
Abstract
Background
Chronic migraine is a common and highly disabling disorder. Functional MRI has indicated that abnormal brain region activation is linked with chronic migraine. Drugs targeting the calcitonin gene-related peptide (CGRP) or its receptor have been reported to be efficient for treating chronic migraine. The CGRP signaling was also shared in two types of chronic migraine models (CMMs). However, it remains unclear whether the activation of specific brain regions could contribute to persistent behavioral sensitization, and CGRP receptor antagonists relieve migraine-like pain in CMMs by altering specific brain region activation. Therefore, it’s of great interest to investigate brain activation pattern and the effect of olcegepant (a CGRP receptor-specific antagonist) treatment on alleviating hyperalgesia by altering brain activation in two CMMs, and provide a reference for future research on neural circuits.
Methods
Repeated administration of nitroglycerin (NTG) or levcromakalim (LEV) was conducted to stimulate human migraine-like pain and establish two types of CMMs in mice. Mechanical hypersensitivity was evaluated by using the von Frey filament test. Then, we evaluated the activation of different brain regions with c-Fos and NeuN staining. Olcegepant was administered to explore its effect on mechanical hyperalgesia and brain region activation.
Results
In two CMMs, acute and basal mechanical hyperalgesia was observed, and olcegepant alleviated mechanical hyperalgesia. In the NTG-induced CMM, the medial prefrontal cortex (mPFC), anterior cingulate cortex (ACC), and the caudal part of the spinal trigeminal nucleus (Sp5c) showed a significant increase of c-Fos expression in the NTG group (p < 0.05), while pre-treatment with olcegepant reduced c-Fos expression compared with NTG group (p < 0.05). No significant difference of c-Fos expression was found in the paraventricular thalamic nucleus (PVT) and ventrolateral periaqueductal gray (vlPAG) between the vehicle control and NTG group (p > 0.05). In the LEV-induced CMM, mPFC, PVT, and Sp5c showed a significant increase of c-Fos expression between vehicle control and LEV group, and olcegepant reduced c-Fos expression (p < 0.05). No significant difference in c-Fos expression was found in vlPAG and ACC (p > 0.05).
Conclusions
Our study demonstrated the activation of mPFC and Sp5c in two CMMs. Olcegepant may alleviate hyperalgesia of the hind paw and periorbital area by attenuating brain activation in CMMs.
Similar content being viewed by others

Background
Migraine is one of the most common types of primary headache disorders and represents a brain state of altered excitability [1,2,3]. Current evidence suggests that about 3% of patients with episodic migraine annually progress to chronic migraine (CM) [4]. Recent neuroimaging studies showed structural and functional changes in the cortex [5,6,7,8,9,10,11,12], basal ganglia [7,8,9,10], thalamus [6, 12], hypothalamus [13, 14], and brainstem [14, 15] in patients with chronic migraine. Thus, it’s of great interest to explore how neurons are activated in these brain regions. However, in chronic migraine models (CMMs), including the repeated dural application of inflammatory soup, and chronic systemic infusion of nitroglycerin (NTG) [16, 17], studies investigating brain activation remain scarce [18].
In experimental models of migraine in humans, migraine could be triggered by various compounds such as nitric oxide (NO) donor NTG, calcitonin gene-related peptide (CGRP), phosphodiesterase 3 (PDE3) inhibitor cilostazol, and ATP-sensitive potassium (KATP) channel opener levcromakalim (LEV) [19,20,21]. NTG causes increased intracellular cGMP and CGRP causes increased intracellular cAMP. Activation of cAMP and cGMP-mediated pathways results in the opening of KATP channels, so the modulation of nociceptive transmission by KATP channel may be a final common pathway in the genesis of a migraine attack [22]. Meanwhile, repeated systemic administration of LEV also induced hind paw and periorbital hyperalgesia in a mouse model, and CGRP signaling was shared in NTG and LEV-induced CMMs [23, 24]. However, whether the activation of specific brain regions could contribute to persistent behavioral sensitization remains unclear, and whether similar activation of brain regions occurs in two CMMs is still unknown. Moreover, it has been reported that gepants (CGRP receptor antagonists) are effective for the acute and preventative treatment of migraine [25]. In CMMs, olcegepant (OLC), a selective CGRP receptor antagonist, significantly alleviates mechanical hypersensitivity [23, 24, 26]. However, it is still unknown whether OLC can alter brain activation to relieve migraine-like pain. To address this question, two CMMs were established to investigate the activation of brain regions by NTG or LEV treatment and the effect of OLC treatment on brain activation by quantifying the expression levels of c-Fos, an immediate-early gene (IEG) widely used for brain activity mapping [27].
Materials and methods
Animals
All experiments with animals were approved by the Animal Ethics Committee in Lanzhou University Second Hospital and carried out according to the criteria outlined in the “Guide for the Care and Use of Laboratory Animals” prepared by the National Academy of Sciences and published by the National Institutes of Health. The sample size of this study was determined based on previous studies [28, 29]. In this experiment, adult male C57BL/6 J mice (18-26 g) were purchased from the Experimental Animal Center of Lanzhou Veterinary Research Institute. All animals were housed under standard conditions with a 12 h light/dark cycle, controlled room temperature, and standard rodent chow diet. In total, 48 male mice were used in the study. Mice were randomly assigned to different experimental groups with 8 male mice. Before all experiments started, mice were given one week to adapt to the experimental environment.
Chronic migraine models
For the establishment of NTG-induced CMM, a stock solution of 5 mg/ml NTG (Beijing Yimin, China), containing 30% propylene glycol and 30% alcohol, was dissolved in water. Prior to the injection, NTG was freshly diluted to 1 mg/ml with 0.9% saline and administered intraperitoneally (i.p.) at a dose of 10 mg/kg every other day for 9 days, based on the literature [28, 29].
For the establishment of LEV-induced CMM, LEV (MedChemExpress, HY-14255), a KATP channel opener, was dissolved in dimethyl-sulfoxide (DMSO) to a final concentration of 5 mg/ml. Prior to the injection, LEV was freshly diluted to 0.1 mg/ml with 0.9% saline by ultrasonic instrument, administered intraperitoneally (i.p.) at a dose of 1 mg/kg every other day for 9 days, as previously described [23, 24].
Drug administration
Olcegepant (MedChemExpress, HY-10095) was dissolved in DMSO to a final concentration of 5 mg/ml. Prior to the injection, OLC was freshly diluted to a final concentration of 0.1 mg/ml with 0.9% saline by ultrasonic instrument. OLC (1 mg/kg) was administered intraperitoneally (i.p.) 15 min prior to the LEV or NTG injection [23, 24]. Accordingly, 2% DMSO + 0.9% saline in the NTG-induced migraine model and 2% DMSO + 2% DMSO in the LEV-induced migraine model were used as the vehicle (VEH) control.
Behavioral tests
All behavioral tests were conducted under low-light conditions between 9:00 and 15:00. Mice were habituated to the behavioral testing room for 2 days prior to behavioral tests. The experiment was double-blinded designed and all data were analyzed by another blinded observer. Previous studies showed that LEV/NTG-induced hyperalgesia was most pronounced 2 h after injection [24, 34]. Thus, the post-treatment mechanical threshold was measured 2 h after NTG or LEV injection (acute hyperalgesia) on each injection day. The basal mechanical threshold (basal hyperalgesia) was measured prior to the VEH, NTG, or LEV injection. For the periorbital mechanical threshold test, the mouse was put into a 4 oz. cup and allowed to adapt for 15 min [28, 29]. Von Frey monofilaments (range from 0.008 to 2 g) were applied perpendicularly to the periorbital region with the up-down method to assess the mechanical threshold. A positive response in the periorbital test was defined as quick retraction of the head from the stimulation or scratching the face with the ipsilateral forepaw. The periorbital test was performed every 4 days to avoid sensitizing the mice [23]. Before the hind paw mechanical threshold test, mice were placed on wire grid floors in clear plexiglass chambers (L X W X H: 10 X 7 X 7 cm) and allowed to habituate for 30 min [23, 24]. Von Frey monofilaments was applied as described above. A positive response in the hind paw test was defined as withdrawal, shaking, or licking of the paw [30, 31]. Finally, a 50% mechanical pain threshold was calculated by the online tool at https://bioapps.shinyapps.io/von-Frey-app/ [32].
Immunofluorescence staining
After the last behavioral test on day 9, in order to reduce bias caused by a small sample size, we randomly selected 5 mice from 8 mice for immunostaining analysis in each group. Mice were deeply anesthetized with 1% pentobarbital and transcardially perfused with 1X phosphate-buffered saline (1 X PBS, pH = 7.4)), followed by 4% paraformaldehyde (PFA). Mouse brains were immediately dissected and postfixed overnight with 4% PFA at 4 °C. Then, the brain tissue was dehydrated in 20% and 30% sucrose solutions sequentially until the tissue sank. Tissue blocks were prepared by embedding them in Tissue-Tek O.C.T. Compound (Sakura 4583). The brain tissue was frozen and sectioned on a cryostat microtome (Leica, CM1950) to obtain 40 μm-thick sections. For immunostaining, the tissue sections were washed with 1 X PBS for a 5- minutes incubation and permeabilized with 0.4% Triton X-100 for 30 min at room temperature. After a wash with clean 1 X PBS buffer for a second 5 min incubation, the tissue sections were followed by incubation with rabbit anti-c-Fos antibody (1:500, Cell Signaling Technology) and mouse anti-NeuN (1:400, Abcam) in antibody dilution buffer (1% BSA, 1 X PBS, 0.4% Tryton X-100) overnight at 4 °C. After primary antibody incubation, the sections were washed for 2 times with 1 X PBS and incubated with following secondary antibodies for 2 h at room temperature: Goat anti-rabbit Alexa Fluor 488 and Goat anti-mouse Alexa Fluor 594 (1:400, Abcam). Cell nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) at room temperature for 10 min. The sections were washed 2 times with 1 X PBS and coverslipped with 50% glycerol for imaging. Images were acquired with a confocal microscope under a 10 X objective (Leica SP8, Germany).
To quantify c-Fos+ and NeuN+ cells, in the medial prefrontal cortex (mPFC), anterior cingulate cortex (ACC), paraventricular thalamic nucleus (PVT), ventrolateral periaqueductal gray (vlPAG), and the caudal part of the spinal trigeminal nucleus (Sp5c) we manually outlined following the atlas of Paxinos (2nd edition from the mouse brain in stereotaxic coordinates) by ImageJ. Every 4 brain slices were selected for the density of c-Fos+ cells (c-Fos+/mm2) and the percentage of c-Fos+ cells in NeuN cells (c-Fos+ NeuN+/NeuN+ cell) in ImageJ. Due to the different depths of specific brain regions, thus the number of sections per brain region was different. In each mouse, 2 brain slices were quantified for mPFC, and 6 brain slices were quantified for PVT, vlPAG, and Sp5c, respectively.
Statistical analysis
All data were presented as mean ± SEM. Data analysis was performed by PRISM 9.3 software (GraphPad, San Diego, CA). For behavioral test data, including drug administration and different time points, two-way ANOVA with the Tukey post hoc test was used. For c-Fos density quantifications data comparisons in three groups, one-way ANOVA followed by the Tukey post hoc test was used to determine statically significance. A p-value < 0.05 was defined as statistical significance.
Results
Olcegepant alleviated acute hyperalgesia but not basal hyperalgesia in the NTG-induced chronic migraine model
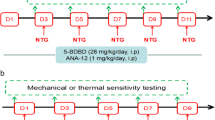
Mice were injected with NTG every other day for 9 days to establish a CMM, and OLC was given 15 min before NTG injection in other treatment groups (Fig. 1A; Fig. S1A). The mechanical thresholds of the hind paw and periorbital area were significantly decreased in the NTG group compared with the VEH group. Interestingly, pre-treatment with OLC in the other NTG group alleviated acute hyperalgesia, but not basal hyperalgesia (n = 8/group, p < 0.01; Fig. 1B, C; Fig. S1B, C). These data indicated the successful establishment of the NTG-induced CMM in mice and OLC could alleviate acute hyperalgesia.

CGRP receptor antagonist (olcegepant) alleviated acute hyperalgesia in the NTG-induced chronic migraine model. A. Representative schematic diagrams and procedures for the behavioral tests. (Created with BioRender.com) B-C. Repeated NTG administration induced mechanical hyperalgesia of periorbital area (B) and hindpaw (C), alleviated by OLC. Two-way ANOVA with the Tukey post hoc tests; * P < 0.05, **P < 0.01, NTG group compared with the VEH group, n = 8/group; #P < 0.05, ## P < 0.01, NTG group compared with the NTG+OLC group, n = 8/group. Abbreviations: VEH, vehicle; NTG, nitroglycerin; OLC, olcegepant
c-Fos activation map of cortical structures, thalamic and brain stem structures in the NTG-induced chronic migraine model
To detect the neural activity of brain regions and the role of CGRP receptors antagonist in the NTG-induced CMM, we evaluated the activation of different brain regions by using c-Fos mapping. After CMM establishment, mPFC and ACC showed a significant increase in the density of c-Fos+ cells and the percentage of c-Fos+ cells in NeuN+ cells, which were alleviated by OLC (n = 5/group, p < 0.01; Fig. 2A-F). In PVT and vlPAG, no significant difference was found by c-Fos mapping in the VEH, NTG, and pre-treatment with OLC group (n = 5/group, p > 0.05; Fig. 3A-C; Fig. 4 A-C). In the NTG group, Sp5c showed a significant increase in the density of c-Fos+ cells and the percentage of c-Fos+ cells in NeuN+ cells, which was also alleviated by OLC (n = 5/group, p < 0.01; Fig. 4 D-F).

c-Fos activation map of cortical structures in the NTG-induced chronic migraine model. A. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the mPFC. Scale bars =100 μm. B-C. mPFC showed a significant increase of c-Fos+ cells density (B) and the percentage of c-Fos + cells in NeuN + cells (C), alleviated by OLC, n = 5/group. D. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the ACC. Scale bars =100 μm. E–F. ACC showed a significant increase of c-Fos+ cells density (E) and the percentage of c-Fos+ cells in NeuN cells (F), alleviated by OLC, n = 5/group. One-way ANOVA with the Tukey post hoc tests, * P < 0.05, **P < 0.01. Abbreviations: mPFC, medial prefrontal cortex; ACC, anterior cingulate cortex; OLC, olcegepant; NTG, nitroglycerin

c-Fos activation map of thalamic structure in the NTG-induced chronic migraine model. A. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the PVT. Scale bars =50 μm. B-C. In PVT, no significant differences in c-Fos+ cell density (B) and the percentage of c-Fos+ cells in NeuN+ cells (C) were found. n = 5/group. One-way ANOVA with the Tukey post hoc tests, * P < 0.05, **P < 0.01. Abbreviations: PVT, paraventricular thalamic nucleus. NTG, nitroglycerin

c-Fos activation map of brain stem structures in the NTG-induced chronic migraine model. A. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the vlPAG. Scale bars =100 μm. B-C. In vlPAG, no significant differences in c-Fos+ cell density (B) and the percentage of c-Fos+ cells in NeuN cells (C) were found, alleviated by OLC, n = 5/group. D. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the Sp5c. Scale bars =50 μm. E–F. Sp5c showed a significant increase of c-Fos+ cell density (E) and the percentage of c-Fos+ cells in NeuN+ cells (F). alleviated by OLC, n = 5/group. One-way ANOVA with the Tukey post hoc tests, * P < 0.05, **P < 0.01. Abbreviations: vlPAG, ventrolateral periaqueductal gray; OLC, olcegepant; Sp5c, caudal part of the spinal trigeminal nucleus
Olcegepant alleviated acute hyperalgesia and basal hyperalgesia in the LEV-induced chronic migraine model
Previous studies indicated that CGRP signaling as a critical factor participated in NTG and LEV-induced CMMs [22, 23]. Accordingly, mice were injected with LEV every other day for 9 days to establish CMM, and OLC was injected 15 min prior to LEV injection in other treatment groups (Fig. 5A; Fig. S1A). In the LEV-induced CMM, ANOVA analysis indicated that the mechanical thresholds of the hindpaw and periorbital area in the LEV group were significantly decreased compared with the VEH group 2 h after injection. Pre-treatment with OLC alleviated acute and basal hyperalgesia (n = 8/group, p < 0.01; Fig. 5B, C; Fig. S1B, C). These data indicated that we established a reliable LEV-induced CMM in mice and OLC alleviated acute and basal hyperalgesia.

CGRP receptor antagonist (olcegepant) alleviated acute hyperalgesia in the LEV-induced chronic migraine model. A. Representative schematic diagrams and procedures for the behavioral tests (Created with BioRender.com). B-C. Repeated LEV administration induced mechanical hyperalgesia of periorbital area (B) and hindpaw (C), alleviated by OLC. Two-way ANOVA with the Tukey post hoc tests, *P < 0.05, **P < 0.01, LEV group compared with the VEH group, n = 8/group; #P < 0.05, # # P < 0.01, LEV group compared with the LEV+OLC group, n = 8/group. Abbreviations: VEH, vehicle; OLC, olcegepant; LEV, levcromakalim
c-Fos activation map of cortical structures, thalamic, and brain stem structures in the LEV-induced chronic migraine model
It remains unknown whether similar activation of brain regions occurs in the LEV-induced CMM. We detected activation in the different brain regions using c-Fos and NeuN staining. After CMM establishment, mPFC showed a significant increase in the density of c-Fos+ cells and the percentage of c-Fos+ cells in NeuN+ cells, which were alleviated by OLC (n = 5/group, p < 0.01; Fig. 6A-C). In the ACC, no significant differences in the density of c-Fos+ cells and the percentage of c-Fos+ cells in NeuN+ cells were found (n = 5/group, p > 0.05; Fig. 6D-F). PVT showed a significant increase in the density of c-Fos+ cells and the percentage of c-Fos+ cells in NeuN+ cells after LEV treatment, which was alleviated by OLC (n = 5/group, p < 0.01; Fig. 7A-C). However, there was no significant difference in the vlPAG among the three groups (n = 5/group, p > 0.05; Fig. 8A-C). Sp5c showed a significant increase in the density of c-Fos+ cells and the percentage of c-Fos+ cells in NeuN+ cells in the LEV group, which was alleviated by OLC (n = 5/group, p < 0.05; Fig. 8 D-F).

c-Fos activation map of cortical structures in the LEV-induced chronic migraine model. A. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the mPFC. Scale bars =100 μm. B-C. mPFC showed a significant increase of c-Fos+ cell density (B) and the percentage of c-Fos+ cells in NeuN+ cells (C). alleviated by olcegepant. n = 5/group. D. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the ACC. Scale bars =100 μm. E–F. In the ACC, no significant differences in c-Fos+ cell density (E) and the percentage of c-Fos+ cells in NeuN cells (F) were found. n = 5/group. One-way ANOVA with the Tukey post hoc tests, * P < 0.05, **P < 0.01. Abbreviations: mPFC, medial prefrontal cortex; ACC, anterior cingulate cortex; OLC, olcegepant; LEV, levcromakalim

c-Fos activation map of thalamic structure in the LEV-induced chronic migraine model. A. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the PVT. Scale bars =50 μm. B-C. PVT showed a significant increase of c-Fos+ cell density (B) and the percentage of c-Fos+ cells in NeuN+ cells (C), alleviated by OLC. n = 5/group. One-way ANOVA with the Tukey post hoc tests, * P < 0.05, **P < 0.01. Abbreviations: PVT, paraventricular thalamic nucleus; OLC, olcegepant. LEV, levcromakalim

c-Fos activation map of brain stem structures in the LEV-induced chronic migraine model. A. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the vlPAG. Scale bars =100 μm. B-C. In vlPAG, no significant differences in c-Fos+ cell density (B) and the percentage of c-Fos + cells in NeuN+ cells (C) were found. n = 5/group. D. Representative pictures of c-Fos (red) and NeuN (green) immunofluorescence labeling in the Sp5c. Scale bars =50 μm. E–F. Sp5c showed a significant increase of c-Fos+ cell density (E) and the percentage of c-Fos+ cells in NeuN+ cells (F), alleviated by OLC. n = 5/group. One-way ANOVA with the Tukey post hoc tests, * P < 0.05, **P < 0.01. Abbreviations: vlPAG, ventrolateral periaqueductal gray; OLC, olcegepant; Sp5c, caudal part of the spinal trigeminal nucleus; LEV, levcromakalim
Discussion
Herein, we provide evidence that both NTG and LEV treatment can result in persistently mechanical hyperalgesia accompanied by alterations of brain activation patterns (Fig. 9 A-C). Meanwhile, OLC alleviated hyperalgesia of the hind paw and periorbital area and decreased activation in several brain regions. These alterations in brain regions help us better understanding the pathogenesis of chronic migraine and providing a potential target for the treatment of CM.

Brain activation in chronic migraine models. A-B. The relative c-Fos+ neuron density in several brain regions (A, red) and the relative c-Fos+ neuron density with OLC treatment (B, green) in the NTG-induced chronic migraine model or LEV-induced chronic migraine model. C. The brain activation and alteration of brain activation with OLC treatment in the NTG-induced chronic migraine model (dark red) and LEV-induced chronic migraine model (pink); No activation (gray); CGRP receptor (green); (Created with BioRender.com). Abbreviations: VEH, vehicle; NTG, nitroglycerin; LEV, levcromakalim; OLC, olcegepant; mPFC, medial prefrontal cortex; ACC, anterior cingulate cortex; PVT, paraventricular thalamic nucleus; vlPAG, ventrolateral periaqueductal gray; Sp5c, caudal part of the spinal trigeminal nucleus
OLC alleviated acute hyperalgesia but no response in alleviating the basal hyperalgesia in the NTG-induced CMM. Recent studies have reported similar results [23, 26]. These results might be caused by a relatively short plasma half-life of OLC in rodents. However, in the LEV-induced CMM, OLC alleviated both acute and basal hyperalgesia. The activation of CGRP signaling and the opening of KATP channels were identified in both CMMs [23, 24, 33]. The different results of OLC treatment might be caused by following reasons. Firstly, NTG can activate the sensory chemoreceptor channel TRPA1 of trigeminal afferents rather than LEV [23, 34]. Secondly, NTG promotes central neuroinflammation by increasing blood–brain barrier (BBB) permeability [35]. These explained the different mechanisms contributing to migraine-like pain in two CMMs. Meanwhile, LEV can cross the BBB [36] and induce KATP channel opening, leading to the triggering of aura and migraine headache through distinct mechanisms in humans [20, 21]. It is widely thought that LEV may increase the extracellular potassium concentration in glial cells, cortical neurons, and cerebral vasculature [37] or lead to stimulation of hyperpolarization-activated cyclic nucleotide-gated cation channels [38], resulting in the generation of cortical spreading depolarization (CSD). CSD has been hypothesized to be the underlying mechanism of the migraine aura [39]. CSD might play an important role in LEV-induced CMM. OLC relieves basal hyperalgesia in the LEV-induced CMM, not in the NTG-induced CMM. A recent study showed CGRP antagonism reduces CSD, supporting the possible use of drugs targeting central CGRP receptors as antimigraine agents [40] and CGRP receptor was widely expressed in the central nervous system, including the cortex, and thalamus, PAG, and Sp5c [41,42,43,44]. Accordingly, a similar activation pattern of mPFC was found in both CMMs, owing to the shared CGRP signaling. Activation of mPFC was reversed by OLC. Meanwhile, in CM patients, resting-state functional connectivity of the default mode network also decreased in the region of interests of the lateral parietal cortex and mPFC, and headache frequency was negatively correlated with the volume of the mPFC [7, 12]. These studies also corroborated that the activation of mPFC was involved in persistent mechanical sensitization in two CMMs. In the NTG-induced CMM, the activation of ACC was consistent with clinical imaging studies and the finding in genetic migraine models. For instance, in CM patients, stronger structural connectivity was found between the caudal ACC and other brain regions [5, 10] and the N-acetyl-aspartate of bilateral thalami and right ACC decreased [6]. In the familial hemiplegic migraine type 2 mouse model, migraine-relevant hypersensitivity triggered by NTG has been attributed to the alteration of neural function in the cingulate cortex [45]. However, unlike NTG, in the LEV-induced CMM, no significant difference of c-Fos expression in ACC was found. Moreover, in the NTG-induced CMM with early growth response gene 1 (Egr1)-enhanced green fluorescent protein transgenic mice, no significant difference in Egr1 expression was found in the ACC [18]. These results were also explained by the analysis of different IEG markers of active neurons.
Thalamic central sensitization maybe contributes to the chronification of migraine [4]. In this study, neuronal activity of the PVT increased in the LEV-induced CMM. It is well-known that PVT plays a critical role in the central processing of chronic pain [46]. However, no alteration in neuronal activity of PVT was found in the NTG-induced CMM. The PVT-CeA-vlPAG circuit reportedly mediates the central mechanisms of descending pain facilitation underlying persistent pain [46]. Although several clinical studies showed that vlPAG, the descending pain-modulating system, was associated with allodynia [15, 47], vlPAG did not exhibit a significant difference between two CMMs. In other rat migraine models, CSD decreased c-Fos expression in PAG [48], and inflammatory soup administration increased c-Fos expression in PAG [49]. Thus, the mechanism of vlPAG and PVT contributing to migraine-like pain was different in CMMs. Interestingly, neurons in Sp5c were activated in two CMMs. CM patients also exhibited cephalic and extracephalic allodynia, corresponding to the sensitization of the second-order neurons in the Sp5c [50]. Recent studies have also reported microglia activation and aberrant synaptic plasticity in the Sp5c contribute to central sensitization in the NTG-induced CMM [29, 51, 52]. Our results further substantiated that the activation of Sp5c may be involved in persistent mechanical sensitization. As expected, OLC decreased the activation of Sp5c. Immunohistochemical studies performed with rat and human tissue revealed that the CGRP receptor was expressed in the medulla-pons region, and cervical spinal cord [41, 44]. NTG increased the gene expression of CGRP and c-Fos in the Sp5c. Our findings suggest that OLC may act on the CGRP receptor of the Sp5c to attenuate neuronal activity.
There were several strengths and limitations in this study. To the best of our knowledge, this is the first study to report the c-Fos activation map of brain regions in NTG and LEV-induced CMMs and provided a reference for neural circuit study in future research. However, we only focused on five representative coronal sections. These results not necessarily were observed in CM patients, and the function of other brain regions in CMM can’t be excluded entirely. Besides, a recent study showed that basal hyperalgesia was maintained for a week after NTG injection on day 9 [30]. We only focused on the c-Fos of expression in brain regions after NTG injection on day 9, but whether these brain regions are continuously activated after the last NTG/LEV injection need further to be explored.
Conclusions
In conclusion, our study demonstrated the activation of mPFC and Sp5c in both CMMs. OLC may alleviate hyperalgesia of the hind paw and periorbital area by attenuating brain activation in CMMs.
Availability of data and materials
The data used and analyzed in this article are available upon reasonable request.
Abbreviations
- ACC:
-
Anterior cingulate cortex
- ANOVA:
-
Analysis of variance
- BBB:
-
Blood–brain barrier
- CM:
-
Chronic migraine
- CMM:
-
Chronic migraine model
- CSD:
-
Cortical spreading depolarization
- CGRP:
-
Calcitonin gene-related peptide
- DMSO:
-
Dimethyl sulfoxide
- Egr1:
-
Early growth response gene 1
- IEG:
-
Immediate-early gene
- KATP :
-
ATP-sensitive potassium channel opener
- LEV:
-
Levcromakalim
- mPFC:
-
Medial prefrontal cortex
- NeuN:
-
Neuronal nuclei
- NTG:
-
Nitroglycerin
- OLC:
-
Olcegepant
- PVT:
-
Paraventricular thalamic nucleus
- Sp5c:
-
Caudal part of the spinal trigeminal nucleus
- SEM:
-
Standard error of the mean
- vlPAG:
-
Ventrolateral periaqueductal gray
- VEH:
-
Vehicle
References
Goadsby PJ, Holland PR, Martins-Oliveira M, Hoffmann J, Schankin C, Akerman S (2017) Pathophysiology of Migraine: A Disorder of Sensory Processing. Physiol Rev 97(2):553–622. https://doi.org/10.1152/physrev.00034.2015
Brennan KC, Pietrobon D (2018) A systems neuroscience approach to migraine. Neuron 97(5):1004–1021. https://doi.org/10.1016/j.neuron.2018.01.029
Maniyar FH, Sprenger T, Monteith T, Schankin C, Goadsby PJ (2014) Brain activations in the premonitory phase of nitroglycerin-triggered migraine attacks. Brain 137(Pt 1): 232–241. https://doi.org/10.1093/brain/awt320
May A, Schulte LH (2016) Chronic migraine: risk factors, mechanisms and treatment. Nat Rev Neurol 12(8):455–464. https://doi.org/10.1038/nrneurol.2016.93
Schwedt TJ, Schlaggar BL, Mar S et al (2013) Atypical resting-state functional connectivity of affective pain regions in chronic migraine. Headache 53(5):737–751. https://doi.org/10.1111/head.12081
Niddam DM, Lai KL, Tsai SY et al (2018) Neurochemical changes in the medial wall of the brain in chronic migraine. Brain 141(2):377–390. https://doi.org/10.1093/brain/awx331
Androulakis XM, Krebs K, Peterlin BL et al (2017) Modulation of intrinsic resting-state fMRI networks in women with chronic migraine. Neurology 89(2):163–169. https://doi.org/10.1212/WNL.0000000000004089
Coppola G, Petolicchio B, Direnzo A et al (2017) Cerebral gray matter volume in patients with chronic migraine: correlations with clinical features. J Headache Pain 18(1):115. https://doi.org/10.1186/s10194-017-0825-z
Neeb L, Bastian K, Villringer K, Israel H, Reuter U, Fiebach JB (2017) Structural Gray Matter Alterations in Chronic Migraine: Implications for a Progressive Disease? Headache 57(3):400–416. https://doi.org/10.1111/head.13012
Planchuelo-Gómez Á, García-Azorín D, Guerrero ÁL, Aja-Fernández S, Rodríguez M, de Luis-García R (2020) Structural connectivity alterations in chronic and episodic migraine: A diffusion magnetic resonance imaging connectomics study. Cephalalgia 40(4):367–383. https://doi.org/10.1177/0333102419885392
Woldeamanuel YW, DeSouza DD, Sanjanwala BM, Cowan RP (2019) Clinical Features Contributing to Cortical Thickness Changes in Chronic Migraine - A Pilot Study. Headache 59(2):180–191. https://doi.org/10.1111/head.13452
Chen XY, Chen ZY, Dong Z, Liu MQ, Yu SY (2020) Regional volume changes of the brain in migraine chronification. Neural Regen Res 15(9):1701–1708. https://doi.org/10.4103/1673-5374.276360
Schulte LH, Allers A, May A (2017) Hypothalamus as a mediator of chronic migraine: Evidence from high-resolution fMRI. Neurology 88(21):2011–2016. https://doi.org/10.1212/WNL.0000000000003963
Lee MJ, Park BY, Cho S, Kim ST, Park H, Chung CS (2019) Increased connectivity of pain matrix in chronic migraine: a resting-state functional MRI study. J Headache Pain 20(1):29. https://doi.org/10.1186/s10194-019-0986-z
Domínguez C, López A, Ramos-Cabrer P et al (2019) Iron deposition in periaqueductal gray matter as a potential biomarker for chronic migraine. Neurology 92(10):e1076–e1085. https://doi.org/10.1212/WNL.0000000000007047
Chou TM, Chen SP (2018) Animal Models of Chronic Migraine. Curr Pain Headache Rep 22(6):44. https://doi.org/10.1007/s11916-018-0693-5
Levine A, Vanderah TW, Largent-Milnes TM (2021) An underrepresented majority: A systematic review utilizing allodynic criteria to examine the present scarcity of discrete animal models for episodic migraine. Cephalalgia 41(3):404–416. https://doi.org/10.1177/0333102420966984
Wang F, Jiang W, Gao L et al (2021) Detecting Abnormal Neuronal Activity in a Chronic Migraine Model by Egr1-EGFP Transgenic Mice. Front Neurosci 15:705938. https://doi.org/10.3389/fnins.2021.705938
Ashina M, Hansen JM, Dunga Á, BO, Olesen J (2017) Human models of migraine - short-term pain for long-term gain. Nat Rev Neurol 13(12):713–724. https://doi.org/10.1038/nrneurol.2017.137
Al-Karagholi MA, Ghanizada H, Nielsen CAW, Hougaard A, Ashina M (2021) Opening of ATP sensitive potassium channels causes migraine attacks with aura. Brain 144(8):2322–2332. https://doi.org/10.1093/brain/awab136
Al-Karagholi MA, Hansen JM, Guo S, Olesen J, Ashina M (2019) Opening of ATP-sensitive potassium channels causes migraine attacks: a new target for the treatment of migraine. Brain 142(9):2644–2654. https://doi.org/10.1093/brain/awz199
Ashina M (2020) Migraine. N Engl J Med 383(19):1866–1876. https://doi.org/10.1056/NEJMra1915327
Christensen SL, Rasmussen RH, Ernstsen C et al (2021) CGRP-dependent signalling pathways involved in mouse models of GTN- cilostazol- and levcromakalim-induced migraine. Cephalalgia 41(14):1413–1426. https://doi.org/10.1177/03331024211038884
Christensen SL, Munro G, Petersen S et al (2020) ATP sensitive potassium (KATP) channel inhibition: A promising new drug target for migraine. Cephalalgia 40(7):650–664. https://doi.org/10.1177/0333102420925513
Ferrari MD, Goadsby PJ, Burstein R, et al (2022) Migraine. Nat Rev Dis Primers 8, 2. https:// doi.org/https://doi.org/10.1038/s41572-021-00328-4
Christensen SL, Petersen S, Kristensen DM, Olesen J, Munro G (2019) Targeting CGRP via receptor antagonism and antibody neutralisation in two distinct rodent models of migraine-like pain. Cephalalgia 39(14):1827–1837. https://doi.org/10.1177/0333102419861726
Silva BA, Burns AM, Gräff J (2019) A cFos activation map of remote fear memory attenuation. Psychopharmacology(Berl) 236(1):369–381. https://doi.org/10.1007/s00213-018-5000-y
Zhang Y, Zhang Y, Tian K et al (2020) Calcitonin gene-related peptide facilitates sensitization of the vestibular nucleus in a rat model of chronic migraine. J Headache Pain 21(1):72. https://doi.org/10.1186/s10194-020-01145-y
Pan Q, Wang Y, Tian R et al (2022) Sphingosine-1 phosphate receptor 1 contributes to central sensitization in recurrent nitroglycerin-induced chronic migraine model. J Headache Pain 23(1):25. https://doi.org/10.1186/s10194-022-01397-w
Pradhan AA, Smith ML, Mcguire B et al (2014) Characterization of a novel model of chronic migraine. Pain 155(2):269–274. https://doi.org/10.1016/j.pain.2013.10.004
Moye LS, Pradhan AAA (2017) Animal Model of Chronic Migraine-Associated Pain. Curr Protoc Neurosci 80:9.60.1–9.60.9. https://doi.org/10.1002/cpns.33
Christensen SL, Hansen RB, Storm MA et al (2020) Von Frey testing revisited: Provision of an online algorithm for improved accuracy of 50% thresholds. Eur J Pain 24:783–790. https://doi.org/10.1002/ejp.1528
Christensen SL, Rasmussen RH, Cour S et al (2022) Smooth muscle ATP-sensitive potassium channels mediate migraine-relevant hypersensitivity in mouse models. Cephalalgia 42(2):93–107. https://doi.org/10.1177/03331024211053570
Marone IM, De Logu F, Nassini R et al (2018) TRPA1/NOX in the soma of trigeminal ganglion neurons mediates migraine-related pain of glyceryl trinitrate in mice. Brain 141(8):2312–2328. https://doi.org/10.1093/brain/awy177
Chen H, Tang X, Li J et al (2022) IL-17 crosses the blood-brain barrier to trigger neuroinflammation: a novel mechanism in nitroglycerin-induced chronic migraine. J Headache Pain 23(1):1. https://doi.org/10.1186/s10194-021-01374-9
Redrobe JP, Pinot P, Bourin M (1996) The effect of the potassium channel activator, cromakalim, on antidepressant drugs in the forced swimming test in mice. Fundam Clin Pharmacol 10(6):524–528. https://doi.org/10.1111/j.1472-8206.1996.tb00610.x
Al-Karagholi MA, Hansen JM, Severinsen J, Jansen-Olesen I, Ashina M (2017) The KATP channel in migraine pathophysiology: a novel therapeutic target for migraine. J Headache Pain 18(1):90. https://doi.org/10.1186/s10194-017-0800-8
Yee AG, Lee SM, Hunter MR, Glass M, Freestone PS, Lipski J (2014) Effects of the Parkinsonian toxin MPP+ on electrophysiological properties of nigral dopaminergic neurons. Neurotoxicology 45:1–11. https://doi.org/10.1016/j.neuro.2014.08.009
Charles AC, Baca SM (2013) Cortical spreading depression and migraine. Nat Rev Neurol 9(11):637–644. https://doi.org/10.1038/nrneurol.2013.192
Bhatt DK, Gupta S, Ploug KB, Jansen-Olesen I, Olesen J (2014) mRNA distribution of CGRP and its receptor components in the trigeminovascular system and other pain related structures in rat brain, and effect of intracerebroventricular administration of CGRP on Fos expression in the TNC. Neurosci Lett 559:99–104. https://doi.org/10.1016/j.neulet.2013.11.057
Tozzi A, de Iure A, Di Filippo M et al (2012) Critical role of calcitonin gene-related peptide receptors in cortical spreading depression. Proc Natl Acad Sci U S A 109(46):18985–18990. https://doi.org/10.1073/pnas.1215435109
Warfvinge K, Edvinsson L, Pickering DS, Sheykhzade M (2019) The Presence of Calcitonin Gene-Related Peptide and Its Receptors in Rat, Pig and Human Brain: Species Differences in Calcitonin Gene-Related Peptide Pharmacology. Pharmacology 104(5–6):332–341. https://doi.org/10.1159/000502471
Warfvinge K, Edvinsson L (2019) Distribution of CGRP and CGRP receptor components in the rat brain. Cephalalgia 39(3):342–353. https://doi.org/10.1177/0333102417728873
Greco R, Demartini C, Zanaboni AM, Tassorelli C (2018) Chronic and intermittent administration of systemic nitroglycerin in the rat induces an increase in the gene expression of CGRP in central areas: potential contribution to pain processing. J Headache Pain 19(1):51. https://doi.org/10.1186/s10194-018-0879-6
Romanos J, Benke D, Pietrobon D, Zeilhofer HU, Santello M (2020) Astrocyte dysfunction increases cortical dendritic excitability and promotes cranial pain in familial migraine. Sci Adv 6(23):eaaz1584. https://doi.org/10.1126/sciadv.aaz1584
Liang SH, Zhao WJ, Yin JB et al (2020) A Neural Circuit from Thalamic Paraventricular Nucleus to Central Amygdala for the Facilitation of Neuropathic Pain. J Neurosci 40(41):7837–7854. https://doi.org/10.1523/JNEUROSCI.2487-19.2020
Schwedt TJ, Larson-Prior L, Coalson RS et al (2014) Allodynia and descending pain modulation in migraine: a resting state functional connectivity analysis. Pain Med 15(1):154–165. https://doi.org/10.1111/pme.12267
Borysovych Bogdanov V, Bogdanova OV, Lombard A et al (2015) Cortical spreading depression decreases Fos expression in rat periaqueductal gray matter. Neurosci Lett 585:138–143. https://doi.org/10.1016/j.neulet.2014.11.026
Zeng X, Niu Y, Qin G, Zhang D, Zhou J, Chen L (2020) Deficiency in the function of inhibitory interneurons contributes to glutamate-associated central sensitization through GABABR2-SynCAM1 signaling in chronic migraine rats. FASEB J 34(11):14780–14798. https://doi.org/10.1096/fj.202001561R
Aurora SK (2009) Spectrum of illness: understanding biological patterns and relationships in chronic migraine. Neurology 72(5Suppl): S8-S13. https://doi.org/10.1212/WNL.0b013e31819749fd
Long T, He W, Pan Q et al (2020) Microglia P2X4R-BDNF signalling contributes to central sensitization in a recurrent nitroglycerin-induced chronic migraine model. J Headache Pain 21(1):4
Wang Y, Pan Q, Tian R et al (2021) Repeated oxytocin prevents central sensitization by regulating synaptic plasticity via oxytocin receptor in a chronic migraine mouse model. J Headache Pain 22(1):84
Acknowledgements
We are sincerely grateful to all the staff of Lanzhou Veterinary Research Institute and Headache Center, Beijing Tiantan Hospital, Capital Medical University. We thank Chengze Wang, and Weiqing Jiang for comments on the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (No: 32170752, No: 91849104, No: 31770800) and the National Natural Science Foundation of Beijing (Z200024).
Author information
Authors and Affiliations
Contributions
This study was designed by YGW, XR, and SYW. SYW, and XJL participated in behavioral testing and immunofluorescence staining. ZCL and XR performed the statistical analysis. SYW, ZCL, and XR write the manuscript. ZLL, KBZ, and WW revised the manuscript. XR and YGW provided supervision and a final check. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All animal experiments performed in this study were approved by the Animal Ethics Committee of Lanzhou University Second Hospital.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
CGRP receptor antagonist (olcegepant) did not alleviate basal hyperalgesia in the NTG-induced chronic migraine model. A. Representative schematic diagrams and procedures for the behavioral tests. B-C. Repeated NTG administration induced basal hyperalgesia of periorbital area (B) and hindpaw (C), but not alleviated by OLC. Two-way ANOVA with the Tukeypost hoc tests; * P<0.05, **P<0.01, NTG group compared with the VEH group, n=8/group; #P<0.05, ## P<0.01, NTG group compared with the NTG+OLC group, n=8/group. Abbreviations: VEH, vehicle; NTG, nitroglycerin; OLC, olcegepant. Figure S2. CGRP receptor antagonist (olcegepant) alleviated basal hyperalgesia in the LEV-induced chronic migraine model. A. Representative schematic diagrams and procedures for the behavioral tests. B-C. Repeated LEV administration induced mechanical hyperalgesia of periorbital area (B) and hindpaw (C) alleviated by OLC. Two-way ANOVA with the Tukey post hoc tests; * P<0.05, **P<0.01, LEV group compared with the VEH group, n=8/group; #P<0.05, ## P<0.01, LEV group compared with the LEV+OLC group, n=8/group. Abbreviations: VEH, vehicle; NTG, nitroglycerin; OLC, olcegepant; LEV, levcromakalim.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wu, S., Ren, X., Zhu, C. et al. A c-Fos activation map in nitroglycerin/levcromakalim-induced models of migraine. J Headache Pain 23, 128 (2022). https://doi.org/10.1186/s10194-022-01496-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10194-022-01496-8