
Abstract
The high temperatures reached during cremation lead to the destruction of organic matter preventing the use of traditional isotopic methods for dietary reconstructions. Still, strontium isotope (87Sr/86Sr) and concentration ([Sr]) analyses of cremated human remains offer a novel way to assess changing consumption patterns in past populations that practiced cremation, as evidenced by a large amount of new data obtained from Metal Ages and Gallo-Roman human remains from Destelbergen, Belgium. The Gallo-Roman results show significantly higher [Sr] and a narrower interquartile range in 87Sr/86Sr (0.7093–0.7095), close to the value of modern-day seawater (0.7092). This contrasts with the Metal Ages results, which display lower concentrations and a wider range in 87Sr/86Sr (0.7094–0.7098). This typical Sr signature is also reflected in other sites and is most likely related to an introduction of marine Sr in the form of salt as a food preservative (e.g. salt-rich preserved meat, fish and fish sauce). Paradoxically, this study highlights caution is needed when using 87Sr/86Sr for palaeomobility studies in populations with high salt consumption.
Similar content being viewed by others


Introduction
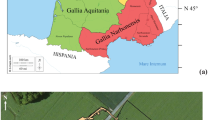
The site of Destelbergen “Eenbeekeinde”, close to Ghent, Belgium, is located on the left bank of the Scheldt river (Fig. 1) and is one of the rare sites where a large number of both Metal Ages and Gallo-Roman cremation burials were excavated1,2,3,4. Steady advancements in Sr isotopic applications prove that cremated bone shows even less contamination than previously favoured tooth enamel5 and provide new methods to gain knowledge on communities who practiced cremation. As such, this site offers a unique opportunity to study shifts in mobility, diet and landscape use from the Iron Age to the Roman period by combining archaeological evidence with state-of-the-art 87Sr/86Sr and [Sr] analyses of cremated human remains. In the Metal Ages cemetery (107 burials), a general chronological evolution from east to west was found6. In the Gallo-Roman cemetery, which was with its 204 burials far more extensive than most common familial cemeteries of this period7, graves were usually simple pits. Besides Destelbergen, both new and published data from three other Belgian sites was analysed for comparison. Blicquy offered just like Destelbergen both Metal Ages and Roman cemeteries at close distance from each other. Additionally, in relatively close proximity to each other and located further inland in the Meuse basin, two more sites serve as reference data. Metal Ages cemetery Herstal “Pré Wigier” delivered already published results8. The cemetery Fize-le-Marsal “Beauflot-Pivache” revealed Gallo-Roman burials on which new analyses were performed. For more information on the sites, see Suppl. Text 1.

Location of the Destelbergen, Blicquy, Fize-le-Marsal and Herstal sites within Belgium on the inset map and the six new plant sampling locations around Destelbergen with median 87Sr/86Sr superimposed on a geological map (map created using QGIS version 3.12.0-București, https://qgis.org/).
Sufficient evidence shows that in western Europe the Iron Age diet was fairly homogeneous, rich in animal protein in the form of meat and dairy and almost exclusively terrestrial e.g.:9,10,11. Although, for Belgium, clear archaeozoological and archaeobotanical evidence on Iron Age food economies is scarce, data from neighbouring areas indicates that the population relied mainly on varying proportions of cattle, sheep, and pig as main suppliers of animal protein, and cereal and, to a lesser extent, pulses as a staple food10,12,13. A total lack of fish bone at Iron Age sites led to the conclusion that fish was not consumed in Belgium, nor in England, but was yet present in some areas such as in Scotland and the Netherlands14. Generally, wild resources, such as game and fish, were not prominent in the protohistoric diet in western Europe12 and are even suggested to have been taboo14.
The diet in the Roman period, however, is expected to have changed significantly under the important influence of the Mediterranean world. Still, cereal and some pulses remained the staple food13,15, with the specific dominant crop and livestock types largely following the local environmental demands16. Seeds and pollen in the Gallo-Roman wells in Destelbergen revealed the presence of nearby yards in which several Roman introductions were cultivated4,17,18. This shows a change in the amount and type of resources available and the implementation of Mediterranean influences in the diet19. In terms of livestock, in Destelbergen, the first century occupation was associated with some sheep bone while the settlement remains from the following centuries delivered mostly cattle with smaller amounts of pig and horse bone2,4.
Carbon (δ13C) and nitrogen (δ15N) isotope studies on cremated individuals are not informative for dietary studies because these isotopes are altered during cremation. The measurable δ13C after burning reflect the cremation conditions rather than the original dietary practices of the deceased during life20,21,22. Still, these analyses on inhumated human bone were able to paint an isotopic picture of Metal Ages and Roman diets. The Romano-British population generally revealed a significant rise in marine protein compared to the Metal Ages as attested by δ13C and δ15N studies9,23,24,25. Due to lacking studies on inhumated bone from Belgium, it is uncertain whether this shift can be extrapolated to Belgium. Fish sauce (a condiment made of salted, fermented fish) and molluscs are suggested as a potential source of this marine protein as these products are easier to transport and preserve than fresh fish9,25,26. In Belgium, the originally imported product from the Mediterranean seems to have been substituted by a locally produced fish sauce in the second and third centuries27,28, confirming an important local demand. Clearly, the consumption of marine food seems a Roman dietary addition in Belgium and England. In Destelbergen, however, no evidence of fish consumption has been uncovered, nor in the pottery, nor in the animal bone record. Absence of this archaeological evidence does not necessarily mean that these foods were not consumed. This lack of affirmation could be caused by the use of perishable (wooden), or else generic, unrecognised types of transport vessels29 and the fact that the applied excavation methods were not aimed at finding tiny fish bone fragments.
Another vital component in the diet for humans and domestic animals alike is salt30. Next to that and its multiple industrial uses, this mineral revolutionised food storage thanks to its antimicrobial properties, increased transportability, and availability of products such as meat (most often pork), fish and dairy (in butter and cheese) throughout the year30,31,32. Inscriptions from a submerged native Nehalennia temple in the Scheldt estuary (the Netherlands) indicate the presence of fish sauce and salt traders in the area33,34. Salt production at the North Sea coast goes back to at least Early Iron Age practices (seventh century BCE)35. This production is thought to have increased in the Roman period in part to sustain the military apparatus defending the northern borders of the Roman Empire, but in the process possibly markedly increasing the availability of salt in the region7,35,36,37. It remains, however, hard to assess to what degree the actual salt consumption in the wider population changed in the transition from Iron Age to Roman period. Van den Broeke sees an overall rise in the number of salt containers in the hinterland as a proxy for an increased salt consumption from the fourth century BCE onwards38. In Britain, intensification of salt production during the Late Iron Age has also been linked to the rise of meat preservation and in the trade of otherwise perishable foods31,32. The presence of salt container fragments in Destelbergen4,35 and many other settlement sites affirms the availability of salt in the Gallo-Roman economy. Isotopically, an increase in salted or brined food may have a significant impact on the 87Sr/86Sr and [Sr] of human bones and teeth39,40.
In the inland area with modest sandy ridges and wide river valleys, mixed subsistence farming including cattle, sheep and pig breeding in varying frequencies depending on the local environment is the expected form of economy during the Late Bronze Age and Early Iron Age41. A general cold episode with extreme wetness during the Early Iron Age (750–400 BCE) led to a shortening of the growing season for crops and contractions in settlement patterns42. In this period, cemeteries were usually not established adjacent to the settlement43,44. To date, indeed no indications for Metal Ages settlements have been found close to the Destelbergen cemetery. This fits in the concept of ‘wandering farmsteads’41,45. This paradigm encompasses that farms moved locations every other generation within a certain territory while maintaining an expanding central communal cemetery over generations. Especially in regions with fast degenerating sandy soils this moving of the farmstead to newly regenerated agricultural land is presumed necessary to keep crop production up to standards. Nevertheless, this model has been nuanced and longer continuity of settlements has been observed in more sustainable environments46 (Suppl. Text 2).
In contrast to the Destelbergen Metal Ages group, the Gallo-Roman group lived on site and as such provides a better idea of the settlement size and type. The number of burials (n = 204, based on fragmentary salvage excavations) covering an estimated period of 230 years (Flavian period until the end of third century3) in combination with the dense associated settlement remains suggest that the site in the Roman period was significantly larger than in the Metal Ages, needing an intensified agricultural production. The last two centuries BCE until around 400 CE experienced drier and warmer conditions, favourable for crop yields42. Recovered seeds and pollen suggest the inhabitants were essentially supporting themselves with staple foods from nearby fields, pastures, and vegetable plots17. The sandy region around Destelbergen, however, is not ideal for large surplus cereal productions to trade, possibly explaining the lack of villa domains and vici, but is expected to have been more suited for animal husbandry34,47. In well-connected places, the locally produced ‘farmer’s diet’ could be supplemented with imported, preservable special goods that could often not be produced locally, such as olive oil, wine, and salted products (e.g. salted meat and fish). Besides a clear consumption of local produce, the archaeobotanical evidence, next to considerable amounts of imported pottery4 and stone products48 in sites like Destelbergen, warns to not underestimate the extent of mobility of consumer goods during the Roman period; more than is reflected in the Metal Ages cemetery. Receiving traded goods, however, does not mean that the inhabitants themselves were more mobile47, but rather that there was a dynamic trade network.
During cremation, all organic matter is destroyed and the carbon present in the mineral fraction (often called bioapatite) is heavily altered, reflecting the cremation conditions rather than the diet49,50. As such, the only proxies currently available to investigate changes in mobility and potentially in diet in calcined human remains are 87Sr/86Sr and [Sr] as they are both unaltered by the cremation process and post-burial diagenesis22,51 (Suppl. Text 3, Suppl. Table 1a and 1b, Suppl. Fig. 2).
Strontium enters the food chain via plants and their 87Sr/86Sr vary depending on the type of underlying bedrock52. 87Sr/86Sr measured in human remains, therefore, reflect the geographical origin of the food and drinks consumed. Figure 1 shows a geological map of Destelbergen. The area is dominated by the Eocene Gentbrugge formation that is locally covered by 10 to 20 m of quaternary sediments of fluvial and aeolian origin53. Within a 15 km radius, several pre-quaternary lithologies occur, but are usually covered by similar quaternary sediments of varying thickness, potentially affecting the bioavailable strontium. As a result, it is difficult to predict the local bioavailable 87Sr/86Sr range without proper sampling of modern plants.
From the various food and drinks that can contribute to the Sr pool in bone and teeth, meat and milk do not contribute much as Sr accumulates mainly in the skeleton and not in soft tissue52,54,55. This also applies for marine organisms56. In contrast, plants (i.e. crops) represent the main dietary source of strontium40,57. Marine resources, especially salt, can also be a major contributor of Sr and heavily alter both 87Sr/86Sr and [Sr] in human remains40,54,56,58,59. This influence can easily be recognised since marine resources (and seawater) have a characteristic 87Sr/86Sr of 0.709260. Demonstrating that salt can be impactful, a study on modern salted hams revealed that the meat took on the Sr isotopic signature of the applied salt61 and the [Sr] of several of the 16 hams raised up to 4.6 ppm61, instead of the usual low content in meat (pork fat and meat were found ranging from 0.1 to 1.1 ppm54). The 15 used marine and rock salts themselves ranged from 10 to 153 ppm and rock salts often have different 87Sr/86Sr than the current seawater value of 0.7092, depending on the age of the sea the evaporite mineral was formed in61. Ten additional commercial unrefined salts contained 95.2 ± 78.5 (2SD) ppm of Sr62.
As plants, and not meat, generally represent the main source of strontium in the human diet, [Sr] depend heavily on trophic level of the tested individual (i.e. herbivores will have higher [Sr] in their bones and teeth compared to carnivores), even more so than on geological variations. [Sr] therefore provides information about dietary habits rather than geographical origin40,55,63. Products high in [Sr] (eg. fish flour (231–280 ppm), kale (109–117 ppm), kelp (98 ppm), thyme (90 ppm), and several spices in general, clam (26 ppm)54) should affect the resulting Sr mix in a consumer to a larger degree than products lower in Sr, such as grains that are in the range of 1–3.8 ppm54, weighted according to the ingested quantity. Furthermore, Sr uptake (by plants, and thus in the rest of the food chain) is enhanced by soil acidity64 and Sr metabolism is highly correlated with calcium (Ca) intake, with Ca preferentially being incorporated over Sr39,40,65. Considering that Sr substitutes for Ca in the skeleton, high Ca levels in the diet prohibit Sr to be incorporated in the skeleton. Accordingly, high dairy consumption—besides being poor in Sr—prevents the incorporation of Sr in bone and actively reduces [Sr]40,54,55,63. Opposed to this, diets high in fibre and phytate available in grains, leafy vegetables and legumes promote Sr uptake and result in higher [Sr]40,54,57. In addition, [Sr] is also highly correlated with salinity66 of the food supplies. High salt intakes promote Ca excretion and eventually cause detrimental Ca degradation in the skeleton67,68,69. This allows for potential replacement by Sr, although this could not yet be observed in (short term) experimental studies67,68. Fenner and Wright (2014) calculated the amount of salt consumption needed to change the 87Sr/86Sr in Mayan individuals39, but did not account for the metabolic effects of the added salt on Ca balances in the bone. They concluded that a daily dose of 9.2 g of dietary salt per day is able to warp the 87Sr/86Sr towards sea levels in a specific diet of lime-treated (and thus Ca rich) produce, which would be considerably higher than the 5 g daily salt intake recommended by the World Health Organization70. Yet, this high salt intake might not be unusual, since an actual salt intake in the current world population up to even 15 g a day is not uncommon in regions such as West and East Asia71. It is important to keep in mind that human diets are multi-component diets meaning that Sr intakes come from a summation of different sources65. The final Sr signature measured in the bone thus has to be seen as a mixture of Sr resources, rather than a direct reflection of one food source.
Results
Radiocarbon dating
Six new dates on the identified individuals of the bone pit (identified as “gx”) (Suppl. Text 4) from the Roman period show a remarkable overlap. A chi-squared test (T = 1.472 (5% 11.071), df = 5) confirms that these individuals are contemporary and could correspond to the same chronological event between 124 and 204 cal. CE (95.4% probability). See Suppl. Text 4.
87Sr/86Sr in plants as bioavailable reference
Since a bioavailable Sr map for Belgium is not yet developed72, only preliminary results based on limited plant sample sizes are available8,73. While around Herstal and Fize-le-Marsal some plant data was already published8,73, around Destelbergen, six new sampling locations were selected (Fig. 1) within 10 km of the site. Plants were collected following the method described in Snoeck et al. 202074. The 87Sr/86Sr of all 18 new plant samples (three per location) around Destelbergen result in an interquartile range (IQR) of 0.7095 to 0.7104 (Suppl. Table 4). For more information on the published local estimates of Herstal (IQR’s 0.7090–0.7099 and 0.7134–0.7141) and Fize-le-Marsal (IQR 0.7098–0.7103), see Suppl. Text 1 and8,73.
87Sr/86Sr and [Sr]
The 87Sr/86Sr of the Destelbergen Metal Ages group vary from 0.7091 to 0.7117 (n = 89) (Suppl. Table 2) and present a wider range (IQR 0.7094–0.7098, 0.0004) compared to the Roman group (IQR 0.7093–0.7095, 0.0002) with values ranging from 0.7090 to 0.7100 (n = 33) (Fig. 2). Although there is a great deal of overlap, a Mann–Whitney U test confirms both groups can be statistically significantly distinguished (U = 1948, p < 0.01). Most results fall within the IQR seen in the plant samples (0.7095–0.7104). Despite their identical baseline, the difference in 87Sr/86Sr between Metal Ages (n = 9) and Roman individuals (n = 5) in Blicquy (Suppl. Table 3b) is very clear (Mann–Whitney U test: U = 43, p < 0.01). The IQR of the Metal Ages individuals (0.7103–0.7105, 0.0001) does even not overlap with the IQR of the Roman individuals (0.7095–0.7099, 0.0003). Despite their relatively close proximity to each other and similar geological environment, the individuals of Herstal (published, see8) and Fize-le-Marsal exhibit distinct 87Sr/86Sr (Suppl. Table 3a and 3b). It has to be admitted though that in the comparison between these two sites, the baselines are not identical as is the case with the previous sites and local presence of more extreme 87Sr/86Sr are likely more at influence in Herstal. The Roman site exhibits lower results (IQR 0.7095–0.7101, 0.0005), slightly more distributed towards the lower side of the expected local bioavailable value of 0.7098, while the Metal Ages site reveals a higher range (IQR 0.7117–0.7126, 0.0008). The higher results of Herstal are not in concordance with the measured value for the local geological background (0.7092), but fall entirely in between this bioavailable range and the nearby Meuse alluvion measured further downstream (0.7136).

Comparison of 87Sr/86Sr and [Sr] between the Metal Ages and Gallo-Roman buried individuals of Destelbergen, Blicquy, Herstal8 and Fize-le-Marsal. The available 87Sr/86Sr baselines are indicated in grey. The baseline of Herstal includes two isotopically very different geological formations. [Sr] normalized to 40 wt% Ca. (graph created using R Studio (R version 4.0.2), www.r-project.org).
The Metal Ages group in Destelbergen shows four statistical outliers on 89 individuals. Both Blicquy groups show very little variability and give the impression of sites without clear mobility. Herstal on the other hand, despite its large variability, does not show statistical outliers, which suggests an absence of mobility in this group. In a region with variable bioavailable 87Sr/86Sr, however, it is difficult to distinguish non-local individuals, since many non-local signals can overlap with the locally occurring values. In the Gallo-Roman group of Destelbergen, no such outliers are present and the 87Sr/86Sr homogeneity is exceptional (IQR 0.0002). Gallo-Roman Fize-le-Marsal contains a single outlier (burial 17, 0.7119), who could be a non-local individual. Three more individuals exhibit higher 87Sr/86Sr, above 0.7106, than the majority of this group (< 0.7102).
While 87Sr/86Sr clearly shows a contraction towards seawater values in Roman times, even more contrast can be seen in [Sr]. Statistically significant differences in the [Sr] between the Metal Ages and Roman group in Destelbergen can be confirmed using the Mann–Whitney U test (U = 191.5, p < 0.0001), in which the Roman individuals contain higher [Sr] (IQR 144–205, 61 ppm) than their Metal Ages counterparts (IQR 87–130, 43 ppm). In Blicquy a similar shift from lower [Sr] in the Metal Ages individuals (IQR 67–92; 25) to higher [Sr] in the Roman individuals (IQR 118–160; 42) can be observed. Both sites in the Meuse valley show the same trend. Significantly higher [Sr] can be seen in the Gallo-Roman group of Fize-le-Marsal (IQR 139–176, 37 ppm), but not in the Metal Ages group of Herstal (IQR 89–110, 21 ppm). Cross-period testing of all the data with the Pairwise Wilcoxon Signed Rank test leads to a p-score < 0.0001, acknowledging them as statistically different.
Three scatterplots of the data (Suppl. Fig. 3) investigate the correlation between 87Sr/86Sr and [Sr] and clearly visualise a clustering of the Gallo-Roman individuals compared to the more dispersed Metal Ages individuals. This difference is most clear with regards to the [Sr] than for the 87Sr/86Sr taking into account the local bioavailable 87Sr/86Sr variation.
Discussion
The 87Sr/86Sr and [Sr] obtained from cremated individuals from the Metal Ages and the Roman period in Destelbergen, especially when compared to the sites of Blicquy, Herstal and Fize-le-Marsal (Fig. 2), show that mobility, dietary practices and/or land use, considerably changed from one period to the next. As the two cemeteries in Destelbergen were found only 120 m apart and the Roman settlement was situated on top of the Metal Ages cemetery, the underlying bedrock does not cause the differences. The same can be said about Blicquy, where the same underlying bedrock did not offer different bio-available 87Sr/86Sr and [Sr] to the Metal Ages and Roman groups.
The lower variability in the 87Sr/86Sr in the Roman period in Destelbergen (and to a lesser extent in Fize-le-Marsal versus Herstal) compared to the Metal Ages groups (Fig. 2) could be explained by a reduction in mobility. However, this is in contradiction with the current archaeological and historical evidence that generally supports an increase in economic and military mobility in the Roman period compared to the Metal Ages29,75,76.
Changing land use strategies are able to considerably influence the Sr variability of a population. The cultivation of different areas of the landscape over time possibly exhibiting various Sr signatures, may result in the uptake of distinct 87Sr/86Sr77. In Destelbergen, a small sedentary group maintaining the cemetery in the Metal Ages, lived within a reasonably diverse ecosystem (dryer sand ridge, wetter fluvial plain and wetlands) supposedly allowing for some settlement continuity. Nevertheless, they likely moved the location of their settlement and crops over the course of the 650–700 year use of the cemetery45, varying their local Sr intake. This area in which the farmstead shifted would not necessarily have been so large, still the geological background around Destelbergen is not entirely homogeneous (Fig. 1), potentially explaining the more variable 87Sr/86Sr. The same observation applies to Herstal. Even though in Blicquy we have no locally measured baseline, the measurements of the Eocene formation in Destelbergen likely can be extrapolated to the Blicquy environment. The main occurring formations are of Ypresian (like in Destelbergen) and Thanetian age. The Thanetian immediately precedes the Ypresian, thus likely differs not much in (heavily age-dependent52) 87Sr/86Sr. The local baseline can be expected to be in the range of 0.7100–0.7104 (IQR Eocene formation Destelbergen). The Blicquy Metal Ages group exactly matches this expected baseline, but the Roman group falls clearly below. The much higher 87Sr/86Sr of the Herstal group than the locally underlying value of 0.7090–0.7099 (IQR), seems to be influenced by the close proximity of the Meuse alluvion (IQR 0.7134–0.7141). A mixed use of alluvion and river terrace would lead to a population with a variable 87Sr/86Sr in between both geological formations. In this site, the large diversity in 87Sr/86Sr is not in the first place connected to the timespan of the cemetery, which is with maximum 400 years shorter than in Destelbergen, but rather to the large geological diversity present locally.
The Roman settlement in Destelbergen on the other hand, located next to the Roman cemetery (on top of the Metal Ages cemetery), might indeed have exploited a more stable portion of land, since the tested sample represents the land use of a shorter period of time (mainly two centuries). Seeds and pollen confirm nearby grain fields, although the exploitation was not on an intensive scale17. The warmer and dryer climate in the Roman period42 may have led to an intensified exploitation extending into lower lying, wetter parts of the landscape, such as the fluvial plain, while during the Early Iron Age, one had to resort to the higher and drier soils. While a slightly changed and more stable agricultural land use for staple food might have been the case, there is still the established idea that in the Roman period, more consumer goods were transported than ever before. Although staple foods must have mostly been provided locally, this diversified supply nevertheless contributed to the individual’s 87Sr/86Sr and [Sr]. As such, a drop in isotopic variability contrasts expectations.
The 87Sr/86Sr in Fize-le-Marsal fits relatively well with the expected locally available 87Sr/86Sr, except for four individuals with higher results. This is not surprising, since the northwest of Belgium appears to be characterised by a large geological diversity often extending in high 87Sr/86Sr, which is reflected in the large 87Sr/86Sr ranges in the studied sites (see e.g. Herstal8 and Echt73) and sampled plants73. It is however apparent that the Roman groups, despite being located in regions with abundant availability of higher 87Sr/86Sr, pick up on this to a lesser extent and display often a narrower 87Sr/86Sr range than the Metal Ages groups. The 87Sr/86Sr warped towards the lower side of the baselines and the marine value of 0.7092, combined with their elevated [Sr], makes it tempting to compare the Gallo-Romans to the coastal populations of the Hebrides and Orkney78. The extreme marine signatures seen in these Scottish groups found on different isles (Suppl. Fig. 4) present as a relatively homogeneous, contracted 87Sr/86Sr around 0.7092 and elevated and variable [Sr], often in excess of 150 ppm. The Belgian Gallo-Roman individuals tend towards patterns shown in these marine-affected people, whereas this is not the case in the Metal Ages individuals. This marine signal in measured human samples from the Scottish Isles is evidently explained by direct salty sea spray, by Sr-rich seaweed used as a fertilizer40,79, and/or direct consumption of marine products58. Whatever the dominant source of this marine signature is, it consistently reflects the same 87Sr/86Sr of the sea and warps the other Sr resources an individual consumes at a rate depending on the amount and [Sr] of the marine resource39.
Since the Belgian Roman individuals tend towards a rather marine Sr signature, one should try to define the cause of this marine influence. Sea spray is not an option for the Belgian inland sites, as this effect fades out after a few kilometres80. Several reasons make frequent seaweed manuring in the Belgian sites unlikely. The distance from the sea meant that the recurring transportation cost to supply seaweed in large amounts were a considerable disadvantage over other types of terrestrial fertiliser. On top of that, the needed amount to alter the Sr signature of a population, once diluted through the local soil and diminished via refraction in crops, is substantial. Finally, the frequent occurrence of Gallo-Roman stables with manure accumulation pits in the sandy regions in Belgium (and even on site in Destelbergen) from the second century onwards4,34, prove that the use of animal manure was an established practice in the Roman period. It therefore seems unlikely that seaweed as a fertilizer would have played an important role in the available Sr budget in Destelbergen, and even less so in Fize-le-Marsal considering its location further inland. A marine-like influence could, however, have come from a change in diet with a marine component or specific landscape use.
In Destelbergen, the 87Sr/86Sr of the plant samples reveal lower values around 0.7095 in the quaternary alluvion, while on the sandy ridge adjacent to the alluvion, somewhat higher 87Sr/86Sr of 0.7102 are measured. Under an agricultural system limited to the alluvion, e.g. under the influence of the improved climate42, this would indeed lead to lower 87Sr/86Sr in the human individuals. Nevertheless, the settlement itself covered both geological formations and to sustain larger groups of inhabitants, one would expect that much of the surrounding landscape, including the ridge with higher 87Sr/86Sr, would be exploited. Although unexpected, it is possible that there was indeed a selective land use that could lead to lowered, contracted 87Sr/86Sr results for the Gallo-Roman group. However, this is not sufficient to explain the elevated [Sr] seen in this group.
Supplementing one’s diet with large proportions of imported food from regions with a lower 87Sr/86Sr (such as parts of the source regions of rivers Scheldt and Lys in the north of France (e.g. 0.708781) could also have a lowering effect on the mixed 87Sr/86Sr. Although the possibility exists, it is highly unlikely that Destelbergen, a rural settlement which presumably lacked the means to purchase all of its grain elsewhere, would be structurally supplemented with imported staple food from other regions. Interestingly however, a seeds and pollen study of Destelbergen revealed a seed of white laceflower (Orlaya grandiflora), which is a Mediterranean arable weed preferring Ca-rich limy soils that are not locally present. The find of this taxon is commonly interpreted as at a certain moment being introduced via imported grain as food or seed17,18, hinting at potential grain transports from nearby regions such as the loamy belt in Belgium, northern France or the German Rhineland. This weed was also found in the grain cargo of a sunken Roman barge at Woerden (NL) at the Rhine limes82 and in Late Roman coastal castellum of Oudenburg83. In the context of Destelbergen, however, the singular Orlaya grandiflora seed likely results from an occasional introduction and not from structural grain imports, which nonetheless ties the site to a larger trade network. Importantly, structural grain imports would not necessarily explain elevated [Sr] in both Gallo-Roman rural groups.
Dietary changes are more likely to explain the elevated [Sr] seen in the Roman population than biosphere differences40,55,63. Increased [Sr] in itself could be caused by diets lower in dairy and/or meat55,84 and higher in plants55, but there is no direct indication that the Gallo-Romans clearly abandoned dairy and/or meat in their diet. This particular dietary change alone would also not result in a contraction to lower 87Sr/86Sr. The staple food in both Metal Ages and Roman period was based on grains and to a lesser extent on pulses12,13. This diet was in both periods supplemented with meat from domesticated cattle, sheep and pig. However, in the Roman period also fish products, imported foods, some newly introduced vegetables and herbs are entirely new additions. Of these, marine resources are of particular interest to explain the Sr signature shift towards more marine values in Gallo-Roman individuals.
Sea fish in itself is unlikely to transmit a marine Sr signature to human consumers very well, since Sr predominantly accumulates in the fish bone and not in soft tissue. A Sr uptake derived from fish would require the fish skeleton to be eaten as well, which is generally less probable but could be the case in fish sauce. High [Sr] measured in clams54 also sound promising as potential source of marine Sr, although it is unclear whether the not eaten shells were included in the measured samples. A risen sea food consumption in the Roman period, if it included fish bone, would definitely have helped in the Sr signature shift towards marine values in the Roman period, but due to the fact that the fish skeletons are usually discarded, the contribution might be not as severe as with sea salt. Due to their naturally high [Sr], salted products usually have a stronger impact on both the 86Sr/86Sr and [Sr]64 of human remains than untreated products. On top of that, an elevated salt consumption has an added metabolic effect. High salt diets been demonstrated to cause detrimental changes in the Ca balance in the bone69, while Ca reducing conditions in the skeleton are understood to lead to elevated [Sr]40. Salt consumption was for instance found to be at the root of clear 87Sr/86Sr shifts in Mayan samples from Tikal, demonstrating that the amount of 87Sr/86Sr warping is a function of the amount of salt consumed39,85. Based on the combined evidence65, we propose that an elevated use of (sea) salt, probably used as a food preservative, best explains the observed changes in the Sr signature from the Late Bronze Age-Early Iron Age to the Roman period in this region. The economic and social significance of salt in the Roman period, especially in connection with the maintenance of a large military force, is well-recognised and was most likely used in larger amounts than in the Metal Ages35,37. Long distance trade and mobile military forces must have required more edible goods to be preserved and might be a potential driver for elevated salt use. This dynamic, applied to meat, possibly developed in the Late Iron Age31. Additionally, a warmer Mediterranean climate that worsened the shelf life of untreated food, could have stimulated this Roman culinary trend, which in turn would influence the Northern provinces. The rise in fish consumption in the Roman period, often in salted fashion, compared to its absence in the Metal Ages, might be an important manner in which salt consumption increased. Additionally, salted hams were a famous regional and even exported product86, which receive much of their Sr properties from the salt used61. Furthermore, it must be stressed that added salt could alter, but never completely erase the Sr signature of other, dominant ingested food sources. In regions with high bioavailable 87Sr/86Sr, individuals with a diet of local produce combined with a high salt consumption would still display high 87Sr/86Sr, but somewhat lower than the bioavailable signal.
Comparing Sr signatures over time makes it possible to examine salt distribution and consumption patterns. This offers a very interesting addition to the study of salt production based on material remains from production sites. Future research should aim at exploring this shifting effect to elevated [Sr] and warped 87Sr/86Sr range on a wider scale, pinpointing when and locating where sites seem to conform to this pattern and in this way determine how trends in diet and food preservation changed.
The results of this study show a significant rise in [Sr] in Gallo-Roman Destelbergen individuals compared to their Metal Ages predecessors. This effect, also clearly observed in Blicquy and to some degree in Fize-le-Marsal, is accompanied by mildly lowered and contracted 87Sr/86Sr range towards 0.7092. These changes are best explained by an augmented salt consumption and its use as a preservative (e.g. for fish and meat). Despite the vast complexity of Sr metabolism in multi-component diets, these analyses offer an indication of connectivity (trade) and diet rather than mobility and reveal to what degree a group was tied in the wider economic and cultural fabric to obtain commodities such as salt. Circumstantial evidence is therefore needed to support such interpretations, as elemental and isotopic Sr in humans are the summation of many dietary and metabolic factors65. Still, this study confirms that in the absence of C, N, S isotope analyses, isotopic and elemental Sr analysis can detect dietary variations even in cremated human remains, and [Sr] can be more revealing than is often assumed. As in clearly marine populations, where locality/mobility of the individuals is often obscured due to the impact of salty sea spray and/or a marine diet, this study demonstrates that in populations with high salt use caution is required when interpreting mobility based on 87Sr/86Sr alone.
Materials and methods
Sampling
One hundred and twenty-two individuals (89 Metal Ages and 33 Roman) from 116 graves were sampled for 87Sr/86Sr and [Sr] analysis. Thirty-four of these sampled individuals have available 14C dates, of which 28 have previously been published87 and six new were obtained for the Roman bone pit (gx) with multiple individuals (Suppl. Text 1). Unfortunately, only a limited number of Roman individual graves could be sampled and of these all bone samples were too small for 14C dating. The same bone fragments used for 14C dating were used for 87Sr/86Sr and [Sr] analyses. In graves that were not 14C dated, calcined diaphysis fragments were selected when available. The fragmentary state of 23 individual graves made sampling of diaphyseal fragments impossible resulting in the selection of two cranial fragments and 21 unidentifiable human bone fragments instead. In the bone pit (gx), seven right parts of mandible were selected (six of which were radiocarbon dated) as these skeletal elements can be used to infer the grave’s minimum number of individuals (MNI). Two of these individuals were identified to be nonadults. Four extra samples from the bone pit were analysed (juvenile rib, adult rib, juvenile diaphysis and adult diaphysis), but as these samples cannot conclusively be identified as separate individuals, these extra data were not included in the statistics and are merely added in Suppl. Table 2.
Nine Metal Ages graves of Blicquy were analysed (eight diaphyseal and one cranial fragment). Five of Blicquy’s Roman graves (diaphysis fragments) were analysed and also 14C dated.
Even though the results of several samples per grave of Herstal were available to study life biographies, bringing to light that in some graves the remains of several individuals were present8, for this study only one sample per grave was (randomly) selected. This ensures that every individual is only accounted for once. This way, Herstal delivered twenty-one samples (all diaphysis fragments) for this study. The selection of these previously published results is listed in Suppl. Table 3a.
Eighteen cremated fragments from Fize-le-Marsal were analysed. All were diaphysis fragments, except for grave 14, for which a cranial fragment was selected in the absence of diaphyseal remains. For reference, four Sr samples from Fize-le-Marsal have been radiocarbon dated (Suppl. Table 3b).
The cremated remains under study are owned by museal and research institutions. The owners have confirmed that limited destructive analysis was allowed for the purpose of this study. Ethical and academic guidelines concerning the study of archaeological human remains set by the Flemish government have been followed during the research. Further administrative and ethical information regarding the cremated remains of these sites can be consulted in Suppl. Text 5.
Sr isotope and concentration analysis
Snoeck et al. (2015) demonstrated that Sr isotope analysis is possible on calcined bone, which holds, thanks to its high crystallinity, even less contamination than previously favoured tooth enamel5. The pretreatment, extraction and mass spectrometry measurements of the samples were performed at the AMGC laboratories at the Vrije Universiteit Brussel (VUB) and G-Time laboratories at the Université Libre de Bruxelles (ULB). The procedures described in Snoeck et al. 20155 were applied. The samples were first mechanically cleaned by drilling off the possibly contaminated outer layer. Next, chemical cleaning was done by three series of 10 min of ultrasonication in milliQ water, followed by one time of 3–10 min of ultrasonication in 1 M acetic acid. Finally, another three times of ultrasonication in milliQ water finishes the cleaning process. The cleaned samples are then dried and powdered. The extraction process is described in Snoeck et al. (2015)5. Columns filled with Sr-specific resin (Eichrom Sr Spec) separated the Sr from the sample. The calculated Sr column recovery rate yields more than 95% of total Sr. As a reference for testing the accuracy of the subsequent analytical measurements, a sample of the standard ‘bone ash SRM1400’ underwent the same extraction process.
Most measurements of the 87Sr/86Sr were executed on a Nu Plasma MC-ICP Mass Spectrometer (Nu015 from Nu Instruments, Wrexham, UK) at ULB while samples 07209 to 07231 were measured at the VUB on a Nu Plasma 3 (PD017 from Nu Instruments, Wrexham, UK). Repeated measurements of the NBS987 and SRM1400 standards yielded 87Sr/86Sr = 0.710246 ± 45 (2SD for > 300 analyses) and 0.713159 ± 30 (2SD; n = 21) respectively. For this research purpose, this is sufficiently consistent with the mean value of 0.710252 ± 13 (2SD for analyses) obtained by Thermal Ionization Mass Spectrometry (TIMS)88. A standard bracketing method with the recommended value of 87Sr/86Sr = 0.710248 was used to normalise all sample measurements88. Procedural blanks were considered negligible (total Sr (V) of max 0.02 versus 7–10 V for analyses, equivalent to ≈ 0.3%). The 87Sr/86Sr is reported with a 2SE for each sample (absolute error of the individual sample analysis—internal error).
An aliquot of 0.5 ml of all dissolved samples was used to measure the Sr and Ca concentrations. Once diluted again with 0.42 M HNO3, a Thermo Scientific Element 2 sector field ICP mass spectrometer at Vrije Universiteit Brussel (VUB) was used to determine the Sr and Ca concentrations in low and medium resolution respectively using indium (In) as an internal standard and external calibration versus various certified reference materials (SRM1400, CCB01). The strontium data were then normalised to 40 wt% Ca to account for the varying loss of organic matter and carbonates during cremation. All [Sr] mentioned in the text and supplementary data refer to Sr concentrations normalised to 40 wt% Ca. To evaluate the accuracy of the procedure, two internal bioapatite standards (ENF and CBA) were analysed in parallel. Based on repeated digestion and measurement of these reference materials, the analytical precision of the procedure is estimated to be better than 5% relative standard deviation (1SD, n = 33 for CBA and n = 5 for ENF). For the procedures used to prepare the plant samples and produce the Sr baseline map, see Veselka et al. 202173 and Snoeck et al. 202074.
14C dating
The 14C dating of the bioapatite in the cremated bone samples, six of Destelbergen, four of Fize-le-Marsal and five for Blicquy, was performed at the Royal Institute for Cultural Heritage (KIK-IRPA, Brussels, Belgium) and followed the KIK-IRPA protocol89. Radiocarbon dates were calibrated using the software OxCal 4.490,91 and the IntCal20 calibration curve92.
Data processing
Data management, plots and statistical tests were performed with R Studio (R version 4.0.2 (2020–06-22). Maps were compiled with QGIS version 3.12.0-București. Data are openly available in the IsoArcH database https://www.isoarch.eu93.
Data availability
All data are available in the main text or the supplementary materials.
References
De Laet, S. J. et al. Oudheidkundige opgravingen en vondsten in Oost-Vlaanderen V 1969. (Provincie Oost-Vlaanderen, 1970).
De Laet, S. J., Thoen, H. & Bourgeois, J. Les fouilles du séminaire d’archéologie de la Rijksuniversiteit te Gent à Destelbergen Eenbeekeinde (1960–1984) et l’histoire la plus ancienne de la région de Gent (Gand). I La période préhistorique (De Tempel, Haarlem, 1986).
De Vos, S. Het Gallo-romeins grafveld van destelbergen-eenbeekeinde. Een studie van 60 brandrestengraven en 4 ustrina uit de opgravingscampagne van 1998. VOBOV-Info 59, 17–26 (2004).
De Logi, A. & Dalle, S. Destelbergen – Panhuisstraat archeologisch onderzoek – 2011. (Ename Expertisecentrum voor Erfgoedontsluiting vzw, 2013).
Snoeck, C. et al. Calcined bone provides a reliable substrate for strontium isotope ratios as shown by an enrichment experiment. Rapid Commun. Mass Spectrom. 29, 107–114 (2015).
De Reu, J., De Mulder, G., Van Strydonck, M., Boudin, M. & Bourgeois, J. 14C dates and spatial statistics: Modeling intrasite spatial dynamics of urnfield cemeteries in Belgium using case study of Destelbergen cemetery. Radiocarbon 54, 635–648 (2012).
De Clercq, W. Roman rural settlements in Flanders. Perspectives on a ‘non-villa’ landscape in extrema Galliarum. In Villa Landscapes in the Roman North. Economy, culture and lifestyles (eds Roymans, N. & Derks, T.) 235–258 (Amsterdam University Press, Amsterdam, 2011).
Sabaux, C. et al. Multi-proxy analyses reveal regional cremation practices and social status at the Late Bronze Age site of Herstal, Belgium. J. Archaeol. Sci. 132, 105437 (2021).
Richards, M. P., Hedges, R. E. M., Molleson, T. I. & Vogel, J. C. Stable isotope analysis reveals variations in human diet at the Poundbury Camp Cemetery site. J. Archaeol. Sci. 25, 1247–1252 (1998).
Jay, M. & Richards, M. P. British iron age diet: Stable isotopes and other evidence. Proc. Prehist. Soc. 73, 169–190 (2007).
Varalli, A. et al. Bronze Age innovations and impact on human diet: A multi-isotopic and multi-proxy study of western Switzerland. PLoS ONE 16, e0245726 (2021).
Roymans, N. Tribal societies in Northern Gaul. An anthropological perspective. Cingula Vol. 12 (Universiteit van Amsterdam, Amsterdam, 1990).
Ferdière, A. et al. Histoire de l’agriculture en Gaule. 500 a. J.-C/ - 1000 apr. J.-C. (Editions Errance, Paris, 2006).
Dobney, K. & Ervynyck, A. To fish or not to fish? evidence for the possible avoidance of fish consumption during the iron age around the North Sea. In The Later Iron Age in Britain and Beyond (eds Haselgrove, C. & Moore, T.) 403–418 (Oxbow Books, Oxford, 2007).
Van Limbergen, D. What Romans ate and how much they ate of it. Old and new research on eating habits and dietary proportions in classical antiquity. Rev. Belge Philol. d’hist. 96, 1049–1092 (2018).
Lepetz, S. & Matterne, V. Élevage et agriculture dans le Nord de la Gaule durant l’époque gallo-romaine: Une confrontation des données archéologiques et carpologiques. Rev. Archéol Picardie 1, 23–35 (2003).
Van Beurden, L. & Verbruggen, F. Archeobotanisch onderzoek aan Romeinse en vroegmiddeleeuwse waterputten in Destelbergen (B). (BIAX Consult, 2013).
Kooistra, L. I. Borderland Farming: Possibilities and Limitations of Farming in the Roman Period and Early Middle Ages Between the Rhine and Meuse (Van Gorcum, Assen, 1996).
van der Veen, M., Livarda, A. & Hill, A. New plant foods in roman britain — dispersal and social access. Environ. Archaeol. 13, 11–36 (2008).
Zazzo, A., Saliège, J.-F., Lebon, M., Lepetz, S. & Moreau, C. Radiocarbon dating of calcined bones: Insights from combustion experiments under natural conditions. Radiocarbon 54, 855–866 (2012).
Hüls, C. M., Erlenkeuser, H., Nadeau, M. J., Grootes, P. M. & Andersen, N. Experimental study on the origin of cremated bone apatite carbon. Radiocarbon 52, 587–599 (2010).
Snoeck, C. A Burning Question: Structural and Isotopic Analysis of Cremated Bone in Archaeological Contexts (University of Oxford, Oxford, 2014).
Müldner, G. Stable isotopes and diet: Their contribution to Romano-British research. Antiquity 87, 137–149 (2013).
Redfern, R. C., Hamlin, C. & Athfield, N. B. Temporal changes in diet: A stable isotope analysis of late Iron Age and Roman Dorset, Britain. J. Archaeol. Sci. 37, 1149–1160 (2010).
Molleson, T. The anthropological evidence for change through Romanisation of the Poundbury population. Antropol. Anz. 50, 179–189 (1992).
Cummings, C. Meat consumption in roman Britain: The evidence from stable isotopes. Theor. Rom. Archaeol. J. https://doi.org/10.16995/TRAC2008_73_83 (2009).
Van Neer, W. & Lentacker, A. New archaeozoological evidence for the consumption of locally-produced fish sauce in the Northern Provinces of the Roman Empire. Archaeofauna 3, 53–62 (1994).
Van Neer, W., Ervynck, A. & Monsieur, P. Fish bones and amphorae: Evidence for the production and consumption of salted fish products outside the Mediterranean region. J. Rom. Archaeol. 23, 161–195 (2010).
Monsieur, P. Roman amphorae in the Civitas Nerviorum: from the late La Tène period to the Flavians. Rei Cr. Romana Fautorum Acta 38, 353–356 (2003).
Kinory, J. Salt production, distribution and use in the British Iron Age. In Salt Production, Distribution and Use in the British Iron Age (University of Oxford, Oxford, 2011).
Maltby, M. Salt and animal products: Linking production and use in iron age Britain. In Integrating zooarchaeology. In Proceedings of the 9th Conference of the International Council of Archaeozoology, Durham, August 2002 (ed. Maltby, M.) 117–122 (Oxbow Books, 2006).
Tsigarida, I. Salt in Roman Britain. In Archaeology of Salt. Approaching an Invisible Past (eds Brigand, R. & Weller, O.) 211–220 (Sidestone Press, Leiden, 2015).
Stuart, P. & Bogaers, J. E. Nehalennia: Römische Steindenkmäler aus der Oosterschelde bei Colijnsplaat (Rijksmuseum van Oudheden, Leiden, 2001).
De Clercq, W. Lokale gemeenschappen in het Imperium Romanum. Transformaties in rurale bewoningsstructuur en materiële cultuur in de landschappen van het noordelijk deel van de civitas Menapiorum. (Provincie Gallia-Belgica, ca. 100 v. Chr. – 400 n. Chr.). (Ghent University, Ghent, 2009).
van den Broeke, P. W. Southern sea salt in the Low Countries. A reconnaissance into the land of the Morini. In Archaeological and Historical Aspects of West-European Societies. Album Amicorum André Van Doorselaer, Acta Archaeologica Lovaniensia Monographiae 8 (eds Lodewijckx, M. & Van Doorselaer, A.) 193–205 (Leuven University Press, Leuven, 1994).
Dekoninck, M. D. Romeinse zoutproductie langs de Menapische kust: Een ingenieus technologische proces. Signa Rom. 7, 61–66 (2018).
van Beek, B. L. Salinatores and sigillata: The coastal areas of North-Holland and Flanders and their economic differences in the 1st century AD. Helinium 23, 3–12 (1983).
van den Broeke, P. W. Iron Age sea salt trade in the Lower Rhine area. In Different Iron Ages: Studies on the Iron Age in temperate Europe (eds Hill, J. D. & Cumberpatch, C. G.) 149–162 (BAR publishing, Oxford, 1995).
Fenner, J. N. & Wright, L. E. Revisiting the strontium contribution of sea salt in the human diet. J. Archaeol. Sci. 44, 99–103 (2014).
Montgomery, J. Passports from the past: Investigating human dispersals using strontium isotope analysis of tooth enamel. Ann. Hum. Biol. 37, 325–346 (2010).
Roymans, N. & Fokkens, H. Een overzicht van veertig jaar nederzettingsonderzoek in de Lage Landen. In Nederzettingen uit de bronstijd en de vroege ijzertijd in de Lage Landen (eds. Fokkens, H. & Roymans, N.) 1–19 (Rijksdienst voor het Oudheidkundig Bodemonderzoek, 1991).
Jones, M. E. The End of Roman Britain (Cornell University Press, New York, 1998).
Roymans, N. & Kortlang, F. P. Urnfield symbolism, ancestors and the land in the Lower Rhine Region. in Land and ancestors. Cultural dynamics in the Urnfield period and the Middle Ages in the Southern Netherlands (eds. Theuws, F. & Roymans, N.) 33–62 (1999).
De Mulder, G. & Bourgeois, J. Shifting centres of power and changing elite symbolism in the scheldt fluvial basin during the Late Bronze Age and the Iron Age. In Atlantic Europe in the First Millennium BC Crossing the Divide (eds Moore, T. & Armada, X. L.) 302–318 (Oxford University Press, Oxford, 2015).
Schinkel, K. Zwervende Erven: Bewoningssporen in Oss-Ussen uit Bronstijd, Ijzertijd en Romeinse tijd: Opgravingen 1976–1986 (Universiteit Leiden, Leiden, 1994).
De Mulder, G. Funeraire Rituelen in Het Scheldebekken Tijdens de Late Bronstijd en de Vroege Ijzertijd. De Grafvelden in Hun Maatschappelijke en Sociale Context (Ghent University, Ghent, 2011).
Groot, M., Evans, J. & Albarella, U. Mobility of cattle in the iron age and Roman Netherlands. J. Archaeol. Sci. Rep. 32, 102416 (2020).
Reniere, S. Sourcing the Stone in Roman Northern Gaul On the Provenance, Use, and Socio-Economics of Stone Tools in a Stoneless Landscape: The Case of the Northern Civitas Menapiorum (Universiteit Gent, Ghent, 2018).
Snoeck, C., Brock, F. & Schulting, R. J. Carbon exchanges between bone apatite and fuels during cremation: Impact on radiocarbon dates. Radiocarbon 56, 591–602 (2014).
Stamataki, E. et al. Is it hot enough? A multi-proxy approach shows variations in cremation conditions during the Metal Ages in Belgium. J. Archaeol. Sci. 136, 105509. https://doi.org/10.1016/j.jas.2021.105509 (2021).
Harbeck, M. et al. Research potential and limitations of trace analyses of cremated remains. Forensic Sci. Int. 204, 191–200 (2011).
Bentley, R. A. Strontium isotopes from the earth to the archaeological skeleton: A review. J. Archaeol. Method Theory 13, 135–187 (2006).
Databank Ondergrond Vlaanderen. https://www.dov.vlaanderen.be/portaal/?module=verkenner.
Schroeder, H. A., Tipton, I. H. & Nason, A. P. Trace metals in man: Strontium and barium. J. Chronic Dis. 25, 491–517 (1972).
Underwood, E. J. Trace Elements in Human and Animal Nutrition (Elsevier, Amsterdam, 1977).
Burton, J. H. & Douglas Price, T. Evaluation of bone strontium as a measure of seafood consumption. Int. J. Osteoarchaeol. 9, 233–236 (1999).
Lambert, J. B. & Weydert-Homeyer, J. M. The fundamental relationship between ancient diet and the organic consituents of bone as derived from feeding experiments. Archaeometry 35, 279–294 (1993).
Lahtinen, M., Arppe, L. & Nowell, G. Source of strontium in archaeological mobility studies—marine diet contribution to the isotopic composition. Archaeol. Anthropol. Sci. 13, 1. https://doi.org/10.1007/s12520-020-01240-w (2021).
Fernández-Crespo, T. et al. Multi-isotope evidence for the emergence of cultural alterity in Late Neolithic Europe. Sci. Adv. 6(4), eaay2169. https://doi.org/10.1126/sciadv.aay2169 (2020).
Hess, J., Bender, M. L. & Schilling, J.-G. Evolution of the ratio of strontium-87 to strontium-86 in seawater from cretaceous to present. Science 231, 979–984 (1986).
Epova, E. N. et al. 87Sr/86Sr isotope ratio and multielemental signatures as indicators of origin of European cured hams: The role of salt. Food Chem. 246, 313–322 (2018).
Kuhnlein, H. V. The trace element content of indigenous salts compared with commercially refined substitutes. Ecol. Food Nutr. 10, 113–121 (1980).
Aufderheide, A. C. Chemical analysis of skeletal remains. In Reconstruction of Life from the Skeleton (eds Iscan, M. Y. & Kennedy, K. A. R.) 237–260 (Alan R. Liss Inc., New York, 1989).
Odum, H. T. Biogeochemical deposition of strontium. Publ. Inst. Mar. Sci. IV, 38–114 (1957).
Sillen, A. & Kavanagh, M. Strontium and paleodietary research: A review. Am. J. Phys. Anthropol. 25, 67–90 (1982).
Limburg, K. E. The biogeochemistry of strontium: A review of H.T. Odum’s contributions. Ecol. Modell. 178, 31–33 (2004).
Evans, C., Chughtai, A. Y., Blumsohn, A., Giles, M. & Eastell, R. The effect of dietary sodium on calcium metabolism in premenopausal and postmenopausal women. Eur. J. Clin. Nutr. 51, 394–399 (1997).
Teucher, B. et al. Sodium and bone health: Impact of moderately high and low salt intakes on calcium metabolism in postmenopausal women. J. Bone Miner. Res. 23, 1477–1485 (2008).
Tiyasatkulkovit, W. et al. Excessive salt consumption causes systemic calcium mishandling and worsens microarchitecture and strength of long bones in rats. Sci. Rep. 11, 1–15 (2021).
World Health Organization (WHO). Guideline: Sodium intake for adults and children. (2012).
Powles, J. et al. Global, regional and national sodium intakes in 1990 and 2010: A systematic analysis of 24 h urinary sodium excretion and dietary surveys worldwide. BMJ Open 3, 1–18 (2013).
Dalle, S. et al. Preliminary results in the collecting of protohistoric cremation samples for the CRUMBEL project. Lunula Archaeol. Protohist. 27, 9–14 (2019).
Veselka, B. et al. Divergence, diet, and disease: The identification of group identity, landscape use, health, and mobility in the fifth- to sixth-century AD burial community of Echt, the Netherlands. Archaeol. Anthropol. Sci. 13, 97 (2021).
Snoeck, C. et al. Towards a biologically available strontium isotope baseline for Ireland. Sci. Total Environ. 712, 136248 (2020).
de Soto, P. Network analysis to model and analyse roman transport and mobility. Comput. Soc. Sci. Simul. Past (2019). doi:https://doi.org/10.1007/978-3-030-04576-0_13.
Pitts, M., Versluys, M. Globalisation and the Roman World. (Cambridge University Press, New York, 2014).
Montgomery, J., Evans, J. A. & Cooper, R. E. Resolving archaeological populations with Sr-isotope mixing models. Appl. Geochem. 22, 1502–1514 (2007).
Evans, J. A., Chenery, C. A. & Montgomery, J. A summary of strontium and oxygen isotope variation in archaeological human tooth enamel excavated from Britain. J. Anal. At. Spectrom. 27, 754–764 (2012).
Evans, J. A., Montgomery, J., Wildman, G. & Boulton, N. Spatial variations in biosphere 87Sr/86Sr in Britain. J. Geol. Soc. Lond. 167, 1–4 (2010).
Alonzi, E., Pacheco-Forés, S. I., Gordon, G. W., Kuijt, I. & Knudson, K. J. New understandings of the sea spray effect and its impact on bioavailable radiogenic strontium isotope ratios in coastal environments. J. Archaeol. Sci. Rep. 33, 102462 (2020).
Willmes, M. et al. Mapping of bioavailable strontium isotope ratios in France for archaeological provenance studies. Appl. Geochem. 90, 75–86 (2018).
Haalebos, J. K. Ein römisches Getreideschiff in Woerden. Jahrb. Römisch-Ger. Zentral. Mainz 43, 475–509 (1996).
Vanhoutte, S. et al. De dubbele waterput uit het laat-Romeinse castellum van Oudenburg (prov. West-Vlaanderen): Tafonomie, chronologie en interpretatie. Relicta 5, 9–142 (2009).
Burton, J. H. & Wright, L. E. Nonlinearity in the relationship between bone Sr/Ca and diet: Paleodietary implications. Am. J. Phys. Anthropol. 96, 273–282 (1995).
Wright, L. E. Identifying immigrants to Tikal, Guatemala: Defining local variability in strontium isotope ratios of human tooth enamel. J. Archaeol. Sci. 32, 555–566 (2005).
Kropff, A. An English translation of the Edict on Maximum Prices, also known asthe Price Edict of Diocletian. (Edictum de pretiis rerum venalium). Version 2.1. https://www.academia.edu/23644199/New_English_translation_of_the_Price_Edict_of_Diocletianus (2016).
De Mulder, G., Van Strydonck, M., Boudin, M. & Deweirdt, E. Een voorlopig overzicht van de 14C-resultaten op gecremeerd bot en houtskool van het urnengrafveld te Destelbergen (provincie Oost-Vlaanderen, België). Lunula. Archaeol. Protohist. 17, 65–71 (2009).
Weis, D. et al. High-precision isotopic characterization of USGS reference materials by TIMS and MC-ICP-MS. Geochemistry, Geophys. Geosystems 7, Q08006. https://doi.org/10.1029/2006GC001283 (2006).
Wojcieszak, M., Van den Brande, T., Ligovich, G. & Boudin, M. Pretreatment protocols performed at the royal institute for cultural heritage (RICH) prior to AMS 14C measurements. Radiocarbon 62, e14–e24 (2020).
Bronk Ramsey, C. Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360 (2009).
Bronk Ramsey, C. Methods for summarizing radiocarbon datasets. Radiocarbon 59, 1809–1833 (2017).
Reimer, P. J. et al. The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757 (2020).
Salesse, K. et al. IsoArcH.eu: An open-access and collaborative isotope database for bioarchaeological samples from the Graeco-Roman world and its margins. J. Archaeol. Sci. Reports 19, 1050-1055 (2018).
Acknowledgements
Many thanks to dr. Patrick Monsieur of the Ghent University Museum for access to the Destelbergen and Roman Blicquy samples. Thank you to the Musée Communal d’Archéologie Hesbignonne for the Fize-le-Marsal samples and to the Archéosite et Musée d’Aubechies-Beloeil asbl for the Metal Ages Blicquy samples. We thank our funding sources: Research Foundation Flanders (FWO) & Fonds de la Recherche Scientifique (FNRS-FRS) Excellence of Science (EOS) grant 30999782 (CS, MB, GDM, MV), Fonds de la Recherche Scientifique (FNRS-FRS), doctoral grant (AS), Research Foundation Flanders (FWO), doctoral grants 11F1120N and 11A6221N (resp. MH and ES), VUB Strategic Research Programme (SRP2), Research Foundation Flanders (FWO-Hercules program) (HERC19). This research is supported by the ERC Starting Grant LUMIERE (Landscape Use and Mobility In EuRope – Bridging the gap between cremation and inhumation), funded by European Union’s Horizon 2020 research and innovation programme under grant agreement number 948913.
Author information
Authors and Affiliations
Contributions
Conceptualisation: S.D., C.S., G.D.M., A.S. & K.S.; Methodology: S.D., C.S., S.G.; Formal analysis: S.D., C.S., S.G., C.G., T.B.; Investigation: S.D., M.H., C.S., A.S., M.B., G.C., T.B., C.G., S.G.; Visualisation: S.D.; Supervision: C.S., G.D.M.; Writing—original draft: S.D., C.S., G.D.M.; Writing—review & editing: S.G., A.S., K.S., R.A., C.S.a., E.S., M.V., B.V., E.W.; Funding acquisition: C.S., M.B., G.D.M., M.V.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dalle, S., Snoeck, C., Sengeløv, A. et al. Strontium isotopes and concentrations in cremated bones suggest an increased salt consumption in Gallo-Roman diet. Sci Rep 12, 9280 (2022). https://doi.org/10.1038/s41598-022-12880-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-12880-4
- Springer Nature Limited