
Abstract
Background
The search for quantitative trait loci (QTL) affecting traits of interest in mammals is frequently limited to autosomes, with the X chromosome excluded because of its hemizygosity in males. This study aimed to assess the importance of the X chromosome in the genetic determinism of 11 complex traits related to milk production, milk composition, mastitis resistance, fertility, and stature in 236,496 cows from three major French dairy breeds (Holstein, Montbéliarde, and Normande) and three breeds of regional importance (Abondance, Tarentaise, and Vosgienne).
Results
Estimates of the proportions of heritability due to autosomes and X chromosome (h²X) were consistent among breeds. On average over the 11 traits, h²X=0.008 and the X chromosome explained ~ 3.5% of total genetic variance. GWAS was performed within-breed at the sequence level (~ 200,000 genetic variants) and then combined in a meta-analysis. QTL were identified for most breeds and traits analyzed, with the exception of Tarentaise and Vosgienne and two fertility traits. Overall, 3, 74, 59, and 71 QTL were identified in Abondance, Montbéliarde, Normande, and Holstein, respectively, and most were associated with the most-heritable traits (milk traits and stature). The meta-analyses, which assessed a total of 157 QTL for the different traits, highlighted new QTL and refined the positions of some QTL found in the within-breed analyses. Altogether, our analyses identified a number of functional candidate genes, with the most notable being GPC3, MBNL3, HS6ST2, and DMD for dairy traits; TMEM164, ACSL4, ENOX2, HTR2C, AMOT, and IRAK1 for udder health; MAMLD1 and COL4A6 for fertility; and NRK, ESX1, GPR50, GPC3, and GPC4 for stature.
Conclusions
This study demonstrates the importance of the X chromosome in the genetic determinism of complex traits in dairy cattle and highlights new functional candidate genes and variants for these traits. These results could potentially be extended to other species as many X-linked genes are shared among mammals.
Similar content being viewed by others


Background
Since the late 2000s, significant efforts have been made to decipher the genetic determinism of complex traits using genome-wide association studies (GWAS) or meta-analyses of GWAS results in different species, in particular in humans [1]. To date, the largest GWAS have focused on human height, which is a highly heritable and easily measured trait and thus one for which very large datasets are available [2, 3]. Despite all these efforts, in all human GWAS results published so far—including a very recent study that examined human height in more than five million individuals [3]—a portion of the heritability of the studied traits remains unexplained [4, 5]. The percentage differs among studies, but even small amounts of unexplained variability can be detrimental to our understanding of the biological mechanisms underlying traits of interest. Furthermore, a lack of information on some of the genetic variants associated with complex traits, such as some diseases for example, can decrease the accuracy of predictions of genetic risk at the population or individual level [6].
In livestock species, the bovine genome was one of the first to be sequenced, in 2009 [7]. As in humans, considerable efforts have been made in the last decade to discover the genes and genomic variants that are involved in the genetic determinism of various complex traits of interest [8, 9], including stature, for which a large-scale meta-analysis was conducted [10] as part of the 1000 Bull Genomes project [11]. One of the expected benefits in cattle, and in livestock species in general, is improving the prediction of complex traits for breeding programs [12].
However, in most GWAS conducted in mammals, and in particular in the large-scale GWAS of stature in cattle [10] and humans [13], the focus has mainly been on autosomes, with the X chromosome often excluded due to its unique mode of inheritance. Genetic studies of the X chromosome are complicated by two factors: male hemizygosity (XY) in the non-pseudoautosomal region (non-PAR), which covers the majority of the X chromosome in mammals [14, 15], and dosage compensation, i.e., the inactivation of one X chromosome, in XX females during early development, which ensures equivalent X-linked gene expression in cells of animals of both sexes [16]. Although it has been much less studied than in humans or model species, the phenomenon of dosage compensation has also been demonstrated in cattle [17]. Consequently, in order to include the X chromosome in a genetic study, researchers must use different or separate treatments for bulls and cows and make assumptions about dosage compensation [18, 19], which is rarely done. However, the X chromosome is the second-largest chromosome in the bovine genome and contains 1132 annotated genes (Ensembl release 107 - Jan 2023 [20]), i.e., more than 4% of all annotated genes in the entire genome, with most (1098) located in the non-PAR region. Therefore, the exclusion of the X chromosome from efforts to decipher the genetic determinism of complex traits or to improve the breeding values of animals may mean that a significant number of relevant genes are missed, and may result in a loss of efficiency in genomic selection [19].
In France, programs of genomic selection have been active since 2009 for the national dairy breeds Montbéliarde, Normande, and Holstein [21], and since 2016 for the regional dairy breeds Abondance, Tarentaise, and Vosgienne [22]. As a result, data are available for several thousand to several hundred thousand cows in these breeds, including phenotypes for various complex traits of economic interest as well as genotypes from the bovine EuroGMD chip [23]. Although X-chromosome variants have not yet been incorporated into genomic prediction equations for estimating the breeding values of animals, more than 1000 SNPs located in this region are included on the bovine Illumina EuroGMD chip. Here, we leveraged this large dataset to assess the relative importance of the X chromosome in the genetic determinism of complex traits in dairy cattle and to identify candidate causative X-linked genes and variants.
Specifically, we investigated 11 traits related to milk production, mastitis resistance, fertility, and stature that were measured in 236,496 cows of the six breeds (Montbéliarde, Normande, Holstein, Abondance, Tarentaise, and Vosgienne), with three objectives: [1] to estimate the respective proportions of heritability due to autosomes and the X chromosome in each breed and for each trait, [2] to perform within-breed association analyses that evaluated the effects of X-chromosome variants on each trait using genotypes imputed at the sequence level, and [3] for each trait, to combine the results obtained in the six different breeds in a meta-analysis.
Results
In the six breeds examined here— Montbéliarde, Normande, Holstein, Abondance, Tarentaise, and Vosgienne —a number of traits are routinely measured for breeding, including traits related to milk production and composition (milk yield (MY), protein yield (PY), fat yield (FY), protein content (PC), and fat content (FC)), udder health (clinical mastitis (MAST) and somatic cell score (SCS)), fertility (interval between calving and first fertilizing artificial insemination (ICFI), heifers’ conception rate (HCR), and lactating cows’ conception rate (CCR)), and stature (STAT). To complement these phenotypes, genotypes are available for a subset of cows, typically based on one of several medium-density SNP chips used in the last 12 years, especially the Illumina EuroGMD chip. The number of cows for which both phenotypes and genotypes are available varies greatly among breeds, from 2555 cows in Vosgienne to 81,815 cows in Holstein (Table 1). Note that, due to computational limitations, we restricted the sample size of the two largest breeds by randomly sampling 61,881 Montbéliarde and 81,815 Holstein cows from the available 162,419 Montbéliarde and 315,674 Holstein cows with both phenotypes and genotypes.
Heritability due to autosomes and X chromosome
To estimate heritabilities due to autosomes (h²AUT) and the X chromosome (h²X), we first used a restricted maximum likelihood (REML) approach that included two random polygenic effects estimated using genomic relationship matrices (GRM), which were derived from 53,469 autosomal and 1147 X-linked SNPs, respectively. In all breeds, the most heritable traits were STAT (0.42–0.72) and milk composition traits, i.e., PC (0.52–0.75) and FC (0.52–0.72), while MAST and fertility traits (ICFI, HCR, and CCR) were the least heritable (0.01–0.09). SCS, MY, PY, and FY presented intermediate heritability, with h² estimates ranging from 0.22 to 0.40 (Table 2). For all breeds and traits, h²AUT ranged from 0.008 (ICFI in Vosgienne) to 0.74 (PC in Vosgienne), while h²X ranged from 0.000 (MAST, HCR, and/or CCR in multiple breeds; STAT in Tarentaise) to 0.04 (PC in Tarentaise). For the most-heritable traits, h² was mainly explained by autosomes; instead, the proportion of genetic variance explained by the X chromosome was higher for SCS, HCR, and CCR in Tarentaise and for ICFI in Vosgienne. The standard error of estimates ranged from 0.002 to 0.035 for h²AUT and from 0.0004 to 0.019 for h²X. On average across all breeds, around 3.5% of the total genetic variance of each trait was explained by the X chromosome.
Within-breed linkage disequilibrium on X chromosome
Linkage disequilibrium data, calculated for chromosomes 2 and X in the 6 breeds, revealed an expected decline in r2 as the distance between markers increased (Fig. S1). Remarkably, the X chromosome generally exhibited higher linkage disequilibrium levels irrespective of marker distance. Moreover, regional breeds (Abondance, Tarentaise, and Vosgienne) exhibited higher levels of linkage disequilibrium, consistent with their lower effective population size.
Within-breed and meta-analyses of association
For all cows with both genotypes and phenotypes, sequence-level genotypes were imputed from the 50k EuroGMD genotypes via an intermediate HD density step; details on the imputation procedure can be found in the Materials and Methods. A multibreed sample of 2712 sequenced bulls, which included between 4 (Vosgienne) and 1019 (Holstein) bulls of each of the six breeds, was used as a reference for sequence-level imputation. After we removed variants with a MAF lower than 0.005 and an imputation accuracy lower than 0.2 (as assessed by the R squared (R²) value generated by Minimac software [24]), the mean imputation accuracy ranged from 0.69 for Abondance to 0.82 for Normande (Table 3).
For each trait, we first conducted within-breed association analyses and then combined the within-breed results in a meta-analysis using the fixed effects method. The number of QTL and their confidence intervals were defined from both within-breed and meta-analysis results using an iterative procedure that evaluated linkage disequilibrium (LD) between the variant with the most significant effect, referred to as the lead variant, and variants located ± 10 Mbp around the lead variant (Fig. 1). The most plausible candidate genes were identified by considering the location of variants with the most significant effects in the QTL peaks.

Iterative procedure for defining QTL and their confidence intervals
Within-breed association analyses detected variants with significant effects (-log10(P) ≥ 7.3) in all breeds except Tarentaise and Vosgienne, and for all traits except HCR and CCR (Table 4, Table S2, Figs. S1 and S2). The number of QTLs identified in each breed varied widely, with 3 QTL detected in Abondance compared to 59, 71, and 72 QTL in Normande, Holstein, and Montbéliarde, respectively. The number of breed x trait combinations with significant effects also varied greatly depending on the trait analyzed: for example, 1 QTL was identified for MAST (in Abondance), while 19 were found for SCS (4, 4, and 11 in Montbéliarde, Normande and Holstein, respectively) and 35 for STAT (8, 13, and 14 in Montbéliarde, Normande and Holstein, respectively). Meta-analyses detected QTL for 8 of the 11 traits: 9 for PC, 15 for SCS and ICFI, 23 for PY, 24 for FY, 26 for STAT, and 27 for MY, but none for MAST, HCR, or CCR (Table 4, Table S3, Figs. S1 and S2).
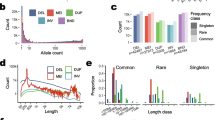
Features of all the QTL detected in within-breed analyses (Table S2) and in meta-analyses (Table S3) are summarized in Table 5. On average, the -log10(P) of the lead variant of a QTL was higher in Holstein, Montbéliarde, and the meta-analyses (13.8–14.2) than in Normande (11.2) and Abondance (8.6). Both in terms of the number of variants included and the length in Mbp, the average size of the confidence intervals of QTL was higher in Abondance (5.7 Mbp and 733 variants) and Montbéliarde (5.0 Mbp and 274 variants), lower in Holstein (2.8 Mbp and 151 variants) and Normande (2.5 Mbp and 96 variants), and lowest in the meta-analyses (1.7 Mbp and 65 variants). For each trait, Fig. 2 depicts the numbers of variants located within the confidence intervals of the QTL that were shared between the different within-breed analyses and the meta-analyses.

UpSet diagrams for variants within the confidence intervals of QTL detected in within-breed and meta-analyses of association. (a) milk yield (MY), (b) fat yield (FY), (c) protein yield (PY), (d) fat content (FC), (e) protein content (PC), (f) somatic cell score (SCS), (g) interval between calving and first insemination (ICFI), and (h) stature (STAT).
Functional annotation of the lead variants for all QTL revealed that, both in within-breed and meta-analyses, the majority were intergenic (121 and 108, respectively) or intronic (61 and 34, respectively). Less frequently, lead variants were located in upstream regions (15 and 6, respectively), downstream regions (5 and 6, respectively), exons (missense: 2 and 1, respectively; synonymous: 2 in within-breed analyses), and 3’UTR regions (1 in within-breed analyses). The missense lead variants affecting ENSBTAG00000006384 (FC in Montbéliarde), NRK (STAT in Normande), and PLXNB3 (STAT in meta-analyses) genes were predicted to have SIFT moderate effects.
Milk production and composition
Across all of the within-breed association analyses, the QTL with the most significant effect was identified for PC in Montbéliarde, with the lead variant located at 15,665,045 bp (-log10(P) = 97.5). In the three other breeds for which QTL were detected, as well as in the meta-analyses, the QTL with the most significant effect was also found for PC, with the lead variant located at 17,846,562 bp in Abondance (-log10(P) = 10.4), 17,977,632 bp in Holstein (-log10(P) = 51.2), 16,169,349 bp in Normande (-log10(P) = 17.1), and 16,429,402 bp in meta-analyses (-log10(P) = 111.9) (Fig. 3). The confidence intervals of these QTLs were small in the meta-analysis (50 kbp with 19 variants), Holstein, and Normande (~ 200 kbp with 18 and 93 variants, respectively), but larger in Montbéliarde (1.9 Mbp with 73 variants) and Abondance (6.2 Mbp with 440 variants). Within the confidence intervals, the respective number of positional candidate genes was 2, 6, 4, 7, and 26. In all analyses but Holstein, the lead variant was located in an intergenic region; in Holstein, instead, it was located in the upstream region of the MIR363 gene. In the vicinity of the lead intergenic variants, we identified the genes GPC3, Metazoa_SRP, OR13H1, ENSBTAG00000051508, RAP2C, MBNL3, and HS6ST2 (Table S2). Across the different analyses, numerous other QTL were detected for PC (Fig. 2) and other milk production (MY, PY, and FY) and composition (FC) traits (Tables S2 and S3; Figs. S1 and S2). Similar to the QTL with the most significant effect on PC, several QTL detected for a given milk trait in different breeds were identified in neighboring regions, but only a few had their lead variants located in the same gene or close to the same gene in different breeds (Tables S2 and S3; Fig. 3). We note three in particular: (1) a QTL for FC detected in Normande and Montbéliarde with the lead variants located in an intronic region of HS6ST2 (at 16,679,506 bp) and 250-kb upstream (at 16,420,927 bp), respectively; (2) a QTL for FY in both Normande and Holstein with lead variants located in introns of DMD, at 110,665,296 and 111,032,759 bp, respectively; and (3) a QTL for PY found in Holstein and Normande with the lead variants located in the same intergenic region between DDX53 and ENSBTAG00000049480, at 120,448,964 and 120,590,087 bp, respectively. Interestingly, HS6ST2, DMD, and DDX53 were also the most plausible positional candidate genes for QTL detected in the meta-analyses for FC, FY, and PY, respectively, but with different lead variants, located at 16,409,330 bp (intergenic), 110,665,443 bp (intronic), and 119,923,321 bp (intergenic), respectively.

Results of within-breed and meta-analyses of the X chromosome for protein content (PC): Manhattan plot for the entire chromosome and LocusZoom graph for the QTL with the most significant effects. Within-breed association analyses in Abondance, Montbéliarde, Normande, and Holstein cows (Manhattan plot in blue); fixed effects meta-analyses (Manhattan plot in gray, variants with effects in the same direction in all within-breed analyses are highlighted in green); and corresponding LocusZoom graphs for the 20-Mb interval centered around the variant with the most significant effect
Udder health
While only one QTL was found for MAST (in Abondance), we detected 4, 4, and 11 QTL for SCS in Montbéliarde, Normande, and Holstein, respectively. These QTL were located in different regions in the different breeds, with ACSL4, TMEM164, LHFPL1, AMOT, and ENOX2 as the most plausible positional candidate genes. Similarly, meta-analyses did not detect any QTL for MAST but did reveal 15 distinct QTL regions for SCS, with the one with the most significant effect located at 63.1 Mbp (intergenic lead variant between bta-mir-1911 and HTR2C). In the vicinity of this region we also found the QTL with the most significant effects in Montbéliarde, with the lead variant located at 61,232,885 bp, 20 kbp downstream AMOT; however, the meta-analysis peak appeared much narrower, with a confidence interval of 671 kbp compared to 11.8 Mbp in Montbéliarde (Fig. 4). Other positional candidate genes that were identified in the meta-analyses for SCS included ATG4A, TMEM47, ENSBTAG00000012533, TMEM187, IRAK1, PPP1R2C, and PPP4R3C.

Results of within-breed and meta-analyses of the X chromosome for somatic cell score (SCS): Manhattan plot for the entire chromosome and LocusZoom graph for the QTL with the most significant effects. Within-breed association analyses in Montbéliarde, Normande, and Holstein cows (Manhattan plot in blue); fixed effects meta-analyses (Manhattan plot in gray, variants with effects in the same direction in all within-breed analyses are highlighted in green); and corresponding LocusZoom graphs for the 20-Mb interval centered around the variant with the most significant effect
Fertility
ICFI was the only fertility trait for which we detected QTL: 7 in Montbéliarde, 3 in Normande, 1 in Holstein, and 15 in the meta-analyses. As we observed in the SCS analyses, although the confidence intervals of some QTL overlapped between Montbéliarde and Normande, the lead variants were distinct. In addition, the QTL with the most significant effects were located in different regions in Montbéliarde, Normande, and Holstein (Fig. 5). In the within-breed analyses, the most significant QTL were highlighted in Montbéliarde: these included one intronic variant in MAMLD1 and two intergenic variants between ENSBTAG00000054511 and PRR32 and between ENSBTAG00000050383 and ENSBTAG00000042114, respectively. The meta-analysis revealed new QTL regions that were not found in within-breed analyses, with the most significant lead variant located between ENOX2 and ARHGAP36.

Results of within-breed and meta-analyses of the X chromosome for interval between calving and first insemination (ICFI): Manhattan plot for the entire chromosome and LocusZoom graph for the QTL with the most significant effects. Within-breed association analyses in Montbéliarde, Normande, and Holstein cows (Manhattan plot in blue); fixed effects meta-analyses (Manhattan plot in gray, variants with effects in the same direction in all within-breed analyses are highlighted in green); and corresponding LocusZoom graphs for the 20-Mb interval centered around the variant with the most significant effect
Stature
STAT was the trait for which the highest number of QTL was detected: 8 in Montbéliarde, 13 in Normande, 14 in Holstein, and 26 in the meta-anayses (Fig. 6). In Normande, Montbéliarde, and Holstein, the QTL with the most significant effects had their lead variant located in the downstream region of IDS, in an intron of COL4A6, and in the intergenic region between ENSBTAG00000000567 and ENSBTAG00000052795, respectively. The COL4A6 gene was also the most plausible positional candidate gene for another QTL detected in Normande, with a synonymous lead variant. Another gene, TRPC5, was highlighted in two breeds, but with two different intronic lead variants, at 60,006,622 bp in Normande and at 60,092,981 bp in Holstein. The most significant QTL detected in the meta-analysis had an intergenic lead variant located at 53,156,112 bp, between ESX1 and ENSBTAG00000053380, and a large confidence interval (52.2–63.1 Mbp) that contained different QTLs than those found in the within-breed analyses, in particular those located near COL4A6 and TRPC5. Certain candidate genes highlighted for STAT were also identified for other traits: two for PC (MBLN3 and GPC3) and one for SCS (PPP4R3C). Finally, as mentioned above, the missense variants in NRK (g.54,703,522 C > T, p.714R > W) and PLXNB3 (g.36994420G > A, p.1600 > H) genes were also noteworthy because for each of the QTL for which they were positional candidates, they are the variants with the most significant effect on STAT.

Results of within-breed and meta-analyses of the X chromosome for stature (STAT): Manhattan plot for the entire chromosome and LocusZoom graph for the QTL with the most significant effects in meta-analyses. Within-breed association analyses in Montbéliarde, Normande, and Holstein cows (Manhattan plot in blue); fixed effects meta-analyses (Manhattan plot in gray, variants with effects in the same direction in all within-breed analyses are highlighted in green); and corresponding LocusZoom graphs for the 20-Mb interval centered around the variant with the most significant effect in both the within-breed Montbéliarde analysis and meta-analysis
Discussion
In mammals, the non-PAR region of the X chromosome differs from autosomes because it is hemizygous in males (XY). In females (XX), mechanisms of dosage compensation inactivate one copy of the X chromosome and thereby ensure the equal expression of these genes in both sexes. The complex mechanisms of X chromosome inactivation have been less extensively studied in cows than in other mammalian species but, in the absence of mutations affecting the Xist locus [25], both copies appear to be equally expressed in the mammary gland, which suggests random inactivation of either copy [17]. Because of these unique features, the X chromosome is usually excluded from genetic analyses. In cattle, previous efforts to test for genetic associations with the X chromosome examined reproductive traits in beef cattle at the 50k [26] and the HD [27,28,29,30,31] level. In the present study, instead, we focused on how the X chromosome affects the genetic determinism of 11 complex traits related to milk production and composition, udder health, fertility, and stature in six dairy cattle breeds. For each trait, we first estimated the proportion of genetic variance explained by this chromosome and then identified potential X-linked candidate genes and variants at the sequence level through within-breed and meta-analyses of association.
For the different traits and the different breeds, overall heritabilities—as estimated with genomic relationship matrices derived from EuroGMD autosomal and X-linked SNPs—were similar or higher to previous estimations obtained from pedigrees [32]. For each trait, the phenotypes used were yield deviations, i.e., mean performances adjusted for environmental effects. One potential issue with this approach is that, when records are repeated, the phenotype has a reduced residual variance and therefore a higher heritability. However, this phenomenon applies to all chromosomes equally and does not affect the proportion of genetic variance explained by the X chromosome.
Depending on the breed and trait in question, the gain in heritability due to inclusion of the X chromosome ranged from 0 to 0.04 points (+ 0.008 on average), indicating that the X chromosome can play an important role in the genetic determinism of traits of interest in dairy cattle. Generally speaking, values of h²X were larger in the regional breeds with the lowest numbers of cows (Tarentaise and Vosgienne), and this was particularly true for the traits with the lowest heritabilities (MAST, CCR, HCR and ICFI). However, these results should be interpreted with caution due to the relatively large degrees of error associated with these estimates (e.g., h²X = 0.011 ± 0.012 for ICFI in Vosgienne). Directly comparable results previously published are scarce for dairy cattle. In Holstein, VanRaden et al. [33] found the X chromosome accounted for about 1% of genetic variance of most of the 27 traits recorded in US and Canadian bulls while Su et al. [19] reported that using X chromosome SNPs resulted in a gain in the reliability of estimated breeding values averaged over 15 milk, fertility, udder health, and type traits (+ 0.3 to + 0.5% depending on the model used). Similarly, the heritability of female and male reproductive traits in beef cattle was estimated to increase by + 0.02 to + 0.09 as a result of the inclusion of X chromosome SNPs [34].
GWAS resolution is influenced by linkage disequilibrium. Overall, we show here higher levels of linkage disequilibrium on the X chromosome compared to an autosome of equivalent length (chromosome 2). Moreover, smaller breeds with reduced effective population size exhibited higher linkage disequilibrium. This distinction from autosomes is expected due to the greater genetic drift experienced by the X chromosome, as supporting by literature (e.g., [36]). Notably, the absence of segregation and recombination in male X chromosomes further impacts the mapping resolution of GWAS.
In our analyses, we considered paternal and maternal X chromosomes to be equally expressed at the population level and did not consider any potential effects of paternal versus maternal origin. In our association analyses, we applied the mixed model used for autosomes, which includes both the additive fixed effect of the variant tested and a vector of random polygenic effects to adjust the data for population structure. To capture the structure of the population and avoid detecting spurious associations, polygenic effects were estimated using a genomic relationship matrix calculated with the 50k SNPs of the autosomes only. We accounted for relatedness using a leave-one-chromosome-out (LOCO) approach, i.e., excluding the chromosome tested from the GRM. Although this approach may inflate test statistics [35], we chose to use it in order to avoid over-correction of the data and a consequent decrease in detection power, which was especially critical here because of the increased long-range LD on the X chromosome compared to autosomes [36]. The best option would have been to employ a leave-one-segment-out, or LOSO [37], approach that excluded the flanking region of the variant tested, but this was not computationally feasible due to the large number of animals and traits analyzed.
Regardless of the region of the genome studied, defining QTL regions (number and confidence intervals) from GWAS results is challenging and relies on a variety of more-or-less arbitrary approaches (e.g., LOD drop-off method [38]). In this study, we implemented a procedure that evaluated LD between the lead variant and variants in the surrounding region to (1) select variants to keep within the confidence interval and (2) search for other QTL on the chromosome after excluding the variants whose effects could be explained by that of the lead variant. This procedure, which can be compared to the COJO approach in GCTA software [39], has the advantage of being easily automated. Compared to autosomes, LD has a particularly strong influence in the non-PAR region of the X chromosome because no recombination occurs in males [36, 40]. Indeed, it is worth noting that the high number of QTL defined for some trait x breed combinations (up to 26 for STAT in meta-analyses), may be inflated due to the extreme LD in this region, which may have extended farther than the length of the window (20 Mbp) we used to define QTL. Increasing the size of the interval would have reduced the number of QTL but, because of the sequence-level resolution of our analyses, would also have caused computational issues. The number of QTL detected in this study is therefore probably an overestimate, but the procedure we developed would likely produce more accurate results in autosomes where the LD is less extensive.
We successfully identified QTL for most breeds and traits, with the exception of the two breeds with the lowest number of cows (Tarentaise and Vosgienne) and two fertility traits (HCR and CCR). As expected, due to the very different numbers of cows analyzed in each breed, the number of QTL was higher in the largest breeds, and these QTL presented more-significant effects. A higher number of QTL with more-significant effects was also found for the most heritable traits (milk production, milk composition, and stature).
Between breeds, we identified QTL with overlapping confidence intervals but the variants with the most significant effects were never the same, and were only rarely located in the same genes. This result has also been observed in autosomes (e.g., [41]), but appears to be more pronounced in the non-PAR region of the X chromosome, probably because of the heightened influence of within-breed LD. By combining different populations with different LD patterns, meta-analyses generally increase both the power and resolution of association analyses (e.g., [42]). Indeed, here this approach detected more breed x trait combinations with significant effects (n = 157), and, on average, more precise locations (65 variants in 1.7 Mbp), than any of the within-breed association analyses, for which the highest number of QTL detected was 74 (in Montbéliarde) and even the smallest confidence interval still contained 96 variants (in 2.5 Mbp in Normande).
As previously mentioned, the X chromosome has often been excluded from association analyses due to its unique pattern of inheritance, but it also suffers from a lower quality assembly and poorer functional annotation compared to autosomes, in particular in the previous UMD3.1 assembly [14, 17]. GWAS that examine the X chromosome are therefore rare, in particular at the sequence level. In the present study, by combining within-breed and meta-analyses of association at the sequence level on a large number of animals from six breeds and for a wide panel of traits, we are able to propose a list of candidate genes that could be responsible for the largest effects observed on the X chromosome.
Based on literature and human databases, the function of the positional candidate genes – or their association with traits similar to those investigated in this study – was examined in order to highlight the best functional candidates. Although we were not able to identify a clear functional link to milk production and composition traits, four of the best positional candidate genes associated with the most significant QTL for these traits warrant special attention: GPC3 (glypican 3), MBNL3 (muscleblind like splicing regulator3), HS6ST2 (heparan sulfate 6-o sulfotransferase 2), and DMD (dystrophin). GPC3, which was one of the best candidates for PC in our study, has been previously associated to longevity [43] and metabolic disorders [44] in Holstein cows; both traits could be related to milk composition. A more recent study, conducted at the whole-genome sequence level, identified a QTL for milk urea nitrogen (which is genetically correlated with milk PC [45]), in which the lead SNP was located between MBNL3 and HS6ST2 at 16,376,624 bp [46], i.e., close to the lead variants we found for several QTL for PC in Normande (at 16,169,349 bp), and FC in Montbéliarde (at 16,420,927 bp) and Normande (at 16,679,506 bp). Furthermore, HS6ST2 knockout mice showed glucose and insulin metabolism disorders [47]. Mutations in the DMD gene, leading to the absence or dysfunction of the dystrophin protein causing muscular dystrophies in humans, were associated with muscle fat replacement [48]. In a Chinese Holstein population, genome-wide association analyses of milk, protein, and fat yields, conducted with SNPs of the Illumina BovineSNP150 BeadChip, identified rs135780687, located at 127,465,011 bp on the X chromosome, as the lead SNP for a QTL detected for FY [49]. The authors identified GRPR, encoding a gastrin-releasing peptide receptor, as a functional candidate gene in this region. In the present, sequence-level, study, we detected QTL in this region for multiple traits but the most plausible positional candidate genes were generally different from GRPR, with the exception of two QTL identified in meta-analyses for MY and PY, for which the lead variants were located in the intergenic region between the GRPR and AP1S2 genes (at 127,509,209 and 127,507,371 bp, respectively).
Notably, among the positional candidate genes identified for udder health, three encode transmembrane proteins (TMEM164, TMEM47, and TMEM187) and two encode protein phosphatase regulators (PPP1R2C and PPP4R3C). The lead variant of the QTL with the most significant effect on SCS in Holstein was located close to TMEM164, which was found to be differentially expressed in mammary infections due to E. coli and S. aureus in cattle [50] and associated with improved survival and increased immune cell infiltration in patients with pancreatic cancer [51]. ACSL4 (acyl-CoA synthetase long chain family member4), which is involved in fatty acid metabolism, is another positional candidate gene for the most significant QTL identified in Holstein. This gene was reported to be upregulated in late lactation, resulting in an increased concentration of triglycerides in bovine mammary epithelial cells [52]; interestingly, Genini et al. [50] demonstrated that lipid metabolism was significantly affected during the cattle-specific response to mastitis infection, suggesting that it could be tightly linked to immune response. Furthermore, ACSL4 was found to be involved in inflammatory responses in mice [53]. Two other genes, candidates for the most significant QTL identified in Normande (ENOX2, ecto-NOX disulfide-thiol exchanger 2) and in meta-analyses (HTR2C, 5-hydroxytryptamine receptor 2 C), have been associated with ruminant health: the former was associated with innate immunity in sheep [54] while the latter was identified as a candidate gene for hyperketonemia in Holstein cows [55]. Finally, two other genes are noteworthy: AMOT (angiomotin), associated with the most significant effects on SCS in Montbéliarde, promotes the proliferation of mammary epithelial cells in women [56] and IRAK1 (interleukin 1 receptor associated kinase 1), one of the best candidates in both Holstein and meta-analyses, was described as a critical signaling mediator of innate immunity [57].
Among the genes identified for ICFI, which was the only fertility trait with significant results in our study, MAMLD1 and COL4A6 appear to be the best functional candidates. MAMLD1 (mastermind-like domain containing 1) has been associated with disorders of sex development in men, as well with ovarian dysfunction in women [58, 59]. This gene has also been highlighted as a candidate for bull fertility [27] and for the number of piglets born alive [60]. COL4A6 (collagen type IV alpha 6 chain) was found to be particularly expressed in the bovine uterus [61] and differentially expressed in the endometrium of high- and low-fertility heifers during the mid-luteal phase of the estrus cycle [62].
Because of its relatively high heritability and ease of measurement, stature (height in humans) has been examined in very large-scale GWAS and meta-analyses in different species, including cattle [10] and humans [3]. However, neither one of these studies considered the X chromosome, which could explain a part of the missing heritability observed in each case. Indeed, in the present study, stature was the trait for which the highest number of QTL was detected (8 in Montbéliarde, 13 in Normande, 14 in Holstein, and 26 in meta-analyses) and we identified a number of functional candidate genes that might explain the effects we observed. For the QTL located around 53 Mbp, the confidence intervals are probably inflated due by the long order inversion between 51.9 and 54.5 Mbp on the ARS-UCD1.2 bovine assembly previously identified by Zhang et al. [40]. In particular, although they were predicted to have only moderate effects, two of the lead variants were associated with missense changes in the proteins encoded by NRK (nik-related protein kinase), at 54,703,522 bp, and PLXNB3 (plexin B3), at 36,994,420 bp. Interestingly, NRK has been previously linked with an X-linked form of short stature in humans [63], which supports its role in the genetic determinism of this trait. In contrast, no functional link with stature has yet been identified for PLXNB3. Furthermore, a larger number of positional candidate genes for stature were shared between breeds than any of the other traits we examined. Among these genes, the best positional candidates were TRPC5 (transient receptor potential cation channel subfamily C member 5), which was shared between Holstein and Normande, and COL4A6 (collagen type IV alpha 6 chain), which was detected in both Montbéliarde and Normande. The latter gene was also associated with fertility in our study, but presented no obvious functional link with stature. Among the within-breed analyses and the meta-analysis, several other genes were highlighted for the QTL with the most significant effects, namely, GPR50 (G protein-coupled receptor 50), VMA21 (vacuolar ATPase assembly factor VMA21), IDS (iduronate 2-sulfatase), GPC4 (glypican 4), ESX1 (ESX homeobox 1), GPC3 (glypican 3), bta-mir-507b, PCDH19 (protocadherin 19), and SLITRK4 (SLIT and NTRK like family member 4). Four of these are involved in growth disorders in humans: ESX1 and GPR50 were found to be involved in growth and pituitary hormone deficiencies [64, 65], while GPC3 and GPC4 were identified as genes causing Simpson-Golabi-Behmel syndrome, a rare X-linked syndrome characterized by pre‐and post‐natal overgrowth [66].
Conclusions
With its large sample size and fine-scale (sequence-level) resolution, this study provides clear evidence for the importance of the X chromosome in the genetic determinism of complex traits in dairy cattle. These new insights support the inclusion of this chromosome in all genetic evaluation models in which it is not currently considered. Based on our results, we would expect that inclusion of the X chromosome would increase the accuracy of estimated breeding values and expedite genetic progress on milk production, milk composition, udder health, and fertility traits, which are all included in the breeding goals of the different dairy cattle breeds. Although the necessary changes to the evaluation software and procedure would not be trivial, the relative gain of 3–5% in the accuracy of genomic estimated breeding values would justify the effort. In addition, because the majority of X-linked genes are shared between mammals, further work on the identification of X-linked genes that are involved in the genetic determinism of traits could be beneficial for our understanding of other species.
Methods
Ethics statement
All analyses were performed using data from routine milk recording and genotyping in commercial herds of French cows. We did not perform any experiments on animals and no ethical approval was required.
Cows, phenotypes, and genotypes
We analyzed 236,496 cows from three national breeds—Holstein (81,815 animals), Montbéliarde (61,881 animals), and Normande (78,472 animals)—and three regional breeds—Abondance (7449 animals), Tarentaise (3969 animals), and Vosgienne (2910 animals)—for which phenotypes and 50k genotypes were available (Table 1).
Phenotypes were obtained for 11 traits:
-
Five milk production traits defined at the level of the individual lactation based on monthly records: milk yield (MY), protein yield (PY), fat yield (FY), protein content (PC), and fat content (FC);
-
Two udder health traits: average somatic cell score (SCS) at lactation level, computed as the mean of monthly records of log-transformed somatic cell counts, and clinical mastitis (MAST; except for Vosgienne) defined as 0/1 (0 = no clinical mastitis and 1 = at least one episode of clinical mastitis in the interval from 10 days before calving to 150 days after calving);
-
Three female reproductive traits: the interval between calving and the first artificial insemination (ICFI) which reflects the ability of a cow to initiate the postpartum cycle, and heifers’ (HCR) and lactating cows’ (CCR) conception rates, which represent the success/failure (1/0) of each artificial insemination;
-
Stature (STAT);
-
To remove the influence of environmental effects, all phenotypes used were yield deviations (YD), i.e., phenotypes adjusted for non-genetic effects and, for repeated records, averaged per cow. YDs are produced by the French national genetic evaluation systems for the Holstein, Montbéliarde, Normande, Abondance, Tarentaise, and Vosgienne populations using the models described at https://interbull.org/ib/geforms [67].
Cows in the six breeds were genotyped with different versions of the 50 K SNP Beadchip, with the most recent being the EuroGMD Beadchip, which is currently used for genomic selection (https://www.eurogenomics.com/actualites/the-eurog-md:-a-unique-genotyping-microarray-for-cattle-.html). The standard EuroGMD Beadchip contains 53,469 autosomal SNPs and 1147 SNPs located in the non-pseudoautosomal region (non-PAR) of the X chromosome; all SNPs passed all quality control filters (individual call rate > 95%; SNP call rate > 90%; minor allele frequency (MAF) > 1%; genotype frequencies in HW equilibrium with P > 10− 4).
Imputation analyses
Missing genotypes of EuroGMD SNPs are routinely imputed in the French evaluation system using FImpute software [68]. For imputations at higher densities (HD and sequence levels), we considered only the X-specific non-PAR region, which covers the majority of the X chromosome (0–133.3 Mbp on the ARS-UCD1.2 reference genome [14]). To account for male hemizygosity for this chromosome, we assumed that all males were homozygous for all non-PAR SNPs, and removed the pedigree information. All imputation analyses were done within-breed. First, HD genotypes of 32,268 SNPs were imputed with FImpute [68] from genotypes of the 1147 non-PAR SNPs of the EuroGMD chip, using the 179 to 804 major ancestors of each breed with HD genotypes as a reference (Table 3). Then, 778,576 sequence variants were imputed using a multi-breed population of 2712 animals from the RUN8 reference panel of the 1000 Bull Genomes consortium [10] and the Minimac algorithm [24]. The reference population for the sequence-level imputation comprised 2712 Bos taurus animals from 28 different breeds, including 1059 Holstein, 63 Montbéliarde, 45 Normande, 9 Abondance, 12 Tarentaise, and 4 Vosgienne (S1 Table).
Linkage disequilibrium
Linkage disequilibrium (r2) were assessed for the X chromosome and for an autosome of equivalent length (chromosome 2) in each breed using a sample of cows. To minimize relatedness and avoid accumulation of the same paternal X chromosome, one daughter per sire was randomly selected. A total of 126 to 2279 individuals were included in the study, depending on the breed. High density SNP genotypes, either true or imputed, were used for a total of 15,892 and 35,723 SNPs on chromosomes X and 2, respectively. Variants with a minor allele frequency (MAF) less than 0.01 were excluded from the analysis. The calculated values were then averaged within bins based on marker distance.
Genomic relationship matrices
Three different genomic relationship matrices were constructed at the 50k density: the first contained 53,469 autosomal SNPs (GA), the second contained 1147 SNPs of the non-PAR X chromosome (GX), and the third contained both autosomal and X-chromosome SNPs, i.e., 54,616 SNPs (GG). As all animals included in this study were females, no assumptions were made regarding dosage compensation for the X chromosome (both X chromosomes active in females). All matrices were therefore constructed using the --make-grm option of GCTA software [69] which was developed for autosomal SNPs.
REML analyses
To estimate the relative proportions of genetic variance explained by the autosomes and the X chromosome, and the corresponding heritabilities, within-breed REML analyses were carried out for each trait using GCTA software [69] and the following model:
where y is the vector of YD; µ is the overall mean; gA~ N(0,GAσ²A) is the vector of random autosomal genetic effects, with GA the autosomal GRM and σ²A the autosomal genetic variance; gX~ N(0,GXσ²X) is the vector of random X-linked genetic effects, with GX the X-chromosome GRM and σ²X the X-linked genetic variance; and e ~ N(0,Iσ²e) is the vector of random residual effects, with I the identity matrix and σ²e the residual variance.
Then, we calculated the overall heritability of the traits (h²=(σ²A+σ²X)/(σ²A+σ²X+σ²e)) and the heritability due to autosomes (h²AUT=σ²A/(σ²A+σ²X+σ²e)) and the X chromosome (h²X=σ²X/(σ²A+σ²X+σ²e)).
Within-breed association analyses
Allele dosages of X-linked variants imputed at the sequence level were evaluated in within-breed association analyses using GCTA software [69]. To adjust data for population structure, we estimated polygenic effects using the GA GRM calculated with autosomal 50k SNPs. All phenotypes were measured on females, which had two copies of the X chromosome. Therefore, we applied the following linear mixed model:
where y is the vector of YD; µ is the overall mean; b is the additive fixed effect of the variant tested; x is the vector of imputed allele dosages; gA~ N(0, GA σ²A) is the vector of random polygenic effects, with GA the GRM based on autosomal SNPs and σ²A the autosomal polygenic variance; and e ~ N(0,Iσ²e) is the vector of random residual effects, with I the identity matrix and σ²e the residual variance.
We analyzed variants with a MAF ≥ 0.005 and with a Minimac imputation R² ≥ 0.20, which resulted in between 154,966 and 201,554 variants depending on the breed (Table 3).
Association meta-analyses
We then conducted meta-analyses for each trait by combining the within-breed association results of the six breeds (five for MAST, which was not measured in Vosgienne). All variants retained after filtering (MAF ≥ 0.005 and Minimac R² ≥ 0.20), i.e., 212,111 for MAST and 224,073 for all other traits, were included in meta-analyses. The fixed effects meta-analysis method was applied as implemented in METAL software [70]. This method assumes that the true effect of each allele is the same across different studies and combines the different effects by weighting them by the inverse of their error variance. Therefore, this meta-analysis method weights the different studies by their sample size.
Identification of QTL regions
We implemented an iterative procedure to identify QTL regions in the within-breed association analyses and meta-analyses based on a threshold corresponding to P = 0.05 after Bonferroni correction for ~ 1 million independent tests (-log10(P) = 7.3) [71].
For each trait, we applied the following six-step iterative procedure:
-
1.
Search for the variant with the maximal -log10(P) (≥ 7.3), hereafter named the lead variant;
-
2.
Select all variants within a 20-Mb window centered around the lead variant;
-
3.
Calculate LD between the lead variant and all other variants in the 20-Mb interval, i.e., correlations between allele dosages (r);
-
4.
Define the confidence interval of the QTL by retaining all variants in high LD with the lead variant (|r| ≥ 0.7);
-
5.
For each variant i outside of the confidence interval and located in the 20-Mb window, determine if its effect could be explained by the effect of the lead variant; for this, we calculated a new test statistic, TNEW = (bi – bLEAD x r (i,LEAD)) / SEi, and the corresponding log10(PNEW) value, and selected variants that satisfied the following two conditions: (1) log10(PNEW) < 7.3 and (2) Ti (bi/SEi) and TNEW had the same sign;
-
6.
Remove all variants identified in steps 4 (variants within the confidence interval) and 5 (variants with effects explained by LD with the lead variant).
This procedure, described in Fig. 1, was repeated until no more significant variants were found on the chromosome.
Functional annotation and visualization.
Variants in the confidence intervals of each QTL were annotated with the Ensembl variant effect predictor (VEP) pipeline v81 [72] and effects of amino-acid changes were predicted using the SIFT tool [73]. Functions of genes were investigated using GeneCards [64] and MalaCards [74]. Visualization of the QTL and of their annotation was performed using the R LocusZoom function [75]. Manhattan plot and UpSet diagrams were created using the R packages qqman [76] and UpSetR [77], respectively.
Data Availability
All GWAS and meta-analyses results are available at the following URL https://entrepot.recherche.data.gouv.fr/privateurl.xhtml?token=fb349842-2a69-49d5-bdca-a3f0ec3d1dbd. Raw genotypes and phenotypes data are part of a reference population used for genomic selection and have commercial value. Therefore, restrictions apply to their availability and they are not publicly available. The authors can be contacted for a reasonable request.
Abbreviations
- CCR:
-
Lactating cows’ conception rate
- FC:
-
Fat content
- FY:
-
Fat yield
- GRM:
-
Genomic relationship matrix
- GWAS:
-
Genome-wide association studies
- HCR:
-
Heifers’ conception rate
- HD:
-
High density
- ICFI:
-
Interval between calving and first fertilizing artificial insemination
- LD:
-
Linkage disequilibrium
- LOCO:
-
Leave-one-chromosome-out
- LOSO:
-
Leave-one-segment-out
- MAF:
-
Minor allele frequency
- MAST:
-
Clinical mastitis
- MY:
-
Milk yield
- non-PAR:
-
Non-pseudoautosomal region
- PC:
-
Protein content
- PY:
-
Protein yield
- QTL:
-
Quantitative trait locus
- REML:
-
Restricted maximum likelihood
- SCS:
-
Somatic cell score
- SNP:
-
Single nucleotide polymorphism
- STAT:
-
Stature
References
Visscher P, Wray N, Zhang Q, Sklar P, McCarthy M, Brown M, et al. 10 years of GWAS Discovery: Biology, function, and translation. Am J Hum Genet. 2017;101:5–22.
Yengo L, Sidorenko J, Kemper K, Zheng Z, Wood A, Weedon M, et al. Meta-analysis of genome-wide association studies for height and body mass index in similar to 700 000 individuals of european ancestry. Hum Mol Genet. 2018;27(20):3641–9.
Yengo L, Vedantam S, Marouli E, Sidorenko J, Bartell E, Sakaue S, et al. A saturated map of common genetic variants associated with human height. Nature. 2022;610(7933):704–12.
Manolio T, Collins F, Cox N, Goldstein D, Hindorff L, Hunter D, et al. Finding the missing heritability of complex diseases. Nature. 2009;461(7265):747–53.
Nolte I, van der Most P, Alizadeh B, de Bakker P, Boezen H, Bruinenberg M, et al. Missing heritability: is the gap closing? An analysis of 32 complex traits in the Lifelines Cohort Study. Eur J Hum Genet. 2017;25(7):877–85.
Wray N, Goddard M, Visscher P. Prediction of individual genetic risk to disease from genome-wide association studies. Genome Res. 2007;17(10):1520–8.
Zimin A, Delcher A, Florea L, Kelley D, Schatz M, Puiu D, et al. A whole-genome assembly of the domestic cow, Bos taurus. Genome Biol. 2009;10(4):R42.
Hayes B, Daetwyler H, Lewin H, Roberts R. 1000 Bull Genomes Project to Map simple and complex genetic traits in cattle: applications and outcomes. Annu Rev Anim Biosci. 2019;7:89–102.
Meuwissen T, Hayes B, MacLeod I, Goddard M. Identification of genomic variants causing variation in quantitative traits: a review. Agriculture. 2022;12(10):1713.
Bouwman AC, Daetwyler HD, Chamberlain AJ, Ponce CH, Sargolzaei M, Schenkel FS, et al. Meta-analysis of genome-wide association studies for cattle stature identifies common genes that regulate body size in mammals. Nat Genet. 2018;50:362–7.
Daetwyler HD, Capitan A, Pausch H, Stothard P, Van Binsbergen R, Brøndum RF, et al. Whole-genome sequencing of 234 bulls facilitates mapping of monogenic and complex traits in cattle. Nat Genet. 2014;46:858–67.
Hayes B, Bowman P, Chamberlain A, Goddard M. Invited review: genomic selection in dairy cattle: Progress and challenges. J Dairy Sci. 2009;92:433–43.
Yengo L, Vedantam S, Marouli E, Sidorenko J, Bartell E, Sakaue S, et al. A saturated map of common genetic variants associated with human height. Nature. 2022;610(7933):704–.
Johnson T, Keehan M, Harland C, Lopdell T, Spelman R, Davis S, et al. Short communication: identification of the pseudoautosomal region in the Hereford bovine reference genome assembly ARS-UCD1.2. J Dairy Sci. 2019;102(4):3254–8.
Rappold G. The pseudoautosomal regions of the human sex-chromosomes. Hum Genet. 1993;92(4):315–24.
Nguyen D, Disteche C. Dosage compensation of the active X chromosome in mammals. Nat Genet. 2006;38(1):47–53.
Couldrey C, Johnson T, Lopdell T, Zhang I, Littlejohn M, Keehan M, et al. Bovine mammary gland X chromosome inactivation. J Dairy Sci. 2017;100(7):5491–500.
Druet T, Legarra A. Theoretical and empirical comparisons of expected and realized relationships for the X-chromosome. Genet Sel Evol. 2020;52(1).
Su G, Guldbrandtsen B, Aamand G, Stranden I, Lund M. Genomic relationships based on X chromosome markers and accuracy of genomic predictions with and without X chromosome markers. Genet Sel Evol. 2014;46.
Cunningham F, Allen J, Allen J, Alvarez-Jarreta J, Amode M, Armean I, et al. Ensembl 2022. Nucleic Acids Res. 2022;50(D1):D988–D95.
Boichard D, Guillaume F, Baur A, Croiseau P, Rossignol M, Boscher M, et al. Genomic selection in french dairy cattle. Anim Prod Sci. 2012;52:115–20.
Sanchez MP, Jonas D, Baur A, Ducrocq V, Hozé C, Saintilan R, et al. editors. Implementation of genomic selection in three french regional dairy cattle breeds. 67 ed. Belfast, Ireland: European Association of Animal Production; 2016.
Boichard D, Boussaha M, Capitan A, Rocha D, Hozé C, Sanchez MP, et al. Experience from large scale use of the EuroGenomics custom SNP chip in cattle. 11 ed. New Zealand: th WCGALP; Auckland; 2018.
Howie B, Fuchsberger C, Stephens M, Marchini J, Abecasis GR. Fast and accurate genotype imputation in genome-wide association studies through pre-phasing. Nat Genet. 2012;44.
Escouflaire C, Rebours E, Charles M, Orellana S, Cano M, Rivière J, et al. Α de novo 3.8-Mb inversion affecting the EDA and XIST genes in a heterozygous female calf with generalized hypohidrotic ectodermal dysplasia. BMC Genom. 2019;18(20):715.
Diaz I, de Camargo G, da Cruz V, Hermisdorff I, Carvalho C, de Albuquerque L, et al. Mapping genomic regions for reproductive traits in beef cattle: inclusion of the X chromosome. Reprod Domest Anim. 2020;55(11):1650–4.
Fortes M, Porto-Neto L, Satake N, Nguyen L, Freitas A, Melo T et al. X chromosome variants are associated with male fertility traits in two bovine populations. Genet Sel Evol. 2020;52(1).
Carvalho C, da Costa Hermisdorff I, Souza I, Junqueira G, Magalhães A, Fonseca L, et al. Influence of X-chromosome markers on reproductive traits of beef cattle. Livest Sci. 2019;220:152–7.
Fortes M, Reverter A, Hawken R, Bolormaa S, Lehnert S. Candidate genes Associated with Testicular Development, sperm quality, and hormone levels of Inhibin, luteinizing hormone, and insulin-like Growth factor 1 in Brahman Bulls. Biol Reprod. 2012;87(3).
Fortes M, Reverter A, Kelly M, McCulloch R, Lehnert S. Genome-wide association study for inhibin, luteinizing hormone, insulin-like growth factor 1, testicular size and semen traits in bovine species. Andrology. 2013;1(4):644–50.
Hermisdorff I, Diaz I, de Camargo G, de Albuquerque L, Costa R. Effect of genomic X-chromosome regions on Nelore bull fertility. J Appl Genet. 2021;62(4):655–9.
Tribout T, Croiseau P, Lefebvre R, Barbat A, Boussaha M, Fritz S, et al. Confirmed effects of candidate variants for milk production, udder health, and udder morphology in dairy cattle. Genet Sel Evol. 2020;52(1):55.
VanRaden PM, Van Tassell CP, Wiggans GR, Sonstegard TS, Schnabel RD, Taylor JF, et al. Invited review: reliability of genomic predictions for north american Holstein bulls. J Dairy Sci. 2009;92(1):16–24.
Diaz I, de Camargo G, da Cruz V, Hermisdorff I, Carvalho C, de Albuquerque L et al. Effect of the X chromosome in genomic evaluations of reproductive traits in beef cattle. Anim Reprod Sci. 2021;225.
Mesbah-Uddin M, Guldbrandtsen B, Capitan A, Lund M, Boichard D, Sahana G. Genome-wide association study with imputed whole-genome sequence variants including large deletions for female fertility in 3 nordic dairy cattle breeds. J Dairy Sci. 2022;105(2):1298–313.
Sandor C, Farnir F, Hansoul S, Coppieters W, Meuwissen T, Georges M. Linkage disequilibrium on the bovine X chromosome: characterization and use in quantitative trait Locus Mapping. Genetics. 2006;173(3):1777–86.
Reynolds E, Neeley C, Lopdell T, Keehan M, Dittmer K, Harland C, et al. Non-additive association analysis using proxy phenotypes identifies novel cattle syndromes. Nat Genet. 2021;53(7):949–54.
Lander E, Botstein D. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics. 1989;121:185–99.
Yang J, Ferreira T, Morris A, Medland S, Madden P, Heath A, et al. Conditional and joint multiple-SNP analysis of GWAS summary statistics identifies additional variants influencing complex traits. Nat Genet. 2012;44:369–75.
Zhang J, Kadri N, Mullaart E, Spelman R, Fritz S, Boichard D, et al. Genetic architecture of individual variation in recombination rate on the X chromosome in cattle. Heredity. 2020;125(5):304–16.
Sanchez MP, Govignon-Gion A, Croiseau P, Fritz S, Hozé C, Miranda G, et al. Within-breed and multi-breed GWAS on imputed whole-genome sequence variants reveal candidate mutations affecting milk protein composition in dairy cattle. Genet Sel Evol. 2017;49:68.
van den Berg I, Xiang R, Jenko J, Pausch H, Boussaha M, Schrooten C et al. Meta-analysis for milk fat and protein percentage using imputed sequence variant genotypes in 94,321 cattle from eight cattle breeds. Genet Sel Evol. 2020;52(1).
Steri R, Moioli B, Catillo G, Galli A, Buttazzoni L. Genome-wide association study for longevity in the Holstein cattle population. Animal. 2019;13(7):1350–7.
Atashi H, Salavati M, Koster J, Crowe M, Opsomer G, Hostens M, et al. Genome-wide association for metabolic clusters in early -lactation holstein dairy cows. J Dairy Sci. 2020;103(7):6392–406.
Beatson P, Meier S, Cullen N, Eding H. Genetic variation in milk urea nitrogen concentration of dairy cattle and its implications for reducing urinary nitrogen excretion. Animal. 2019;13(10):2164–71.
van den Berg I, Ho P, Nguyen T, Haile-Mariam M, MacLeod I, Beatson P et al. GWAS and genomic prediction of milk urea nitrogen in australian and New Zealand dairy cattle. Genet Sel Evol. 2022;54(1).
Nagai N, Habuchi H, Sugaya N, Nakamura M, Imamura T, Watanabe H, et al. Involvement of heparan sulfate 6-O-sulfation in the regulation of energy metabolism and the alteration of thyroid hormone levels in male mice. Glycobiology. 2013;23(8):980–92.
Veeger T, van Zwet E, Al Mohamad D, Naarding K, van de Velde N, Hooijmans M, et al. Muscle architecture is associated with muscle fat replacement in Duchenne and Becker muscular dystrophies. Muscle Nerve. 2021;64:576–84.
Liu L, Zhou J, Chen C, Zhang J, Wen W, Tian J et al. GWAS-based identification of new loci for milk yield, fat, and protein in Holstein cattle. Animals. 2020;10(11).
Genini S, Badaoui B, Sclep G, Bishop S, Waddington D, Pinard-van der Laan M et al. Strengthening insights into host responses to mastitis infection in ruminants by combining heterogeneous microarray data sources. BMC Genom. 2011;12.
Liu J, Liu Y, Wang Y, Li C, Xie Y, Klionsky D et al. TMEM164 is a new determinant of autophagy-dependent ferroptosis. Autophagy. 2022:1–12.
Fan Y, Han Z, Lu X, Zhang H, Arbab A, Loor J et al. Identification of milk fat metabolism-related pathways of the bovine mammary gland during mid and late lactation and functional verification of the ACSL4 gene. Genes. 2020;11(11).
Kuwata H, Nakatani E, Shimbara-Matsubayashi S, Ishikawa F, Shibanuma M, Sasaki Y, et al. Long-chain acyl-CoA synthetase 4 participates in the formation of highly unsaturated fatty acid-containing phospholipids in murine macrophages. Biochim Biophys Acta Mol Cell Biol Lipids. 2019;1864(11):1606–18.
Yang H, Yang Y, Li G, Yu Q, Yang J. Identifications of immune-responsive genes for adaptative traits by comparative transcriptome analysis of spleen tissue from Kazakh and Suffolk sheep. Sci Rep. 2021;11(1).
Yepes F, Nydam D, Mann S, Caixeta L, McArt J, Overton T et al. Longitudinal phenotypes improve genotype association for hyperketonemia in dairy cattle. Animals. 2019;9(12).
Ranahan W, Han Z, Smith-Kinnaman W, Nabinger S, Heller B, Britney-Shea H, et al. The adaptor protein AMOT promotes the proliferation of mammary epithelial cells via the prolonged activation of the extracellular signal-regulated kinases. Cancer Res. 2011;71:2203–11.
Gottipati S, NL R, Fung-Leung W. IRAK1: a critical signaling mediator of innate immunity. Cell Signal. 2007;20:269–76.
Miyado M, Fukami M, Ogata T. MAMLD1 and Differences/Disorders of Sex Development: an update. Sex Dev. 2022;16(2–3):126–37.
Fukami M, Wada Y, Miyabayashi K, Nishino I, Hasegawa T, Nordenskjold A, et al. CXorf6 is a causative gene for hypospadias. Nat Genet. 2006;38(12):1369–71.
Chen S, Freitas P, Oliveira H, Lazaro S, Huang Y, Howard J et al. Genotype-by-environment interactions for reproduction, body composition, and growth traits in maternal-line pigs based on single-step genomic reaction norms. Genet Sel Evol. 2021;53(1).
Fang L, Cai W, Liu S, Canela-Xandri O, Gao Y, Jiang J, et al. Comprehensive analyses of 723 transcriptomes enhance genetic and biological interpretations for complex traits in cattle. Genome Res. 2020;30(5):790–801.
Killeen A, Morris D, Kenny D, Mullen M, Diskin M, Waters S. Global gene expression in endometrium of high and low fertility heifers during the mid-luteal phase of the estrous cycle. BMC Genom. 2014;15.
Wit J, van Duyvenvoorde H, van Klinken J, Caliebe J, Bosch C, Lui J, et al. Copy number variants in short children born small for gestational age. Horm Res Paediat. 2014;82(5):310–8.
Stelzer G, Rosen R, Plaschkes I, Zimmerman S, Twik M, Fishilevich S et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analysis. Curr Protoc Bioinformatics. 2016;54:1.30.1 -1.3.
Li Y, Behringer R. Esx1 is an x-chromosome-imprinted regulator of placental development and fetal growth. Nat Genet. 1998;20:309–11.
Cottereau E, Mortemousque I, Fau - Moizard M-P, Moizard Mp Fau - Bürglen L, Bürglen L, Fau - Lacombe D, Lacombe D, Fau - Gilbert-Dussardier B, Gilbert-Dussardier B, Fau - Sigaudy S, et al. Phenotypic spectrum of Simpson-Golabi-Behmel syndrome in a series of 42 cases with a mutation in GPC3 and review of the literature. Am J Med Genet. 2013;163 C(1552–4876):92–105.
https://. interbull.org/ib/geforms. Int Bull Evaluation Service Official Website 2023 [.
Sargolzaei M, Chesnais J, Schenkel F. A new approach for efficient genotype imputation using information from relatives. BMC Genom. 2014;15:478.
Yang J, Lee S, Goddard M, Visscher P. GCTA: A Tool for Genome-wide Complex Trait Analysis. Am J Hum Genet. 2011;88:76–82.
Willer C, Li Y, Abecasis G. METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics. 2010;26(17):2190–1.
Chen Z, Boehnke M, Wen X, Mukherjee B. Revisiting the genome-wide significance threshold for common variant GWAS. G3. 2021;11(2):jkaa056.
McLaren W, Gil L, Hunt S, Riat H, Ritchie G, Thormann A, et al. The Ensembl variant effect predictor. Genome Biol. 2016;17:122.
Kumar P, Henikoff S, Ng P. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat Protoc. 2009;4:1073–82.
Rappaport N, Twik M, Plaschkes I, Nudel R, Iny Stein T, Levitt J, et al. MalaCards: an amalgamated human disease compendium with diverse clinical and genetic annotation and structured search. Nuc Acids Res. 2016;45:D877–D87.
Pruim R, Welch R, Sanna S, Teslovich T, Chines P, Gliedt T, et al. LocusZoom: regional visualization of genome-wide association scan results. Bioinformatics. 2010;26(18):2336–7.
Turner S. qqman: an R package for visualizing GWAS results using Q-Q and manhattan plots. J Open Source Software. 2018;3(25):731.
Conway J, Lex A, Gehlenborg N. UpSetR: an R package for the visualization of intersecting sets and their properties. Bioinformatics. 2017;33(18):2938–40.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
MPS, CE, CH, SF, AC, and DB conceptualized the study. MPS, CH, DB, AB, MB, and SF performed pre-analyses or provided resources for analyses. MPS, AC, and DB supervised the analyses. MPS, CE, and FB performed formal analyses. MPS wrote the original draft. All the authors reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate.
Not applicable.
Consent for publication.
Not applicable.
Additional information
Acknowledgements.
The authors gratefully acknowledge the French National Database (CTIG) and Valogene for providing phenotype and genotype data, respectively, as well as the contribution of the 1000 Bull Genomes consortium. We also thank Dr. Tom Druet (University of Liège, Belgium) for his helpful advice regarding the analyses.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

{kind=link}
Cite this article
Sanchez, MP., Escouflaire, C., Baur, A. et al. X-linked genes influence various complex traits in dairy cattle. BMC Genomics 24, 338 (2023). https://doi.org/10.1186/s12864-023-09438-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-023-09438-7