acting elements (Fig. 5, Table S4). A number of elements associated with stress response or hormone signaling were identified, including abscisic acid-response (ABRE), salicylic acid response (TCA), MeJA-response (TGACG/CGTCA), drought stress-response (MBS), low temperature response (LTR), and defense response (TC-rich), which are widely distributed in the promoters of the CiWRKYs. However, only the ABRE cis-acting element was present in all members of Group I, suggesting that these WRKYs gene may be involved in the ABA signaling pathway and have a functional role in ABA-mediated stress response. In addition, gibberellin responsive (GARE/P-box/TATC-box) elements were detected in the promoters of several CiWRKYs, implying that they are likely to regulate growth and development.

Cis-acting elements in the promoters of CiWRKYs. Ten cis-acting element motifs within the 2 kb of CiWRKYs promoter regions are highlighted and represented by boxes with different colors. ABRE: Abscisic acid response element; CGTCA-motif/ TGACG-motif: MeJA response; GARE-motif/ TATC-box: gibberellin response; LTR: low-temperature response; MBS: MYB binding site; TCA-element: salicylic acid response; TC-rich repeats: defense and stress response; W-box: specific binding motif for WRKYs. The details of cis-acting elements were illustrated in supplemental Table S4
Gene ontology annotation of CiWRKY proteins
Functional annotation and predicted protein-protein interactions could gain insight into regulatory functions of the examined proteins. Gene Ontology (GO) annotations of the 52 CiWRKY proteins were scrutinized using the Dr. Tom web-based solution, an in-house customized data mining system of the BGI (Table S5). Among the protein sequences annotated in the GO database, CiWRKY proteins were categorized into three main groups, cellular component, molecular function, and biological process (Fig. 6). Six terms were greatly enriched in the cellular component category (Fig. 6A), including intracellular anatomical structure (GO:0005622), organelle (GO:0043226), intracellular organelle (GO:0043229), membrane-bounded organelle (GO:0043227), intracellular membrane-bounded organelle (GO:0043231) and nucleus (GO:0005634). In the biological process category (Fig. 6B), most CiWRKYs were found to be involved in the biological regulation (GO:0065007), response to stimulus (GO:0050896), cellular process (GO:0009987). A majority of CiWRKYs were predicted to be involved in the responses against various stimuli, indicating the crucial role of the WRKY members in modulation of stress tolerance. As for the molecular function terms the CiWRKYs were predominantly associated with binding (GO:0005488) and transcription regulator activity (GO:0140110) (Fig. 6C). The top three enriched GO terms are ‘sequence-specific DNA binding’, ‘DNA-binding transcription factor activity’ and ‘transcription regulator activity’ (Fig. 7). These results demonstrate that the CiWRKY proteins are closely involved in various biological processes, in particular the stress response.

Gene ontology (GO) annotation and term enrichment of CiWRKY proteins. (A–C) Enriched GO terms of the Cellular Component (CC, A) Biological Process (BP, B) and Molecular Function (MF, C). GO terms with a Q-value ≤ 0.05 were considered as overrepresented. The details of GO terms were illustrated in supplemental Table S5

The top 20 enriched GO terms among CiWRKY genes. The data were analyzed using the R package and made FDR correction with Q-value ≤ 0.05. The bubble diagram was performed using the Dr. Tom Multi-omics Data Mining System. Gene number was illustrated by the size of circles. Q-value was indicated by the color shading of the bar chart
Protein interaction analysis of CiWRKY proteins
To systematically investigate the CiWRKY protein interactions, we utilized STRING 12.0 software to construct a protein interaction network (Fig. 8). As a result, interactions were identified between 20 CiWRKYs. In the network, CiWRKY33, CiWRKY26, CiWRKY29, and CiWRKY22 were associated with the plant-pathogen interactions (ath04626) and plant MAPKs (ath04016) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways. Furthermore, the interaction among CiWRKY30, CiWRKY40, CiWRKY54, CiWRKY70, and CiWRKY1 is considerably enriched with the GO terms of salicylic acid (GO:0009751), response to oxygen-containing compounds (GO:1,901,700), and chemical response (GO:0042221). Moreover, six CiWRKYs, as well as their corresponding orthologs, participated in a narrower protein interaction network.

Prediction of protein-protein interactions between CiWRKYs. Protein-protein interaction of CiWRKY protein was performed by STRING online search tool. The line thickness represents the level of protein interaction
Expression patterns of CiWRKY genes in response to cold treatment
To better understand the role of C. ichangensis WRKYs in cold response, leaves of three-month-old plants were sampled at five time points (0 h, 6 h, 1 d, 3 d, and 5 d) after cold treatment and subjected to RT-qPCR analysis. All of the 52 CiWRKY genes had detectable expression under cold stress, but displayed disparate expression patterns (Fig. 9, Table S7). Fourteen genes were substantially induced by cold stress, with stable upregulation being observed in CiWRKY71, CiWRKY29, CiWRKY27, and CiWRKY30. On the other hand, the expression levels of CiWRKY74, CiWRKY58, and CiWRKY20 were markedly decreased upon exposure to cold, suggesting that they may have negative functions in modulation of cold tolerance. In addition, 16 CiWRKY genes exhibited a slight induction, while no change was detected in 19 genes under the cold stress. These results indicate that the CiWRKY genes may perform distinct biological functions in response to cold stress. To confirm the RT-qPCR analysis, the transcript profile of 14 cold-responsive CiWRKY genes in RNA-seq data [36] from Ichang papeda at four time points (0 h, 12 h, 24 h, 72 h) after cold treatment were examined (Fig. 10, Table S6), which showed that the gene expression trend was consistent between the two methods, supporting the cold-responsive nature of these genes.

Expression profiles of CiWRKYs under cold treatment. Heatmap of WRKY gene expression in leaves subjected to cold treatment at five time points (0 h, 6 h, 24 h, 72 h, 120 h). The blue color corresponds to low expression levels, and red color corresponds to high expression levels. The heatmap was generated by TBtools. Genes that are substantially induced by cold are labeled by small red circle

The expression patterns of 14 cold-induced CiWRKY genes. The relative expression of cold-induced CiWRKY genes was assessed by RT-qPCR (red lines) and RNA-seq (yellow columns). RT-qPCR data are means ± SD (n = 3)
Silencing of CiWRKY reduced cold resistance in C. ichangensis
CiWRKY31 was found to be one of the genes that was most remarkably induced by cold stress, suggesting that it possibly plays a significant role in cold resistance. To confirm this assumption, VIGS (virus-induced gene silencing) was employed to knock down CiWRKY31 in Ichang papeda. RT-qPCR analysis showed that the transcript levels of CiWRKY31 were markedly down-regulated in the VIGS plants (designated as TRV-CiWRKY31) relative to the control plants (TRV) (Fig. S2). When subjected to freezing treatment (− 4 °C, 8 h), the VIGS plants displayed seriously visible leaf wilting, while this phenotype was not observed for the control (Fig. 11A). In agreement with the plant phenotype, EL was dramatically increase in the VIGS plants relative to the control (Fig. 11B). Meanwhile, chlorophyll fluorescence was considerably impaired, accompanied by lower Fv/Fm ratio, in the VIGS plants when compared to the control plants (Fig. 11C-D). These data indicate that knockdown of CiWRKY31 substantially compromised the cold tolerance of Ichang papeda, indicating that CiWRKY31 is a positive regulator of cold tolerance in Ichang papeda.

Silencing of CiWRKY31 enhances cold sensitivity in Ichang papeda. (A) Phenotype of control plants (TRV), VIGS plants (TRV-CiWRKY31) before and after cold treatment. Scale bars = 1 cm. (B–D) Electrolyte leakage (B), chlorophyll fluorescence imaging (C) and Fv/Fm ratios (D) of tested lines before and after the cold treatment. The false color scale between 0 and 1 is shown beside the imaging. Error bars indicate ± SE (n = 3). Asterisks indicate significant differences between the control and VIGS line under the same growth condition (*P < 0.05; **P < 0.01; ***P < 0.001)
Discussion
CiWRKY genes exhibit both conserved and divergent characteristics
WRKYs, one of the largest transcription factor families in plants, are well recognized for their crucial roles in regulation of plant growth and development and response to adverse environmental stimuli [41]. Due to their valuable biological roles, WRKY members from several plant species have been extensively explored with the release of their whole genome sequences [42,43,44,45]. In the present study, a comprehensive analysis of the WRKY gene family was conducted in C. ichangensis, leading to identification of a total of 52 full-length WRKY genes. Based on phylogenetic analysis in comparison with the Arabidopsis AtWRKYs [19], the C. ichangensis WRKY members were designated CiWRKY1 to CiWRKY75. Notably, the CiWRKY family has only 52 members, which is fewer than other angiosperms such as rice [20], soybean [21] and maize [22]. It has been reported that both C. sinensis and C. ichangensis have the same number of WRKY genes [46]. Furthermore, Poncirus trifoliata, a relative of C. ichangensis, also possesses a comparable number of 51 WRKY genes. These findings indicate that WRKY gene evolution in Rutaceae is probably conserved and that the WRKY family has not undergone significant expansion.
Physicochemical properties of the CiWRKY genes, including their CDS length, protein length, molecular weight, and isoelectric point, were comprehensively analyzed. Interestingly, the 52 CiWRKY genes exhibited substantial differences in terms of gene size. For instance, CiWRKY58 was the largest gene, with a CDS length of 2736 bp, while CiWRKY11 was the smallest one, whose CDS is only 240 bp. Moreover, significant variation was also observed in other properties of CiWRKYs, such as molecular weight and isoelectric point (Table S1). These results indicate that the CiWRKY family exhibits considerable diversity and is potentially involved in various cellular and developmental processes of C. ichangensis. To further elucidate the evolution of CiWRKYs, MUSCLE alignment of CiWRKY and AtWRKY domains was carried out in MEGA7 to form the neighbor-joining trees (Fig. 1). According to the phylogenetic analysis, the CiWRKY genes were classified into three major groups (Group I, II, III), and Group II was further group into five subfamilies (II a, II b, II c, II d, II e), which is in line with other plants [23]. The majority of the CiWRKYs (35) were classified into Group II (Fig. 2). The structure of conserved domains is highly similar in the members within a given group, implying that the WRKY family genes remained relatively stable during C. ichangensis evolution. Moreover, chromosomal duplication was detected, emerging throughout the C. ichangensis evolutionary process. Overall, 22 segmental duplication events involving 28 WRKY genes were identified, while there was a lack of tandem duplication events (Fig. 3). This may explain the smaller number of CiWRKY genes in this plant, while segmental duplications were the major driver for WRKY gene expansion during the evolutionary process of C. ichangensis.
WRKY domains, the typical structural feature of WRKY transcription factors, serve as a crucial point of reference in their evolutionary process [10]. It should be noted that although there is an extraordinary conservation of the WRKY domain in the C. ichangensis WRKY family, as 50 members possess the WRKYGQK heptapeptide. By contrast, CiWRKY50 possesses the WRKYGKK variant, implying that there is a possibility that this gene may perform a distinct biological function (Table S1). Previous research has indicated that AtWRKY50 promotes the production of salicylic acid and plays a role in countering defense responses induced by JA [47]. CiWRKY50 is a cold stress responsive gene, and the unique motif structure implies that C. ichangensis is consistently evolving WRKY genes to withstand a wide range of environmental stresses.
Gene ontology annotations and protein interactions shed light on CiWRKY functions
Gene Ontology (GO) is a standardized system for gene function classification, providing a continuously updated and comprehensive vocabulary to describe the features of genes and gene products within an organism [48]. A total of 29 GO terms were identified related to cellular component (CC), biological process (BP), and molecular function (MF) (Fig. 6). In the BP class, the top three overrepresented terms were associated with “response to stimulus”, including “defense response to bacterium”; “response to bacterium” and “response to chitin”. WRKY transcription factors play a crucial role in regulating various plant defense response, acting as transcriptional activators or repressors in a differential and graded manner [18]. Previous studies have shown their involvement in plant defense response in rice [49], pepper [50] and cotton [51], which is consistent with the GO annotation in C. ichangensis. This further confirms that WRKY transcription factors contribute to environmental stress tolerance. Within the CC class, six GO terms were enriched, including “nuclear”, “intracellular anatomical structure”, “intracellular organelle”, “membrane-bound organelle”, “intracellular membrane-bound organelle”, and “organelle”, suggesting that CiWRKY proteins perform highly complex functions in regulating various plant cellular processes. Within the MF category, the enriched GO terms included ‘binding’ and ‘transcription regulatory activity’, which is consist with the natures of transcription factors that have the ability to binding to specific DNA sequences. Overall, our findings suggest that CiWRKYs are closely involved in regulation of cellular responses to various environmental stimuli.
Protein-protein interactions can indicate the potential regulatory functions of proteins [52]. The interaction of CiWRKY proteins were identified using the STRING12 program, with the confidence parameter set at 0.4. Of these, 20 CiWRKY proteins displayed extensive interactions with other family members (Fig. 8). Five proteins, including CiWRKY9, CiWRKY32, CiWRKY40, CiWRKY54, and CiWRKY70, were positioned at the center of the network, forming a key regulatory hub that might play crucial roles in various biological processes related to the CiWRKYs. Furthermore, a significant interaction was observed between CiWRKY29 and CiWRKY22, two members in the subfamily IIe. These results implying that WRKY proteins have the potential to form homodimers or heterodimers to exert diverse functional roles. In recent years, a substantial number of WRKY-interacting proteins have been identified. For example, the interaction between WRKY33 and WRKY12 in Arabidopsis can elevate the expression of RAP2.2, leading to enhanced tolerance to hypoxia stress [53]. In addition, GhWRKY41 was found to facilitate a positive feedback regulation loop to improve the resistance of V. dahliae in cotton [54]. The extensive interactions between WRKY proteins allow for cooperative functions and promote a dynamic regulation of target genes.
The CiWRKY genes may play a crucial role in cold tolerance
Recent evidence has revealed the vital functions of WRKY genes in response to cold stress. For instance, VaWRKY33 was upregulated by VaERF092, which was shown to enhance cold tolerance in Amur grape [55]. SmWRKY26 and SmWRKY32 of eggplant exhibited positive regulatory functions in response to cold stress [56]. In rice, the cold-responsive OsWRKY71 was reported to act as a transcriptional repressor that positively affected cold tolerance [57]. While the WRKY functions under cold stress have been investigated in various plant species, their involvement in cold stress resistance in C. ichangensis is not fully understood. In this study, the expression levels of CiWRKY genes were evaluated under cold stress, which revealed that the genes showed various degree of cold response. Of the genes, 14 were drastically induced by cold, 16 were slightly induced, while three genes were repressed. It is worth mentioning that 19 genes underwent negligible alterations of expression levels in the presence of cold treatment. The homologous genes of WRKY40 and WRKY33 in Arabidopsis have been previously shown to play significant roles in regulation of response to both biotic stresses, such as response to bacteria [58,59,60] or fungi [61, 62] and abiotic stresses, such as salt [37], osmotic [53], cold [63], and drought stress [64]. Some CiWRKY genes like CiWRKY31 were consistently up-regulated in the presence of cold stress, implying it is possible to function in adaptation to cold treatment. The remaining highly induced genes exhibited rapid up-regulation at the onset of cold stress, peaking at either 24–72 h after the cold treatment. The diversity of expression patterns appears to be intricately linked to gene function. Due to their significance, the genes that exhibited persistent up-regulation could serve as valuable candidate to improve cold tolerance. It should be noted that three CiWRKY genes (CiWRKY20, 58, and 74) were significantly repressed by cold stress. WRKY20 of wild soybean has been confirmed to positively regulate wax biosynthesis-related genes for enhancing drought tolerance [65]. Meanwhile, OsWRKY74 has also been found to be suppressed by cold stress but strongly induced by phosphate and Fe deficiency, indicating that WRKY74 may play a crucial role in response to nutrient starvation [66]. Additionally, SlWRKY58 was shown to confer drought tolerance in tomato [67]. These findings corroborate the essential roles of WRKY genes in response to various environmental cues. To verify the expression analysis and our assumption, CiWRKY31 was knocked down by VIGS. Silencing CiWRKY31 significantly decreased the cold tolerance of Ichang papeda, indicating that this gene function as a positive regulator of cold stress response. This result indicates that the cold-induced WRKYs may play a positive role in modulation of cold tolerance, and extra work is required to elucidate the functions of other cold-responsive WRKYs. In addition to cold stress, WRKY31 has been demonstrated to participate in modulation of drought tolerance [68], suggesting that WRKY31 might have a significant biological function in combating multiple stresses.
Conclusions
In summary, the Citrus ichangensis WRKY gene family was comprehensively analyzed in terms of its functional and structural properties. A total of 52 full-length WRKY genes were characterized and further classified into three main groups. The structure of exon-introns and the composition of motifs were similar in the same group and subgroup. Synteny and comparative phylogenetic analyses among the WRKY genes provided valuable insights into the evolutionary characteristics of C. ichangensis WRKY genes. Additionally, Gene Ontology annotation and protein-protein interaction prediction provide a foundation for further elucidate the functional properties of CiWRKY genes. Based on the expression patterns of CiWRKY genes under cold treatment we identified 14 cold-induced members, among which CiWRKY31 was verified to be a positive regulator of cold tolerance. These findings gain valuable insights into understanding the function of CiWRKYs in cold stress responses, and unravel potential WRKY members that can be selected as candidates for improving cold tolerance in Citrus.
Data availability
The sequence information of Citrus ichangensis and Arabidopsis WRKY family genes were collected from Citrus Pan-genome to Breeding Database (http://citrus.hzau.edu.cn/index.php) and The Arabidopsis information Resource (https://www.arabidopsis.org/). All data analyzed during this study are included in this article and its additional files.
References
Wu GA, Terol J, Ibanez V, López-García A, Pérez-Román E, Borredá C, Domingo C, Tadeo FR, Carbonell-Caballero J, Alonso R, et al. Genomics of the origin and evolution of Citrus. Nature. 2018;554(7692):311–6.
Yang X, Li H, Yu H, Chai L, Xu Q, Deng X. Molecular phylogeography and population evolution analysis of Citrus ichangensis (Rutaceae). Tree Gene Genomes. 2017;13(1):29.
Rieger M, Krewer G, Lewis P, Linton M, McClendon T. Field evaluation of cold hardy citrus in Coastal Georgia. Horttechnology. 2003;13(3):540–4.
Dietz K-J, Vogelsang L. A general concept of quantitative abiotic stress sensing. Trends Plant Sci. 2024;29(3):319–28.
Kilian J, Peschke F, Berendzen KW, Harter K, Wanke D. Prerequisites, performance and profits of transcriptional profiling the abiotic stress response. Biochim Biophys Acta. 2012;1819(2):166–75.
Agarwal P, Reddy MP, Chikara J. WRKY: its structure, evolutionary relationship, DNA-binding selectivity, role in stress tolerance and development of plants. Mol Biol Rep. 2011;38(6):3883–96.
Javed T, Gao SJ. WRKY transcription factors in plant defense. Trends Genet. 2023;39(10):787–801.
Chen F, Hu Y, Vannozzi A, Wu K, Cai H, Qin Y, Mullis A, Lin Z, Zhang L. The WRKY transcription factor family in model plants and crops. Crit Rev Plant Sci. 2018;36:311–35.
Eulgem T, Rushton PJ, Robatzek S, Somssich IE. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000;5(5):199–206.
Rinerson CI, Rabara RC, Tripathi P, Shen QJ, Rushton PJ. The evolution of WRKY transcription factors. BMC Plant Biol. 2015;15:66.
Phukan UJ, Jeena GS, Shukla RK. WRKY transcription factors: molecular regulation and stress responses in plants. Front Plant Sci. 2016;7:760.
Yin W, Wang X, Liu H, Wang Y, Nocker S, Tu M, Fang J, Guo J, Li Z, Wang X. Overexpression of VqWRKY31 enhances powdery mildew resistance in grapevine by promoting salicylic acid signaling and specific metabolite synthesis. Hortic Res. 2022;9:uhab064.
Huang X, Cao L, Fan J, Ma G, Chen L. CdWRKY2-mediated sucrose biosynthesis and CBF-signalling pathways coordinately contribute to cold tolerance in bermudagrass. Plant Biotechnol J. 2022;20(4):660–75.
Zhang M, Zhao R, Huang K, Huang S, Wang H, Wei Z, Li Z, Bian M, Jiang W, Wu T, et al. The OsWRKY63-OsWRKY76-OsDREB1B module regulates chilling tolerance in rice. Plant J. 2022;112(2):383–98.
Chen S, Cao H, Huang B, Zheng X, Liang K, Wang GL, Sun X. The WRKY10-VQ8 module safely and effectively regulates rice thermotolerance. Plant Cell Environ. 2022;45(7):2126–44.
Lim C, Kang K, Shim Y, Yoo SC, Paek NC. Inactivating transcription factor OsWRKY5 enhances drought tolerance through abscisic acid signaling pathways. Plant Physiol. 2022;188(4):1900–16.
Bo C, Cai R, Fang X, Wu H, Ma Z, Yuan H, Cheng B, Fan J, Ma Q. Transcription factor ZmWRKY20 interacts with ZmWRKY115 to repress expression of ZmbZIP111 for salt tolerance in maize. Plant J. 2022;111(6):1660–75.
Jiang J, Ma S, Ye N, Jiang M, Cao J, Zhang J. WRKY transcription factors in plant responses to stresses. J Integr Plant Biol. 2017;59(2):86–101.
Dong J, Chen C, Chen Z. Expression profiles of the Arabidopsis WRKY gene superfamily during plant defense response. Plant Mol Biol. 2003;51(1):21–37.
Ross CA, Liu Y, Shen QJ. The WRKY gene family in rice (Oryza sativa). J Integr Plant Biol. 2007;49(6):827–42.
Yu Y, Wang N, Hu R, Xiang F. Genome-wide identification of soybean WRKY transcription factors in response to salt stress. Springerplus. 2016;5(1):920.
Zhang T, Tan D, Zhang L, Zhang X, Han Z. Phylogenetic analysis and drought-responsive expression profiles of the WRKY transcription factor family in maize. Agri Gene. 2017;3:99–108.
Huang S, Gao Y, Liu J, Peng X, Niu X, Fei Z, Cao S, Liu Y. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum. Mol Genet Genomics. 2012;287(6):495–513.
Chen C, Chen X, Han J, Lu W, Ren Z. Genome-wide analysis of the WRKY gene family in the cucumber genome and transcriptome-wide identification of WRKY transcription factors that respond to biotic and abiotic stresses. BMC Plant Biol. 2020;20(1):443.
Meng D, Li Y, Bai Y, Li M, Cheng L. Genome-wide identification and characterization of WRKY transcriptional factor family in apple and analysis of their responses to waterlogging and drought stress. Plant Physiol Biochem. 2016;103:71–83.
Huang X, Li K, Xu X, Yao Z, Jin C, Zhang S. Genome-wide analysis of WRKY transcription factors in white pear (Pyrus Bretschneideri) reveals evolution and patterns under drought stress. BMC Genom. 2015;16:1104.
Xie T, Chen C, Li C, Liu J, Liu C, He Y. Genome-wide investigation of WRKY gene family in pineapple: evolution and expression profiles during development and stress. BMC Genom. 2018;19(1):490.
Liu H, Wang X, Liu S, Huang Y, Guo YX, Xie WZ, Liu H, Tahir Ul Qamar M, Xu Q, Chen LL. Citrus pan-genome to breeding database (CPBD): a comprehensive genome database for citrus breeding. Mol Plant. 2022;15(10):1503–5.
Mistry J, Chuguransky S, Williams L, Qureshi M, Salazar GA, Sonnhammer ELL, Tosatto SCE, Paladin L, Raj S, Richardson LJ, et al. Pfam: the protein families database in 2021. Nucleic Acids Res. 2021;49(D1):D412–9.
Potter SC, Luciani A, Eddy SR, Park Y, Lopez R, Finn RD. HMMER web server: 2018 update. Nucleic Acids Res. 2018;46(W1):W200–4.
Letunic I, Khedkar S, Bork P. SMART: recent updates, new developments and status in 2020. Nucleic Acids Res. 2021;49(D1):D458–60.
Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, Xia R. TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020;13(8):1194–202.
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33(7):1870–4.
Wang Y, Tang H, Debarry JD, Tan X, Li J, Wang X, Lee TH, Jin H, Marler B, Guo H, et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012;40(7):e49.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2–∆∆CT method. Methods. 2001;25(4):402–8.
Xiao P, Qu J, Wang Y, Fang T, Xiao W, Wang Y, Zhang Y, Khan M, Chen Q, Xu X et al. Transcriptome and metabolome atlas reveals contributions of sphingosine and chlorogenic acid to cold tolerance in Citrus. Plant Physiol. 2024; kiae327.
Dai W, Wang M, Gong X, Liu JH. The transcription factor FcWRKY40 of Fortunella crassifolia functions positively in salt tolerance through modulation of ion homeostasis and proline biosynthesis by directly regulating SOS2 and P5CS1 homologs. New Phytol. 2018;219(3):972–989.
Dahro B, Wang F, Peng T, Liu JH. PtrA/NINV, an alkaline/neutral invertase gene of Poncirus trifoliata, confers enhanced tolerance to multiple abiotic stresses by modulating ROS levels and maintaining photosynthetic efficiency. BMC Plant Biol. 2016;16:76.
Wang H, Chen W, Xu Z, Chen M, Yu D. Functions of WRKYs in plant growth and development. Trends Plant Sci. 2023;28(6):630–645.
Wittkopp PJ, Kalay G. Cis-regulatory elements: molecular mechanisms and evolutionary processes underlying divergence. Nat Rev Genet. 2011;13(1):59–69.
Li W, Pang S, Lu Z, Jin B. Function and mechanism of WRKY transcription factors in abiotic stress responses of plants. Plants (Basel). 2020;9(11):1515.
Song Y, Cui H, Shi Y, Xue J, Ji C, Zhang C, Yuan L, Li R. Genome-wide identification and functional characterization of the Camelina sativa WRKY gene family in response to abiotic stress. BMC Genom. 2020;21(1):786.
Sun S, Chen H, Yang Z, Lu J, Wu D, Luo Q, Jia J, Tan J. Identification of WRKY transcription factor family genes in Pinus massoniana Lamb. And their expression patterns and functions in response to drought stress. BMC Plant Biol. 2022;22(1):424.
Wen F, Wu X, Li T, Jia M, Liao L. Characterization of the WRKY gene family in Akebia trifoliata and their response to Colletotrichum Acutatum. BMC Plant Biol. 2022;22(1):115.
Wu W, Zhu S, Xu L, Zhu L, Wang D, Liu Y, Liu S, Hao Z, Lu Y, Yang L, et al. Genome-wide identification of the Liriodendron chinense WRKY gene family and its diverse roles in response to multiple abiotic stress. BMC Plant Biol. 2022;22(1):25.
Dai WS, Peng T, Wang M, Liu JH. Genome-wide identification and comparative expression profiling of the WRKY transcription factor family in two Citrus species with different Candidatus Liberibacter asiaticus susceptibility. BMC Plant Biol. 2023;23(1):159.
Hussain RMF, Kim HK, Khurshid M, Akhtar MT, Linthorst HJM. Overexpression of AtWRKY50 is correlated with enhanced production of sinapic derivatives in Arabidopsis. Metabolomics. 2018;14(3):25.
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet. 2000;25(1):25–9.
Zheng C, Zhou J, Yuan X, Zheng E, Liu X, Cui W, Yan C, Wu Y, Ruan W, Yi K, et al. Elevating plant immunity by translational regulation of a rice WRKY transcription factor. Plant Biotechnol J. 2024;22(4):1033–48.
Dang F, Wang Y, She J, Lei Y, Liu Z, Eulgem T, Lai Y, Lin J, Yu L, Lei D, et al. Overexpression of CaWRKY27, a subgroup IIe WRKY transcription factor of Capsicum annuum, positively regulates tobacco resistance to Ralstonia solanacearum infection. Physiol Plant. 2014;150(3):397–411.
Wang L, Guo D, Zhao G, Wang J, Zhang S, Wang C, et al. Group IIc WRKY transcription factors regulate cotton resistance to Fusarium oxysporum by promoting GhMKK2-mediated flavonoid biosynthesis. New Phytol. 2022;236(1):249–265.
Chi Y, Yang Y, Zhou Y, Zhou J, Fan B, Yu JQ, et al. Protein–protein interactions in the regulation of WRKY transcription factors. Mol Plant. 2013;6(2):287–300.
Tang H, Bi H, Liu B, Lou S, Song Y, Tong S, et al. WRKY33 interacts with WRKY12 protein to up‐regulate RAP2.2 during submergence induced hypoxia response in Arabidopsis thaliana. New Phytol. 2020;229(1):106–125.
Xiao S, Ming Y, Hu Q, Ye Z, Si H, Liu S, Zhang X, Wang W, Yu Y, Kong J, et al. GhWRKY41 forms a positive feedback regulation loop and increases cotton defence response against Verticillium Dahliae by regulating phenylpropanoid metabolism. Plant Biotechnol J. 2023;21(5):961–78.
Sun X, Zhang L, Wong DCJ, Wang Y, Zhu Z, Xu G, et al. The ethylene response factor VaERF092 from Amur grape regulates the transcription factor VaWRKY33, improving cold tolerance. Plant J. 2019;99(5):988–1002.
Yang Y, Liu J, Zhou X, Liu S, Zhuang Y. Identification of WRKY gene family and characterization of cold stress-responsive WRKY genes in eggplant. PeerJ. 2020;8:e8777.
Kim CY, Vo KTX, Nguyen CD, Jeong DH, Lee SK, Kumar M, et al. Functional analysis of a cold-responsive rice WRKY gene, OsWRKY71. Plant Biotech Reports. 2016;10:13–23.
Chakraborty J, Ghosh P, Sen S, Das S. Epigenetic and transcriptional control of chickpea WRKY40 promoter activity under Fusarium stress and its heterologous expression in Arabidopsis leads to enhanced resistance against bacterial pathogen. Plant Sci. 2018;276:250–67.
Verma V, Srivastava AK, Gough C, Campanaro A, Srivastava M, Morrell R, Joyce J, Bailey M, Zhang C, Krysan PJ, et al. SUMO enables substrate selectivity by mitogen-activated protein kinases to regulate immunity in plants. Proc Natl Acad Sci U S A. 2021;118(10):e2021351118.
Geilen K, Böhmer M. Dynamic subnuclear relocalisation of WRKY40 in response to Abscisic acid in Arabidopsis thaliana. Sci Rep. 2015;5:13369.
Zheng Z, Qamar SA, Chen Z, Mengiste T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2006;48(4):592–605.
Bao W, Wang X, Chen M, Chai T, Wang H. A WRKY transcription factor, PcWRKY33, from Polygonum cuspidatum reduces salt tolerance in transgenic Arabidopsis thaliana. Plant Cell Rep. 2018;37(7):1033–1048.
Guo M, Yang F, Liu C, Zou J, Qi Z, Fotopoulos V, Lu G, Yu J, Zhou J. A single-nucleotide polymorphism in WRKY33 promoter is associated with the cold sensitivity in cultivated tomato. New Phytol. 2022;236(3):989–1005.
Wang C, Ru J, Liu Y, Yang J, Li M, Xu Z, Fu J. The maize WRKY transcription factor ZmWRKY40 confers drought resistance in transgenic Arabidopsis. Int J Mol Sci. 2018;19(9):2580.
Luo X, Bai X, Sun X, Zhu D, Liu B, Ji W, Cai H, Cao L, Wu J, Hu M, et al. Expression of wild soybean WRKY20 in Arabidopsis enhances drought tolerance and regulates ABA signaling. J Exp Bot. 2013;64(8):2155–69.
Dai X, Wang Y, Zhang WH. OsWRKY74, a WRKY transcription factor, modulates tolerance to phosphate starvation in rice. J Exp Bot. 2016;67(3):947–60.
Karkute SG, Gujjar RS, Rai A, Akhtar M, Singh M, Singh B. Genome wide expression analysis of WRKY genes in tomato (Solanum lycopersicum) under drought stress. Plant Gene. 2018;13:8–17.
Ge M, Tang Y, Guan Y, Lv M, Zhou C, Ma H, Lv J. TaWRKY31, a novel WRKY transcription factor in wheat, participates in regulation of plant drought stress tolerance. BMC Plant Biol. 2024;24(1):27.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (32330095), National Key Research and Development Program of China (2022YFD1200503) and the Key Research and Development Program of Jiangsu Province (BE2023328).
Author information
Authors and Affiliations
Contributions
J.Q. and J.H.L. designed the study. J.Q. Y.K.Z. and X.Z. performed the experiments. J.Q. P.X. Z.Q.Z and Y.L.W. analyzed the data and interpreted the results. J.Q. wrote the manuscript. J.H.L. revised and finalized the writing. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Qu, J., Xiao, P., Zhao, ZQ. et al. Genome-wide identification, expression analysis of WRKY transcription factors in Citrus ichangensis and functional validation of CiWRKY31 in response to cold stress. BMC Plant Biol 24, 617 (2024). https://doi.org/10.1186/s12870-024-05320-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-024-05320-0