
Abstract
Background
Aegilops geniculata Roth is closely related to common wheat (Triticum aestivum L.) and is a valuable genetic resource for improvement of wheat.
Results
In this study, the W19513 line was derived from the BC1F10 progeny of a cross between wheat ‘Chinese Spring’ and Ae. geniculata SY159. Cytological examination showed that W19513 contained 44 chromosomes. Twenty-two bivalents were formed at the first meiotic metaphase I in the pollen mother cellsand the chromosomes were evenly distributed to opposite poles at meiotic anaphase I. Genomic in situ hybridization demonstrated that W19513 carried a pair of alien chromosomes from the M genome. Fluorescence in situ hybridization confirmed detection of variation in chromosomes 4A and 6B. Functional molecular marker analysis using expressed sequence tag–sequence-tagged site and PCR-based landmark unique gene primers revealed that the alien gene belonged to the third homologous group. The marker analysis confirmed that the alien chromosome pair was 3Mg. In addition, to further explore the molecular marker specificity of chromosome 3Mg, based on the specific locus amplified fragment sequencing technique, molecular markers specific for W19513 were developed with efficiencies of up to 47.66%. The W19513 line was inoculated with the physiological race E09 of powdery mildew (Blumeria graminis f. sp. tritici) at the seedling stage and showed moderate resistance. Field inoculation with a mixture of the races CYR31, CYR32, CYR33, and CYR34 of the stripe rust fungus (Puccinia striiformis f. sp. triticii) revealed that the line W19513 showed strong resistance.
Conclusions
This study provides a foundation for use of the line W19513 in future genetic research and wheat improvement.
Similar content being viewed by others

Background
Wheat (Triticum aestivum L.) is an important grain crop cultivated worldwide. The grains are a major source of starch, dietary fiber and nutrients and provides raw materials for industrial food production [1]. Among them, wheat is the main food source in many countries, which provides more than 20% of the energy and protein for human beings around the world [2]. China is the largest wheat producer and consumer in the world [3]. Continued population growth, serious environmental degradation, sharp reduction in the area of effective cultivated land and the ongoing conflict in resource demands all represent severe challenges to food production [4]. Therefore, wheat breeding faces important challenges globally and in China in the twenty-first century [5]. The loss of genetic diversity of cultivated wheat reduces the quality and yield of grains and increases vulnerability to biotic and abiotic environmental stresses [6]. Maintaining the sustainable production and stability of wheat grain yields is closely associated with China’s food security and socio-economic development strategies [7, 8]. Wild relatives of wheat (including wild and cultivated forms) are a reservoir for a large number of genes for desirable agronomic traits, and thus, for example, are an important source of disease resistance genes for cultivated wheat [9].
Aegilops is closely related and genetically similar to Triticum [10]. Almost all species of Aegilops are cross-compatible with wheat and, therefore, are an excellent genetic resource for improvement of wheat quality and other agronomic traits [11]. A large number of disomic addition lines (DALs) of wheat and its wild relatives were established by distant hybridization and chromosome engineering [12]. Aegilops geniculata Roth (2n = 4x = 28) is an annual self-pollinating allotetraploid species classified in Aegilops sect. Aegilops. Its chromosome configuration is UgUgMgMg of which the Ug and Mg subgenomes are derived from Ae. umbellulata Zhuk. (UU, 2n = 2x = 14) [13] and Ae. comosa Sm. (MM, 2n = 2 x = 14), respectively [14, 15]. In the course of long-term evolution, Ae. geniculata has evolved a wealth of morphological variation and harbors a large number of stress-adaptation genes that confer traits such as disease and insect resistance, salt tolerance [16], cold and heat tolerance, and precocious maturity [17]. Therefore, Ae. geniculata is an important wild gene source for wheat improvement [18].
Powdery mildew of wheat, caused by Blumeria graminis f. sp. tritici, is a serious airborne leaf disease worldwide [10, 19], and mainly occurs in cool areas with a maritime climate. Stripe rust is a recurring disease caused by the fungus Puccinia striiformis f. sp. tritici (Pst) [20] and, together with powdery mildew, causes massive losses to agricultural production [21]. Breeding and planting of resistant cultivars is the most economical, effective, safe and reliable means to control wheat diseases. Identification of disease-resistant germplasm is not only beneficial for the breeding of disease-resistant cultivars, but also broadens the diversity of disease-resistance genes [22]. The screening of germplasm for identification of sources of disease resistance is a common focus of research into disease resistance. Disease-resistance genes (Yr8, Lr9, and Pm29) have been transferred from the U and M genomes of Aegilops to common wheat [23,24,25]. The use of wild relatives of wheat to generate, screen, and identify novel genetic lines carrying disease-resistance genes [26] can lay a foundation for the transfer of additional disease-resistance genes to wheat.
Next-generation sequencing technology enables large-scale application of molecular and genome technology for crop improvement, and is important in the development of molecular markers, including markers for RNA-sequencing and other technologies [27, 28], such as whole-genome sequencing(WGS), genotyping-by-sequencing (GBS) and specific-locus amplified fragment sequencing (SLAF-seq) [29,30,31]. SLAF-seq is a method of simplified genome sequencing intended for large-scale genotyping, which has the advantages of long effective reads, high throughput, and flexible scheme design. In addition, SLAF-seq plays a fundamental role in molecular breeding and, for example, has been used to develop specific markers on chromosome 7E of Thinopyrum elongatum with high efficiency of up to 65.9% [32]. To date, most of the specific markers for Aegilops were derived from wheat markers, which were inefficiently developed. New markers need to be developed urgently to accelerate the process of theoretical research and gene editing for utilization in wheat breeding. The success of SLAF-seq in the development of chromosome-specific molecular markers provides strong technical support for rapid screening of exogenous materials and shortening of the breeding cycle.
In previous research done by our research group, ‘Chinese Spring’ (CS) was used as the female parent that was crossed with Ae. geniculata ‘SY159’ as the male parent. The F1 progeny were backcrossed with CS and repeated selfing was performed to generate the BC1F10 population. In the present study, molecular cytological analysis, in situ hybridization, functional molecular markers, development-specific molecular markers based on SLAF-seq, assessment of morphological traits and stress resistance were employed to genetically characterize the disomic addition line W19513 from this population. We identified the configuration and number of chromosomes as well as the homologous group relationship of the alien chromosomes, and confirmed the desirable disease resistance and agronomic traits of the line. The results provide a foundation for use of the W19513 line in future genetic research and breeding improvement.
Results
Cytological observations
The W19513 line was derived from the cross between CS and SY159, followed by one backcross to CS and repeated selfing to generate the BC1F10 population. Cytological examination revealed that the number of mitotic metaphase chromosomes (44) in the root tip of indoor- and field-cultivated plants was identical (Fig. 1A). Twenty-two pairs of chromosomes were observed in the pollen mother cells at meiosis, the chromosomes were evenly paired, and no unpaired chromosomes were observed (Fig. 1B). At meiotic anaphase chromosome separation was uniform, the chromosomes moved to the opposite poles synchronously, and no laggard chromosomes were observed (Fig. 1C). Therefore, W19513 showed high cytological stability.

Root tip and meiotic analysis of chromosome characteristics of W19513, 2n = 22I + 22I Root tip cells at mitotic metaphase (A). Pollen mother cell chromosomal configurations at meiotic metaphase I, 2n = 44 (B). Pollen mother cell chromosomal configurations at prophase II, n = I (C)
Genomic DNA of SY159 (as the probe DNA) and that of CS (as the blocking DNA) were extracted using a modified cetyltrimethylammonium bromide (CTAB) method [33] for genomic in situ hybridization (GISH). Two chromosomes showed the probe signal (Fig. 2A). This result implied that a pair of alien chromosomes were added to the genome of W19513. To further verify this result, GISH was conducted using CS as the blocking DNA, and Ae. umbellulata (Fig. 2B), and Ae. comosa (Fig. 2C) as the probe DNA. These results indicated that W19513 contained a pair of additional chromosomes from the M genome.

Karyotypes with the genomic composition variation of W19513, which was obtained by using GISH analysis. A SY159 genomic DNA. B The DNA of Ae. umbellulata. C The DNA of Ae. comosa. The red arrow indicates the introduction of alien chromosomes of Ae. geniculate. Chromosomes were counterstained by DAPI (blue). Bar indicates 10 μm
Functional molecular markers analysis
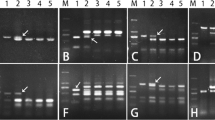
To determine the homologous group relationship of the alien chromosomes pair, 156 pairs of expressed sequence tag–sequence-tagged site (EST-STS) primers and 173 pairs of PCR-based landmark unique gene (PLUG) primers distributed across the seven homologous groups were analyzed using molecular markers. The pair of alien chromosomes was then compared with the amplified bands of the parents SY159 and CS. Five primers (BM134465, CD343475, BQ169491, CD452844, and BF429203) amplified specific bands (Fig. 3H–L). Specific bands of Ae. geniculata were amplified by five PLUG primers (TNAC1300, TNAC1341, TNAC1383, TNAC1627, and TNAC1364) (Fig. 3A–G), all of which belonged to the third homologous group (Table 1). Thus, the alien chromosomes belong to the third homologous group of the M genome (3 M).

PLUG and EST–STS functional molecular marker analysis of W19513. (M). DL2000 (2 kb DNA ladder), (1). CS, (2). SY159, (3). W19513. (A-G). The PLUG markers amplification results with TNAC1364, TNAC1383, TNAC1300, TNAC1627, TNAC1364, TNAC1627, TNAC1341. A–D. The TaqI digestion; E–G. The HAEII digestion. H–L. The EST–STS markers amplification results with BM134465, CD343475, BQ169491, CD452844, BF429203. The red arrows indicate the SY159 specific bands
In situ hybridization
The oligonucleotide probes Oligo-PTa535 and Oligo-pSc119.2 were used for fluorescence in situ hybridization (FISH) analysis, and then the pair of alien chromosomes was compared with the CS standard karyotype map [34]. The karyotype of W19513 was essentially identical to that of CS, comprising 42 chromosomes with an additional pair of alien chromosomes (Fig. 4A). The results of sequential FISH-GISH analysis further confirmed that the line W19513 carried a pair of alien chromosomes from Ae. geniculata. Comparison of the extra pair of chromosomes on the FISH map showed signal from the Ae. geniculata probe (Fig. 4B). Interestingly, comparison of the FISH karyotype of W19513 with that of CS (Fig. 4C) revealed variations in chromosomes 4A and 6B (Fig. 4D). Collectively, the aforementioned results showed that W19513 carried an additional pair of chromosomes 3Mg derived from Ae. geniculata.

Karyotypes with the genomic composition variation of W19513, which was obtained by using FISH and sequential FISH-GISH analysis. FISH used Oligo-pSc119.2 (green) and Oligo-pTa535 (red) as probes. The SY159 genomic DNA was used as probe for sequential FISH- GISH. The red arrow indicates the introduction of alien chromosomes of SY159. White arrows indicated the structural variation of the chromosomes 4A and 6B. A FISH of W19513. B GISH of W19513 in the same cell. C Analysis of CS by FISH. D FISH signal comparison between CS and W19513 on chromosomes 4A, 6B. Chromosomes were counterstained by DAPI (blue). Bar indicates 10 μm
Development of specific molecular markers
The SLAF-seq data comprised a total of 4,733,453, 8,752,434, and 6,943,541 raw reads for CS, SY159, and W19513, respectively. The average Q30 score was 94.15% and the average GC content was 48.66%. After filtering out low-depth data, the final numbers of SLAF-seq reads were 314,571, 193,701, and 371,647 for CS, SY159, and W19513, respectively, and the average sequencing depth was 6.3850. Using the Burrows–Wheeler Alignment (BWA) tool, 2888 reads were observed to show 50% similarity to the CS reference genome (IWGSC RefSeq v1.0). Among these reads, 634 reads showed at least 90% similarity to SY159 and were considered to be specific fragments of chromosome 3Mg. To develop chromosome 3 M-specific markers, 128 primers were designed based on 128 randomly selected fragments and were used to amplify the sequence of CS, SY159, Ae. comosa, Ae. umbellulata and W19513. Among these markers, 61 markers amplified SY159-specific bands with a maximum success rate of 47.66% (Table 2). Ultimately, 61 sequences of chromosome 3 M were obtained (data not shown). The 61 3 M-specific markers were further divided into three categories, namely W19513, SY159 and Ae. comosa and Ae. umbellulata, of which four markers (Fig. 5) showed the same amplification patterns in W19513, SY159, Ae. comosa and Ae. umbellulata (Fig. 5Type 1); 13 markers amplified the same bands in W19513 and SY159, but not the corresponding bands in Ae. comosa and Ae. umbellulata (Fig. 5Type 2); and 44 markers amplified the same bands in W19513, SY159, and Ae. comosa, but not Ae. umbellulata (Fig. 5Type 3).

Amplification patterns of Aegilops accessions using W19513. SLAF markers M124, M39, M52 and M23. (M) DL2000 (2 kb DNA ladder). (1) CS. (2) SY159. (3) Ae. comosa. (4) Ae. umbellulate. (5) W19513. The red arrows indicate the specific bands
Assessment of agronomic traits and disease resistance
The agronomic traits of W19513 and the parents CS and SY159 were analyzed. No significant differences in plant height, number of grains per spikelet, spikelet number, spikelet length, and 1000-grain weight were observed between W19513 and CS. However, notable differences in the lower spike nodes between W19513 and CS were noted. Significant differences were observed in plant height, spikelet number, tiller number, and 1000-grain weight between W19513 and SY159. These results inferred that the W19513 line showed overall morphological intermediary between the parents and produced long panicle nodes and stable agronomic traits (Fig. 6A–D, Table 3).

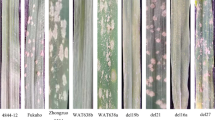
Morphological characteristics (A-D), Powdery mildew (E) and Stripe rust reaction (F) of W19513. (1) CS; (2) SY159; (3) W19513; (4) Shaanyou225; (5) Xuixianhong (A) plants. (B) spikes. (C) florets. (D) grain width. (E) Identification of resistance to powdery mildew at seedling stage. (F) Symptoms in response to inoculation with the mixture of Pst races at the adult stages
The degree of resistance to powdery mildew was determined by inoculation with powdery mildew physiological race E09 at the seedling stage (Fig. 6E). In a bonsai experiment in an artificial climate box, the mortality of CS and Shanyou 225 (the susceptible control) was scored as 4. The resistance of Ae. geniculata was scored as 0, which indicated immunity. The W19513 line showed moderate resistance. In a field experiment to assess resistance to stripe rust, four physiological races (CYR31, CYR32, CYR33, and CYR34) of stripe rust were mixed with talc and used as a powder to inoculate plants. Huixianhong (HXH) was used as the susceptible control. The resistance of Ae. geniculata was scored as 0, which thus indicated immunity. The mortality of HXH and CS was scored as 4; the resistance type of the W19513 line was scored as 1, which thus indicated higherresistance (Fig. 6F). These results indicated that the resistance gene in the W19513 line was derived from SY159.
Discussion
Since the founding of the People’s Republic of China in 1949, substantial progress has been achieved in increasing wheat production. From 1949 to 2000, the average wheat yield per hectare has increased from 0.70 tons to 3.86 tons [35]. The International Maize and Wheat Improvement Centre (CIMMYT) views the development of hybrid cultivars as a promising means to boost crop yields because of the growing global demand for wheat [36]. Hybrids can be grown in marginal environments and at low sowing densities because the plants are more vigorous, robust, and stress resistant than pure-line cultivars under marginal conditions [37]. Alien chromosomes, which carry potentially useful agronomic traits, have been introduced into the common background of wheat and have become a useful resource for wheat breeding. In addition, disomic lines are an important bridge for the introduction of desirable alien genes into common wheat [38,39,40].
Ae. geniculata shows excellent resistance to stress and introduction of its favorable traits into common wheat will improve wheat yield and quality. Aegilops and Triticum are closely related genera and show strong genetic similarities [10]. The method of C-banding analysis [41] is used to identify the chromosome of the whole set of Aegilops and its telomeres that are added to the common wheat [42]. Therefore, the transfer of genes from Aegilops species to common wheat can be achieved readily by hybridization. Chromosome 7Mg of SY159 was incorporated as a disomic addition and substitution line in common wheat, which not only enhanced wheat resistance to powdery mildew, but also increased the 1000-grain weight [6, 43]. In practical application, many disease-resistance genes are transferred by the introduction of alien chromosomes, which plays an important role in genetic breeding. Previous experiments have shown that the stripe rust resistance gene Yr40 and the leaf rust resistance gene Lr57 were both derived from chromosome 5Mg of Ae. geniculata [44]. Similarly, the powdery mildew resistance gene pm29 located on chromosome 7D was derived from Ae. geniculata [25]. In the present experiment, assessment of agronomic traits demonstrated that the line W19513 is genetically stable and morphologically intermediate between the parents CS and SY159. The yield of the W19513 line was higher than that of the parents. In addition, the desirable panicle traits are important for improvement of wheat yield, which was previously achieved through the introduction of chromosomes from wild relatives of wheat [45]. With regard to disease resistance, the line W19513 was moderately resistant to race E09 of powdery mildew and highly resistant to four stripe rust races in the field (Fig. 6E, F). These favorable characters are suitable for introduction into wheat. The W19513 line can be used as a donor source to introduce new genes in wheat breeding. In addition, wheat breeding must coordinate the interaction between yield and yield components, which is of great importance for future wheat production and breeding.
Geneticists and an increasing number of plant breeders consider that molecular marker-assisted selection is a valuable tool for germplasm selection to optimize selection efficiency in plant breeding programs [46,47,48]. Molecular markers are based on nucleotide sequence variation between individuals and thus are a direct reflection of genetic polymorphism at the DNA level. Analysis of wheat–Aegilops derivative lines using EST-STS and PLUG molecular markers can distinguish homologous groups of heterogeneous chromosomes and track exogenous chromosomes [49]. In the present study, using these functional molecular markers, we quickly detected that the alien chromosomes belonged to the third homologous group, thus reducing detection time and cost (Fig. 3). At present, the development efficiency of conventional specific markers is low, the cost is high, and an alien chromosome cannot be quickly screened, which causes great inconvenience in breeding [50]. SLAF-seq technology exhibits a high success rate, high specificity and stabilityand low cost for the development of plant chromosome-specific molecular markers. For example, based on SLAF-seq technology, Bourtzis et al. [32] randomly developed 135 pairs of primers from chromosome 7E in Th. elongatum with efficiency of 65.9%. Markers specific to Aegilops biuncialis were developed with a success rate of up to 40.33% [51]. The success rate of 1JS-specific marker development from an alien chromosome of Th. ponticum was 52.98% [52]. The development of SLAF-seq technology has certain practical importance [32]. In the current study, 634 fragments specific for the 3 M chromosome of SY159 were obtained using SLAF-seq technology based on 128 randomly selected fragments. Sixty-one pairs of molecular markers were screened with an effective rate of 47.66%. The development of 3Mg-specific markers with SLAF-seq technology was beneficial for identification of the disomic addition line derived from Ae. geniculata and the development of related probes, which lays a foundation for development of a karyotype map of Ae. geniculata in the future. Interestingly, by applying SLAF-seq technology to chromosome 3Mg of Ae. biuncialis, we observed that the same bands were detected in W19513, SY159, Ae. comosa and Ae. umbellulata but not CS (Fig. 5). We hypothesized that the chromosomes of Ae. comosa and Ae. umbellulata were homologous. It will be extremely valuable to develop many chromosome-specific molecular markers for Ae. geniculata, not only to quickly identify an alien chromosome introduced into common wheat, but also to accelerate the exploration and utilization of its useful agronomic traits or disease resistance in practical wheat production.
FISH is a powerful tool for the identification of distant hybrids, and to elucidate the origin and evolution of allopolyploids [53]. Tang et al. [54] designed 120 oligonucleotide probes based on new tandem repeat sequences, 29 of which showed specific signals on wheat chromosomes. It was important to use new oligonucleotide probes to identify wheat chromosomes or specific segments of wheat chromosomes. We applied GISH technology as described by Shu-Lan et al. [55] with minor modifications. GISH proved that only one pair of alien chromosomes was introduced in the W19513 line and that the chromosomes belonged to the M genome. Further evidence showed the presence and signal morphology of the alien chromosomes. Furthermore, by combining molecular markers, we demonstrated that W19513 contained one additional pair of alien chromosomes designated 3Mg. Chromosomal rearrangement plays an important role in plant evolution. Huang et al. [56] conducted a FISH analysis of CS and 373 Chinese wheat cultivars and observed that 14 structural rearrangements had occurred in 148 (39.7%) cultivars. Badaeva et al. [57] used the C-banding technique to show that chromosomal rearrangement had occurred in 208 tetraploid wheat,252 hexaploid wheat, and 39 triticale accessions. The introduction of alien chromosomes may lead to changes in the structure of chromosomes of common wheat and affect the expression of a suite of genes [52, 58, 59]. Przewieslik-Allen et al. [60] analyzed 471 materials and found that chromosome rearrangement plays an important role in the genetic diversity of varieties. In the present study, FISH analysis suggested that 38 chromosomes were consistent with the standard karyotype map of CS, and variation in 4A and 6B signals were observed. The green signal at the tip of the chromosome 4A long arm was highly weakened and a strong green signal was observed on the short arm of chromosome 6B. These signal changes may be caused by structural rearrangement of wheat chromosomes or polymorphism of FISH signals. It can be inferred from the aforementioned results that chromosomal rearrangement and polymorphism often occur in widely cultivated wheat cultivars and experimental breeding materials and are essential in breeding new cultivars. The introduction of alien chromosomes may also stabilize their structure through chromosomal rearrangement and polymorphism, which can be utilized in genetic breeding. However, the utilization of such rearrangements requires further exploration.
Conclusions
In this research, W19513 was, derived from BC1F10 progeny of a cross between CS and SY159. This line was characterized by molecular cytological, in situ hybridization, functional molecular markers, development specific molecular markers based on SLAF-Seq, morphological and stress resistance identification. The results showed that line W19513 was a wheat– Ae. geniculata 3Mg alien addition line, with excellent endowing characteristics. The line with resistance genes was created, screened and identified by using wild related species of wheat, which could lay a foundation for further shifting of resistance genes to wheat.
Methods
Plant materials
The wheat – Ae. geniculata disomic addition line W19513 was derived from a cross between common wheat ‘Chinese Spring’ (AABBDD, 2n = 42) and Ae. geniculata ‘SY159’ (UUMM, 2n = 28). The F1 progeny were backcrossed with CS, and then the progenies were repeatedly selfed to generate the BC1F10 population. Wheat ‘Shaanyou 225’ was used as the susceptible control for assessment of powdery mildew resistance and ‘Huixianhong’ was used as the susceptible control for assessment of stripe rust resistance. They were all preserved at the College of Agronomy, Northwest A&F University, Yangling, China. Ae. geniculata SY159, Ae. umbellulata (UU, 2n = 2x = 14), and Ae. comosa (MM, 2n = 2x = 14) were provided by Professor Lihui Li and Xinming Yang, Chinese Academy of Agricultural Sciences, Beijing, China.
Cytogenetic analysis
The seeds were soaked in a petri dish with filter paper for one day. The water was poured from the petri dish and then the seeds were placed in an incubator in the dark. After the root length attained 2–3 cm, the root tip was excised and treated with nitrous oxide for ~ 2 h. The root tip was then fixed in 90% acetic acid, 70% ethanol was added, and the root tips were stored at− 20 °C. Following the method of Han et al. [61], the white film was made, and the chromosome number in the root tip cells was observed. In accordance with the method of Wang et al. [43], a typical anther was fixed with Carnoy’s reagent (ethanol:chloroform:acetic acid, 6:3:1) for 24 h prepared in 1% magenta acetate. A fluorescence microscope (BX53, Olympus, Tokyo, Japan) and CCD imaging system were used to observe and photograph mitosis in the root tip cells and meiosis in the pollen mother cells.
FISH, GISH, and sequential FISH–GISH
Following the method of Tang et al. [34], the oligonucleotide probes Oligo-PTa535 (red) and Oligo-pSc119.2 (green) were used to perform FISH. The signal was compared with the existing karyotype map for CS to identify the alien chromosomes. For sequential FISH–GISH analysis, the film prepared for FISH were soaked in alcohol for 2–3 days and then naturally dried. For GISH, genomic DNA of Ae. geniculata was used as a probe and mixed with different concentrations of CS genomic DNA as the blocking DNA. Using a ratio of 1:300, the probe and blocking DNA were hybridized at 56 °C. The fluorescence signal from the chromosomes was observed with a fluorescence microscope (Olympus BX-53). By combining the results from these two techniques, variation in the alien chromosomes could be further determined. To aid with chromosome observation, the chromosomes were stained with the blue-fluorescent DNA stain 4′,6-diamidino-2-phenylindole (DAPI).
Functional molecular markers analysis
Molecular markers are powerful tools for detection of alien chromosomes or fragments [62]. Genomic DNA of all plant materials was extracted using a modified CTAB method [33]. The molecular markers used for homologous group screening were amplified using EST-STS and PLUG primers selected from among the PCR Primers for Grain Genes (http: //wheat.pw.usda.gov/SNP/new/pcr_primers.shtml). The homologous group to which the alien chromosomes belonged was preliminarily determined using this method. The specificity and homoeologous group of the EST-STS primers was assessed by differential separation of the products by polyacrylamide gel electrophoresis according to the DNA size. The PLUG markers were separated by 2% agarose gel electrophoresis in 1% TAE buffer solution; these markers not only showed a high level of polymorphism, but also detected variation and homologous chromosome groups.
Development of specific molecular markers
The SLAF-seq method is a simplified genome sequencing technology independently developed by Biomarker [63] and shows three distinguishing characteristics: (1) deep sequencing to ensure genotyping accuracy, (2) a reduced representation strategy to reduce sequencing costs, and (3) a predesigned reduced representation scheme to optimize marker efficiency [31]. To increase marker specificity and efficiency, sequences that showed 50% similarity to CS (http://www.wheatgenome.org/News/Latest-news/IWGSC-Reference-Sequence-v1.0-browser-now-available-at-URGI) were selected using the BWA tool [51]. From these markers, sequences that showed at least 90% similarity to SY159 were selected. After PCR amplification, specific bands were detected in 1% agarose gel [63]. All primers were synthesized by the Beijing Aoke Ding Sheng Biotechnology Co., Ltd. (Beijing, China).
Assessment of agronomic traits and disease resistance
The plant materials were grown at the Northwest A&F University in late September 2019. The row length was 1.0 m, the row spacing was 0.25 m, and the spacing between plants was 5–10 cm. Ten plants were randomly selected at harvesting in 2020 to compare the agronomic traits of W19513, CS, and SY159. In the field, following the assessment method of Wang et al. [43], we recorded plant height, tiller number, spike length, spikelet number, grain number per spikelet, and spikelet node characters, and the 1000-grain weight and grain width were measured after maturity.
The degree of resistance to powdery mildew was assessed at the seedling stage. Ten plants were planted in each pot with three replicates. Shaanyou 225 was used as the susceptible control. At the two-leaf stage, the plants were inoculated with B. graminis f. sp. tritici physiological race E09 by artificial shaking. About two weeks after inoculation, the resistance to E09 was evaluated in accordance with the method of Wang et al. [64] using a 0–4 scale, 0 = immunity, 0; = near immunity, 1 = highly resistant, 2 = moderately resistant, 3 = moderately susceptible, and 4 = highly susceptible. The assessment of resistance to stripe rust in the field was conducted in the experimental field of the College of Agronomy, Northwest A&F University. The physiological races CYR31, CYR32, CYR33, and CYR34 were mixed in equal proportions with talc 1:1:1:1. The mixture of strains was used to inoculate plants by shaking the powder onto the leaves. HXH was used as the susceptible control. The reaction type was recorded in accordance with the standard 0–4 scale [65].
Availability of data and materials
All data and materials are available in the article.
Abbreviations
- Pst :
-
Puccinia striiformis f. sp. Tritici
- SLAF-seq:
-
specific-locus amplified fragment sequencing
- CS:
-
Chinese Spring
- Ae. geniculate :
-
Aegilops geniculate
- CTAB:
-
cetyltrimethylammonium bromide
- GISH:
-
genomic in situ hybridization
- EST-STS:
-
expressed sequence tag–sequence-tagged site
- PLUG:
-
PCR-based landmark unique gene
- BWA:
-
Burrows–Wheeler Alignment
- FISH:
-
fluorescence in situ hybridization
- HXH:
-
Huixianhong
- CIMMYT:
-
International Maize and Wheat Improvement Centre
- DAPI:
-
4′,6-diamidino-2-phenylindole.
References
Peña RJ, Trethowan R, Pfeiffer WH, Ginkel MV. Quality (end-use) improvement in wheat. J Crop Prod. 2008;5(1–2):1–37.
Brinton J, Uauy C. A reductionist approach to dissecting grain weight and yield in wheat. J Integr Plant Biol. 2019;61(3):337–58.
You-Liang YE, Han YL, Tan JF. Cui ZLJJoTC: wheat production and fertilizer application in China. Journal of Triticeae Crops. 2007:127–33.
Liu Y, Pan X, Li J. Current agricultural practices threaten future global food production. J Agric Environ Ethics. 2014;28(2):203–16.
Li Z, Li B, Tong Y. The contribution of distant hybridization with decaploid Agropyron elongatum to wheat improvement in China. J Genet Genomics. 2008;35(8):451–6.
Wang Y, Long D, Wang Y, Wang C, Liu X, Zhang H, Tian Z, Chen C, Ji W: Characterization and Evaluation of Resistance to Powdery Mildew of Wheat-Aegilops geniculata Roth 7M(g) (7A) Alien Disomic Substitution Line W16998. Int J Mol Sci 2020, 21(5).
Tiancai G. Countermeasures and suggestions for the development of wheat production in China. Review of China Agricultural Science. 2001;3(04):27–31.
Yang Y, Zhang X. Cost-benefit analysis of wheat production in Shandong Province. Agricultural Outlook. 2017:36–9.
Schneider A, Molnár I, Molnár-Láng M. Utilisation of Aegilops (goatgrass) species to widen the genetic diversity of cultivated wheat. Euphytica. 2007;163(1):1–19.
Bowden MW. The taxonomy and nomenclature of the wheats, barleys and ryes and their wild relatives. Can J Bot. 1959;37(4):657–84.
Zhang XY, Wang RRC, Dong YS. RAPD polymorphisms in Aegilops geniculata Roth (Ae-ovata auct non L). Genet Resour Crop Evol. 1996;43(5):429–33.
Cui L, Ren YK, Murray TD, Yan WZ, Guo Q, Niu YQ, et al. Development of perennial wheat through hybridization between wheat and wheatgrasses: a review. Engineering. 2018;4(4):507–13.
Schneider A, Molnar I, Molnar-Lang M. Selection of U and M genome-specific wheat SSR markers using wheat-Aegilops biuncialis and wheat-Ae. geniculata addition lines. Euphytica. 2010;175(3):357–64.
Molnar I, Simkova H, Leverington-Waite M, Goram R, Cseh A, Vrana J, et al. Syntenic relationships between the U and M genomes of Aegilops, wheat and the model species Brachypodium and rice as revealed by COS markers. PLoS One. 2013;8(8):e70844.
Schneider A, Molnárláng M. Detection of various U and M chromosomesin wheat-Aegilops biuncialis hybrids and derivatives using fluorescence in situ hybridisation and molecular markers. Journal of Genetics Plant Breeding. 2012;48(4):169–77.
Siddiqui KA, Yousufzai MN. Natural and induced variation for endomorphic traits in the tribe Triticeae. In: Proceedings of the Seventh International Wheat Genetics Symposium. Cambridge: Published by the Institute of Plant Science Research, Cambridge Laboratory; 1988. p. 139–43.
Zaharieva M, Monneveux P, Henry M, Rivoal R, Valkoun J, Nachit MM. Evaluation of a collection of wild wheat relative Aegilops geniculata Roth and identification of potential sources for useful traits. Euphytica. 2001;119(1–2):33–8.
Slageren MV. Wild wheats: a monograph of Aegilops L. and Amblyopyrum (Jaub. & Spach) Eig (Poaceae). ICARDA / In.; 1994: 94(94).
Ullah KN, Li N, Shen T, Wang P, Tang W, Ma S, et al. Fine mapping of powdery mildew resistance gene Pm4e in bread wheat (Triticum aestivum L.). Planta. 2018;248(5):1319–28.
Yang FP, Yang WX, Shang XW. A study on the effect of wheat stripe rust on wheat yield. Journal of Gansu Agricultural University. 2008;43(2):51–5.
Everts KL, Leath S. Effect of early season powdery mildew on development, survival, and yield contribution of tillers of winter-wheat. Phytopathology. 1992;82(11):1273–8.
Li T, Zhang Z, Hu Y, Duan X, Xin Z. Identification and molecular mapping of a resistance gene to powdery mildew from the synthetic wheat line M53. J Appl Genet. 2011;52(2):137–43.
Ralph R, Victor C, Roy J. The incorporation of alien disease resistance in wheat by genetic interference with the regulation of meiotic chromosome synapsis. Genet Res. 1968;12(2):199–219.
Sears ER. The transfer of leaf- rust resistance from Aegilops umbellulate to wheat. Brookhaven Symp Biol. 1956;9.
Zeller FJ, Kong L, Hartl L, Mohler V, Hsam SLK. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.) 7. Gene Pm29 in line Pova. Euphytica. 2002;123(2):187–94.
Friebe B, Jiang J, Raupp WJ, McIntosh RA, Gill BS. Characterization of wheat-alien translocations conferring resistance to diseases and pests: current status. Euphytica. 1996;91(1):59–87.
Varshney RK, Terauchi R, McCouch SR. Harvesting the promising fruits of genomics: applying genome sequencing technologies to crop breeding. PLoS Biol. 2014;12(6):e1001883.
Okada M, Yoshida K, Nishijima R, Michikawa A, Motoi Y, Sato K, et al. RNA-seq analysis reveals considerable genetic diversity and provides genetic markers saturating all chromosomes in the diploid wild wheat relative Aegilops umbellulata. BMC Plant Biol. 2018;18(1):271.
Tiwari VK, Wang S, Danilova T, Koo DH, Vrana J, Kubalakova M, et al. Exploring the tertiary gene pool of bread wheat: sequence assembly and analysis of chromosome 5M(g) of Aegilops geniculata. Plant J. 2015;84(4):733–46.
Elshire RJ, Glaubitz JC, Sun Q, Poland JA, Kawamoto K, Buckler ES, et al. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS One. 2011;6(5):e19379.
Sun X, Liu D, Zhang X, Li W, Liu H, Hong W, et al. SLAF-seq: an efficient method of large-scale de novo SNP discovery and genotyping using high-throughput sequencing. PLoS One. 2013;8(3):e58700.
Chen S, Huang Z, Dai Y, Qin S, Gao Y, Zhang L, et al. The development of 7E chromosome-specific molecular markers for Thinopyrum elongatum based on SLAF-seq technology. PLoS One. 2013;8(6):e65122.
Doyle J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 1987;19.
Tang Z, Yang Z, Fu S. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J Appl Genet. 2014;55(3):313–8.
He ZH, Rajaram S, Xin ZY, Huang GZ. A history of wheat breeding in China. J Comp Neurol. 2001;523(5):805–13.
Reynolds MP, Beem JV, Ginkel MV, Hoisington DA. Breaking the yield barriers in wheat: a brief summary of the outcomes of an international consultation. In: CIMMYT; 1997. p. 1–10.
Jordaan JP, Engelbrecht SA, Malan JH, Knobel H.A.: Wheat and Heterosis; 1999.
Du WL, Wang J, Lu M, Sun SG, Chen XH, Zhao JX, et al. Molecular cytogenetic identification of a wheat-Psathyrostachys huashanica Keng 5Ns disomic addition line with stripe rust resistance. Mol Breed. 2013;31(4):879–88.
Wang G, Ji J, Wang YB, Hu H, King IP, Snape JW. The genetic characterisation of novel multi-addition doubled haploid lines derived from triticale x wheat hybrids. Theor Appl Genet. 1993;87(5):531–6.
Shin T, TJBe K. Production and characterization of a complete set of wheat-wild barley (Hordeum vulgare ssp. spontaneum) chromosome addition lines. Breed Sci. 2001;51(3):199–206.
Gill BS, Friebe B, Endo TR. Standard karyotype and nomenclature system for description of chromosome bands and structural aberrations in wheat ( Triticum aestivum ). Genome. 1991;34(5):830–9.
Friebe BR, Tuleen NA, Gill BS. Development and identification of a complete set of Triticum aestivum Aegilops geniculata chromosome addition lines. Genome. 1999;42(3):374–80.
Wang Y, Quan W, Peng N, Wang C, Yang X, Liu X, et al. Molecular cytogenetic identification of a wheat–Aegilops geniculata Roth 7Mg disomic addition line with powdery mildew resistance. Mol Breed. 2016;36(4).
Kuraparthy V, Chhuneja P, Dhaliwal HS, Kaur S, Bowden RL, Gill BS. Characterization and mapping of cryptic alien introgression from Aegilops geniculata with new leaf rust and stripe rust resistance genes Lr57 and Yr40 in wheat. Theor Appl Genet. 2007;114(8):1379–89.
Wu J, Yang X, Wang H, Li H, Liu W. The introgression of chromosome 6P specifying for increased numbers of florets and kernels from Agropyron cristatum into wheat. Theoretical Applied Genetics. 2007;114(1):13–20.
Xu YB, Crouch JH. Marker-assisted selection in plant breeding: from publications to practice. Crop Sci. 2008;48(2):391–407.
Dwivedi SL, Crouch JH, Mackill DJ, Xu Y, Blair MW, Ragot M, et al. The molecularization of public sector crop breeding: Progress, problems, and prospects. Adv Agron. 2007;95:163–318.
Dwivedi SL, Crouch JH, Mackill DJ, Xu Y, Blair MW, Ragot M, et al. The Molecularization of public sector crop breeding: Progress, problems, and prospects. In: Advances in Agronomy; 2007. p. 163–318.
Kong LN, Li Q, Wang HY, Cao AZ, Chen PD, Wang XE. Molecular marker analysis of wheat-Roegneria ciliaris additions lines. Yi Chuan. 2008;30(10):1356–62.
Li XM, Lee BS, Mammadov AC, Koo BC, Mott IW, Wang RR. CAPS markers specific to Eb, Ee, and R genomes in the tribe Triticeae. Genome. 2007;50(4):400–11.
Song L, Zhao H, Zhang Z, Zhang S, Liu J, Zhang W, et al. Molecular Cytogenetic Identification of Wheat-Aegilops Biuncialis 5M(b) Disomic Addition Line with Tenacious and Black Glumes. Int J Mol Sci. 2020;21(11).
Wang Y, Cao Q, Zhang J, Wang S, Chen C, Wang C, et al. Cytogenetic analysis and molecular marker development for a new wheat-Thinopyrum ponticum 1J(s) (1D) disomic substitution line with resistance to stripe rust and powdery mildew. Front Plant Sci. 2020;11:1282.
Mukai Y, Endo TR, Gill BS. Physical mapping of the 5S rRNA multigene family in common wheat. Heredity. 1990;4:4.
Tang S, Tang Z, Qiu L, Yang Z, Li G, Lang T, et al. Developing New Oligo Probes to Distinguish Specific Chromosomal Segments and the A, B, D Genomes of Wheat (Triticum aestivum L.) Using ND-FISH. Front Plant Sci. 2018;9:1104.
Fu SL, Yang MY, Ren ZL, Yan BJ, Tang ZX. Abnormal mitosis induced by wheat-rye 1R monosomic addition lines. Genome. 2014;57(1):21–8.
Huang X, Zhu M, Zhuang L, Zhang S, Wang J, Chen X, et al. Structural chromosome rearrangements and polymorphisms identified in Chinese wheat cultivars by high-resolution multiplex oligonucleotide FISH. Theor Appl Genet. 2018;131(9):1967–86.
Badaeva ED, Dedkova OS, Gay G, Pukhalskyi VA, Zelenin AV, Bernard S, et al. Chromosomal rearrangements in wheat: their types and distribution. Genome. 2007;50(10):907–26.
Rey E, Abrouk M, Keeble-Gagnere G, Karafiatova M, Vrana J, Balzergue S, et al. Transcriptome reprogramming due to the introduction of a barley telosome into bread wheat affects more barley genes than wheat. Plant Biotechnol J. 2018;16(10):1767–77.
Wang SW, Wang CY, Wang YZ, Wang YJ, Chen CH, Ji WQ. Molecular cytogenetic identification of two wheat-Thinopyrum ponticum substitution lines conferring stripe rust resistance. Mol Breed. 2019;39(11):143.
Przewieslik-Allen AM, Wilkinson PA, Burridge AJ, Winfield MO, Dai X, Beaumont M, et al. The role of gene flow and chromosomal instability in shaping the bread wheat genome. Nat Plants. 2021;7(2):172–83.
Han F, Lamb JC, Birchler JA. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc Natl Acad Sci U S A. 2006;103(9):3238–43.
Ma ZQ, Sorrells ME, Tanksley SD. RFLP markers linked to powdery mildew resistance genes Pm1, Pm2, Pm3, and Pm4 in wheat. Genome. 1994;37(5):871–5.
Liu L, Luo Q, Teng W, Li B, Li H, Li Y, et al. Development of Thinopyrum ponticum-specific molecular markers and FISH probes based on SLAF-seq technology. Planta. 2018;247(5):1099–108.
Wang Y, Wang C, Quan W, Jia X, Fu Y, Zhang H, et al. Identification and mapping of PmSE5785, a new recessive powdery mildew resistance locus, in synthetic hexaploid wheat. Euphytica. 2015;207(3):619–26.
Bariana HS, Mcintosh RA. Cytogenetic studies in wheat. XV. Location of rust resistance genes in VPM1 and their genetic linkage with other disease resistance genes in chromosome 2A. Genome. 1993;36(3):476–82.
Acknowledgments
We thank PhD Robert McKenzie for editing the English text of a draft of this manuscript.
Funding
This work was supported by the project of science and technology of Shaanxi province of China (2021NY-081), and Crop Germplasm Resources Protection (No. 2019NWB036–02-1).
Author information
Authors and Affiliations
Contributions
Y.W. (Yongfu Wang), Y.W. (Yajuan Wang) and W.J. designed the study, analyzed the data and wrote the article. Y.W. (Yajuan Wang) and Y.W. (Yongfu Wang) contributed to the development of material. X.C. and X.Y contributed to cytological analysis. P.D contributed to the SLAF-seq data analysis. C.W., H.Z., X.L. and C.C contributed to powdery mildew resistance and agronomic trait evaluation. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, Y., Cheng, X., Yang, X. et al. Molecular cytogenetics for a wheat–Aegilops geniculata 3Mg alien addition line with resistance to stripe rust and powdery mildew. BMC Plant Biol 21, 575 (2021). https://doi.org/10.1186/s12870-021-03360-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-021-03360-4