
Abstract
Cholestatic liver diseases (CLD) are characterized by impaired normal bile flow, culminating in excessive accumulation of toxic bile acids. The majority of patients with CLD ultimately progress to liver cirrhosis and hepatic failure, necessitating liver transplantation due to the lack of effective treatment. Recent investigations have underscored the pivotal role of the gut microbiota-bile acid axis in the progression of hepatic fibrosis via various pathways. The obstruction of bile drainage can induce gut microbiota dysbiosis and disrupt the intestinal mucosal barrier, leading to bacteria translocation. The microbial translocation activates the immune response and promotes liver fibrosis progression. The identification of therapeutic targets for modulating the gut microbiota-bile acid axis represents a promising strategy to ameliorate or perhaps reverse liver fibrosis in CLD. This review focuses on the mechanisms in the gut microbiota-bile acids axis in CLD and highlights potential therapeutic targets, aiming to lay a foundation for innovative treatment approaches.
Similar content being viewed by others

Introduction
The gastrointestinal tract constitutes the largest mucosal surface within human bodies (de Vos et al. 2022). An intricate assembly of diverse cell types of epithelial and endothelial origin and innate and adaptive immunity system is located in the gastrointestinal mucosal barrier, empowering the body to promptly confront challenges from the complex environment (de Vos et al. 2022; Lee et al. 2022; Martel et al. 2022). The portal vein gathers blood from the small and large intestine, spleen, and pancreas and directs it into the liver, establishing a crucial connection between the liver and the intestines (Tilg et al. 2022). In healthy individuals, the liver and its immune system can transform potentially pathogenic or toxic compounds from the gastrointestinal tract into lower toxic metabolites, thereby maintaining the host’s overall health (Guilliams et al. 2022). In turn, liver secretion through the biliary tree into the intestine improves intestinal flora diversity and mucosal barrier integrity, establishing bidirectional crosstalk along the gut-liver axis (Wahlstrom et al. 2016). Disruptions within the gut microbiota-bile acid axis have been recognized for their role in exacerbating the progression of hepatic fibrosis across a spectrum of hepatic pathologies, such as non-alcoholic or alcoholic fatty liver disease, cholestatic liver diseases (CLD), hepatocellular carcinoma, and cholangiocarcinoma (Fuchs and Trauner 2022). This study provides a comprehensive overview of the mechanistic role of the gut microbiota-bile acid in the progression of CLD.
Cholestatic liver diseases emerge due to various intrahepatic and extrahepatic factors impeding bile formation, secretion, or excretion, which results in the accumulation of bile acids (BAs) within the liver and an elevated concentration of BAs in the circulatory system (Chiang and Ferrell 2020). The most common types of CLD include primary biliary cholangitis (PBC), primary sclerosing cholangitis (PSC), biliary atresia, and progressive familial intrahepatic cholestasis (PFIC) (Li et al. 2017). In the early stages of certain CLD, the clinical manifestations might be asymptomatic, with only alkaline phosphatase (ALP) and gamma-glutamyl transpeptidase (GGT) elevating (Shi et al. 2022). Nevertheless, as the disease progresses, hyperbilirubinemia might develop, culminating in liver fibrosis, cirrhosis, and failure, necessitating eventual liver transplantation (Chen et al. 2018; Zeng et al. 2023). Over the past two decades, a global surge in the incidence and prevalence of CLD has been observed. However, the development of efficacious therapeutic strategies to mitigate CLD progression remains a formidable challenge (Zeng et al. 2023).
There is growing attention on the disruption of the gut microbiota-bile acid axis as a culprit of liver fibrosis in CLD (Albillos et al. 2020; Zhang et al. 2022). Studies have demonstrated that under conditions of cholestasis, the impediment to bile drainage impairs the intestinal mucosal barrier, leading to bacteria translocation (Li et al. 2018; Yang et al. 2020). Gut microbiota can also modify the BAs pool, perturbing BAs synthesis and metabolism (Guzior and Quinn 2021). The interplay between the gut microbiota and BAs is bidirectional (Gijbels et al. 2021). Improving the detrimental loop of the gut microbiota-bile acid axis presents an innovative therapeutic strategy for CLD (Li et al. 2017; Laue and Baumann 2022). This review will provide a comprehensive summary of the gut microbiota-bile acid axis in the progression of CLD and present potential therapeutic approaches to ameliorate CLD progression by targeting this axis.
Advances in the regulation of gut microbiota in CLD
The gut microbiota, characterized as a large and diverse community with trillions of microorganisms, colonizes the gastrointestinal tract, spanning from the oral cavity to the rectum (Tilg et al. 2022). In healthy individuals, the gut microbiota’s vast array of microbial species contributes to the resilience and stability of the ecosystem, providing the host with greater resilience to environmental challenges and pathogenic invasion. Gut microbiota dysbiosis is involved in liver fibrosis progression in CLD through multiple mechanisms, such as inflammation, immune activation, and metabolic disturbances (Tilg et al. 2022; Fuchs and Trauner 2022; Li et al. 2018; Wang et al. 2019).
Alteration of gut microbiota in CLD
A series of studies have shown that cholestasis condition led to a reduction in gut microbiota diversity, an increase in potentially pathogenic bacteria, and a decrease in beneficial bacteria (Yang et al. 2020). Lynch et al. (2023) observed a robust positive correlation between the post-menstrual age and the variability in gut microbiome composition in neonates without cholestatic conditions. In contrast, no such correlation was evident in neonates with cholestasis, suggesting an impaired developmental trajectory of the gut microbiome in cholestasis status. Furthermore, they identified that Clostridium perfringens were depleted in stool from cholestatic neonates, leading to impaired deconjugation of BAs (Lynch et al. 2023).
Several studies have observed that the gut microbial composition altered in CLD, such as biliary atresia, primary sclerosing cholangitis (PSC), and primary biliary cholangitis (PBC) (Kummen and Hov 2019; Rager and Zeng 2023). Interestingly, the alterations in gut microbiota in CLD have some similarities, such as the increase in the abundance of Streptococcus and Veillonella genera, and the decrease in the abundance of Faecalibacterium (Thibaut and Bindels 2022; Wang et al. 2022). However, as shown in Table 1, there are also certain variations at the phylum or genus level among different types of CLD (Bajer et al. 2017; Lv et al. 2016; Ostadmohammadi et al. 2021). Biliary atresia, characterized as progressive and fibrotic biliary obstruction, is one of the most devastating hepatobiliary diseases in the neonatal period (Harpavat et al. 2020). In a comparative analysis by Wang et al. (2019) involving 34 patients with biliary atresia and 34 healthy controls (HC), a significant disparity in gut microbiota diversity and composition was observed. Patients with biliary atresia exhibited lower diversity and significant structural segregation compared to HC. At the phylum level, there was an increase in the abundance of Proteobacteria and a reduction in Bacteroidetes in biliary atresia. At the genus level, some potential bacteria, such as Streptococcus and Klebsiella, thrived in biliary atresia, while beneficial bacteria, such as Bifidobacterium and several butyrate-producing bacteria, diminished. In addition, the abundance ratio of Streptococcus/Bacteroides displayed great promise for diagnosing biliary atresia. Song et al. (2021) found Streptococcus, Klebsiella, Veillonella, and Enterococcus were dominant bacteria in biliary atresia. Klebsiella and Veillonella were closely associated with elevated liver enzymes (p < 0.05), while Enterococcus positively correlated with lithocholic acid derivatives (p < 0.05). Yang et al. (2022) reported that post-Kasai jaundice clearance was associated with Campylobacter and Rikenellaceae (p < 0.05). This implies that the alteration of gut microbiota can serve as a diagnostic tool for biliary atresia and a prognostic indicator for predicting long-term outcomes.
So far, numerous high-quality studies have identified alterations in gut microbiota in PSC and PBC (Abe et al. 2018; Lemoinne et al. 2019). Compared to PBC, PSC shows a strong association with inflammatory bowel disease (IBD) and is considered more representative of typical gut-liver axis disorder (Kummen and Hov 2019). Although there might be variations in the specific microbial change observed across different studies on PSC, they consistently reveal distinctions in gut microbial composition between PSC/PSC-IBD patients and HC or IBD patients. These distinctions include the following aspects: (a) a reduction in bacterial diversity and diminished bacterial richness; (b) an enrichment of pathogenic bacterial genera in PSC patients, such as Streptococcus, Veillonella, and Enterococcus. For instance, Kummen et al. (2017) found a 4.8-fold increase of Veillonella in PSC compared to HC, and it could be applied as a promising predictive marker for diagnosing PSC. Sabino et al. (2016) demonstrated a positive correlation between Enterococcus and elevated levels of alkaline phosphatase (ALP) (p = 0.048), a clinical marker for cholestasis; (c) the presence of IBD has a negligible effect on the composition of gut microbiota (Sabino et al. 2016). In addition, Tang et al. (2018) observed a significant reduction in gut microbiota diversity in PBC compared to HC. Importantly, the alterations in gut microbiota can distinguish PBC from HC (area under curve = 0.86, 0.84 in exploration and validation area, respectively). Subsequent research revealed that treatment with ursodeoxycholic acid (UDCA) in PBC patients partially alleviated the dysbiosis of gut microbiota, suggesting that gut microbiota could become a potential therapeutic target for PBC (Tang et al. 2018).
In summary, it is reasonable to infer significant alternations in gut microbiota in CLD, and the specificity of alterations in gut microbiota might become a new diagnostic tool for CLD (Tilg et al. 2022). Besides, the intervention of gut microbiota might become a potential therapeutic target for CLD (Guzior and Quinn 2021). However, there are still some problems to be solved. For example, despite the apparent overlapping microbial features in the microbiomes of patients with CLD, suggesting certain similarities in CLD, such as changes in Streptococcus and Veillonella genera in PSC/PBC/biliary atresia, the etiology of these similar microbial changes remains unclear. Therefore, further investigation is required to explore the role of gut microbiota in the pathogenesis of CLD.
The effect of gut microbiota dysbiosis on liver fibrosis in CLD
Despite alterations in gut microbiota in CLD, the specific mechanisms of how gut microbiota dysbiosis promotes liver fibrosis are still under exploration. Zhou et al. (2023) confirmed that the depletion of gut microbiota exacerbated cholestatic liver injury in mice under bile duct ligation (BDL). The elevated plasma ALT and ALP levels are associated with diminished gut microbiota diversity and increased Gram-negative bacteria, suggesting the significant role of gut microbiota in the progression of CLD (Schneider et al. 2021). Gut microbiota dysbiosis promotes liver fibrosis progression through multiple mechanisms, including disturbances in the intestinal mucosal barrier, immune response, and metabolic disorders (Tilg et al. 2022; Zhang et al. 2022). Intestinal mucosal barrier dysfunction and increased intestinal permeability provided channels for the entry of gut microbiota and metabolites into the liver (Tilg et al. 2022; Qi et al. 2020). Subsequently, the gut microbiota and their metabolites can act as messages to activate hepatic inflammatory signaling pathways, leading to liver fibrosis progression (Qin et al. 2014).
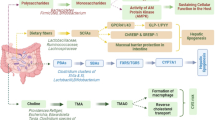
The intestinal mucosal barrier serves as the primary defensive interface against the translocation of luminal contents into the hepatic circulation (Fig. 1). It mainly consists of gut microbiota, intestinal mucus layer, and intestinal epithelial cells (Chopyk and Grakoui 2020). The integrity of the intestinal mucosal barrier is crucial to preventing microbial translocation (Schnabl and Brenner 2014). Gut microbiota plays a vital role in the intestinal mucosal barrier function through various mechanisms (Zhou et al. 2023; Schnabl and Brenner 2014). Firstly, gut microbiota dysbiosis disrupts tight junctions between intestinal epithelial cells, inhibits mucus production, and reduces antimicrobial peptide release (Hiippala et al. 2018; Wells et al. 2017). Tight junctions are integral components between adjacent intestinal epithelial cells, controlling the transport across the epithelium and intestinal epithelial cell permeability (Tilg et al. 2022; Albillos et al. 2020; Turner 2009). Sorriba et al. (2019) have reported that, in comparison to healthy controls (HC), there is a reduction in the ileal epithelial mucosal thickness, loss of goblet cells, bacterial overgrowth, and increased vascular endothelial permeability, all of which contribute to an impaired intestinal epithelial barrier. Once the intestinal mucosal barrier is disrupted, gut bacteria and pathogen-associated molecular patterns (PAMPs) such as lipopolysaccharide (LPS), lipoteichoic acid (LTA), peptidoglycans, endotoxins, can penetrate the intestinal mucosal barrier and enter the liver through the portal vein, inducing innate immune responses activated by Toll-like receptors (TLRs) and nucleotide-binding oligomerization domain-like receptors (NLRs), as well as adaptive immune responses mediated by T cells and B cells, promoting liver fibrosis progression (Wang et al. 2022; Mridha et al. 2017; Vaishnava et al. 2011). The TLR family consists of transmembrane proteins in hepatic cells that recognize gut microbiota and their metabolites, activating the innate immune system. Among them, TLR4 has been studied extensively, and PAMPs such as LPS can bind to TLR4, activating the NF-κB signaling pathway, upregulating inflammatory factors such as TNF-α, IL-1β, and IL-6, inducing hepatic stellate cells (HSCs) activation, and promoting extracellular matrix synthesis, leading to liver fibrosis progression (Mridha et al. 2017). NLRs are a class of pattern recognition receptors that can induce the expression of NOD-like receptor protein 3 (NLRP3) in the cytoplasm. NLRP3 can recognize pathogenic substances from damaged cells and activate the caspase-1 signaling pathway through the innate immune pathway, which can produce IL-1 and IL-18 and stimulate the aviation of hepatic stellate cells (Assimakopoulos and Charonis 2013; Liao et al. 2019; Wree et al. 2014). Furthermore, after entering the portal circulation, gut microbiota and their metabolites can be captured by antigen-presenting cells, activating adaptive immune responses, such as T and B cells, regulating hepatic stellate cell activation, and promoting liver fibrosis progression (Tilg et al. 2022; Hammerich et al. 2014; Novobrantseva et al. 2005).

Abnormal gut microbiota-bile acids in CLD. (A) Abnormal BAs signaling promotes the progression of CLD, and the mechanisms are as follows: (i) the aviation of immune response associated with released numerous pro-inflammatory signals and increased shedding of inflammatory mediators, including cytokines, chemokines, and adhesion molecules, leading to systematic inflammation, (ii) the aviation of hepatic stellate cells (HSCs) leading to extracellular matrix (ECM) deposition, endothelial dysfunction of liver sinusoidal epithelial cells (LSECs), and fibrogenesis formation, (iii) Induction of hepatocytes death through multiple death pathways, including apoptosis, necrosis, and pyroptosis, (iv) gut bacteria and pathogen-associated molecular patterns (PAMPs) enter the liver through the portal vein and promote immune responses, (v) the disruption of tight junctions between cholangiocytes leading to inflammatory and fibrotic response, and cholangiocytes proliferation and senescence. (B) Bacterial translocation participates in liver fibrosis progression in CLD, comprising (i) gut microbiota dysbiosis, (ii) reduced excretion of mucosal thickness and antimicrobial peptide, (iii) disrupted tight junctions between intestinal epithelial cells, (iv) vascular endothelial permeability, (v) the decreasing synthesis of short-chain fatty acids (SCFAs), which enabled the disruption of intestinal mucosal barrier and translocation of bacteria and pathogen-associated molecular patterns (PAMPs) into the liver through portal vein. BA, bile acids; DCs, dendritic cells; TJ, tight junction; KC, Kupffer cells; HSCs, hepatic stellate cells; PAMPs, pathogen-associated molecular patterns; LPS, lipopolysaccharide; SCFAs, short-chain fatty acids;
Dysbiosis of gut microbiota can also disrupt microbial metabolism, resulting in decreased short-chain fatty acids (SCFAs) synthesis. Jee et al. (2022) found that feeding with butyrate salts in BDL mice could alleviate the progression of liver fibrosis and improve the survival of biliary epithelial cells. SSCFAs, predominantly derived from the fermentation of dietary fibers by the gut microbiota, include acetate, butyrate, propionate, and formate, which collectively account for 90-95% of the SCFA pool (Rios-Covian et al. 2016). SCFAs not only act as an energy source for intestinal epithelial cells, but also regulate immune cells, maintaining the integrity of the intestinal epithelial mucosal barrier (Kim 2018). Acetate can bind to surface receptors of dendritic cells (DCs), inducing the release of infection-free globules (IgA) through B cells aviation. Propionate and butyrate can inhibit dendritic cells (DCs) CD40 expression, reducing IL-6 and IL-12 expression (Nastasi et al. 2015). Butyrate can also regulate Treg cell differentiation through dendritic cells to modulate immune responses (Singh et al. 2014). Treg cells are a CD4 + T cell subset that serves as the only negative regulator of immune cells in the body, inhibiting the proliferation of Th cells (such as Th1, Th2, and Th17 cells) and thereby suppressing inflammatory responses (Dong et al. 2015). In conclusion, the dysbiosis of gut microbiota promotes the progression of liver fibrosis through multiple pathways, such as disruption of the intestinal mucosal barrier, immune response, and decreased synthesis of SCFAs. Maintaining gut microbiota homeostasis plays a vital role in improving the progression of liver fibrosis (Zhang et al. 2022; Juanola et al. 2021).
Role of bile acids in the CLD progress
BAs homeostasis
BAs are primarily synthesized from cholesterol, with the synthesis pathways involving two main enzymatic processes: the classical pathway, predominantly initiated by cholesterol 7α-hydroxylase (CYP7A1) (responsible for the majority of bile acid synthesis), and the alternative pathway, mediated by sterol 27-hydrolase (CYP27A1) (accounting for approximately 10% of total bile acid synthesis) (Zeng et al. 2023). These pathways yield two free primary BAs: cholic acid (CA) and chenodeoxycholic acid (CDCA) (Chiang 2004). Irrespective of the initial pathway, the last step in BA synthesis involves the conjugation of primary BAs with glycine or taurine, which is facilitated by bile acid coenzyme A amino acid N-acetyltransferase (BAAT). This enzyme-catalyzed reaction maintains the amphipathic structure and allows BAs to be impermeable through the membrane (Dawson et al. 2009; Monte et al. 2009). Subsequently, the majority of BAs are actively transported into bile canaliculi via the bile salt export pump (BSEP). In the terminal ileum, primary conjugated BAs undergo microbial enzymatic conversion and are converted into secondary BAs, such as deoxycholic acid (DCA), lithocholic acid (LCA), and ursodeoxycholic acid (UDCA). The majority of conjugated BAs are actively taken up into enterocytes via the apical sodium-dependent BA transporter (ASBT), transported through the basolateral membrane by the ileal BA-binding protein (IBABP), and then enter the portal vein via the organic solute transporter-α/β (OSTα/β). Unconjugated BAs can be taken up by passive diffusion through the intestine. In the liver, most BAs are reabsorbed into hepatocytes via sodium taurocholate co-transporting polypeptide (NTCP). A minor fraction of unconjugated BAs returned to hepatocytes through organic anion-transporting polypeptides (OATPs) (Zhang et al. 2022). Approximately 95% of BAs are reabsorbed into the liver by the gut-liver axis, while the remaining unabsorbed portion is excreted with feces and urine (Fig. 2) (Guzior and Quinn 2021).

Bile acids transport and signaling along the gut-liver axis. (A) Hepatocellular BA homeostasis. Most conjugated BAs are reabsorbed via sodium taurocholate co-transporting polypeptide (NTCP) and unconjugated BAs through organic anion-transporting polypeptides (OATPs). BAs are taken up from the portal vein and synthesized by two main enzymatic processes: the classical pathway, predominantly initiated by cholesterol 7α-hydroxylase (CYP7A1) (responsible for the majority of bile acid synthesis), and the alternative pathway, mediated by sterol 27-hydrolase (CYP27A1) (not shown here), which are excreted into bile canaliculi via the bile salt export pump (BSEP). In addition, the FXR signaling pathway (activated by BAs) can inhibit CYP7A1 and CYP8B1 gene expression, suppress NTCP expression, and induce BSEP expression, thus protecting hepatocytes from BAs toxicity. BAs synthesis is also inhibited by fibroblast growth factor 19 (FGF19), which is transported back to the liver via fibroblast growth factor receptor 4 (FGFR4) and suppresses the activity of CYP7A1. (B) In the terminal ileum, primary conjugated BAs are actively taken up into enterocytes via the apical sodium-dependent BA transporter (ASBT), transported through the basolateral membrane by the ileal bile acid-binding protein (IBABP), and enter the portal vein via the organic solute transporter-α/β (OSTα/β). Unconjugated BAs can be taken up by passive diffusion through the intestine. Intestinal BAs enhance the expression of the FXR/FGF19 signaling pathway, which is transported back to the liver and suppresses BA synthesis. In addition, the aviation of FXR signaling downregulates ASBT expression and induces OSTα/β expression, protecting enterocytes from BAs toxicity. NTCP, sodium taurocholate co-transporting polypeptide; OATPs, organic anion-transporting polypeptides; FGF19, fibroblast growth factor 19; FGFR4, fibroblast growth factor receptor 4; ASBT, apical sodium-dependent BA transporter; IBABP, ileal bile acid-binding protein; CYP7A1, cholesterol 7α-hydroxylase
The synthesis and transport of BAs are also regulated by a sophisticated network of hormone-like signaling (Chiang 2009; Lu et al. 2000). BAs can bind and activate members of the cell surface and nuclear receptors family, collectively referred to as BA-activated receptors (Fiorucci et al. 2020). Key receptors include the Farnesoid X receptor (FXR), Takeda G protein-coupled receptor 5 (TGR5), Liver X receptor (LXR), and sphingosine-1-phosphate receptor 2 (S1PR2) (Wan and Sheng 2018). Among these receptors, FXR is the most crucial receptor that regulates BA homeostasis (Yan et al. 2021). FXR signaling regulates BAs synthesis through two principal pathways (Fig. 2). Firstly, the FXR signaling pathway regulates the gene expression of crucial enzymes involved in BAs synthesis (Goodwin et al. 2000). FXR aviation promotes the expression of the target gene SHP, leading to the downregulation of CYP7A1 and CYP8B1 gene expression (Xiang et al. 2021). Notably, loss of SHP in mice fed BAs showed that inhibition of CYP7A1 gene expression was retained, indicating the presence of compensatory repression pathways of BA signaling independent of SHP (Kerr et al. 2002; Wang et al. 2002). Moreover, intestinal BAs aviation enhances the expression of FGF15/19, which is transported back to the liver via the portal vein system, activating the JNK signaling pathway by binding with fibroblast growth factor receptor 4 (FGFR4), thereby inhibiting CYP7A1 expression (Li et al. 2017). Additionally, studies revealed that the expression of FGF19 in the liver tissue of patients with biliary atresia or PBC was significantly higher than FGF19 in serum, suggesting that intrahepatic auto-/paracrine FGF19 feedback signaling might exist (Al-Khaifi et al. 2018; Nyholm et al. 2023).
Secondly, FXR signaling also plays a pivotal role in regulating bile acid transporters. Experimental studies demonstrated that the aviation of the FXR/SHP pathway downregulates ASBT expression in the ileum (Chen et al. 2003; Neimark et al. 2004). The subsequent suppression of ASBT has been associated with a reduction in the expression of pro-fibrogenic genes, thereby attenuating the progression of liver fibrosis (Baghdasaryan et al. 2016; Miethke et al. 2016). FXR aviation also induced OSTα/β expression, which protected enterocytes from BA toxicity. Elevated OSTα/β expression has been observed in BDL mice as well as patients with PBC. The upregulation of OSTα/β might serve as a baolateral “pressure relieve valve” for consistently elevated intracellular BA concentration (Boyer et al. 2006; Gulamhusein and Hirschfield 2020). In addition, the FXR/SHP signaling pathway represses the expression of NTCP in hepatocytes in BDL mice, which protects hepatocytes from BA toxicity (Geier et al. 2005; Robin et al. 2018; Zollner et al. 2002). NTCP inhibitor is promising to become a novel therapy for HBV/HDV and has demonstrated liver protection in cholestatic mice (Bogomolov et al. 2016; Slijepcevic et al. 2018). FXR aviation could induce BSEP expression, facilitating BA secretion into bile canaliculi (Ananthanarayanan et al. 2001; Plass et al. 2002). Inherited BSEP deficiency leads to cholestasis in humans, underscoring the critical role of BSEP in BA synthesis (Fuchs et al. 2017).
Changes in the BAs profile in CLD progress
Based on their concentration, BAs might exhibit either beneficial or cytotoxic effects in hepatic and gastrointestinal diseases (Jansen et al. 2017). BAs at physiological concentrations can not only facilitate the metabolism of fats and cholesterols but also enhance immune response and maintain the balance of gut microbiota (Tilg et al. 2022). However, excessive levels of BAs may trigger inflammatory pathways and kill hepatocytes (Fuchs and Trauner 2022). BAs toxicity might not only be dependent on their concentration but also on their hydrophobicity and conjugation status, with the following increasing order of toxicity: UDCA< CA< CDCA< DCA< LCA (Hegade et al. 2016). BAs with greater hydrophobicity are more prone to penetrate cell membranes, thus disrupting intracellular structures and processes, leading to cellular damage and toxicity (Fuchs and Trauner 2022; Zeng et al. 2023). Hydrophobic BAs have been implicated in liver fibrosis through various pathways (Fig. 2). They can initiate inflammation response by stimulating the production of mediators, such as cytokines, chemokines, and adhesion molecules (Allen et al. 2011). Studies have demonstrated that hydrophobic BAs could induce the hepatocellular expression of adhesion factors and neutrophil chemoattractants, including ICAM1, CXCL1, CXCL2, and IL-17, leading to neutrophil aggregation (Fuchs and Trauner 2022). Additionally, BAs can promote the expression of the transcription factor early growth factor (EGR1), releasing pro-inflammatory signals in hepatocytes (Allen et al. 2010). The activation of EGR1-dependent pro-inflammatory signaling can induce the expression of IL-23 in hepatocytes, which is integral to the infiltration and activation of T helper 17 (Th17) cells (Licata et al. 2013; O’Brien et al. 2013). Hepatic accumulation of BAs converts DCs into an immunogenic myeloid phenotype with an enhanced ability to prime allogeneic and syngeneic T lymphocytes and secretes pro-inflammatory cytokines (Carambia et al. 2014; Rahman and Aloman 2013). Macrophage necrosis plays a pivotal role in liver fibrosis. In cholestatic conditions, conjugated BA glycodeoxycholate (GDCA) upregulates the Z-DNA-binging protein 1 (ZBP1)/phosphorylated-mixed lineage kinase domain-like pseudokinase (p-MLKL) signaling pathway and mediates macrophage necrosis, exacerbating liver fibrosis progression (Yang et al. 2023).
High BAs concentrations can induce cell death through multiple pathways in CLD, including apoptosis, necrosis, and pyroptosis (Jansen et al. 2017; Chen et al. 2001). Firstly, Hydrophobic BAs can activate mitochondrial oxidative stress, leading to alterations in mitochondrial permeability, the release of cytochrome C, and the initiation of a caspase cascade reaction, culminating in hepatocyte apoptosis (Lemasters et al. 1998; Yin and Ding 2003). Additionally, BAs can activate death receptors through an exogenous pathway, facilitating the formation of death receptor signaling complexes, activation of Caspase 8, and translocation of pro-apoptotic proteins to the mitochondria, leading to irreversible cell death (Faubion et al. 1999; Malhi and Kaufman 2011). Secondly, hydrophobic BAs can trigger necrosis through oxidative-stress-induced lipid peroxidation and dissolution of the hepatocellular plasma membrane (Shojaie et al. 2020). Afonso et al. (2016) found a high level of MLKL and receptor-interacting protein kinase (RIPK3) expression in patients diagnosed with PBC, confirming the presence of hepatic necrosis in CLD. Using the BDL animal model, further investigation showed a significantly increased pMLKL/MLKL after three days of ligation, suggesting that hepatocyte necrosis might represent an early event in the pathogenesis of CLD. Thirdly, in recent years, studies have revealed that pyroptosis might play a vital role in the progression of liver fibrosis. Gan et al. (2022; Xiao et al. 2023). Pyroptosis is a recently described programmed cell death with the release of numerous inflammatory cytokines (Gan et al. 2022; Yu et al. 2021). Chen et al. found BAs targeting mitofusin 2 (MFN2) differentially modulated innate immunity in physiological versus cholestatic states. BAs at a physiological concentration can promote mitochondrial fusion and enhance phagocytosis of macrophages, while BAs at high concentrations promote mitochondrial tethering to the endoplasmic reticulum, leading to mitochondrial calcium overload, which activates NLRP3 inflammasome and pyroptosis (Che et al. 2023). Xu et al. further analyzed the mechanisms of BAs-induced pyroptosis. Different from the LPS-induced GSDMD-dependent canonical pyroptosis pathway, BAs mainly aviated caspase-11 and GSDME-mediated non-canonical pyroptosis pathway underlies cholestatic liver damage (Xu et al. 2021).
Compared to elevated serum or hepatocellular BAs concentration, bile leakage from canaliculi and bile duct might play a more significant role in developing cholestatic liver injury (Jansen et al. 2017). High concentrations of BAs can disrupt tight junctions between biliary epithelial cells, resulting in bile leakage in the periductal region and the subsequent activation of inflammatory and fibrotic responses (Zeng et al. 2023). The accumulation of BAs can promote cholangiocyte proliferation and periportal fibrosis. Cholangiocyts proliferation, known as ductular reaction (DR), represents an adaptive response of cholangiocyts (Bertolini et al. 2022; Wang et al. 2017). Cholangiocyts often exhibit a neuroendocrine-like phenotype after DR, expressing numerous anti-apoptotic genes, adhesion molecules, cytokines, chemokines, and growth factors, which promotes the aviation, migration, and proliferation of myofibroblasts (Zeng et al. 2023). In BDL mice, DR can be observed, accompanied by the secretion of growth factors and inflammatory cytokines such as epidermal growth factor (EGF), vascular endothelial growth factor (VEGF), IL-6, and tumor necrosis factor (TNFα) (Munshi et al. 2011). While early DR-induced bile duct injury might regress, persistent BA concentration and inflammatory response can induce infiltration of the mesenchymal cell, promote periductal inflammatory response, induce cholangiocyte apoptosis or senescence, leading to the progression of portal fibrosis and even potential malignant transformation (Meadows et al. 2021; Yokoda and Rodriguez 2020). Consequently, further research is warranted to explore the critical pathways involved in DR in CLD.
BAs can also exert anti-inflammatory pathways via interacting with BA nuclear receptors and inhibiting immune responses (Fuchs and Trauner 2022; Wang et al. 2008). FXR is expressed in various immune cells. The aviation of FXR inhibits Toll-receptor 9 (TLR9)-dependent expression in macrophages and suppresses pro-inflammatory activity (Renga et al. 2013). FXR activation can also downregulate cytoplasmic NLRP3 expression, reducing inflammatory responses (Hao et al. 2017). FXR signaling pathway antagonizes NF-kB target gene expression in dendritic cells, including TNFα and inducible nitric oxide synthase (iNOS) (Wang et al. 2008). In natural killer T cells, FXR aviation suppresses the production of osteopontin, IL-1β, and IFN-γ (Gadaleta et al. 2011).
Takeda G protein-coupled receptor 5 (TGR5) is identified in macrophages as the first G-protein-linked receptor (GPCR) activated by BAs (Keitel et al. 2009). TGR5 exhibits high expression in non-parenchymal liver cells, including hepatic sinusoidal endothelial cells, hepatic stellate cells, and cholangiocytes (Reich et al. 2021). The aviation of TGR5 promotes intracellular bucolic adenosine monophosphate (cAMP) production and protein kinase A (PKA) aviation, thereby inhibiting NF-kB signal transduction and reducing inflammatory response (Guo et al. 2016; Keitel and Häussinger 2018). More importantly, TGR5 seems necessary for biliary tree development (Reich et al. 2016). TGR5 aviation promotes cholangiocyte regeneration, which is crucial for maintaining the integrity of the biliary tree and mitigating BA toxicity through stimulating bicarbonate secretion (Reich et al. 2021; Trauner and Fuchs 2022). TGR5-knockout BDL mice appeared more susceptible to cholestatic liver injury than wild-type mice (Reich et al. 2021; Pean et al. 2013).
The body employs compensatory defense mechanisms to alleviate the toxic effects of bile stasis in hepatic and gastrointestinal cells (Zollner et al. 2006). These mechanisms include adaptive changes in BAs receptors and activation of anti-inflammatory responses. However, these adaptations prove insufficient in countering the sustained liver damage induced by prolonged cholestasis. Moreover, cholestatic status can lead to downregulation and functional impairment of BA receptors and transport systems (Li et al. 2017). Consequently, further research is essential to investigate the pivotal pathways involved in liver fibrosis and bile duct injury, aiming to develop potential therapeutic strategies for mitigating the progression of CLD.
The gut microbiota-bile acid crosstalk in CLD
Bidirectional interactions between gut microbiota and BAs
Regulation of BAs profiles by gut microbiota
The intricate interplay of the gut microbiota encompasses a diverse array of mechanisms that govern the metabolism of BAs. The gut microbes are instrumental in the conversion of primary BAs into secondary BAs through multiple metabolic pathways, such as deconjugation, dehydroxylation, oxidation, isomerization, esterification, and desulfation (Li et al. 2017; Cai and Boyer 2021). These bioactive secondary BAs participate in the enterohepatic circulation, where they modulate the composition of BA pools and contribute to the maintenance of the host’s metabolic homeostasis (Zhang et al. 2022; Li et al. 2021). The initial phase of microbial BAs metabolism hinges on the enzymatic activity of bile salt hydrolase (BSH), which catalyzes the deconjugation reaction, hydrolyzing glycine- and taurine-conjugated BAs into their free forms (Li et al. 2017; Guzior and Quinn 2021). Gut microbiota exhibiting the BSH enzyme are varied and encompass multiple species, including Bacteroides, Clostridium, Lactobacillus, Bifidobacterium, Enterococcus, Ruminococcaceae, and Listeria (Chascsa et al. 2017). Lynch and colleagues reported a substantial reduction in BSH gene abundance and enzyme activity in neonates with bile stasis, leading to the impaired synthesis of unconjugated BAs synthesis and diminished gut microbiota diversity (Lynch et al. 2023). Further downstream, the gut microbiota facilitates unconjugated BAs dehydroxylation, oxidation, and isomerization processes (Guzior and Quinn 2021). Particularly noteworthy is the role of the Bai operon of Clostridium in orchestrating 7α-dehydroxylation, which transforms cholic acid (CA) into deoxycholic acid (DCA) and chenodeoxycholic acid (CDCA) into lithocholic acid (LCA) (Thibaut and Bindels 2022). Additional modifications enacted by bacteria encompass oxidation, epimerization, desulfation, and esterification (Zhang et al. 2022; Guzior and Quinn 2021). For instance, Oxalobaceraceae, Enterobacteriaceae, Clostridiaceae, Escherichia, and Lachnospiraceae contribute to BA oxidation and isomerization (Guzior and Quinn 2021). In parallel, Oxalabacteraceae, Enterobacteriaceae, and Lactobacillus are involved in bile acid esterification, and Clostridiaceae, Stretociccaeae, and Pseudomonadaceae are implicated in BA desulfation (Guzior and Quinn 2021; Jia et al. 2018; Just et al. 2018).
Given the potential for oxidation, epimerization, and dehydroxylation at multiple hydroxyl groups in BAs, the diversity of human bile acids extends beyond 2800 distinct variants (Guzior and Quinn 2021). This diversity exerts a pronounced influence on gut microbiota and the host. Alteration in oxidation and isomerization status modulate BA’s hydrophobicity and toxicity profiles, thereby protecting the hepatocytes and enterocytes from more hydrophobic and hazardous BAs (Guzior and Quinn 2021; Kisiela et al. 2012). In addition to stimulating the conversion from primary BA to secondary forms, the gut microbiota governs BA synthesis by regulating BA synthetic enzymes. Sayin and collaborators unveiled that gut microbiota could downregulate the expression of taurine-β-muri cholate sodium salt and upregulate FGF15 expression through FXR receptor aviation, thereby enhancing hepatic CYP7A1 expression and promoting BAs synthesis (Sayin et al. 2013). Moreover, investigations by Kim et al. confirmed that in the absence of gut microbiota, FXR antagonists remained unmetabolized, underscoring the potential role of the gut microbiota in the regulation of BAs synthesis via the FXR-FGF15/19 signal pathway (Schneider et al. 2021; Kim et al. 2007).
BAs as regulators of gut microbiota
BAs also assume a pivotal role in modulating the gut microbiota composition and upholding the integrity of the intestinal mucosal barrier (Albillos et al. 2020). Compromised integrity of this barrier can establish a conduit for intestinal bacteria and their metabolites to access the liver via the portal vein, thereby contributing to the progression of liver fibrosis (Tilg et al. 2022; Yang et al. 2020). In cholestatic conditions, the depletion of BAs within the intestinal lumen could trigger microbial dysbiosis and disruption of the intestinal epithelial mucosal barrier, culminating in microbial translocation (Li et al. 2017). Nevertheless, oral administration of BAs has demonstrated the capacity to mitigate excessive bacterial proliferation and microbial translocation in mice afflicted with liver cirrhosis (Simbrunner et al. 2021; Verbeke et al. 2015). Under normal physiological conditions, the intestinal mucosal barrier is meticulously regulated by intricate mechanisms that prevent the translocation of bacteria and PAMPs from the mucosal stratum (D’Aldebert et al. 2009). The occurrence of microbial translocation suggests a disruption of these mechanisms, including encompassing the downregulation of antimicrobial peptides, reduction in mucosal thickness, diminished expression of tight junction proteins, and impairment of intestinal vascular carriers (Xiang et al. 2021). Ongoing research underscores the potential of BAs metabolism to impinge upon the integrity of the intestinal mucosal barrier through the following mechanisms: Firstly, the deficiency of intestinal BAs could alter the composition of gut microbiota. BAs can stimulate the proliferation of intestinal flora, such as Enterococcus faecalis, Escherichia coli, and Listeria (Devkota and Chang 2015; Gahan and Hill 2014). Furthermore, previous studies have indicated that the absence of BAs could lead to a significant decline in enzyme activity and quantity of 7α-dehydroxylation, underscoring the indispensable role of BAs in fostering the growth of gut microbial communities (Guzior and Quinn 2021). The second mechanism involves the impairment of the intestinal mucosal barrier. Ubeda et al. discerned a downregulation of the FXR/SHP signaling pathway in the context of cirrhotic rats (Ubeda et al. 2016). Activation of the FXR pathway led to an augmentation in the expression of tight junction-related proteins and antimicrobial peptides, thereby preserving the integrity of the intestinal mucosal barrier and effectively ameliorating the progression of liver fibrosis (Sorribas et al. 2019). Meanwhile, Verbeke et al. observed a significant reduction in the expression of the FXR pathway in the ileum and jejunum in experimental BDL mice, resulting in the alteration of the intestinal mucosal barrier and the consequent translocation of gut microbiota. Conversely, FXR agonists enhanced the integrity of tight junctions and improved the intestinal mucosal barrier (Xiang et al. 2021). A recent investigation illuminated that Tropifexor (an FXR against) could elevate the expression of FGF19 within the ileum in BDL piglets, which enhanced the abundance of bile caid-biotransforming bacteria in the distal ileum, ameliorated intestinal barrier injury and suppressed BDL-induced liver injury, fibrosis, and ductular reaction (Xiao et al. 2021). These studies conveyed the pivotal role of BAs in shaping the gut microbiota and preserving the integrity of the intestinal mucosal barrier (Shi et al. 2022).
The gut microbiota-bile acid axis as a target for CLD treatment
FXR ligands
Current studies have provided substantial evidence that multiple FXR agonists could ameliorate the gut microbiota, suppress BA synthesis and metabolism, and relieve liver fibrosis progression in animal models with cholestasis (Yan et al. 2021; Xiang et al. 2021). Currently, FXR agonists are in clinical trials for treating CLD (Table 2). Obeticholic acid, characterized as a potent FXR receptor activator, has completed phase II and III clinical trials on CLD in multiple studies (Kowdley et al. 2020; Trauner et al. 2019a, b). Remarkably, obeticholic acid exhibits the capability to attenuate serum hepatic enzyme levels and reduce bilirubin and alkaline phosphatase levels (ALP) in patients with PBC with inadequate response to or intolerance to ursodeoxycholic acid, suggesting long-term efficacy and safety in treating CLD. However, this drug has some adverse effects. The preeminent adverse effect was dose-dependent pruritus, thus demanding heightened attention in prospective research (Kowdley et al. 2020; Trauner et al. 2019a, b). Other types of FXR agonists, such as Cilofexor and Tropifexor, have also accomplished phase II clinical trials, which exhibited significant reduction in liver enzymes and gamma-glutamyl transferase (GGT) levels and possessed predictable pharmacokinetics and an acceptable safety-tolerability profile, thereby supporting further clinical exploration for FXR against in the treatment of CLD (Schramm et al. 2022; Trauner et al. 2019a). In addition, FGF19 acted as a downstream molecule after FXR aviation (Johansson et al. 2020). Using FGF19 analog to treat CLD could potently and robustly suppress hydrophobic BA levels and protect hepatocellular and intestinal epithelial from toxicity (Sanyal et al. 2021).
ASBT inhibitors
ASBT inhibitor, characterized by suppressors of the ileal bile acid transport protein, could impede the hepatic-enteric circulation of BA and reduce serum BA concentration (Fuchs and Trauner 2022; Thibaut and Bindels 2022). Several ASBT inhibitor drugs, such as Odevixibat, Maralixibat, and Linerixibat, have been utilized in phase II and phase III clinical trials targeting pediatric CLD (Table 2) (Baumann et al. 2021; Bowlus et al. 2023; Gonzales et al. 2021; Hegade et al. 2017; Levy et al. 2023; Loomes et al. 2022; Thompson et al. 2022). ASBT inhibitors demonstrated the ability to reduce serum bile acid levels and alleviate pruritus markedly and represented a pioneering group of agents that manifest sustained and effective improvement in pediatric cholestasis (Gonzales et al. 2021). The most commonly observed adverse effects were associated with gastrointestinal disease, primarily self-limiting manifestations, such as diarrhea. Although ASBT inhibitors might become a novel therapeutic paradigm for CLD, further clinical studies are needed to observe the safety and efficacy of the drugs and expand the application of ASBT inhibitors in the realm of CLD (Gonzales et al. 2021; Levy et al. 2023).
Norursodeoxycholid acid
Since the 1980s, Norursodeoxycholid acid (UDCA) has been shown to partially alleviate liver injury in PBC (Poupon et al. 1987). Subsequently, several studies have applied UDCA in a wide range of cholestatic disorders, making it a first-line drug for treating CLD (Beuers et al. 2015; Rost et al. 2004). However, long-term follow-up studies of UDCA in CLD have found that while UDCA might relieve the progression of liver cirrhosis and reduce the requirement for liver transplantation in some patients, approximately 40% of CLD patients are insensitive or irresponsive to UDCA treatment. Besides, no clear-cut survival benefit with UDCA in CLD has been displayed (Beuers et al. 2015; Cheung et al. 2019). 24-Norursodeoxycholid acid (norUDCA), a side chain shortened homolog of UDCA, presents several advantages over its parent compound: (a) norUDCA can be passively absorbed through cholangiocytes and undergo “choleheaptic shunting”, promoting HCO3-rich hypercholeresis, thus counteracting the toxic effects of BAs; (b) norUDCA has a stronger anti-inflammatory, antifibrotic and antiproliferative effect, directing regulating immune responses; (c) norUDCA is more hydrophilic and thereby less toxicity to hepatocytes and cholangiocytes (Fuchs and Trauner 2022; Beuers et al. 2015). Fickert et al. (Fickert et al. 2017) conducted a double-blind, randomized, multicenter study to evaluate the safety and efficacy of different doses of oral norUDCA in PSC patients. They found that norUDCA significantly reduced ALP levels in a dose-dependent manner with good safety. Phase III clinical trials are currently undergone to further evaluate the therapeutic efficacy.
Fecal microbial transplantation
Fecal microbial transplantation (FMT) refers to a procedure in which fecal matter, or stool is collected from a healthy donor, processed, and transplanted into the gastrointestinal tract of a recipient, aiming to improve the structure and function of the gut microbiota (Liu et al. 2024). Numerous studies have suggested that FMT might be a promising strategy for ulcerative colitis (Costello et al. 2019; Haifer et al. 2022). In CLD, alternations of gut microbiota are also common, and they can influence BAs metabolism through the gut-live axis. Therefore, FMT might have potential therapeutic effects in the treatment of CLD (Fang et al. 2022; Gerussi et al. 2020). Allegretti et al. (2019) conducted a pilot clinical trial of FMT for treating PSC and found the diversity of gut microbiota increased post-FMT. Additionally, 30% of patients experienced a ≥ 50% decrease in ALP levels with no related adverse events, which suggested that FMT might be safe for treating PSC. However, the safety and efficacy of FMT in treating CLD require further evaluation through larger prospective, multicenter clinical trials.
Conclusion
In summary, the interaction between gut microbiota and BA defies unidirectionality. BAs can mold the gut microbiota; conversely, the gut microbiota can alter BA composition. This bidirectional dynamic, a reciprocal interplay, assumes a pivotal role in CLD. In CLD, the elevated BA concentrations within the liver can incite inflammatory cascades, thereby instigating hepatocyte death and effectuating the progression of liver fibrosis. In addition, impediments of bile flow into the intestines can incite gut microbiota dysbiosis, culminating in the disruption of the intestinal mucosal barrier and microbial translocation. Bacteria translocation can trigger immune response and propel the progression of CLD. Currently, clinical trials are investigating pharmaceutical agents that target the gut microbiota-bile acid axis. However, given the potential side effects and the possible lack of specificity to certain tissues or cell types, there is a clear necessity for further research to evaluate the safety and efficacy of these drugs in clinical practice. Moreover, there is an urgent need for a deeper understanding of the mechanisms of the gut microbiota-bile acid axis in CLD for exploring potential therapeutic targets.
Data availability
All data generated or analyzed during this study are included in this manuscript.
Abbreviations
- CLD:
-
Cholestatic liver diseases
- Bas:
-
Bile acids
- PBC:
-
Primary biliary cholangitis
- PSC:
-
Primary sclerosing cholangitis
- PFIC:
-
Progressive familial intrahepatic cholestasis
- DCs:
-
Dendritic cells
- TGR5:
-
Takeda G protein-coupled receptor 5
- ALP:
-
Alkaline phosphatase
- GGT:
-
Gamma-glutamyl transpeptidase
- HC:
-
Healthy control
- IBD:
-
Inflammatory bowel disease
- TJ:
-
Tight junction
- KC:
-
Kupffer cells
- HSCs:
-
Hepatic stellate cells
- PAMPs:
-
Pathogen-associated molecular patterns
- LPS:
-
Lipopolysaccharide
- LTA:
-
Lipoteichoic acid
- TLR:
-
Toll-like receptor
- NLR:
-
Domain-like receptor
- IgA:
-
Infection-free globule
- BSEP:
-
Bile salt export pump
- SCFAs:
-
Short-chain fatty acids
- NTCP:
-
Sodium taurocholate co-transporting polypeptide
- OATPs:
-
Organic anion-transporting polypeptides
- FGF19:
-
Fibroblast growth factor 19
- FGFR4:
-
Fibroblast growth factor receptor 4
- ASBT:
-
Apical sodium-dependent BA transporter
- IBABP:
-
Ileal bile acid-binding protein
- OSTα/β:
-
Organic solute transporter-α/β
- LXR:
-
Liver X receptor
- S1PR2:
-
Sphingosine-1-phosphate receptor 2
- ZBP1:
-
Z-DNA-binging protein 1
- p-MLKL:
-
Phosphorylated-mixed lineage kinase domain-like pseudokinase
- CYP7A1:
-
Cholesterol 7α-hydroxylase
- GDCA:
-
Conjugated BA glycodeoxycholate
- EGF:
-
Epidermal growth factor
- VEGF:
-
Vascular endothelial growth factor
- TNFα:
-
Tumor necrosis factor
- DR:
-
Ductular reaction
- GPCR:
-
G-protein-linked receptor
- cAMP:
-
Adenosine monophosphate
- NLRP3:
-
NOD-like receptor protein 3
- DCA:
-
Deoxycholic acid
- LCA:
-
Lithocholic acid
- BSH:
-
Bile salt hydrolase
- CDCA:
-
Chenodeoxycholic acid
- NUDCA:
-
Norursodeoxycholid acid
- norUDCA:
-
24-Norursodeoxycholid acid
- FMT:
-
Fecal microbial transplantation
References
Abe K, Takahashi A, Fujita M, Imaizumi H, Hayashi M, Okai K, et al. Dysbiosis of oral microbiota and its association with salivary immunological biomarkers in autoimmune liver disease. PLoS ONE. 2018;13:e0198757.
Afonso MB, Rodrigues PM, Simao AL, Ofengeim D, Carvalho T, Amaral JD, et al. Activation of necroptosis in human and experimental cholestasis. Cell Death Dis. 2016;7:e2390.
Al-Khaifi A, Rudling M, Angelin B. An FXR agonist reduces bile acid synthesis independently of increases in FGF19 in healthy volunteers. Gastroenterology. 2018;155:1012–16.
Albillos A, de Gottardi A, Rescigno M. The gut-liver axis in liver disease: pathophysiological basis for therapy. J Hepatol. 2020;72:558–77.
Allegretti JR, Kassam Z, Carrellas M, Mullish BH, Marchesi JR, Pechlivanis A, et al. Fecal microbiota transplantation in patients with primary sclerosing cholangitis: a Pilot Clinical Trial. Am J Gastroenterol. 2019;114:1071–9.
Allen K, Kim ND, Moon JO, Copple BL. Upregulation of early growth response factor-1 by bile acids requires mitogen-activated protein kinase signaling. Toxicol Appl Pharmacol. 2010;243:63–7.
Allen K, Jaeschke H, Copple BL. Bile acids induce inflammatory genes in hepatocytes. Am J Pathol. 2011;178:175–86.
Ananthanarayanan M, Balasubramanian N, Makishima M, Mangelsdorf DJ, Suchy FJ. Human bile salt export pump promoter is transactivated by the farnesoid x receptor/bile acid receptor. J Biol Chem. 2001;276:28857–65.
Assimakopoulos SF, Charonis AS. Uncovering the molecular events associated with increased intestinal permeability in liver cirrhosis: the pivotal role of enterocyte tight junctions and future perspectives. J Hepatol. 2013;59:1144–46.
Baghdasaryan A, Fuchs CD, Osterreicher CH, Lemberger UJ, Halilbasic E, Pahlman I, et al. Inhibition of intestinal bile acid absorption improves cholestatic liver and bile duct injury in a mouse model of sclerosing cholangitis. J Hepatol. 2016;64:674–81.
Bajer L, Kverka M, Kostovcik M, Macinga P, Daorak J, Stehlikova Z, et al. Distinct gut microbiota profiles in patients with primary sclerosing cholangitis and ulcerative colitis. World J Gastroenterol. 2017;23:4548–58.
Baumann U, Sturm E, Lacaille F, Gonzales E, Arnell H, Fischler B, et al. Effects of odevixibat on pruritus and bile acids in children with cholestatic liver disease: phase 2 study. Clin Res Hepatol Gastroenterol. 2021;45:101751.
Bertolini A, Fiorotto R, Strazzabosco M. Bile acids and their receptors: modulators and therapeutic targets in liver inflammation. Semin Immunopathol. 2022;44:547–64.
Beuers U, Trauner M, Jansen P, Paupon R. New paradigms in the treatment of hepatic cholestasis: from UDCA to FXR, PXR and beyond. J Hepatol. 2015;62:S25–37.
Bogomolov P, Alexandrov A, Voronkova N, Macievich M, Kokina K, Petrachenkova M, et al. Treatment of chronic hepatitis d with the entry inhibitor myrcludex b: first results of a phase II/III study. J Hepatol. 2016;65:490–8. https://doi.org/10.1016/j.jhep.2016.04.016.
Bowlus CL, Eksteen B, Cheung AC, Thorburn D, Moylan CA, Pockros PJ, et al. Safety, tolerability, and efficacy of maralixibat in adults with primary sclerosing cholangitis: open-label pilot study. Hepatol Commun. 2023;7:e0153.
Boyer JL, Trauner M, Mennone A, Soroka CJ, Cai SY, Moustafa T, et al. Upregulation of a basolateral FXR-dependent bile acid efflux transporter ostalpha-ostbeta in cholestasis in humans and rodents. Am J Physiol Gastrointest Liver Physiol. 2006;290:1124–30.
Cai S, Boyer JL. The role of bile acids in cholestatic liver injury. Ann Transl Med. 2021;9:737.
Carambia A, Freund B, Schwinge D, Heine M, Laschtowitz A, Huber S, et al. TGFβ-dependent induction of cd4 (+)cd25 (+)foxp3 (+) tregs by liver sinusoidal endothelial cells. J Hepatol. 2014;61:594–9.
Chascsa D, Carey EJ, Lindor KD. Old and new treatments for primary biliary cholangitis. Liver Int. 2017;37:490–9.
Che Y, Xu W, Ding C, He T, Xu X, Shuai Y, et al. Bile acids target mitofusin 2 to differentially regulate innate immunity in physiological versus cholestatic conditions. Cell Rep. 2023;42:112011.
Chen W, Owsley E, Yang Y, Stroup D, Chiang JY. Nuclear receptor-mediated repression of human cholesterol 7alpha-hydroxylase gene transcription by bile acids. J Lipid Res. 2001;42:1402–12.
Chen F, Ma L, Dawson PA, Sinal CJ, Sehayek E, Gonzalez FJ, et al. Liver receptor homologue-1 mediates species- and cell line-specific bile acid-dependent negative feedback regulation of the apical sodium-dependent bile acid transporter. J Biol Chem. 2003;278:19909–16.
Chen HL, Wu SH, Hsu SH, Liou BY, Chen HL, Chang MH. Jaundice revisited: recent advances in the diagnosis and treatment of inherited cholestatic liver diseases. J Biomed Sci. 2018;25:75.
Cheung AC, Lammers WJ, Murillo Perez CF, van Buuren HR, Gulamhusein A, Trivedi PJ, et al. Effects of Age and sex of response to Ursodeoxycholic Acid and Transplant-free survival in patients with primary biliary cholangitis. Clin Gastroenterol Hepatol. 2019;17:2076–84.
Chiang JY. Regulation of bile acid synthesis: pathways, nuclear receptors, and mechanisms. J Hepatol. 2004;40:539–51.
Chiang JY. Bile acids: regulation of synthesis. J Lipid Res. 2009;50:1955–66.
Chiang J, Ferrell JM. Bile acid biology, pathophysiology, and therapeutics. Clin Liver Dis (Hoboken). 2020;15:91–4.
Chopyk DM, Grakoui A. Contribution of the intestinal microbiome and gut barrier to hepatic disorders. Gastroenterology. 2020;159:849–63.
Costello SP, Hughes PA, Waters O, Bryant RV, Vincent AD, Blathford P, et al. Effect of fecal microbiota transplantation on 8-Week remission in patients with Ulcerative Colitis: a Randomized Clinical Trial. JAMA. 2019;321:156–64.
D’Aldebert E, Biyeyeme BMM, Mergey M, Wendum D, Firrincieli D, Coilly A, et al. Bile salts control the antimicrobial peptide cathelicidin through nuclear receptors in the human biliary epithelium. Gastroenterology. 2009;136:1435–43.
Dawson PA, Lan T, Rao A. Bile acid transporters. J Lipid Res. 2009;50:2340–57.
de Vos WM, Tilg H, Van Hul M, Cani PD. Gut microbiome and health: mechanistic insights. Gut. 2022;71:1020–32.
Devkota S, Chang EB. Interactions between diet, bile acid metabolism, gut microbiota, and inflammatory bowel diseases. Dig Dis. 2015;33:351–6.
Dong H, Qu S, Chen X, Zhu H, Tai X, Pan J. Changes in the cytokine expression of peripheral treg and th17 cells in children with rotavirus enteritis. Exp Ther Med. 2015;10 (2):679–82.
Fang S, Wang T, Li Y, Xue H, Zou J, Cai J, et al. Gardenia jasminoides Ellis polysaccharide ameliorates cholestatic liver injury by alleviating gut microbiota dysbiosis and inhibiting the TLR4/NF-kB signaling pathway. Int J Biol Macromol. 2022;205:23–36.
Faubion WA, Guicciardi ME, Miyoshi H, Bronk SF, Roberts PJ, Svingen PA, et al. Toxic bile salts induce rodent hepatocyte apoptosis via direct activation of FAS. J Clin Invest. 1999;103:137–45.
Fickert P, Hirschfield GM, Denk G, Marschall HU, Altorjay I, Färkkilä M, et al. norUrsodeoxycholic acid improves Cholestasis in Primary Sclerosing Cholangitis. J Hepatol. 2017;67:549–58.
Fiorucci S, Baldoni M, Ricci P, Zampella A, Distrutti E, Biagioli M. Bile acid-activated receptors and the regulation of macrophages function in metabolic disorders. Curr Opin Pharmacol. 2020;53:45–54.
Fuchs CD, Trauner M. Role of bile acids and their receptors in gastrointestinal and hepatic pathophysiology. Nat Rev Gastroenterol Hepatol. 2022;19:432–50.
Fuchs CD, Paumgartner G, Wahlstrom A, Schwabl P, Reiberger T, Leditznig N, et al. Metabolic preconditioning protects bsep/abcb11 (-/-) mice against cholestatic liver injury. J Hepatol. 2017;66:95–101.
Gadaleta RM, van Erpecum KJ, Oldenburg B, Willemsen EC, Renooij W, et al. Farnesoid X receptor activation inhibits inflammation and preserves the intestinal barrier in inflammatory bowel disease. Gut. 2011;60:463–72.
Gahan CG, Hill C. Listeria monocytogenes: survival and adaptation in the gastrointestinal tract. Front Cell Infect Microbiol. 2014;4:9.
Gan C, Cai Q, Tang C, Gao J. Inflammasomes and pyroptosis of liver cells in liver fibrosis. Front Immunol. 2022;13:896473.
Geier A, Zollner G, Dietrich CG, Wagner M, Fickert P, Denk H, et al. Cytokine-independent repression of rodent ntcp in obstructive cholestasis. Hepatology. 2005;41:470–7.
Gerussi A, D’Amato D, Cristoferi L, O’Donnell SE, Carbone M, Invernizzi P, et al. Multiple therapeutic targets in rare cholestatic liver diseases: time to redefine treatment strategies. Ann Hepatol. 2020;19:5–16.
Gijbels E, Pieters A, De Muynck K, Vinken M, Devisscher L. Rodent models of cholestatic liver disease: a practical guide for translational research. Liver Int. 2021;41:656–82.
Gonzales E, Hardikar W, Stormon M, Baker A, Hierro L, Gliwicz D, et al. Efficacy and safety of maralixibat treatment in patients with alagille syndrome and cholestatic pruritus (iconic): a randomised phase 2 study. Lancet. 2021;398:1581–92.
Goodwin B, Jones SA, Price RR, Watson MA, Mckee DD, Moore LB, Galardi C, Wilson JG, Lewis MC, Roth ME, et al. A regulatory cascade of the nuclear receptors fxr, shp-1, and lrh-1 represses bile acid biosynthesis. Mol Cell. 2000;6:517–26. https://doi.org/10.1016/s1097-2765(00)00051-4.
Guilliams M, Bonnardel J, Haest B, Vanderborght B, Wagner C, Remmerie A, et al. Spatial proteogenomics reveals distinct and evolutionarily conserved hepatic macrophage niches. Cell. 2022;185:379–96.
Gulamhusein AF, Hirschfield GM. Primary biliary cholangitis: pathogenesis and therapeutic opportunities. Nat Rev Gastroenterol Hepatol. 2020;17:93–110.
Guo C, Chen WD, Wang YD. TGR5, not only a metabolic regulator. Front Physiol. 2016;7:646.
Guzior DV, Quinn RA. Review: microbial transformations of human bile acids. Microbiome. 2021;9:140.
Haifer C, Paramsothy S, Kaakoush NO, Saikal A, Ghaly S, Yang T, et al. Lyophilised oral faecal microbiota transplantation for ulcerative colitis (LOTUS): a randomised, double-blind, placebo-controlled trial. Lancet Gastroenterol Hepatol. 2022;7:141–51.
Hammerich L, Bangen JM, Govaere O, Zimmermann HW, Gassler N, Huss S, et al. Chemokine receptor CCR6-dependent accumulation of gammadelta T cells in injured liver restricts hepatic inflammation and fibrosis. Hepatology. 2014;59:630–42.
Hao H, Cao L, Jiang C, Che Y, Zhang S, Takahashi S, et al. Farnesoid X receptor regulation of the nlrp3 inflammasome underlies cholestasis-associated sepsis. Cell Metab. 2017;25:856–67.
Harpavat S, Garcia-Prats JA, Anaya C, Brandt ML, Lupo PJ, Finegold MJ, et al. Diagnostic yield of newborn screening for biliary atresia using direct or conjugated bilirubin measurements. JAMA. 2020;323 (12):1141.
Hegade VS, Speight RA, Etherington RE, Jones DE. Novel bile acid therapeutics for the treatment of chronic liver diseases. Th Adv Gastroenterol. 2016;9:376–91.
Hegade VS, Kendrick SF, Dobbins RL, Miller SR, Thompson D, Richards D, et al. Effect of ileal bile acid transporter inhibitor GSK2330672 on pruritus in primary biliary cholangitis: a double-blind, randomised, placebo-controlled, crossover, phase 2a study. Lancet. 2017;389:1114–23.
Hiippala K, Jouhten H, Ronkainen A, Hartikainen A, Kainulainen V, Jalanka J, et al. The potential of gut commensals in reinforcing intestinal barrier function and alleviating inflammation. Nutrients. 2018;10:988.
Jansen PLM, Ghallab A, Vartak N, Reif R, Schaap FG, Hampe J, et al. The ascending pathophysiology of cholestatic liver disease. Hepatology. 2017;65:722–38.
Jee JJ, Yang L, Shivakumar P, Xu PP, Mourya R, Thanekar U, et al. Maternal regulation of biliary disease in neonates via gut microbial metabolites. Nat Commun. 2022;13:18.
Jia W, Xie G, Jia W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat Rev Gastroenterol Hepatol. 2018;15:111–28.
Johansson H, Svensson JF, Almström M, Van Hul N, Rudling M, Angelin B, et al. Regulation of bile acid metabolism in biliary atresia: reduction of FGF19 by Kasai portoenterostomy and possible relation to early outcome. J Intern Med. 2020;287:534–45.
Juanola O, Hassan M, Kumar P, Yilmaz B, Keller I, Simillion C, et al. Intestinal microbiota drives cholestasis-induced specific hepatic gene expression patterns. Gut Microbes. 2021;13 (1):1–20.
Just S, Mondot S, Ecker J, Wegner K, Rath E, Gau L, et al. The gut microbiota drives the impact of bile acids and fat source in diet on mouse metabolism. Microbiome. 2018;6:134.
Keitel V, Häussinger D. Role of TGR5 (gpbar1) in liver disease. Semin Liver Dis. 2018;38:333–9.
Keitel V, Cupisti K, Ullmer C, Knoefel WT, Kubitz R, Haussinger D. The membrane-bound bile acid receptor TGR5 is localized in the epithelium of human gallbladders. Hepatology. 2009;50:861–70.
Kerr TA, Saeki S, Schneider M, Schaefer K, Berdy S, Redder T, et al. Loss of nuclear receptor shp impairs but does not eliminate negative feedback regulation of bile acid synthesis. Dev Cell. 2002;2:713–20.
Kim CH. Immune regulation by microbiome metabolites. Immunology. 2018;154:220–9.
Kim I, Ahn SH, Inagaki T, Choi M, Ito S, Guo GL, et al. Differential regulation of bile acid homeostasis by the farnesoid x receptor in liver and intestine. J Lipid Res. 2007;48:2664–72. https://doi.org/10.1194/jlr.M700330-JLR200.
Kisiela M, Skarka A, Ebert B, Maser E. Hydroxysteroid dehydrogenases (hsds) in bacteria: a bioinformatic perspective. J Steroid Biochem Mol Biol. 2012;129:31–46.
Kowdley KV, Vuppalanchi R, Levy C, Floreani A, Andreone P, Larusso NF, et al. A randomized, placebo-controlled, phase ii study of obeticholic acid for primary sclerosing cholangitis. J Hepatol. 2020;73:94–101.
Kummen M, Hov JR. The gut microbial influence on cholestatic liver disease. Liver Int. 2019;39:1186–96.
Kummen M, Holm K, Anmarkrud JA, Nygård S, Vesterhus M, Høivik ML, et al. The gut microbial profile in patients with primary sclerosing cholangitis is distinct from patients with ulcerative colitis without biliary disease and healthy controls. Gut. 2017;66 (4):611–9.
Laue T, Baumann U. Odevixibat: an investigational inhibitor of the ileal bile acid transporter (IBAT) for the treatment of biliary atresia. Expert Opin Investig Drugs. 2022;31:1143–5020.
Lee JY, Tsolis RM, Baumler AJ. The microbiome and gut homeostasis. Science. 2022;377:eabp9960.
Lemasters JJ, Nieminen AL, Qian T, Trost LC, Elmore SP, Nishimura Y, et al. The mitochondrial permeability transition in cell death: a common mechanism in necrosis, apoptosis and autophagy. Biochim Biophys Acta. 1998;1366:177–96.
Lemoinne S, Kemgang A, Ben Belkacem K, Straube M, Jegou S, Corpechot C, et al. Fungi participate in the dysbiosis of gut microbiota in patients with primary sclerosing cholangitis. Gut. 2019;69 (1):92–102.
Levy C, Kendrick S, Bowlus CL, Tanaka A, Jones D, Kremer AE, et al. Glimmer: a randomized phase 2b dose-ranging trial of linerixibat in primary biliary cholangitis patients with pruritus. Clin Gastroenterol Hepatol. 2023;21:1902–12.
Li Y, Tang R, Leung PSC, Gershwin ME, Ma X. Bile acids and intestinal microbiota in autoimmune cholestatic liver diseases. Autoimmun Rev. 2017;16:885–96.
Li B, Selmi C, Tang R, Gershwin ME, Ma X. The microbiome and autoimmunity: a paradigm from the gut-liver axis. Cell Mol Immunol. 2018;15:595–609.
Li N, Zhan S, Tian Z, Liu C, Xie Z, Zhang S, et al. Alterations in bile acid metabolism associated with inflammatory bowel disease. Inflamm Bowel Dis. 2021;27:1525–40.
Liao L, Schneider KM, Galvez EJC, Frissen M, Marschall H, Su H, et al. Intestinal dysbiosis augments liver disease progression via nlrp3 in a murine model of primary sclerosing cholangitis. Gut. 2019;68:1477–92.
Licata LA, Nguyen CT, Burga RA, Falanga V, Espat NJ, Ayala A, et al. Biliary obstruction results in pd-1-dependent liver T cell dysfunction and acute inflammation mediated by Th17 cells and neutrophils. J Leukoc Biol. 2013;94:813–23.
Liu X, Li J, Shi M, Fu J, Wang J, Kang W, et al. Melatonin improves cholestatic liver disease via the gut-liver axis. J Pineal Res. 2024;76:e12929.
Loomes KM, Squires RH, Kelly D, Rajwal S, Soufi N, Lachaux A, et al. Maralixibat for the treatment of PFIC: long-term, ibat inhibition in an open-label, phase 2 study. Hepatol Commun. 2022;6:2379–90.
Lu TT, Makishima M, Repa JJ, Schoonjans K, Kerr TA, Auwerx J, et al. Molecular basis for feedback regulation of bile acid synthesis by nuclear receptors. Mol Cell. 2000;6 (3):507–15.
Lv L, Fang D, Shi D, Chen DY, Yan R, Zhu YX, et al. Alterations and correlations of the gut microbiome, metabolism and immunity in patients with primary biliary cirrhosis. Environ Microbiol. 2016;18:2272–86.
Lynch LE, Hair AB, Soni KG, Yang H, Gollins LA, Narvaez-Rivas M, et al. Cholestasis impairs gut microbiota development and bile salt hydrolase activity in preterm neonates. Gut Microbes. 2023;15:2183690.
Malhi H, Kaufman RJ. Endoplasmic reticulum stress in liver disease. J Hepatol. 2011;54:795–809.
Martel J, Chang SH, Ko YF, Hwang TL, Young JD, Ojcius DM. Gut barrier disruption and chronic disease. Trends Endocrinol Metab. 2022;33:247–65.
Meadows V, Baiocchi L, Kundu D, Sato K, Fuentes Y, Wu C, et al. Biliary epithelial senescence in liver disease: there will be sasp. Front Mol Biosci. 2021;8:803098.
Miethke AG, Zhang W, Simmons J, Taylor AE, Shi T, Shanmukhappa SK, et al. Pharmacological inhibition of apical sodium-dependent bile acid transporter changes bile composition and blocks progression of sclerosing cholangitis in multidrug resistance 2 knockout mice. Hepatology. 2016;63:512–23.
Monte MJ, Marin JJ, Antelo A, Vazquez-Tato J. Bile acids: chemistry, physiology, and pathophysiology. World J Gastroenterol. 2009;15:804–16.
Mridha AR, Wree A, Robertson A, Yeh MM, Johnson CD, Van Rooyen DM, et al. Nlrp3 inflammasome blockade reduces liver inflammation and fibrosis in experimental Nash in mice. J Hepatol. 2017;66:1037–46.
Munshi MK, Priester S, Gaudio E, Yang F, Alpini G, Mancinelli R, et al. Regulation of biliary proliferation by neuroendocrine factors: implications for the pathogenesis of cholestatic liver diseases. Am J Pathol. 2011;178:472–84.
Nastasi C, Candela M, Bonefeld CM, Geisler C, Hansen M, Krejsgaard T, et al. The effect of short-chain fatty acids on human monocyte-derived dendritic cells. Sci Rep. 2015;5:16148.
Neimark E, Chen F, Li X, Shneider BL. Bile acid-induced negative feedback regulation of the human ileal bile acid transporter. Hepatology. 2004;40:149–56.
Novobrantseva TI, Majeau GR, Amatucci A, Kogan S, Brenner I, Casola S, et al. Attenuated liver fibrosis in the absence of B cells. J Clin Invest. 2005;115:3072–82.
Nyholm I, Hukkinen M, Pihlajoki M, Davidson JR, Tyraskis A, Lohi J, et al. Serum FGF19 predicts outcomes of kasai portoenterostomy in biliary atresia. Hepatology. 2023;77:1263–73.
O’Brien KM, Allen KM, Rockwell CE, Towery K, Luyendyk JP, Copple BL. Il-17A synergistically enhances bile acid-induced inflammation during obstructive cholestasis. Am J Pathol. 2013;183:1498–507.
Ostadmohammadi S, Azimirad M, Houri H, Naseri K, Javanmard E, Mirjalali H, et al. Characterization of the gut microbiota in patients with primary sclerosing cholangitis compared to inflammatory bowel disease and healthy controls. Mol Biol Rep. 2021;48:5519–29.
Pean N, Doignon I, Garcin I, Besnard A, Julien B, Liu B, et al. The receptor tgr5 protects the liver from bile acid overload during liver regeneration in mice. Hepatology. 2013;58:1451–60.
Plass JR, Mol O, Heegsma J, Geuken M, Faber KN, Jansen PL, Muller M. Farnesoid X receptor and bile salts are involved in transcriptional regulation of the gene encoding the human bile salt export pump. Hepatology. 2002;35:589–96.
Poupon R, Chretien Y, Poupon RE, Ballet F, Calmus Y, Darnis F. Is ursodeoxycholic acid an effective treatment for primary biliary cirrhosis? Lancet. 1987;1:834–6.
Qi X, Yang M, Stenberg J, Dey R, Fogwe L, Alam MS, et al. Gut microbiota mediated molecular events and therapy in liver diseases. World J Gastroenterol. 2020;26:7603–18.
Qin N, Yang F, Li A, Prifti E, Chen Y, Shao L, et al. Alterations of the human gut microbiome in liver cirrhosis. Nature. 2014;513:59–64.
Rager SL, Zeng MY. The gut-liver axis in pediatric liver health and disease. Microorganisms. 2023;11:597.
Rahman AH, Aloman C. Dendritic cells and liver fibrosis. Biochim Biophys Acta. 2013;1832:998–1004.
Reich M, Deutschmann K, Sommerfeld A, Klindt C, Kluge S, Kubitz R, et al. Tgr5 is essential for bile acid-dependent cholangiocyte proliferation in vivo and in vitro. Gut. 2016;65:487–501.
Reich M, Spomer L, Klindt C, Fuchs K, Stindt J, Deutschmann K, et al. Downregulation of TGR5 (gpbar1) in biliary epithelial cells contributes to the pathogenesis of sclerosing cholangitis. J Hepatol. 2021;75:634–46.
Renga B, Mencarelli A, Cipriani S, Amore DC, Carino A, Bruno A, et al. The bile acid sensor fxr is required for immune-regulatory activities of tlr-9 in intestinal inflammation. PLoS ONE. 2013;8:e54472.
Rios-Covian D, Ruas-Madiedo P, Margolles A, Gueimonde M, de Los RC, Salazar N. Intestinal short chain fatty acids and their link with diet and human health. Front Microbiol. 2016;7:185.
Robin M, Appelman MD, Vos HR, van Es RM, Paton JC, Paton AW, et al. Calnexin depletion by endoplasmic reticulum stress during cholestasis inhibits the na (+)-taurocholate cotransporting polypeptide. Hepatol Commun. 2018;2:1550–66.
Rost D, Rudolph G, Kloeters-Plachky P, Stiehl A. Effect of high-dose ursodeoxycholic acid on its biliary enrichment in primary sclerosing cholangitis. Hepatology. 2004;40:693–8.
Sabino J, Vieira-Silva S, Machiels K, Joossens M, Falony G, Ballet V, et al. Primary sclerosing cholangitis is characterized by intestinal dysbiosis independent from ibd. Gut. 2016;65 (10):1681–9.
Sanyal AJ, Ling L, Beuers U, Depaoli AM, Lieu HD, Harrison SA, et al. Potent suppression of hydrophobic bile acids by aldafermin, an FGF19 analogue, across metabolic and cholestatic liver diseases. JHEP Rep. 2021;3:100255.
Sayin SI, Wahlstrom A, Felin J, Jantti S, Marschall HU, Bamberg K, et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring fxr antagonist. Cell Metab. 2013;17:225–35.
Schnabl B, Brenner DA. Interactions between the intestinal microbiome and liver diseases. Gastroenterology. 2014;146:1513–24.
Schneider KM, Candels LS, Hov JR, Myllys M, Hassan R, Schneider CV, et al. Gut microbiota depletion exacerbates cholestatic liver injury via loss of fxr signalling. Nat Metab. 2021;3:1228–41.
Schramm C, Wedemeyer H, Mason A, Hirschfield GM, Levy C, Kowdley KV, et al. Farnesoid X receptor agonist tropifexor attenuates cholestasis in a randomised trial in patients with primary biliary cholangitis. JHEP Rep. 2022;4:100544.
Shi T, Malik A, Yang Vom Hofe A, Matuschek L, Mullen M, Lages CS, et al. Farnesoid X receptor antagonizes macrophage-dependent licensing of effector T lymphocytes and progression of sclerosing cholangitis. Sci Transl Med. 2022;14:eabi4354.
Shojaie L, Iorga A, Dara L. Cell death in liver diseases: a review. Int J Mol Sci. 2020;21:9682.
Simbrunner B, Trauner M, Reiberger T. Review article: therapeutic aspects of bile acid signalling in the gut-liver axis. Aliment Pharmacol Ther. 2021;54:1243–62.
Singh N, Gurav A, Sivaprakasam S, Brady E, Padia R, Shi H, et al. Activation of GPR109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity. 2014;40:128–39.
Slijepcevic D, Roscam AR, Fuchs CD, Haazen L, Beuers U, Trauner M, Oude ER, van de Graaf S. Na (+) -taurocholate cotransporting polypeptide inhibition has hepatoprotective effects in cholestasis in mice. Hepatology. 2018;68:1057–69.
Song W, Sun L, Zhu Z, Wei L, Qu W, Zeng Z, et al. Association of gut microbiota and metabolites with disease progression in children with biliary atresia. Front Immunol. 2021;12:698900.
Sorribas M, Jakob MO, Yilmaz B, Li H, Stutz D, Noser Y, et al. FXR modulates the gut-vascular barrier by regulating the entry sites for bacterial translocation in experimental cirrhosis. J Hepatol. 2019;71:1126–40.
Tang R, Wei Y, Li Y, Chen W, Chen H, Wang Q, et al. Gut microbial profile is altered in primary biliary cholangitis and partially restored after UDCA therapy. Gut. 2018;67 (3):534–41.
Thibaut MM, Bindels LB. Crosstalk between bile acid-activated receptors and microbiome in entero-hepatic inflammation. Trends Mol Med. 2022;28:223–36.
Thompson RJ, Arnell H, Artan R, Baumann U, Calvo PL, Czubkowski P, et al. Odevixibat treatment in progressive familial intrahepatic cholestasis: a randomised, placebo-controlled, phase 3 trial. Lancet Gastroenterol Hepatol. 2022;7:830–42.
Tilg H, Adolph TE, Trauner M. Gut-liver axis: pathophysiological concepts and clinical implications. Cell Metab. 2022;34:1700–18.
Trauner M, Fuchs CD. Novel therapeutic targets for cholestatic and fatty liver disease. Gut. 2022;71:194–209.
Trauner M, Nevens F, Shiffman ML, Drenth JPH, Bowlus CL, Vargas V, et al. Long-term efficacy and safety of obeticholic acid for patients with primary biliary cholangitis: 3-year results of an international open-label extension study. Lancet Gastroenterol Hepatol. 2019a;4:445–53.
Trauner M, Gulamhusein A, Hameed B, Caldwell S, Shiffman ML, Landis C, et al. The nonsteroidal farnesoid x receptor agonist cilofexor (GS-9674) improves markers of cholestasis and liver injury in patients with primary sclerosing cholangitis. Hepatology. 2019b;70:788–801.
Turner JR. Intestinal mucosal barrier function in health and disease. Nat Rev Immunol. 2009;9:799–809.
Ubeda M, Lario M, Munoz L, Borrero MJ, Rodriguez-Serrano M, Sanchez-Diaz AM, et al. Obeticholic acid reduces bacterial translocation and inhibits intestinal inflammation in cirrhotic rats. J Hepatol. 2016;64 (5):1049–57.
Vaishnava S, Yamamoto M, Severson KM, Ruhn KA, Yu X, Koren O, et al. The antibacterial lectin regime promotes the spatial segregation of microbiota and host in the intestine. Science. 2011;334:255–8.
Verbeke L, Farre R, Verbinnen B, Covens K, Vanuytsel T, Verhaegen J, et al. The FXR agonist obeticholic acid prevents gut barrier dysfunction and bacterial translocation in cholestatic rats. Am J Pathol. 2015;185:409–19.
Wahlstrom A, Sayin SI, Marschall HU, Backhed F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metab. 2016;24:41–50.
Wan YY, Sheng L. Regulation of bile acid receptor activity. Liver Res. 2018;2:180–5.
Wang L, Lee YK, Bundman D, Han Y, Thevananther S, Kim CS, et al. Redundant pathways for negative feedback regulation of bile acid production. Dev Cell. 2002;2:721–31.
Wang Y, Aoki H, Yang J, Peng K, Liu R, Li X, et al. The role of sphingosine 1-phosphate receptor 2 in bile-acid-induced cholangiocyte proliferation and cholestasis-induced liver injury in mice. Hepatology. 2017;65:2005–18.
Wang J, Qian T, Jiang J, et al. Gut microbial profile in biliary atresia: a case-control study. J Gastroenterol Hepatol. 2019;35:334–42.
Wang L, Cao ZM, Zhang LL, Li JM, Lv WL. The role of gut microbiota in some Liver diseases: from an immunological perspective. Front Immunol. 2022;13:923599.
Wang YD, Chen WD, Wang M, Yu D, Forman BM, Huang W. Farnesoid X receptor antagonizes nuclear factor kappab in hepatic inflammatory response. Hepatology. 2008;48:1632-43.
Wells JM, Brummer RJ, Derrien M, Macdonald TT, Troost F, Cani PD, et al. Homeostasis of the gut barrier and potential biomarkers. Am J Physiol Gastrointest Liver Physiol. 2017;312:171–93.
Wree A, Eguchi A, Mcgeough MD, Pena CA, Johnson CD, Canbay A, et al. Nlrp3 inflammasome activation results in hepatocyte pyroptosis, liver inflammation, and fibrosis in mice. Hepatology. 2014;59 (3):898–910.
Xiang J, Zhang Z, Xie H, Zhang C, Bai Y, Cao H, et al. Effect of different bile acids on the intestine through enterohepatic circulation based on FXR. Gut Microbes. 2021;13:1949095.
Xiao Y, Wang Y, Liu Y, Wang W, Tian X, Chen S, et al. A nonbile acid farnesoid X receptor agonist tropifexor potently inhibits cholestatic liver injury and fibrosis by modulating the gut-liver axis. Liver Int. 2021;41:2117–31.
Xiao Y, Zhao C, Tai Y, Li B, Lan T, Lai E, et al. Sting mediates hepatocyte pyroptosis in liver fibrosis by epigenetically activating the nlrp3 inflammasome. Redox Biol. 2023;62:102691.
Xu W, Che Y, Zhang Q, Huang H, Ding C, Wang Y, et al. Apaf-1 pyroptosome senses mitochondrial permeability transition. Cell Metab. 2021;33 (2):424–36.
Yan N, Yan T, Xia Y, Hao H, Wang G, Gonzalez FJ. The pathophysiological function of non-gastrointestinal farnesoid X receptor. Pharmacol Ther. 2021;226:107867.
Yang X, Lu D, Zhuo J, Lin Z, Yang M, Xu X. The gut-liver axis in immune remodeling: new insight into liver diseases. Int J Biol Sci. 2020;16 (13):2357–66.
Yang T, Yang S, Zhao J, Wang P, Li S, Jin Y, et al. Comprehensive analysis of gut microbiota and fecal bile acid profiles in children with biliary atresia. Front Cell Infect Microbiol. 2022;12:914247.
Yang S, Chang N, Li W, Yang T, Xue R, Liu J, et al. Necroptosis of macrophage is a key pathological feature in biliary atresia via GDCA/S1PR2/ZBP1/p-MLKL axis. Cell Death Dis. 2023;14:175.
Yin XM, Ding WX. Death receptor activation-induced hepatocyte apoptosis and liver injury. Curr Mol Med. 2003;3:491–508.
Yokoda RT, Rodriguez EA. Review: pathogenesis of cholestatic liver diseases. World J Hepatol. 2020;12:423–35.
Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Signal Transduct Target Ther. 2021;6:121–8.
Zeng J, Fan J, Zhou H. Bile acid-mediated signaling in cholestatic liver diseases. Cell Biosci. 2023;13:77.
Zhang Y, Li Z, Gou H, Song X, Zhang L. The gut microbiota-bile acid axis: a potential therapeutic target for liver fibrosis. Front Cell Infect Microbiol. 2022;12:945368.
Zhou X, Zhang X, Zhao N, Zhang L, Qiu W, Song C, et al. Gut microbiota deficiency exacerbates liver injury in bile duct-ligated mice via inflammation and lipid metabolism. Int J Mol Sci. 2023;24:3180.
Zollner G, Fickert P, Silbert D, Fuchsbichler A, Stumptner C, Zatloukal K, et al. Induction of short heterodimer partner 1 precedes downregulation of ntcp in bile duct-ligated mice. Am J Physiol Gastrointest Liver Physiol. 2002;282:184–91.
Zollner G, Marschall HU, Wagner M, Trauner M. Role of nuclear receptors in the adaptive response to bile acids and cholestasis: pathogenetic and therapeutic considerations. Mol Pharm. 2006;3:231–51.
Funding
This work was supported by the National Natural Science Foundation of China grants (82300574).
Author information
Authors and Affiliations
Contributions
All authors read and confirmed the final manuscript. Dayan Sun and Chuanping Xie participated in drafting the article; Dayan Sun, Yong Zhao, Junmin Liao, Shuangshuang Li, Yanan Zhang, Dingding Wang, Kaiyun Hua, Jingbin Du, Jinshi Huang contributed to reviewing the article. All authors read and approved the final manuscript.
Authors’ statement
The authors hereby confirm that neither the manuscript nor any part of it has been published or is being considered for publication elsewhere. We acknowledge that all authors participated sufficiently in the work and take public responsibility for its content.
Corresponding author
Ethics declarations
Ethic approval and consent to participation
Not applicable.
Consent for publication
The authors agree to the publication of all the data involved in this article. No data from other entities are used in this study.
Conflict of interest
No potential conflict of interest was reported by the author (s).
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
sun, D., Xie, C., Zhao, Y. et al. The gut microbiota-bile acid axis in cholestatic liver disease. Mol Med 30, 104 (2024). https://doi.org/10.1186/s10020-024-00830-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10020-024-00830-x