
Abstract
This study aimed to investigate the relationship between moisture dynamics of sorbitol and citric acid (SorCA) modified wood and its biological durability. Specifically, the research aimed to determine the chemical loading needed for effective protection against wood-destroying basidiomycetes, while also improving the understanding of the moisture behavior of SorCA-modified wood. The SorCA modification process is relatively new, and thus, there is limited knowledge on its moisture behavior and its impact on biological durability. The research focused on Scots pine sapwood (Pinus sylvestris L.) and used the EN 113-2 standard to investigate its durability against wood-destroying basidiomycetes. Moisture behavior was analyzed through short-term water uptake and release tests, capillary water uptake and CEN/TS 16818. Results showed a significant reduction in liquid and water vapor uptake, likely due to a reduction in the maximum moisture capacity within the wood cell wall. The study confirmed that high chemical loadings (i.e., weight percent gain, WPG) are necessary for adequate decay protection.
Similar content being viewed by others


Introduction
Wood is widely recognized as the most carbon–neutral construction material and new political initiatives such as the European Green Deal are highly encouraging its use in load-bearing structures, façade claddings and terrace deckings. However, most wood species grown in Europe are susceptible to biological degradation and require preventive protection. With the implementation of the Biocidal Products Regulation [1], many biocidal agents in wood preservatives have become restricted to certain applications. Therefore, wood modification has gained increasing interest as a biocide-free treatment that enhances the biological durability of wood [2] while also improving other properties, such as dimensional stability [3, 4] and hydrophobicity [5, 6].
Chemical wood modification with sorbitol and citric acid (SorCA) has emerged as a promising alternative to already existing treatment methods [7, 8]. In March 2022, a commercial entity, Ciol AS, was formed as a result of a joint collaboration between the Norwegian Institute of Bioeconomy Research (NIBIO) and a technology transfer office, Ard Innovation [9]. Ciol® and SorCA are the same processes based on the same chemicals, molar ratios of citric acid (CA) to sorbitol and reaction temperatures. Both chemicals used for impregnation are bio-derived and affordable, no catalyst is required for the reaction and the only by-product is water. The modification is based on a polyesterification reaction of citric anhydride and sorbitol, which follows a nucleophilic addition mechanism [10]. As a result, a cross-linked network of SorCA polyesters is formed inside the wood cell wall matrix. These newly formed esters may also react with the hydroxyl groups of wood polymer constituents [7, 10].
Until now, only a limited number of studies have investigated the biological durability of SorCA-modified wood, with a particular focus on its resistance to wood-destroying basidiomycetes [8, 11,12,13]. Larnøy et al. [8] conducted a mini-block [14] durability trial, in which Scots pine sapwood was incubated with Rhodonia placenta (R. placenta) and Trametes versicolor (T. versicolor). The results showed that the amount of chemicals remaining in the wood after the leaching procedure was still sufficient for decay protection, regardless of the curing temperature used (103 °C and 140 °C). However, fixation was significantly higher at 140 °C. This finding was later confirmed by Alfredsen et al. [11], who showed that a temperature of 140 °C was necessary to prevent leaching and provide high durability. In this study, Scots pine sapwood samples with a wide range of weight percent gain (WPG) values were tested. The results showed that to achieve the highest durability class (DC) 1 “very durable” according to EN 350 standard [15], WPGs of 21% and 62% were, respectively, required for an adequate protection against T. versicolor and R. placenta. Similar decay protection thresholds were determined when sterile soil was used as a fungal growth medium instead of malt agar. In another study on Scots pine sapwood, Beck [12] established the durability threshold for R. placenta at a WPG of 46% and for T. versicolor at a WPG of 20%. Even lower WPG values were reported for European beech (Fagus sylvatica L., [13]), where only a WPG of 22% was required for adequate protection against both brown rot and white rot fungi (C. puteana and T. versicolor). However, none of the mentioned studies followed the EN 113–2 [16], as different incubation times, sample sizes or growth media were used. Brischke et al. [17] showed that the assessment of the DC might be incorrect if any alternations are made in regard to the sample dimensions or incubation times recommended in the intended standard. Considering, that the Bravery test [14] is commonly used as a quick alternative for the standardized EN 113–2 [16] procedure, the study by Brischke et al. put into question all studies done based on the altered test procedures.
Fungi require a nutrient source (such as wood), water and oxygen to successfully colonize wood [18]. So far, only one study [12] attempted to explore the relationship between the decay protection and wood moisture behavior. The study found that SorCA treatment increased the moisture content (MC) within the cell wall, which is contrary to other wood modification methods that aim to lower the fiber saturation point (FSP) as a mechanism of wood protection against fungal degradation [19, 20]. In general, fungal hyphae are small enough to penetrate the cell lumen. Therefore, to create favorable decay conditions, the MC of wood must be above the FSP (approx. 30% MC, [21]) so that enzymes and metabolites secreted by fungi can diffuse to the wood cell wall. Free water is necessary for this process [18, 20, 22]. However, a recent study suggested that the intra-cell-wall diffusion of inorganic ions in wood is a Fickian diffusion process, occurring when the hemicellulose reaches its glass transition temperature (at approx. 75% RH, [21]), not an aqueous process as previously thought [23]. It increases the hypothesis that it is a rubbery state of amorphous polysaccharides which is required for decay, not a certain amount of free water in lumen. The same study determined that this transition (i.e. softening) occurred when wood reached ca. 10–15% MC and was not related to the amount of free water in the wood cavities. That would suggest that the brown rot and white rot decay can proceed when the MC is significantly below the FSP. Modified wood needs a higher RH for softening [24], which can potentially delay the onset of decay.
Although there is no consensus on what the mode of protection in chemically modified wood is, existing data suggest that the degradation mechanism of brown rot and white rot decay follow the same path as for untreated wood, including similar effects on water relations [25]. For the brown rot fungi, a modus operandi is presumably the moisture exclusion via the reduction of cell wall voids [26, 27], resulting in the reduction of maximum moisture capacity of the wood cell wall [28]. The reduction in available space for water molecules inside the wood cell wall inhibits the diffusion of fungal reductants, hydrogen peroxide and oxalic acid and, thereby, impeding the formation of hydroxyl radicals, which are essential for the oxidative degradation. The transport of cations in chemically modified wood is known to be restricted, depending on the treatment level (WPG) and on the relative humidity (RH) surrounding the specimen [29]. Therefore, if the oxidation is prevented, the hydrolyzing enzymes cannot penetrate the wood cell walls, as they are too large and require prior depolymerization of hemicelluloses by the hydroxyl radicals [26].
All these considered, obtaining an in-depth understanding of the moisture behavior of SorCA-modified wood appears to be crucial in comprehending its protective mechanism against wood-destroying fungi. So far, the mode of protective action of SorCA-modified wood has been not fully explained. To address this gap, this study examined a series of Scots pine sapwood samples for both durability against basidiomycetes and moisture-related properties. The study attempted to establish a relationship between the durability and moisture dynamics data. Additionally, given the discrepancies previously described by Brischke et al. [17], this study was designed to perform the standardized biological durability procedure to confirm the necessity of high chemical loadings (i.e., WPG) for protection against decay.
Material and methods
Wood material and treatment
Scots pine (Pinus sylvestris L.) sapwood specimens in various dimensions (specified in Table 1) were treated with aqueous solutions of technical grade CA monohydrate (approx. 97% purity) from BÜFA Chemikalien GmbH & Co. KG (Oldenburg, Germany) and technical grade sorbitol (approx. 98% purity) from Ecogreen Oleochemicals GmbH (Dessau-Roßlau, Germany).
Prior to the treatment, all specimens were oven-dried at 103 (± 2) °C for 48 h to determine the initial dry mass (MC = 0%). The oven-dried specimens were impregnated in an autoclave in a vacuum-pressure process (5 kPa for 1 h, followed by 1200 kPa for 2 h) with 10%, 20%, 30% and 50% SorCA solutions (molar ratio of polyol to acid was 1:3). Impregnated specimens were pre-dried under room climate conditions (ca. 20 °C, 65% RH) for 168 h and afterwards heat-cured at 140 °C for 24 h (“dry-curing”). However, the specimens for the CEN/TS 16818 test [32], required a different pre-drying procedure ((room climate conditions (168 h), 60 °C (24 h), 80 °C (24 h), and 103 °C (24 h)), due to their bigger dimensions and to avoid drying failures such as cracking. The latter would most likely affect the uptake of liquid water and, thus, the wood–water interaction studies. The amount of deposited chemicals within the wood structure, expressed by the WPG was calculated based on the dry mass before (M1) and after (M2) the treatment, according to the Eq. 1.
Prior testing, all specimens which were foreseen for biological decay testing and half of each collective for moisture behavior testes, underwent a cold-water leaching procedure with deionized water according to EN 84 [30]. Changes in dry mass after leaching (M3) were calculated according to Eq. 2.
Table 1 summarizes all wood specimens, dimensions and treatment levels, together with the mean weight-percent-gain (WPG) and standard deviation of each collective before and after leaching according to EN 84 [30].
Decay tests
Durability tests with basidiomycetes
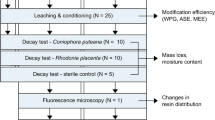
A decay test with basidiomycete monocultures was performed according to EN 113–2 [16] and durability classes assigned according to EN 350 [15]. Used specimens of 25 × 15 × 50 (L) mm had the longitudinal faces parallel to the direction of the grain and the growth-ring orientation was ca. 45°. For each treatment and fungus, the test was conducted with n = 30 specimens. However, for the untreated control group, only 10 specimens per fungus were tested. Prior incubation, sterilization of untreated and modified wood samples was conducted in an autoclave at 121 °C for 20 min. After 16-week incubation, wood specimens were cleaned from adhering mycelium, then weighed to the nearest 10–3 g directly after the test (wet state, Mw) and after step-wise drying (oven-dry state, M4) and their MC upon incubation and mass loss (ML) caused by a fungal decay calculated according to Eqs. 3–4.
The following basidiomycete monocultures were used:
-
•
Coniophora puteana (Cp) = (Schum.:Fr.) Karst. BAM Ebw.
-
•
Rhodonia placenta (Rp) = (Fr.) Niemelä, K.H. Larsson & Schigel 229.
-
•
Trametes versicolor (Tv) = (L.:Fr.) Pilát CTB 863a.
Moisture behavior
Short-term water uptake and release tests (according to Meyer-Veltrup et al. [31])
For each treatment level, 10 non-leached and 10 leached specimens according to the EN 84 standard [30] of 10 (R) × 5 (T) × 100 (L) mm with growth-rings parallel to the specimens’ edges were dried at 103 ˚C until constant mass (m01). Then, oven-dry specimens were submerged in a plastic container with deionized water, separated from each other with a metal grid, and placed in a conditioned surrounding (20 °C, 65% RH). After 24 h of submersion, the specimens were weighted (msubmerged) and their liquid water uptake (W24submersion) determined according to:
After submersion, the specimens were oven-dried until constant mass again (m01) at 103 ˚C. Dried specimens were then placed above water surface with distance to each other on stainless-steel plates in a sealed plastic box filled with deionized water. After 24 h storage in the climate room the specimens were weighted (m100%RH) and the water vapor uptake in water-saturated atmosphere (W24100%RH) calculated, as in Eq. 6.
After the water vapor uptake test the specimens were put back to the boxes and stored above deionized water (approx. 100% RH) until constant mass and reaching almost cell wall saturation (mCWS). Then the specimens were placed on freshly activated silica gel (moisture absorbent, approx. 0% RH) on the 10 × 100 mm surface in sealed boxes and weighted after 24 h (m0%RH).
Water vapor release (W240%RH) was calculated based on the Eq. 7.
Capillary water uptake test
The capillary water uptake (CWU) was measured using a tailor-made device (CWU-meter). For each treatment level, 20 specimens of 10 × 5 × 100 (ax.) mm were conditioned at 20 ˚C 65% RH until constant mass (m65%RH). Half of each collective was leached prior to CWU measurement according to the standard EN 84 [30]. Subsequently, the specimens were fixed in the tensiometer and their cross-section submerged in water. The specimen’s mass was recorded to the nearest 10–3 g after 200 s (m200s). The CWU was related to the cross-sectional area in the conditioned state (A):
Liquid water uptake and water vapor release—CEN/TS 16818 (2018)
Specimens from 3 different boards were used for the CEN/TS 16818 standard [32] test. Overall, 18 specimens (6 from each board) were used per treatment level, of which half were leached according to the EN 84 standard [30]. As a sealing agent, polyurethane sealant Sikaflex® 221i (Sika Austria GmbH) was used to seal the edges of the specimens (see Fig. 1). The moisture performance was tested during a water uptake (absorption) and release (desorption) cycle following the CEN/TS 16818 standard [32]. All measured mass values were registered to the nearest 10–2 g.

Test specimens for the moisture behavior tests: floating 25 (R) × 50 (T) × 50 (L) mm and submersion test 25 (R) × 50 (T) × 150 (L) mm according to the CEN/TS 16818 standard [32], short-term water uptake and release tests and capillary water uptake test 10 (R) × 5 (T) × 100 (L) mm. The sides of floating and submersion specimens (grey) were sealed with polyurethane sealant Sikaflex.® 221i from Sika Austria GmbH (Austria)
Floating test
Specimens of 25 (R) × 50 (T) × 50 (L) mm were dried at 103 ˚C until constant mass (m0). Prior testing, specimens were conditioned at 20 °C (65% RH). After conditioning, four sides (excluding the tangential sections) were sealed with a polyurethane sealant, the weight of the sealant and the initial specimen mass (mi) recorded. Afterwards, sealed specimens were placed in plastic containers filled with deionized water and stored at 20 ˚C and 65% RH. One of the 50 × 50 surfaces of the specimens was placed under water at approx. 10 mm. Tested specimens were removed from the boxes and weighted after 1, 4, 8, 24, 48, 72 and 144 h. The excess water was removed from the wooden surface with a paper tissue prior weighing. After the absorption cycle, the samples were put on the drying grids on their sealed sides, with sufficient distance between them, and left to dry at 20 °C, 65% RH. During the desorption cycle, the specimens were weighted after 1, 4, 8, 24, 48, 72, 96 and 168 h.
Submersion test
Specimens of 25 (R) × 50 (T) × 150 (L) mm were dried at 103 ˚C until constant mass (m0). Prior testing, specimens were conditioned at 20 °C and 65% RH. After conditioning, specimens were sealed as for the floating test and the respective masses (m0 and mi) recorded. Sealed specimens were placed in the containers filled with deionized water and fully submerged throughout the absorption cycle, by covering them with stainless steel meshes to prevent them from emerging onto the surface. Tested specimens were removed from the boxes and weighted after 1, 4, 8, 24, 48, 72 and 144 h. After the absorption cycle, a desorption cycle was performed for 168 h, as described for the floating test procedure.
Parameters calculated based on the CEN/TS 16818 standard (CEN, 2018)
The conditioned mass of each specimen before the test (mi) and after each time interval (mx), was registered and used for calculating the MC at certain point of the test based on the oven-dry weight (m0) of the specimen, as in the Eq. 9:
where: m0 is the mass of the specimen before floating test (oven-dry) with sealant, mi is the mass of the conditioned specimen before floating test with sealant and MCi is the moisture content of the conditioned specimen before the test. The MC of each specimen after x hours of exposure (e.g., after 72 h, where x = 72), was calculated as follows:
where: mx is the mass of the specimen after x hours during test with sealant and MCx is the MC of the specimen after x hours of exposure.
For the SorCA-modified specimens, the corrected MC (MCcor, %) after 144 h and 312 h of test duration was calculated, to exclude the mass added by the impregnation chemicals:
The amount of absorbed moisture after 144 h (a144) and released moisture after 168 h of desorption (d168) was calculated as in the following equations:
Also, the remaining moisture after water uptake and release test (rm168) after 144 h of uptake and 168 h of release (312 h of testing) was calculated:
And contributed to determining the residue as a percentage of the absorbed moisture (res312) after 144 h of uptake and 168 h of release (312 h of testing):
The illustration of the MC upon absorption and desorption phase of the non-leached Scots pine sapwood reference sample, together with other parameters calculated based on CEN/TS 16818 [32] is presented in Fig. 2.

Moisture content (MC) development over 144 h absorption and 168 h desorption of water determined in a floating test with the characteristic values: the initial moisture content (MCi), the MC after 144 of water absorption (MC144) and after 168 h of desorption (MC312), amount of absorbed moisture (a144) and released moisture (d168), as well as the remaining MC after the floating test (rm168) of the non-leached Scots pine sapwood sample (Emmerich et al. [45], adapted)
Results and discussion
Decay test
Durability tests with basidiomycetes
Overall, the results from this study confirmed that the WPG is a crucial parameter which correlates with the decay resistance of chemically modified wood. As shown in Fig. 3, each of the registered ML values were assigned to a corresponding DC according to the durability classification against basidiomycetes following EN 350 [15] and EN 113-2 [16]. Maximum ML registered for all three tested fungi species was selected as a measure of DC for each treatment level. In this study, the untreated Scots pine sapwood control registered 27% and 43% ML for tested brown rot fungi (R. placenta and C. puteana, respectively) and 17% ML for investigated white rot fungi (T. versicolor). For treated specimens, the ML due to incubation with R. placenta was in a range of 21–24% and only the highest treatment experienced a reduced fungal degradation in comparison to untreated control with merely 5% ML. For the second type of tested brown rot fungi the ML decreased as the WPG increased, from 26% (10% WPG) to barely 2% ML (57% WPG). The ML upon exposure to white rot fungi was overall low and amounted to 6% (the lowest treatment level) and 3% for all higher treatments.

Mean mass loss (ML) after 16-week incubation with the brown rot fungi Rhodonia placenta (a) and Coniophora puteana (b) and the white rot fungus Trametes versicolor (c) on a malt substrate, after leaching according to the EN 84 standard [30]. The following concentrations of the impregnation solution (in wt%) were tested and are given on the x-axis: 10%, 20%, 30%, 50% and 0% for the untreated reference. Beech and pine—virulence controls. The number of specimens can be found in the test descriptions
The decay protection threshold required to achieve the highest durability class (DC 1) against both studied brown rot fungi (R. placenta and C. puteana) and white rot fungi (T. versicolor) was 57% WPG. All other treatments (WPG < 57%) were assigned to DC 4, due to a high ML upon the exposure to R. placenta (Fig. 3). The other type of tested brown rot, C. puteana, experienced more pronounced correlation between the WPG value and measured ML.
Overall, the study confirmed that the results for R. placenta are in line with the previous durability studies on SorCA-modified wood [11, 12], in which the WPG above 50% was claimed to provide a full protection against this fungal strain (DC 1). In this particular case, both EN 113-2 [16] (this study) and Bravery tests [14] (studies by [11, 12]) indicated the necessity of assuring high WPG values (> 50%) for reaching the DC 1. Nevertheless, despite the fact that this study found that different specimen sizes and incubation times resulted in the same durability class, it is important to consider that variations in these parameters could yield different durability classifications [17].
According to the most widely accepted mode of decay protection of chemically modified wood (moisture exclusion via reduction of cell wall voids [26]), the enhanced durability of SorCA-treated wood is attributed to two key factors: the cell wall bulking (CWB) and the cross-linking of SorCA polyesters with wood polymers [7]. These processes significantly limit the spatial availability for water molecules in the wood cell wall, thereby impending the diffusion of low molecular weight compounds, enzymes and ions from hyphae in lumen. On the other hand, the glass transition of hemicelluloses might have been altered as a result of the modification, which could have contributed to the decay resistance. Furthermore, biopolyesters like cutin and suberin, which are composed of long carbon-chains, are recognized as natural decay-protection agents in plants [33, 34]. It is possible that SorCA polyesters may also function in a similar capacity, as they might consist of very long carbon chains under favorable conditions.
Based on the results of ML after 16 weeks of incubation, there is a clear indication of a difference in the wood degradation mechanism by various types of brown rot fungi. While brown rot fungi typically modify lignin structure via depolymerization and rearrangements without degrading it [35], the decay pattern of C. puteana has some similarities to white rot fungi, as it erodes and thins the cell wall and produces laccase, a lignin-degrading enzyme [36]. However, since the activity of laccase has also been noted for R. placenta [37], the presence of this enzyme cannot solely explain the different degradation mechanisms of R. placenta and C. puteana. Additionally, and in contrast to the majority of brown rot fungi, C. puteana produces cellobiose dehydrogenases, which enhance the degradation of cellulose. This unique characteristic introduces some uncertainty regarding its mode of action and positions it at the boundary between brown rot and white rot fungi related to its degradation mechanism [38]. It has been also shown that certain secondary metabolites may act as extracellular ferric ion reductants in R. placenta. As this fungi species has an extensive set of secondary metabolic genes, it may also facilitate the enzymatic stage of decay [38] and lead to ML differences between C. puteana and R. placenta.
On the other hand, it is worth considering that the chemicals used for wood modification are not toxic in use and do not inhibit fungal growth, allowing the colonization of the timber under fungal attack [27, 39, 40]. Although the decay might be significantly delayed, the fungus may eventually modify the environment and be able to degrade wood [41]. The creation of such favorable growth conditions may differ between fungal types or strains, which could explain the less severe degradation by C. puteana. However, this only holds true for higher treatment levels (WPG > 20%). In either way, further investigations are needed to understand the difference in decay rate among various types of brown rot fungi.
It is interesting to note that yellow droplets were observed in the Petri dishes of C. puteana and T. versicolor for the collectives from high WPG values after the basidiomycetes trial. Also, Beck [12] noted a presence of such droplets for R. placenta. He suggested that this droplet formation might be attributed to the “translocation of the hydrolysis products out of the wood sample”. It appears, however, that these are the guttation droplets, caused by a removal of water and dissolved substances from the fungal tissue [42].
Moisture behavior
Short-term water uptake and release tests (according to Meyer-Veltrup et al. [31])
As stated by Van Acker et al. [43]: “Additional to the material resistance parameter ‘biological durability’, there is a need for a [service life prediction] classification based on moisture dynamics”. Therefore, this study aimed to investigate the moisture behavior of identically treated specimens which underwent a biological decay test according to EN 113–2 standard [16] and use it to discuss the effect of wood–water interactions on the biological durability. The short-term water uptake and release test [31] showed an overall decrease of the liquid water uptake (W24submersion) and water vapor uptake (W24100%) for all treated specimens in comparison to the untreated controls, whereas the water vapor release (W240%) increased for all tested collectives (Table 2). After being stored above water for three weeks (before the water vapor release stage), the MC of the untreated collective was approx. 27%, while that of all treated collectives was approx. 20%. This suggests a reduction in the FSP of SorCA-modified specimens.
Corresponding studies on Scots pine sapwood modified with cyclic N-methylol (DMDHEU, mDMDHEU) and N-methyl (DMeDHEU) compounds indicated a decrease of all three W24 parameters [44] in comparison to the untreated reference samples. Similar to the improved resistance against wood-destroying basidiomycetes, a decreased water uptake of treated specimens might be attributed to a reduced spatial availability for water molecules due to CWB and cross-linking. On the other hand, the increased water vapor desorption after SorCA-treatment might be related to the reaction of SorCA polyesters with wood polymer constituents, which modified the cell wall and, therefore, facilitated the transport of water vapor outside the wood structure.
As expected, the wood–water interactions in treated specimens were affected by cold-water leaching. Previous study of the DMDHEU modification system has shown that non-fixed chemical components provide additional sorption sites for water and increase water sorption at certain RH levels [45]. In the present study, water uptake increased after leaching, presumably due to the greater availability of space for water molecules in cell wall matrix and lumen. Surprisingly, however, the leaching did not affect the water vapor release (W240%), which cannot be explained based on our current knowledge.
Capillary water uptake (CWU) test
As wood macropores are open in the cross-section [46], the easiest way to measure CWU of wood is by submerging the end-grains in water. In the present study, all tested collectives experienced a considerable decrease in the CWU in comparison to the untreated reference (Table 2). Similar decreases in the CWU were also observed in wood modified with cyclic N-methylol and N-methyl compounds [44]. Although the leaching procedure caused higher uptake of liquid water, it was still lower than that of the untreated control. This is because the SorCA-modification process fills the wood cell wall matrix with polyesters and creates a dimensionally confined, cross-linked network that reduces water uptake, even in the capillaries along the cross-section. Furthermore, the CWU decreased with increasing chemical loading (WPG), as seen in both liquid water uptake (W24submersion) and vapor water uptake (W24100%). This is presumably due to the higher content of SorCA polyesters and their denser, cross-linked network in the wood macro-voids at higher treatment levels.
Liquid water uptake and water vapor release—CEN/TS 16818 [32]
Liquid water uptake and water vapor release were further tested in a standardized test procedure CEN/TS 16818 [32]. This test was initially designed to be used together with fungal decay test data and as a procedure for service life predictions for certain wooden substrates [43]. The MC development upon the water absorption and desorption can be followed in Fig. 4 (a and b for floating; c and d for submersion test). In general, it can be seen that SorCA-modification critically decreased an average MC of each collective and this decrease was more pronounced for the higher WPG values. A decrease in MC (at high RH values) in comparison to untreated reference was already reported for SorCA-treated wood measured in the dynamic vapor sorption (DVS, [7]). After leaching, as expected based on the W24 and CWU tests, the mean MC increased, but was still considerably reduced when compared to untreated control.

Moisture content (MC) development over 144 h absorption and 168 h desorption of water determined in a floating (a, b) and submersion (c, d) tests of non-leached (a, c) and leached (b, d) specimens
The standard parameters according to the CEN/TS 16818 [32] were calculated and summarized in Tables 3 (floating test) and Table 4 (submersion test). For both tests, a decrease in remaining moisture after 144 h water uptake (a144) and 168 h of water vapor release (d168), as well as after 144 h uptake and 168 h release (rm168) was observed after the SorCA-treatment. Only the residue as a percentage of the absorbed moisture (res312) increased after the SorCA treatment. This decrease (or increase) of the mentioned parameters was overall proportional to the WPG value of the collective. The determined parameters (a144, d168, rm168) were still lower than those of untreated reference after the cold-water leaching, though, the decrease was less pronounced. Even when the additional mass of the impregnation chemicals was considered and the corrected MC (MC144cor and MC312cor) calculated, the advantage of the modification was still significant. Nevertheless, due to a very low water desorption in comparison to absorption for both tests, the residue as a percentage of the absorbed moisture (res312) was significantly higher for treated specimens than for untreated reference.
Emmerich et al. [45] reported a decrease in several tested parameters, except for rm168 for submersion and res312 for floating and submersion, in DMDHEU-treated Scots pine sapwood. They also found that overall, submersion resulted in higher water uptake than the floating test. This was also observed in our study and can be explained by the test setup, as previously noted by Brischke et al. [47]. During absorption, water uptake is higher in the submersion test as the sample is in direct contact with water on both non-sealed surfaces. Then, during the desorption stage, water is released from both sides exposed to water, whereas for floating specimens, water can only be released from the one submerged side. As a result, moisture gradually moves from the submerged to the dry side, becoming located in the inner part of the specimen instead of evaporating.
Mode of protective action against decay
Several studies (e.g., [18, 27, 48]) have demonstrated a correlation between moisture behavior and decay resistance of wood, particularly when it is subjected to a modification treatment. In chemically modified wood, reducing the MC of the cell wall is the primary factor contributing to increased decay resistance. To increase biological durability against wood-destroying basidiomycetes, bulking agents must be introduced into the wood cell wall. Once the bulking threshold is reached (which varies depending on the wood and fungi species), the moisture exclusion level rises to a point where the wood is protected against decay [49]. Papadopoulos and Hill [50] also indicated that decay protection is based on the degree of the CWB; however, they did not mention a specific CWB threshold. In the present study, we found that a WPG of 57% was necessary to achieve the highest decay protection. Based on previous investigations [51], this corresponds to a CWB of approximately 11%. This high value is reflected by a substantial reduction in liquid and vapor water uptake. Predicted on the same publication [51], the following CWB values were obtained for the treatment with lower concentrations of the impregnation solutions (expressed as wt%): 3.5% (10%), 6.5% (20%) and 8.5% (30%).
It is important to emphasize that only filling the cell wall with chemicals may result in long-term decay protection. Wax-based treatments, which only fill the lumen and do not change wood chemistry, have been shown to delay decay onset but eventually fail to provide long-term decay protection during outdoor exposure [52]. This is because the wax treatment creates a firm barrier in the lumen and on the surface of wood specimens, which can temporarily act as a moisture repellent and hinder the diffusion of wood-degrading enzymes from fungal hyphae to the cell wall matrix [52].
The study by Brischke and Melcher [53] investigated the limited durability of wax-treated wood when exposed to outdoor conditions. They conducted tests in lap-joint (UC 3.1, no water accumulation) and double-layer (UC 3.2, water may accumulate) configurations to compare the above-ground performance according to the EN 335 standard [54]. Their findings indicated that wax-treated wood had lower durability in the double-layer test due to moisture trapping. While filling wood macropores inhibited decay, it was not sufficient to inhibit fungal degradation when water accumulated over time.
Further investigation showed that fungi could colonize and degrade wood below the FSP if a moisture source was present in the vicinity [55], or even in the absence of available liquid water [56]. This raises questions about the mechanism of intra-cell-wall diffusion of fungal metabolites. Recent research demonstrated that when the MC of wood reached 10–15%, hemicelluloses passed their glass transition, facilitating Fickian diffusion between fungal hyphae and the wood cell wall matrix, even in the absence of liquid water [23]. This could explain why fungal degradation of SorCA-modified wood could still proceed despite a high moisture exclusion and a significant decrease of the FSP. Furthermore, the diffusion constant varies depending on the type of ion, which could explain the difference in decay rate for different fungal species that release slightly different ions.
The findings presented in this study clearly demonstrate variations in the rate of biological degradation, not only between white rot and brown rot, but also within different species of brown rot fungi. It has previously been suggested that these differences could be attributed to variations in the enzymes and secondary metabolites produced by the fungi. Riley et al. [38] emphasized that the traditional classification into brown rot and white rot does not align with how fungi degrade wood and acquire nutrients. Moreover, there is a scarcity of research on the precise progression of the CMF (chelator-mediated Fenton) stage during brown rot decay in different fungal species. Recent studies have shed light on the formation of CMF constituents in Gloeophyllum trabeum and their interaction with lytic polysaccharide monooxygenase (LPMO), a type of carbohydrate-active enzyme [57]. Interestingly, Umezewa et al. [58] investigated gene expression patterns in Gloeophyllum trabeum exposed to different lignocellulosic media, such as cellulose and untreated wood. It was observed that the expression levels of genes varied depending on the growth substrate. Furthermore, specific gene expression pathways must be active to facilitate the CMF reaction in this particular fungal species [58]. SorCA-modified wood, being a distinct growth substrate for fungi, could potentially impact gene expression in the tested Basidiomycetes, impeding the non-enzymatic stage of brown rot decay. It should be also considered that both citric acid and sorbitol, as well as citric acid- and sorbitol-based compounds are highly effective chelating agents [59, 60]. Consequently, they may compete with oxalate (also a potent chelating agent) for iron at the non-enzymatic decay stage [35]. In theory, the chelation of iron by impregnation chemicals or their esters could potentially stimulate excessive production of oxalate by fungi. The impact of this phenomenon on the decay rate cannot be definitively determined with our current understanding. However, a recent study [61] demonstrated the binding of oxalate to β-glucan, a component of the fungal extracellular matrix, and revealed the formation of a hydrogel with varying viscosity. The study further revealed that the concentration of oxalate affects the diffusion of fungal metabolites with higher molecular weights. If an excessive amount of oxalate is formed, e.g., when it competes with impregnation chemicals and their esters, it could potentially impede decay.
The present study emphasizes the dynamic nature of wood MC, which is influenced by processes such as adsorption, diffusion, capillary condensation and desorption [62]. Impregnation chemicals deposited in the cell wall matrix cause bulking and reduce the space available for water molecules. Thus, a decrease in liquid and vapor water uptake was observed in this study, which delayed or even inhibited decay. Moreover, the SorCA treatment might have changed the glass transition of hemicelluloses, inhibiting the diffusion of ions to the cell wall at the exposure conditions [23].
Conclusions
In this study, the effect of a sorbitol and citric acid (SorCA) modification on the biological durability of Scots pine sapwood specimens exposed to brown rot (C. puteana, R. placenta) and white rot (T. versicolor) fungi according to the EN 113-2 standard [16] was investigated. In general, only the highest treatment level (WPG = 57%) provided a decay protection corresponding to DC 1 [15]. It confirmed the findings of the previous studies stating that high SorCA loadings are required for a full decay protection. The observed differences in the correlation between WPG and ML caused by C. puteana and R. placenta were attributed to variations in the enzymatic decay mechanisms plus the enzymes themselves and secondary metabolites produced, to changes in gene expression or to fluctuations in oxalate concentration. The moisture dynamics investigation revealed that increasing treatment levels (higher WPG values) led to decreased liquid and water vapor uptake of wood, resulting in lower MLs upon fungal exposure. This increased decay protection of SorCA-modified wood might be explained by the combined effect of CWB and cross-linking of SorCA polyesters within the wood matrix. These processes ultimately reduce the spatial availability for water, inhibiting fungal degradation, even if fungal hyphae successfully colonize the cell lumina. We discuss these phenomena in terms of protection against decay in modified wood, based on recent literature studies.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- SorCA:
-
Sorbitol and citric acid
- NIBIO:
-
The Norwegian Institute of Bioeconomy Research
- CA:
-
Citric acid
- WPG:
-
Weight percent gain
- DC:
-
Durability class
- MC:
-
Moisture content
- FSP:
-
Fiber saturation point
- RH:
-
Relative humidity
- ML:
-
Mass loss
- CWU:
-
Capillary water uptake
- CWB:
-
Cell wall bulking
- DVS:
-
Dynamic vapor sorption
- CMF:
-
Chelator-mediated Fenton
- LPMO:
-
Lytic polysaccharide monooxygenase
References
The Biocidal Products Regulation (BPR, Regulation (EU) 528/2012) (2012) European Chemicals Agency (ECHA), Helsinki
Hill C (2006) Wood modification: chemical, thermal and other processes. John Wiley & Sons Ltd, Chichester
Hill CAS, Jones D (1996) The dimensional stabilisation of corsican pine sapwood by reaction with carboxylic acid anhydrides. The effect of chain length. Holzforschung 50:457–462. https://doi.org/10.1515/hfsg.1996.50.5.457
Xiao Z, Xie Y, Militz H, Mai C (2010) Effects of modification with glutaraldehyde on the mechanical properties of wood. Holzforschung 64:475–482
Emmerich L, Altgen M, Rautkari L, Militz H (2020) Sorption behavior and hydroxyl accessibility of wood treated with different cyclic N-methylol compounds. J Mater Sci 55:16561–16575. https://doi.org/10.1007/s10853-020-05224-y
Himmel S, Mai C (2015) Effects of acetylation and formalization on the dynamic water vapor sorption behavior of wood. Holzforschung 69:633–643. https://doi.org/10.1515/hf-2014-0161
Kurkowiak K, Emmerich L, Militz H (2021) Sorption behavior and swelling of citric acid and sorbitol (SorCA) treated wood ABC. Holzforschung 75:1136. https://doi.org/10.1515/hf-2021-0068
Larnøy E, Karaca A, Gobakken LR, Hill CAS (2018) Polyesterification of wood using sorbitol and citric acid under aqueous conditions. Int Wood Prod J 9:66–73
Treu A, Larnøy E, Biørnstad J (2022) CIOL®-protection of wood—an update. Bled, Slovenia, p 9
Kurkowiak K, Hentges D, Dumarçay S, Gérardin P, Militz H (2022) Understanding the mode of action of sorbitol and citric acid (SorCA) in wood. Wood Mater Sci Eng 18:67–75. https://doi.org/10.1080/17480272.2022.2125340
Alfredsen G, Larnøy E, Beck G, Biørnstad J, Ross L, Hill C, Treu A (2020) A summary of decay performance with citric acid and sorbitol modification. In: The International Research Group on Wood Protection. p IRG/WP/20–40898
Beck G (2020) Leachability and decay resistance of wood polyesterified with sorbitol and citric acid. Forests 11:650
Mubarok M, Militz H, Dumarçay S, Gérardin P (2020) Beech wood modification based on in situ esterification with sorbitol and citric acid. Wood Sci Technol 54:479–502
Bravery AF (1978) A miniaturised wood-block test for the rapid evaluation of wood preservative fungicides. Int Res Group Wood Preserv IRGWP78–2113
EN 350 (2016). Durability of wood and wood-based products—testing and classification of the durability to biological agents of wood and wood-based materials, European Committee for Standardization
EN 113–2 (2021). Durability of wood and wood-based products - Test method against wood destroying basidiomycetes—part 2: assessment of biocidal efficacy of wood preservatives, European Committee for Standardization
Brischke C, Grünwald LK, Bollmus S (2020) Effect of size and shape of specimens on the mass loss caused by Coniophora puteana in wood durability tests. Eur J Wood Wood Prod 78:811–819. https://doi.org/10.1007/s00107-020-01559-0
Goodell B, Winandy JE, Morrell JJ (2020) Fungal degradation of wood: emerging data, new insights and changing perceptions. Coatings 10:1210. https://doi.org/10.3390/coatings10121210
Ibach RE, Rowell RM (2000) Improvements in decay resistance based on moisture exclusion. Mol Cryst Liq Cryst Sci Technol Sect Mol Cryst Liq Cryst 353:23–33. https://doi.org/10.1080/10587250008025645
Brischke C, Bayerbach R (2006) Decay-influencing factors: a basis for service life prediction of wood and wood-based products. Wood Mater Sci Eng 1:91–107
Engelund ET, Thygesen LG, Svensson S, Hill CAS (2013) A critical discussion of the physics of wood–water interactions. Wood Sci Technol 47:141–161. https://doi.org/10.1007/s00226-012-0514-7
Wälchli O (1980) Der echte Hausschwamm—Erfahrungen fiber Ursachen und Wirkungen seines Auftretens. Holz als Roh- Werkst 38:169–174
Jakes JE, Zelinka SL, Hunt CG, Ciesielski P, Frihart CR, Yelle D, Passarini L, Gleber SC, Vine D, Vogt S (2020) Measurement of moisture-dependent ion diffusion constants in wood cell wall layers using time-lapse micro X-ray fluorescence microscopy. Sci Rep 10:9919. https://doi.org/10.1038/s41598-020-66916-8
Salmén L, Larsson PA (2018) On the origin of sorption hysteresis in cellulosic materials. Carbohydr Polym 182:15–20. https://doi.org/10.1016/j.carbpol.2017.11.005
Thybring EE (2017) Water relations in untreated and modified wood under brown-rot and white-rot decay. Int Biodeterior Biodegrad 118:134–142. https://doi.org/10.1016/j.ibiod.2017.01.034
Ringman R, Pilgård A, Brischke C, Richter K (2014) Mode of action of brown rot decay resistance in modified wood: a review. Holzforschung 68:239–246
Alfredsen G, Ringman R, Pilgård A, Fossdal CG (2015) New insight regarding mode of action of brown rot decay of modified wood based on DNA and gene expression studies: a review. Int Wood Prod J 6:5–7. https://doi.org/10.1179/2042645314Y.0000000085
Thybring EE (2013) The decay resistance of modified wood influenced by moisture exclusion and swelling reduction. Int Biodeterior Biodegrad 82:87–95. https://doi.org/10.1016/j.ibiod.2013.02.004
Hunt CG, Zelinka SL, Frihart CR, Lorenz L, Yelle D, Gleber SC, Vogt S, Jakes JE (2018) Acetylation increases relative humidity threshold for ion transport in wood cell walls—a means to understanding decay resistance. Int Biodeterior Biodegrad 133:230–237. https://doi.org/10.1016/j.ibiod.2018.06.014
EN 84 (2020). Wood preservatives - Accelerated ageing of treated wood prior to biological testing - Leaching procedure, European Committee for Standardization
Meyer-Veltrup L, Brischke C, Alfredsen G, Humar M, Flæte PO, Isaksson T, Larsson Brelid P, Westin M, Jermer J (2017) The combined effect of wetting ability and durability on outdoor performance of wood—development and verification of a new prediction approach. Wood Sci Technol 51:615–637
CEN/ TS 16818 (2018). Durability of wood and wood-based products - Moisture dynamics of wood and wood-based products, European Committee for Standardization
Thomas R, Fang X, Ranathunge K, Anderson TR, Peterson CA, Bernards MA (2007) Soybean root suberin: Anatomical distribution, chemical composition, and relationship to partial resistance to Phytophthora sojae. Plant Physiol 144:299–311. https://doi.org/10.1104/pp.106.091090
Fich EA, Segerson NA, Rose JKC (2016) The plant polyester cutin: biosynthesis, structure, and biological roles. Annu Rev Plant Biol 67:207–233. https://doi.org/10.1146/annurev-arplant-043015-111929
Arantes V, Jellison J, Goodell B (2012) Peculiarities of brown-rot fungi and biochemical Fenton reaction with regard to their potential as a model for bioprocessing biomass. Appl Microbiol Biotechnol 94:323–338. https://doi.org/10.1007/s00253-012-3954-y
Lee KH, Wi SG, Singh AP, Kim YS (2004) Micromorphological characteristics of decayed wood and laccase produced by the brown-rot fungus Coniophora puteana. J Wood Sci 50:281–284. https://doi.org/10.1007/s10086-003-0558-2
Wei D, Houtman CJ, Kapich AN, Hunt CG, Cullen D, Hammel KE (2010) Laccase and its role in production of extracellular reactive oxygen species during wood decay by the brown rot basidiomycete Postia placenta. Appl Environ Microbiol 76:2091–2097. https://doi.org/10.1128/AEM.02929-09
Riley R, Salamov AA, Brown DW, Nagy LG, Floudas D, Held BW, Levasseur A, Lombard V, Morin E, Otillar R, Lindquist EA, Sun H, LaButti KM, Schmutz J, Jabbour D, Luo H, Baker SE, Pisabarro AG, Walton JD, Blanchette RA, Henrissat B, Martin F, Cullen D, Hibbett DS, Grigoriev IV (2014) Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi. Proc Natl Acad Sci 111:9923–9928. https://doi.org/10.1073/pnas.1400592111
Verma P, Dyckmans J, Militz H, Mai C (2008) Determination of fungal activity in modified wood by means of micro-calorimetry and determination of total esterase activity. Appl Microbiol Biotechnol 80:125. https://doi.org/10.1007/s00253-008-1525-z
Ringman R (2016) Biochemical mechanism of brown rot decay: A study on the mode of action of modified wood. Doctoral Thesis, Technical University of Munich, Munich, Germany
Beck G, Thybring EE, Thygesen LG (2018) Brown-rot fungal degradation and de-acetylation of acetylated wood. Int Biodeterior Biodegrad 135:62–70. https://doi.org/10.1016/j.ibiod.2018.09.009
Krain A, Siupka P (2021) Fungal guttation, a source of bioactive compounds, and its ecological role—a review. Biomolecules 11:1270. https://doi.org/10.3390/biom11091270
Acker JV, Windt ID, Li W, Bulcke JV den (2014) Critical parameters on moisture dynamics in relation to time of wetness as factor in service life prediction, In: The International Research Group on Wood Protection. IRG/WP 20–40898
Emmerich L, Ehrmann A, Brischke C, Militz H (2021) Comparative studies on the durability and moisture performance of wood modified with cyclic N-methylol and N-methyl compounds. Wood Sci Technol 55:1531–1554. https://doi.org/10.1007/s00226-021-01303-8
Emmerich L, Brischke C, Sievert M, Schulz MS, Jaeger AC, Beulshausen A, Humar M (2020) Predicting the outdoor moisture performance of wood based on laboratory indicators. Forests 11:1001. https://doi.org/10.3390/f11091001
Carlquist S (2013) Comparative wood anatomy: systematic, ecological, and evolutionary aspects of dicotyledon wood. Springer Science & Business Media, Berlin
Brischke C, Meyer L, Hesse C, Van Acker J, De Windt I, Van den Bulcke J, Conti E, Humar M, Viitanen H, Kutnik M, Malassenet L (2014) Moisture dynamics of wood and wood-based products - Results from an inter-laboratory test. In: The International Research Group on Wood Protection. IRG/WP 14–20539
Meyer L, Brischke C (2015) Fungal decay at different moisture levels of selected European-grown wood species. Int Biodeterior Biodegrad 103:23–29. https://doi.org/10.1016/j.ibiod.2015.04.009
Biziks V, Bicke S, Koch G, Militz H (2020) Effect of phenol-formaldehyde (PF) resin oligomer size on the decay resistance of beech wood. Holzforschung 75:574–583. https://doi.org/10.1515/hf-2020-0020
Papadopoulos AN, Hill CAS (2002) The biological effectiveness of wood modified with linear chain carboxylic acid anhydrides against Coniophora puteana. Holz Als Roh- Werkst 60:329–332. https://doi.org/10.1007/s00107-002-0327-8
Kurkowiak K, Mayer AK, Emmerich L, Militz H (2022) Investigations of the chemical distribution in sorbitol and citric acid (SorCA) treated wood—development of a quality control method on the basis of electromagnetic radiation. Forests 13:151. https://doi.org/10.3390/f13020151
Lesar B, Humar M (2011) Use of wax emulsions for improvement of wood durability and sorption properties. Eur J Wood Wood Prod 69:231–238. https://doi.org/10.1007/s00107-010-0425-y
Brischke C, Melcher E (2015) Performance of wax-impregnated timber out of ground contact: results from long-term field testing. Wood Sci Technol 49:189–204. https://doi.org/10.1007/s00226-014-0692-6
EN 335 (2013). Durability of wood and wood-based products - Use classes: definitions, application to solid wood and wood-based products, European Committee for Standardization
Stienen T, Schmidt O, Huckfeldt T (2014) Wood decay by indoor basidiomycetes at different moisture and temperature. Holzforschung 68:9–15. https://doi.org/10.1515/hf-2013-0065
Brischke C, Soetbeer A, Meyer-Veltrup L (2017) The minimum moisture threshold for wood decay by basidiomycetes revisited. A review and modified pile experiments with Norway spruce and European beech decayed by Coniophora puteana and Trametes versicolor. Holzforschung 71:893–903. https://doi.org/10.1515/hf-2017-0051
Zhu Y, Plaza N, Kojima Y, Yoshida M, Zhang J, Jellison J, Pingali SV, O’Neill H, Goodell B (2020) Nanostructural analysis of enzymatic and non-enzymatic brown rot fungal deconstruction of the lignocellulose cell wall. Front Microbiol 11:1389. https://doi.org/10.3389/fmicb.2020.01389
Umezawa K, Niikura M, Kojima Y, Goodell B, Yoshida M (2020) Transcriptome analysis of the brown rot fungus Gloeophyllum trabeum during lignocellulose degradation. PLoS ONE 15:e0243984. https://doi.org/10.1371/journal.pone.0243984
Cigala RM, Crea F, De Stefano C, Irto A, Milea D, Sammartano S (2020) Thermodynamic behavior of polyalcohols and speciation studies in the presence of divalent metal cations. J Chem Eng Data 65:2805–2812. https://doi.org/10.1021/acs.jced.0c00120
Karimi Z, Goli M (2021) The effect of chelating agents including potassium tartrate and citrate on the maximum reduction of lead and cadmium during soaking and cooking from some different varieties of rice available in Iran. Food Sci Nutr 9:5112–5118. https://doi.org/10.1002/fsn3.2473
Perez-Gonzalez G, Tompsett GA, Mastelarz K, Timko MT, Goodell B (2023) Interaction of oxalate with β-glucan: Implications for the fungal extracellular matrix, and metabolite transport. iScience 26:106851. https://doi.org/10.1016/j.isci.2023.106851
Brischke C, Alfredsen G (2020) Wood-water relationships and their role for wood susceptibility to fungal decay. Appl Microbiol Biotechnol 104:3781–3795. https://doi.org/10.1007/s00253-020-10479-1
Acknowledgements
Not applicable.
Funding
Open Access funding enabled and organized by Projekt DEAL. The authors acknowledge support by the Open Access Publication Funds of the Göttingen University.
Author information
Authors and Affiliations
Contributions
KK and LE designed the experiments. KK performed the experiments, analyzed the data and wrote the first version of the manuscript. All authors contributed to the interpretation and discussion of the results. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Kurkowiak, K., Emmerich, L. & Militz, H. Biological durability and wood–water interactions of sorbitol and citric acid (SorCA) modified wood. J Wood Sci 69, 34 (2023). https://doi.org/10.1186/s10086-023-02108-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10086-023-02108-y